- Роль, распределение и миграция кальция в клетке

Содержание
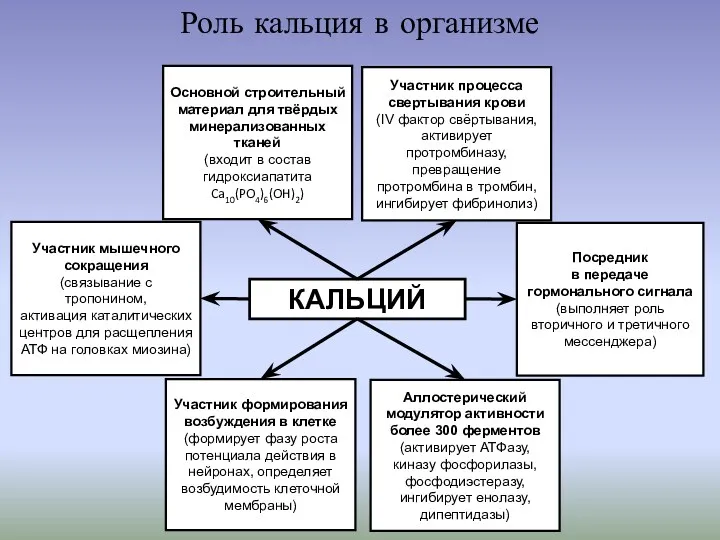
- 2. Роль кальция в организме КАЛЬЦИЙ Основной строительный материал для твёрдых минерализованных тканей (входит в состав гидроксиапатита
- 3. Распределение кальция I пул – экстрацеллюлярный кальций (кальций внеклеточного пространства). II пул – интрацеллюлярный кальций (внутриклеточный
- 4. Кальцийсвязывающие белки Кальцийсвязывающие белки Каналообразующие белки (Са2+-АТФаза) Функциональные белки (тропонин С, кальмодулин) Депонирующие белки (кальсеквестрин, кальрегулин,
- 5. Миграция кальция 1. Медленные потенциалозависимые кальциевые каналы сарколеммы L-типа 2. Кальциевые каналы саркоплазматического ретикулума, связанные с
- 6. Потенциалозависимые кальциевые каналы Состав: Входные ворота Пора Виды: L-типа N-типа P-типа T-типа Цитоплазматическая петля, формирующая ножку,
- 7. Рианодиновый рецептор Рианодиновые рецепторы (RyR) - особый тип хемоактивируемых Са2+-каналов, имеющихся в мембране саркоплазматического ретикулума. Выполняют
- 8. Рианодиновый рецептор
- 9. Рианодиновый рецептор Механизм высвобождения Ca2+ из цистерн саркоплазматического ретикулума. А - рецептор рианодина в мембране цистерны
- 10. Рианодиновый рецептор Механизм высвобождения Ca2+ из цистерн саркоплазматического ретикулума. А - рецептор рианодина в мембране цистерны
- 11. Ca2+-АТФаза Механизм работы Са2+-АТФазы. (1) - Когда протеин находится в исходной конформации, два иона Ca2+ по
- 12. Na+/Ca2+-обменник Натрий-кальциевый обменник - трансмембранный белок цитоплазматической мембраны, транспортирующий ионы кальция из клетки в обмен на
- 13. Тропомиозин и тропонин в отсутствие ионов кальция блокируют связывание миозина с актином. При связывании ионов кальция
- 14. Сокращение кардиомиоцита
- 15. Расслабление кардиомиоцита Активация обмена ионами на сарколемме: ионы Са2+ выходят из клетки, а ионы Na+ входят
- 16. Механизм сокращения В общем объёме кальция, обеспечивающего сокращение, на долю межклеточного кальция приходится 20%, на долю
- 18. Скачать презентацию
Роль кальция в организме
КАЛЬЦИЙ
Основной строительный материал для твёрдых минерализованных тканей
(входит
Роль кальция в организме
КАЛЬЦИЙ
Основной строительный материал для твёрдых минерализованных тканей
(входит

Распределение кальция
I пул
– экстрацеллюлярный кальций
(кальций внеклеточного пространства).
II пул
Распределение кальция
I пул
– экстрацеллюлярный кальций
(кальций внеклеточного пространства).
II пул
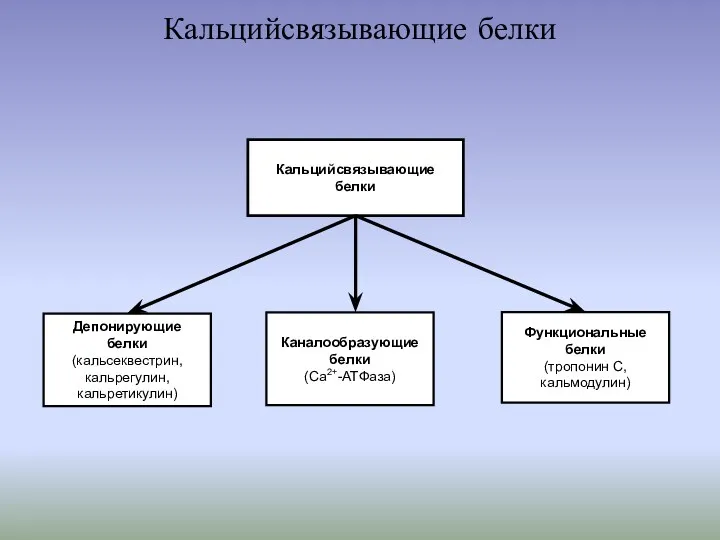
Кальцийсвязывающие белки
Кальцийсвязывающие
белки
Каналообразующие белки
(Са2+-АТФаза)
Функциональные
белки
(тропонин С, кальмодулин)
Депонирующие
белки
(кальсеквестрин, кальрегулин, кальретикулин)
Кальцийсвязывающие белки
Кальцийсвязывающие
белки
Каналообразующие белки
(Са2+-АТФаза)
Функциональные
белки
(тропонин С, кальмодулин)
Депонирующие
белки
(кальсеквестрин, кальрегулин, кальретикулин)
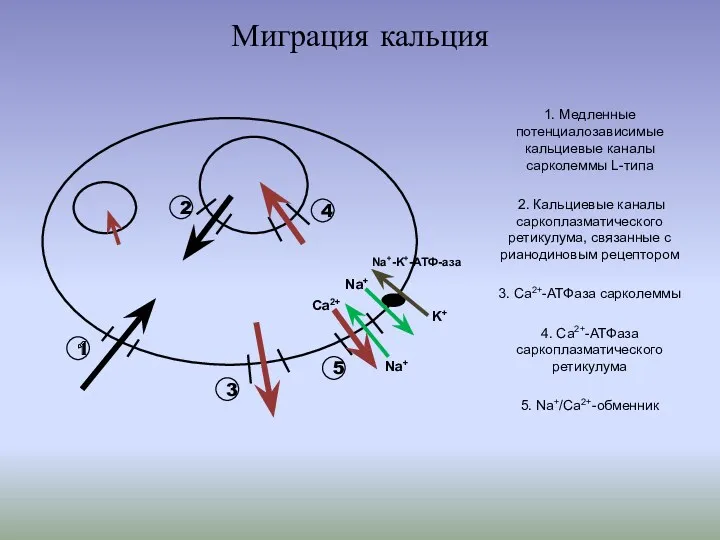
Миграция кальция
1. Медленные потенциалозависимые кальциевые каналы сарколеммы L-типа
2. Кальциевые каналы
Миграция кальция
1. Медленные потенциалозависимые кальциевые каналы сарколеммы L-типа
2. Кальциевые каналы
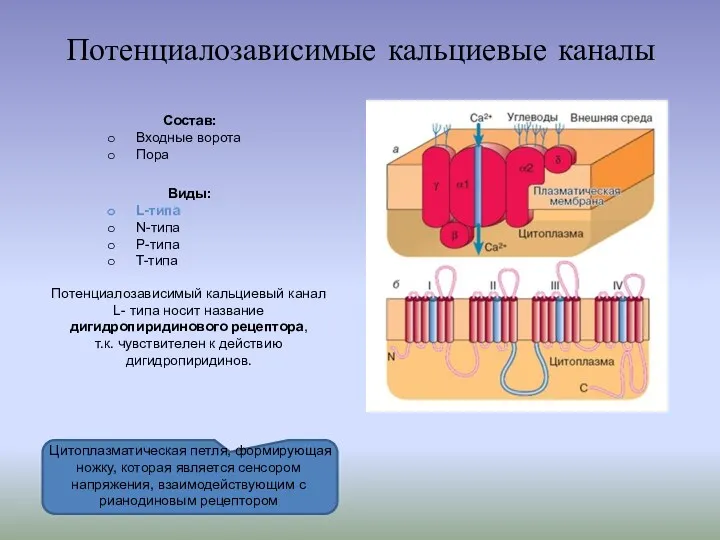
Потенциалозависимые кальциевые каналы
Состав:
Входные ворота
Пора
Виды:
L-типа
N-типа
P-типа
T-типа
Цитоплазматическая петля, формирующая ножку, которая является сенсором
Потенциалозависимые кальциевые каналы
Состав:
Входные ворота
Пора
Виды:
L-типа
N-типа
P-типа
T-типа
Цитоплазматическая петля, формирующая ножку, которая является сенсором

Рианодиновый рецептор
Рианодиновые рецепторы (RyR) -
особый тип хемоактивируемых Са2+-каналов,
имеющихся в мембране саркоплазматического
Рианодиновый рецептор
Рианодиновые рецепторы (RyR) -
особый тип хемоактивируемых Са2+-каналов,
имеющихся в мембране саркоплазматического
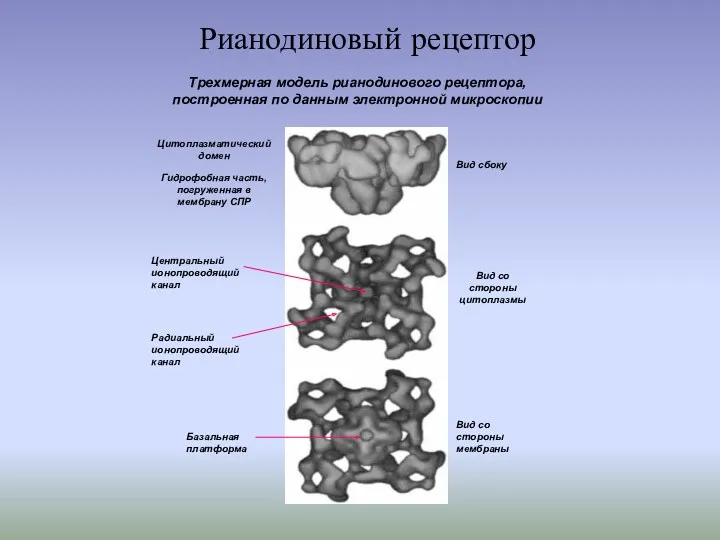
Рианодиновый рецептор
Рианодиновый рецептор
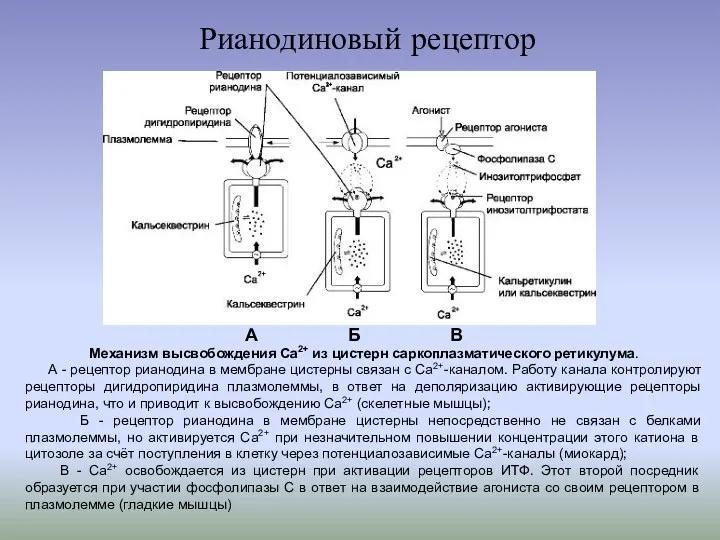
Рианодиновый рецептор
Механизм высвобождения Ca2+ из цистерн саркоплазматического ретикулума.
А - рецептор
Рианодиновый рецептор
Механизм высвобождения Ca2+ из цистерн саркоплазматического ретикулума.
А - рецептор

Рианодиновый рецептор
Механизм высвобождения Ca2+ из цистерн саркоплазматического ретикулума.
А - рецептор
Рианодиновый рецептор
Механизм высвобождения Ca2+ из цистерн саркоплазматического ретикулума.
А - рецептор
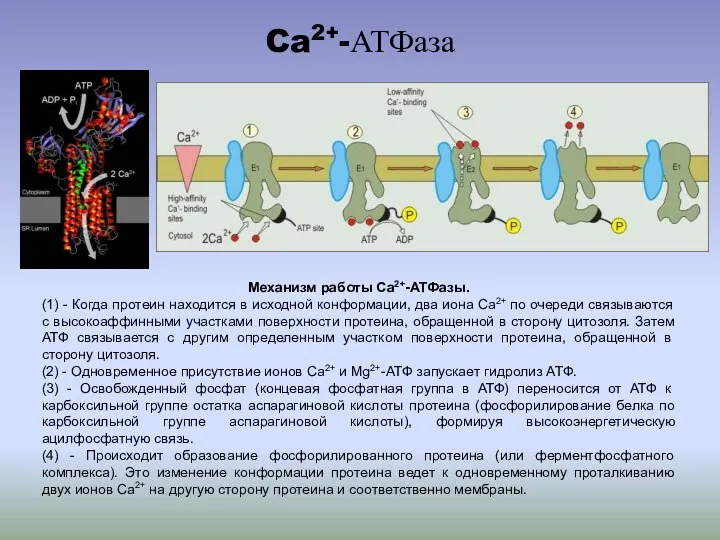
Ca2+-АТФаза
Механизм работы Са2+-АТФазы.
(1) - Когда протеин находится в исходной конформации, два
Ca2+-АТФаза
Механизм работы Са2+-АТФазы.
(1) - Когда протеин находится в исходной конформации, два

Na+/Ca2+-обменник
Натрий-кальциевый обменник -
трансмембранный белок цитоплазматической мембраны, транспортирующий ионы кальция из клетки
Na+/Ca2+-обменник
Натрий-кальциевый обменник -
трансмембранный белок цитоплазматической мембраны, транспортирующий ионы кальция из клетки
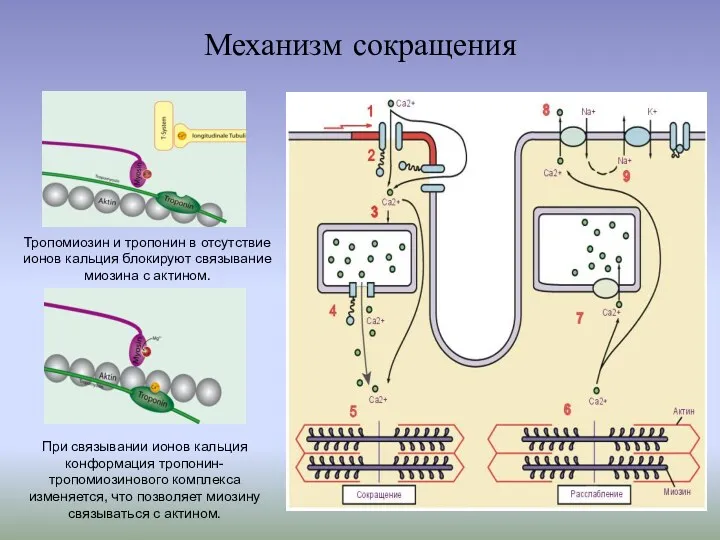
Тропомиозин и тропонин в отсутствие ионов кальция блокируют связывание миозина с
Тропомиозин и тропонин в отсутствие ионов кальция блокируют связывание миозина с
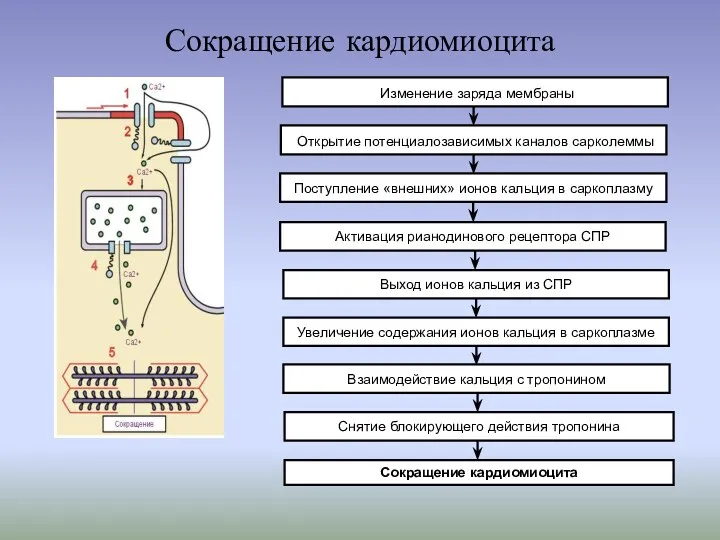
Сокращение кардиомиоцита
Сокращение кардиомиоцита
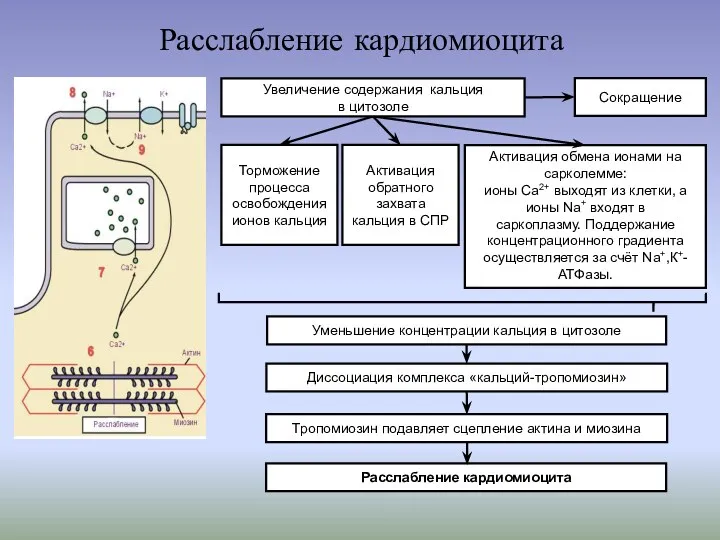
Расслабление кардиомиоцита
Активация обмена ионами на сарколемме:
ионы Са2+ выходят из клетки,
Расслабление кардиомиоцита
Активация обмена ионами на сарколемме:
ионы Са2+ выходят из клетки,
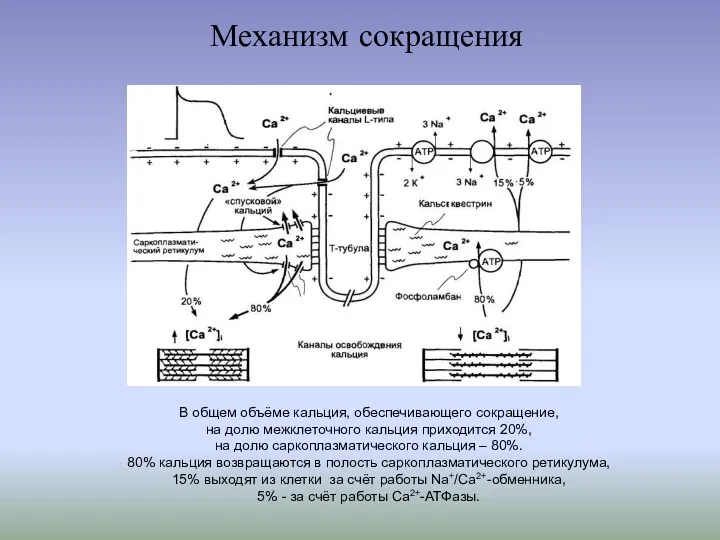
Механизм сокращения
В общем объёме кальция, обеспечивающего сокращение,
на долю межклеточного кальция
Механизм сокращения
В общем объёме кальция, обеспечивающего сокращение,
на долю межклеточного кальция
 Вид. Критерии вида
Вид. Критерии вида Биологическая продукция, скорость накопления биомассы в экосистеме
Биологическая продукция, скорость накопления биомассы в экосистеме Амурский тигр
Амурский тигр Плесневые грибы и дрожжи
Плесневые грибы и дрожжи Строение и функции головного мозга
Строение и функции головного мозга Anatomy of the human brain
Anatomy of the human brain Рибонуклеиновые кислоты АТФ. Химический состав клетки. Рибонуклеиновые кислоты
Рибонуклеиновые кислоты АТФ. Химический состав клетки. Рибонуклеиновые кислоты Органы чувств
Органы чувств Минеральное питание растений. Ассимиляция элементов
Минеральное питание растений. Ассимиляция элементов УРОК1_Живая и неживая природа
УРОК1_Живая и неживая природа Анатомия человека
Анатомия человека Домашние животные.
Домашние животные. Тип Кишечнополостные
Тип Кишечнополостные Презентация к бинарному уроку по физике и биологии в 8 классе Глаз и зрение
Презентация к бинарному уроку по физике и биологии в 8 классе Глаз и зрение Семейство кошачьих
Семейство кошачьих Макроэволюция. Главные направления эволюционного процесса
Макроэволюция. Главные направления эволюционного процесса Что представляет собой хорда?
Что представляет собой хорда? Систематика водных позвоночных-2
Систематика водных позвоночных-2 Развитие жизни на Земле
Развитие жизни на Земле Информационные технологии на уроках биологии
Информационные технологии на уроках биологии Постэмбриональный период
Постэмбриональный период Эмбриогенез и клиническая анатомия мочевыделительной и мочеполовой систем
Эмбриогенез и клиническая анатомия мочевыделительной и мочеполовой систем Геном человека. Мультифакториальные болезни
Геном человека. Мультифакториальные болезни Губки и кишечнополостные
Губки и кишечнополостные Гормоны щитовидной и поджелудочной желез. (Лекция 13)
Гормоны щитовидной и поджелудочной желез. (Лекция 13) Формы размножения организмов
Формы размножения организмов Плоды и семена
Плоды и семена Жасушалардың көбеюі
Жасушалардың көбеюі