- Спинной мозг. Физиология ствола мозга

Содержание
- 2. Сегменты спинного мозга 8 шейных (C1 - C8) 12 грудных (Th1 - Th12) 5 поясничных (L1
- 3. ПРИНЦИП МЕТАМЕРИИ
- 4. Принципы иннервации спинного мозга: Сегментарный принцип Морфологических границ между сегментами нет, поэтому деление является – функциональным.
- 5. Закон Белла - Мажанди Вентральные (передние) корешки содержат эфферентные двигательные (выходящие) волокна, а дорсальные (задние) корешки
- 6. Нейроны спинного мозга (13 млн.): 1. Двигательные или мотонейроны (3%): - альфа-мотонейроны - гамма-мотонейроны 2. Вставочные
- 7. Нейроны. В сером веществе спинного мозга находятся тела двигательных, вставочных и вегетативных нейронов. 1) Мотонейроны. Различают
- 8. По эффекторному действию: Возбуждающие (3%) Тормозные
- 9. ФУНКЦИИ СПИННОГО МОЗГА: АФФЕРЕНТНАЯ ПРОВОДНИКОВАЯ РЕФЛЕКТОРНАЯ
- 10. Основные рефлексы спинного мозга Рефлексы растяжения (миотатические) - в основном разгибательные - рефлексы позы, толчковые (прыжок,
- 11. Регуляция двигательной активности спинным мозгом Регуляция фазной активности мышц: Сгибательные рефлексы Локомоции (автоматизмы) Регуляция тонуса мышц:
- 12. Мышцы содержат 3 типа рецепторов растяжения: Певичные и вторичные окончания Сухожильные рецепторы
- 13. Первичные афференты реагирует на степень и скорость растяжения мышц (динамику), а вторичное — только на степень
- 14. 1. Миотатические рефлексы (сухожильные, Т-рефлексы, от тендон) – рефлексы на растяжение мышцы. Вызываются ударом по сухожилию
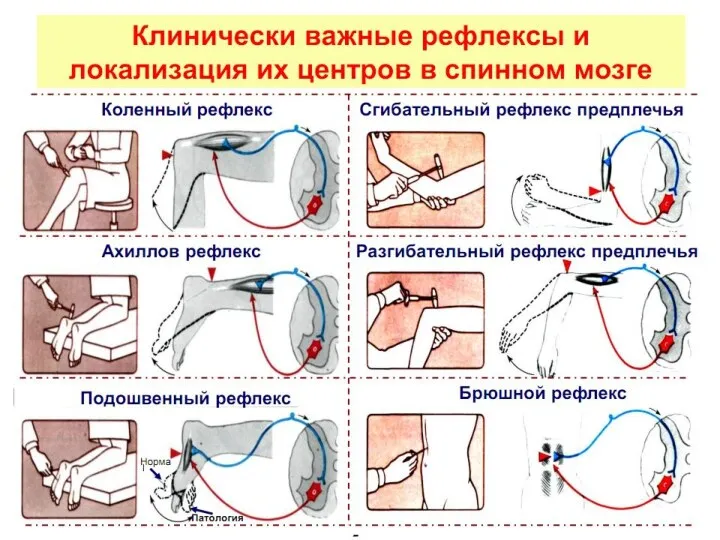
- 16. Облегчение Т–рефлексов. Если коленный и другие Т–рефлексы нижней конечности ослаблены, их можно усилить: для этого больной,
- 17. 2. Постуральные рефлексы (от английск. postur—поза, положение), термин, предложенный Шеррингтоном для обозначения рефлексов, обеспечивающих сохранение определенного
- 18. ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
- 19. Участие спинного мозга в регуляции произвольных движений От супраспинальных отделов управляющие сигналы приходят одновременно на α
- 20. 1. Рефлексы мышц-антагонистов При стимуляции мышечных волокон одной мышцы происходит одновременное торможение мышцы-антагониста.
- 21. 2. Сгибательные рефлексы (защитные) 1. Рефлекторная дуга полисинаптическая. 2. Осуществляются при раздражении различных рецепторных полей: кожных
- 22. 3. Перекрестный сгибательно-разгибательный рефлекс
- 23. Функции сухожильных органов. Рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы. У каждой мышцы
- 24. Активация телец Гольджи приводит к торможению α-мотонейронов и обеспечивает расслабление сокращенной мышцы. 4. Рефлексы ограничивающие напряжение
- 25. Спинальная локомоция (автоматизмы) Основные характеристики локомоции, т.е. перемещения человека или животного в окружающей среде при помощи
- 26. Активность центров координируется проприоспинальными системами и трактами, пересекающими спинной мозг в пределах отдельных сегментов. У более
- 27. Предполагают, что у человека тоже есть спинальные локомоторные центры. По–видимому, их активация при раздражении кожи проявляется
- 28. Шагательный автоматизм Поднимите малыша под мышки ( стопы должны лишь прикасаться к поверхности ). Затем слегка
- 29. СОСАТЕЛЬНЫЙ РЕФЛЕКС
- 30. Автоматизм Бауэра Когда ребенок лежит на животе, подставьте свою ладонь ему под стопы. Он оттолкнется ножками
- 31. Рефлекс Бабинского Грудничок разводит пальчики на ногах веером лишь тогда, когда вы касаетесь внешней стороны стопы
- 32. Рефлекс Моро Положите малыша на спинку и хлопните ладонями по кровати рядом с его головой. Он
- 33. РЕФЛЕКС ГАЛАНТА Проведите пальцем вдоль позвоночника с правой стороны ( на расстоянии сантиметра от него ).
- 34. Хватательный автоматизм Робинсона. Угасает этот рефлекс к 4-му месяцу. А на смену ему приходит сознательное хватание
- 35. Полная параплегия возникает при разрыве спинного мозга в грудном отделе– от Т2 до Т12 и сопровождается:
- 36. В последующие недели и месяцы двигательные рефлексы восстанавливаются. Выделяют следующие четыре стадии: полная арефлексия (обычно 4–6
- 38. СПИНАЛЬНЫЙ ШОК Обратимое угнетение двигательных и вегетативных рефлексов после разрыва спинного мозга называется спинальным шоком. Главный
- 39. Патологические кистевые рефлексы Сгибательные: Рефлекс Жуковского — сгибание пальцев кисти при ударе молоточком по её ладонной
- 40. Патологические стопные рефлексы Разгибательные: Рефлекс Оппенгейма — разгибание I пальца стопы при проведении пальцами по гребню
- 41. Сухожильные (миотатические) рефлексы спинного мозга
- 42. Сгибательные: Нижний рефлекс Россолимо — сгибание пальцев стопы при быстром касательном ударе по их подушечкам. Рефлекс
- 43. Частичная параплегия - синдром Броун-Секара
- 44. Зоны поражения у человека при повреждении спинного мозга на уровне грудных сегментов (синдром Броун-Секара)
- 45. Вегетативные (интероцептивные ) рефлексы спинного мозга
- 46. Висцеромоторные рефлексы Проявляются в двигательных реакциях мышц грудной клетки и брюшной стенки, разгибателей спины при раздражении
- 47. рефлекс Гольца рефлекс Данини-Ашнера раздражение - удар в эпигастральную область, реакция— рефлекторная остановка сердца
- 48. Зоны Захарьина-Геда
- 50. Афферентная функция Восприятие возбуждений, поступающих от афферентных клеток в ответ на раздражение из внешней и внутренней
- 51. ПРОВОДЯЩИЕ СИСТЕМЫ СПИННОГО МОЗГА: ВОСХОДЯЩИЕ ПУТИ (ЭКСТЕРО- ПРОПРИО-ИНТЕРОЦЕПТИВНАЯ ЧУВСТВИТЕЛЬНОСТЬ) НИСХОДЯЩИЕ ПУТИ (ЭФФЕКТОРНЫЕ, ДВИГАТЕЛЬНЫЕ) СОБСТВЕННЫЕ (ПРОПРИОСПИНАЛЬНЫЕ) ПУТИ
- 52. Тонкий пучок Голля (fasciculus gracilis) - от нижней части тела - проприоцепторы сухожилий и мышц, часть
- 53. Латеральный кортикоспинальный пирамидный тракт - двигательные зоны коры - перекрест в продолговатом мозге - мотонейроны передних
- 54. ПРОДОЛГОВАТЫЙ МОЗГ
- 55. Функции продолговатого мозга Рефлекторная Проводниковая Тоническая
- 56. Ядра продолговатого мозга 1. ЯДРА ЧЕРЕПНОМОЗГОВЫХ НЕРВОВ XII пара - ПОДЪЯЗЫЧНОГО НЕРВА - n. hypoglossus -
- 57. IX пара - ЯЗЫКОГЛОТОЧНОГО НЕРВА: 1) двигательное ядро - рот и глотка 2) чувствительное ядро -
- 58. ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ
- 59. Ядра продолговатого мозга - 2 ПЕРЕКЛЮЧАЮЩИЕ ЯДРА: - Голля и Бурдаха - к таламусу - Ретикулярной
- 60. РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА Жизненно-важные рефлексы Защитные рефлексы Рефлексы поддержания позы Вегетативные рефлексы Вестибуло-вегетативные рефлексы
- 61. Центры продолговатого мозга ЖИЗНЕННОВАЖНЫЕ: ЗАЩИТНЫЕ: Слезоотделения 1) Дыхательный Чиха тельный Кашлевой 2) Сердечно-сосудистый Мигательный Рвотный 3)
- 62. Постуральные рефлексы (поддержания позы) СТАТИЧЕСКИЕ - от рецепторов преддверия - рефлексы положения - рефлексы выпрямления (установочные)
- 63. Постуральные (статические) рефлексы (Р. Магнус): 1. шейные тонические - запускаются при возбуждении проприоцепторов мышц шеи: голова
- 64. 2. вестибулярные тонические рефлексы связаны с возбуждением рецепторов преддверия перепончатого лабиринта, неразрывно связаны с шейными тоническими
- 65. Мост В мосту расположены ядра ЧМН: V пара - тройничный нерв, VI пара -отводящий нерв, VII
- 66. Средний мозг
- 67. Основные ядра среднего мозга Ядра черепномозговых нервов: --- III пара - глазодвигательного нерва --- IV пара
- 68. Схема расположения двигательных центров в стволе мозга (продолговатом мозге, мосте и среднем мозге).
- 69. Двигательные рефлексы среднего мозга: СТАТИЧЕСКИЕ - от рецепторов преддверия - рефлексы выпрямления (установочные) переход животного из
- 70. СТАТОКИНЕТИЧЕСКИЕ – от рецепторов полукружных каналов - рефлексы прямолинейного ускорения - рефлексы углового ускорения
- 71. Красное ядро Участвует в поддержании мышечного тонуса. Тесно связано с мозжечком, вестибулярными ядрами, бледным шаром, ретикулярной
- 72. децеребрационная ригидность В случае перерезки головного мозга ниже красного ядра возникает децеребрационная ригидность, которая проявляется в
- 73. Механизм: ядро Дейтерса находится под тормозным влиянием красного ядра. После перерезки ниже красного ядра тормозное влияние
- 74. Чёрная субстанция Тесно связана с четверохолмием, красным ядром, ретикулярной формацией ствола мозга, таламусом и стриопаллидарной системой.
- 75. медиальный продольный пучок В среднем мозге проходит медиальный продольный пучок , который соединяет между собой в
- 77. водопровод среднего мозга Водопровод среднего мозга окружает центральное серое вещество, относящееся к ретикулярной формации. В её
- 78. четверохолмие Верхние бугры являются подкорковыми центрами зрительного анализатора. Нижние бугры – слухового анализатора. Связаны соответственно с
- 79. Ретикулярная формация среднего мозга
- 80. Ретикулярная формация Филогенетически древняя сердцевина мозга — занимает медиовентральную область продолговатого мозга, моста и среднего мозга.
- 81. Неспецифичность системы Ретикулярная формация — комплекс полисинаптических путей. Аксоны входят в неё не только из восходящих
- 82. Проекции восходящей ретикулярной активирующей системы На рисунке приведена общая система контроля активности мозга. Центральный компонент этой
- 83. Благодаря работам Г. Мегуна и Дж. Моруцци наряду с неспецифическими нисходящими влияниями ретикулярной формации ствола были
- 84. Уровень активности ретикулярной формации и, следовательно, уровень активности большого мозга в значительной мере определяется количеством и
- 85. Стволовые центры, выделяющие нейрогормоны
- 86. Голубое пятно и норадреналиновая система. Нервные волокна из этой области распространяются к базальным ганглиям, обонятельной области,
- 87. Ядра шва и серотонинергическая система. Нервные волокна идут к промежуточному мозгу, коре мозга и спинному мозгу.
- 88. Основные системы ретикулярной формации АФФЕРЕНТНЫЕ СИСТЕМЫ: от спинного мозга, от мозжечка, от четверохолмия, от коры, от
- 89. Схема облегчающих (+) и тормозящих (-) зон РФ и ее связи с корой и подкоркой ТОРМОЗЯЩИЕ
- 90. Афферентные и эфферентные связи ретикулярной формации стволовой части мозга
- 92. Скачать презентацию
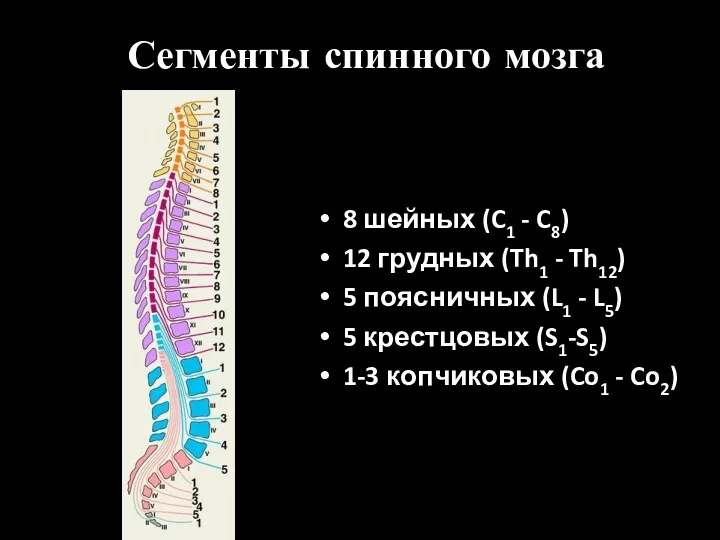
Сегменты спинного мозга
8 шейных (C1 - C8)
12 грудных (Th1 - Th12)
5
Сегменты спинного мозга
8 шейных (C1 - C8)
12 грудных (Th1 - Th12)
5
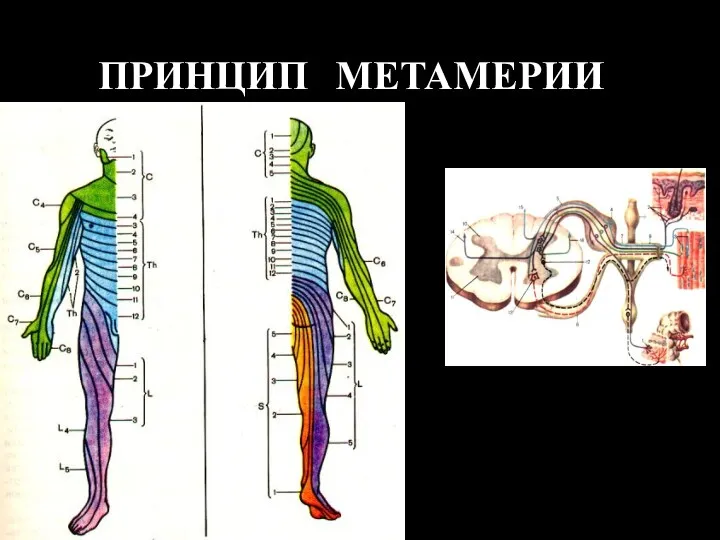
ПРИНЦИП МЕТАМЕРИИ
ПРИНЦИП МЕТАМЕРИИ

Принципы иннервации спинного мозга:
Сегментарный принцип
Морфологических границ между сегментами нет, поэтому
Принципы иннервации спинного мозга:
Сегментарный принцип
Морфологических границ между сегментами нет, поэтому

Закон Белла - Мажанди
Вентральные (передние) корешки содержат эфферентные двигательные (выходящие) волокна,
Закон Белла - Мажанди
Вентральные (передние) корешки содержат эфферентные двигательные (выходящие) волокна,

Нейроны спинного мозга
(13 млн.):
1. Двигательные или мотонейроны (3%):
- альфа-мотонейроны
Нейроны спинного мозга
(13 млн.):
1. Двигательные или мотонейроны (3%):
- альфа-мотонейроны

Нейроны. В сером веществе спинного мозга находятся тела двигательных, вставочных и
Нейроны. В сером веществе спинного мозга находятся тела двигательных, вставочных и

По эффекторному действию:
Возбуждающие (3%)
Тормозные
По эффекторному действию:
Возбуждающие (3%)
Тормозные
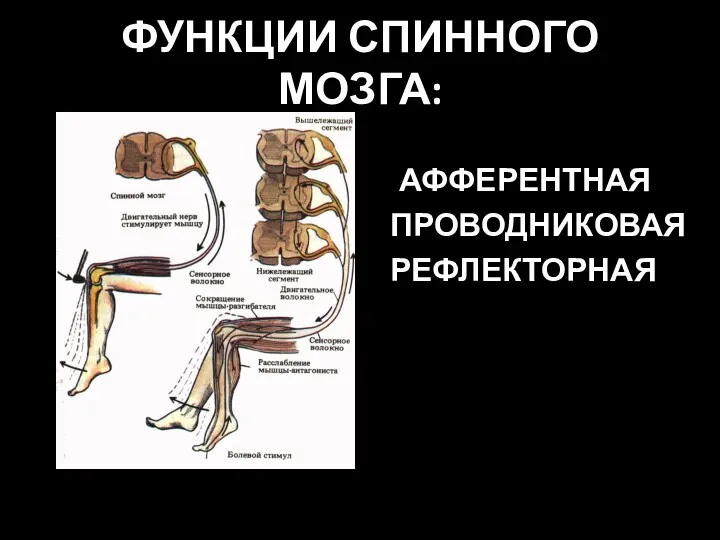
ФУНКЦИИ СПИННОГО МОЗГА:
АФФЕРЕНТНАЯ
ПРОВОДНИКОВАЯ
РЕФЛЕКТОРНАЯ
ФУНКЦИИ СПИННОГО МОЗГА:
АФФЕРЕНТНАЯ
ПРОВОДНИКОВАЯ
РЕФЛЕКТОРНАЯ

Основные рефлексы спинного мозга
Рефлексы растяжения (миотатические) - в основном разгибательные -
Основные рефлексы спинного мозга
Рефлексы растяжения (миотатические) - в основном разгибательные -

Регуляция двигательной активности спинным мозгом
Регуляция фазной активности мышц:
Сгибательные рефлексы
Локомоции (автоматизмы)
Регуляция тонуса
мышц:
Миотатические
Регуляция двигательной активности спинным мозгом
Регуляция фазной активности мышц:
Сгибательные рефлексы
Локомоции (автоматизмы)
Регуляция тонуса
мышц:
Миотатические
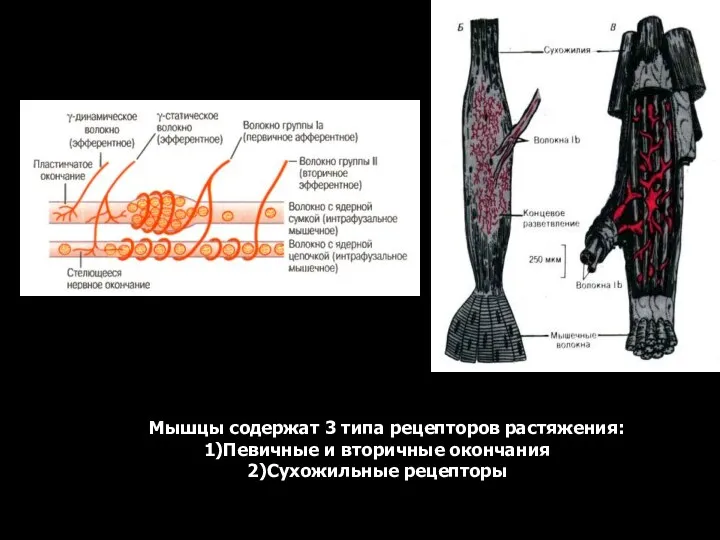
Мышцы содержат 3 типа рецепторов растяжения:
Певичные и вторичные окончания
Сухожильные рецепторы
Мышцы содержат 3 типа рецепторов растяжения:
Певичные и вторичные окончания
Сухожильные рецепторы

Первичные афференты реагирует на степень и скорость растяжения мышц (динамику), а
Первичные афференты реагирует на степень и скорость растяжения мышц (динамику), а
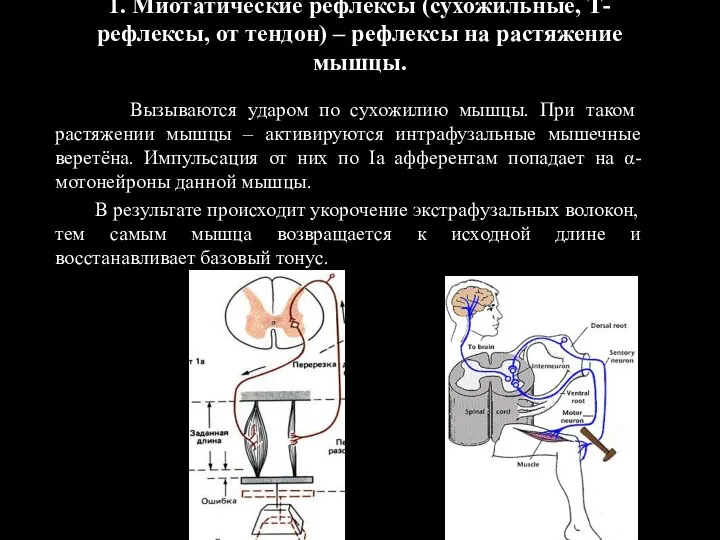
1. Миотатические рефлексы (сухожильные, Т-рефлексы, от тендон) – рефлексы на растяжение
1. Миотатические рефлексы (сухожильные, Т-рефлексы, от тендон) – рефлексы на растяжение

Облегчение Т–рефлексов.
Если коленный и другие Т–рефлексы нижней конечности ослаблены, их можно
Облегчение Т–рефлексов.
Если коленный и другие Т–рефлексы нижней конечности ослаблены, их можно

2. Постуральные рефлексы
(от английск. postur—поза, положение), термин, предложенный Шеррингтоном для обозначения
2. Постуральные рефлексы
(от английск. postur—поза, положение), термин, предложенный Шеррингтоном для обозначения
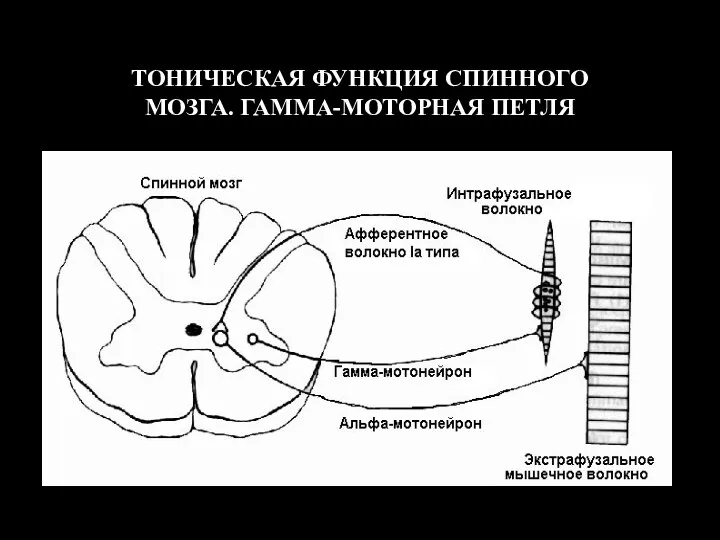
ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ
ТОНИЧЕСКАЯ ФУНКЦИЯ СПИННОГО МОЗГА. ГАММА-МОТОРНАЯ ПЕТЛЯ

Участие спинного мозга в регуляции произвольных движений
От супраспинальных отделов управляющие сигналы
Участие спинного мозга в регуляции произвольных движений
От супраспинальных отделов управляющие сигналы
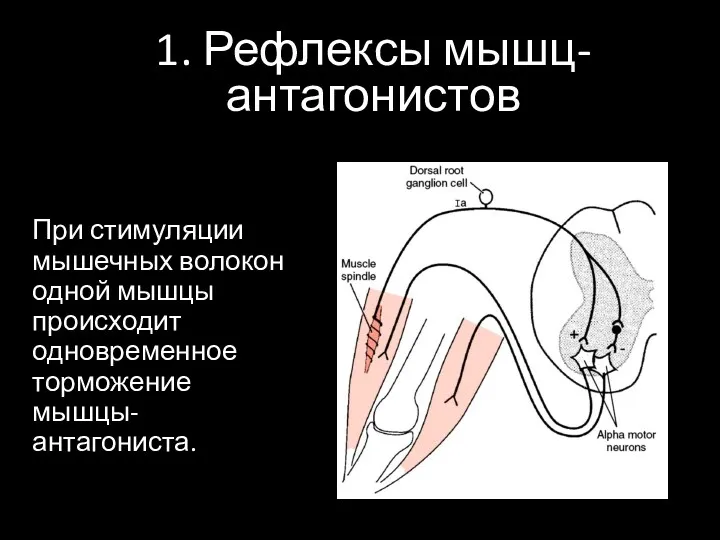
1. Рефлексы мышц-антагонистов
При стимуляции мышечных волокон одной мышцы происходит одновременное торможение
1. Рефлексы мышц-антагонистов
При стимуляции мышечных волокон одной мышцы происходит одновременное торможение
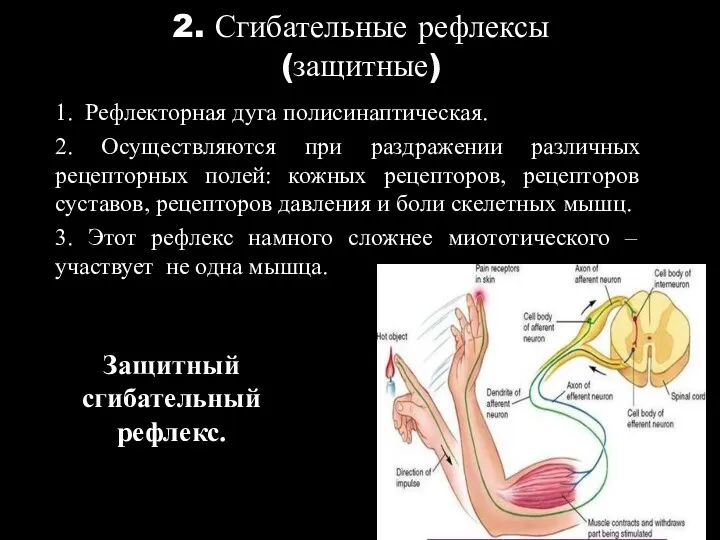
2. Сгибательные рефлексы
(защитные)
1. Рефлекторная дуга полисинаптическая.
2. Осуществляются при раздражении различных рецепторных
2. Сгибательные рефлексы
(защитные)
1. Рефлекторная дуга полисинаптическая.
2. Осуществляются при раздражении различных рецепторных
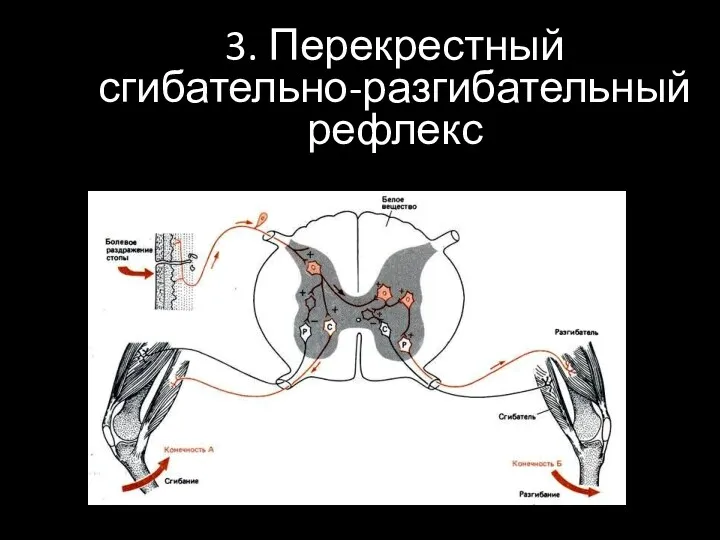
3. Перекрестный сгибательно-разгибательный рефлекс
3. Перекрестный сгибательно-разгибательный рефлекс

Функции сухожильных органов.
Рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы.
У
Функции сухожильных органов.
Рефлекторная дуга сухожильных органов служит для поддержания постоянства напряжения мышцы.
У
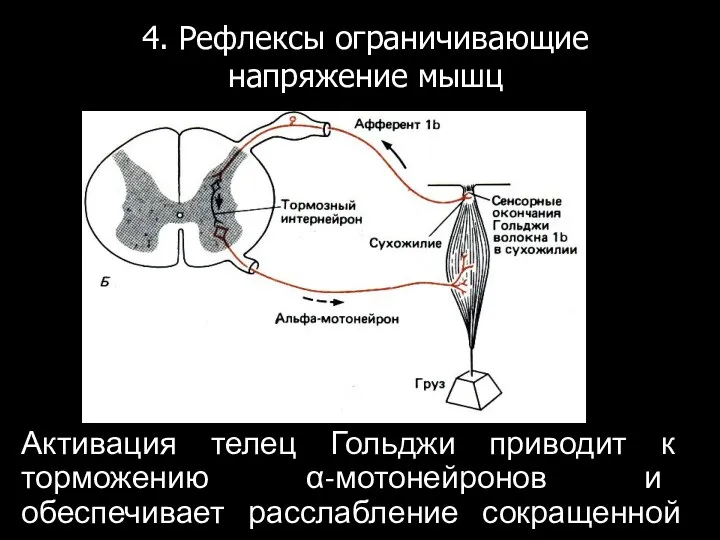
Активация телец Гольджи приводит к торможению α-мотонейронов и обеспечивает расслабление сокращенной
Активация телец Гольджи приводит к торможению α-мотонейронов и обеспечивает расслабление сокращенной

Спинальная локомоция
(автоматизмы)
Основные характеристики локомоции, т.е. перемещения человека или животного в окружающей
Спинальная локомоция
(автоматизмы)
Основные характеристики локомоции, т.е. перемещения человека или животного в окружающей

Активность центров координируется проприоспинальными системами и трактами, пересекающими спинной мозг
Активность центров координируется проприоспинальными системами и трактами, пересекающими спинной мозг

Предполагают, что у человека тоже есть спинальные локомоторные центры.
По–видимому,
По–видимому,

Шагательный автоматизм
Поднимите малыша под мышки ( стопы должны лишь прикасаться к
Шагательный автоматизм
Поднимите малыша под мышки ( стопы должны лишь прикасаться к

СОСАТЕЛЬНЫЙ РЕФЛЕКС
СОСАТЕЛЬНЫЙ РЕФЛЕКС

Автоматизм Бауэра
Когда ребенок лежит на животе, подставьте свою ладонь ему
Автоматизм Бауэра
Когда ребенок лежит на животе, подставьте свою ладонь ему

Рефлекс Бабинского
Грудничок разводит пальчики на ногах веером лишь тогда, когда вы
Рефлекс Бабинского
Грудничок разводит пальчики на ногах веером лишь тогда, когда вы

Рефлекс Моро
Положите малыша на спинку и хлопните ладонями по кровати
Рефлекс Моро
Положите малыша на спинку и хлопните ладонями по кровати

РЕФЛЕКС ГАЛАНТА
Проведите пальцем вдоль позвоночника с правой стороны ( на расстоянии
РЕФЛЕКС ГАЛАНТА
Проведите пальцем вдоль позвоночника с правой стороны ( на расстоянии

Хватательный автоматизм Робинсона.
Угасает этот рефлекс к 4-му месяцу. А на смену ему
Хватательный автоматизм Робинсона.
Угасает этот рефлекс к 4-му месяцу. А на смену ему

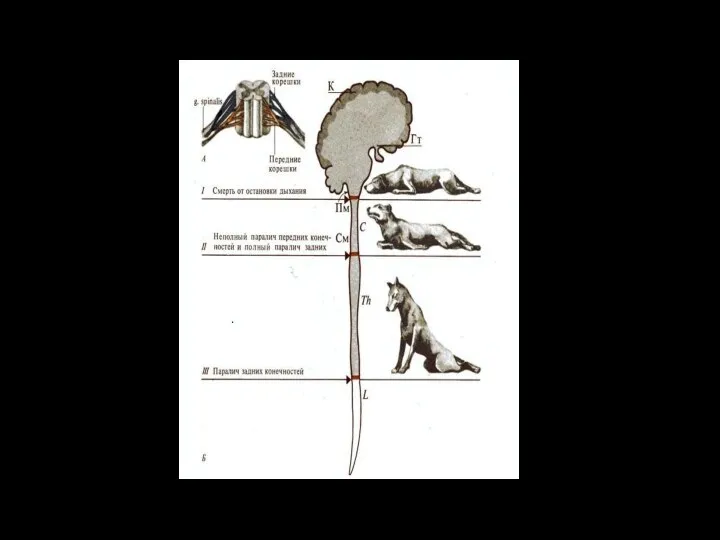
Полная параплегия
возникает при разрыве спинного мозга в грудном отделе–
от Т2
Полная параплегия возникает при разрыве спинного мозга в грудном отделе– от Т2

В последующие недели и месяцы двигательные рефлексы восстанавливаются.
Выделяют следующие четыре стадии:
В последующие недели и месяцы двигательные рефлексы восстанавливаются.
Выделяют следующие четыре стадии:

СПИНАЛЬНЫЙ ШОК
Обратимое угнетение двигательных и вегетативных рефлексов после разрыва спинного мозга
СПИНАЛЬНЫЙ ШОК
Обратимое угнетение двигательных и вегетативных рефлексов после разрыва спинного мозга

Патологические кистевые рефлексы
Сгибательные:
Рефлекс Жуковского — сгибание пальцев кисти при ударе молоточком по
Патологические кистевые рефлексы
Сгибательные:
Рефлекс Жуковского — сгибание пальцев кисти при ударе молоточком по

Патологические стопные рефлексы
Разгибательные:
Рефлекс Оппенгейма — разгибание I пальца стопы при проведении
Патологические стопные рефлексы
Разгибательные:
Рефлекс Оппенгейма — разгибание I пальца стопы при проведении
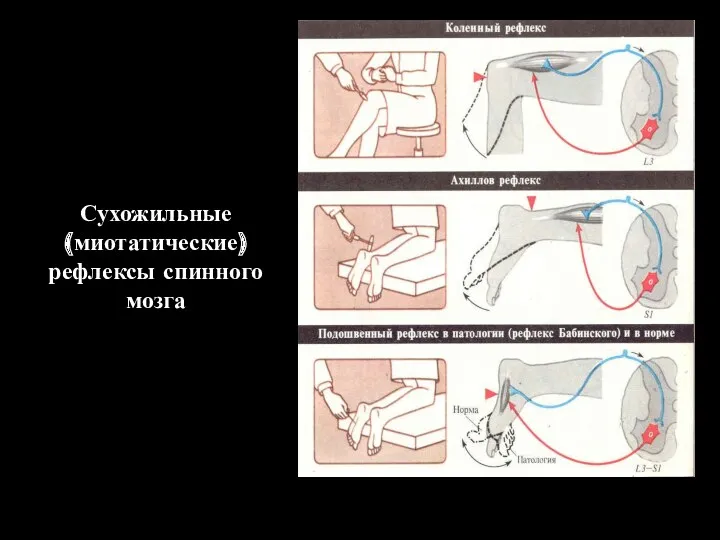
Сухожильные
(миотатические)
рефлексы спинного мозга
Сухожильные
(миотатические)
рефлексы спинного мозга

Сгибательные:
Нижний рефлекс Россолимо — сгибание пальцев стопы при быстром касательном ударе
Сгибательные:
Нижний рефлекс Россолимо — сгибание пальцев стопы при быстром касательном ударе

Частичная параплегия - синдром Броун-Секара
Частичная параплегия - синдром Броун-Секара
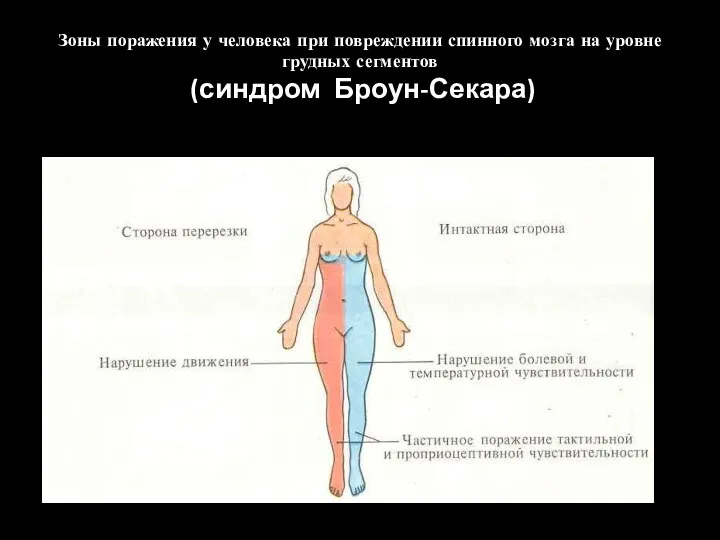
Зоны поражения у человека при повреждении спинного мозга на уровне грудных
Зоны поражения у человека при повреждении спинного мозга на уровне грудных
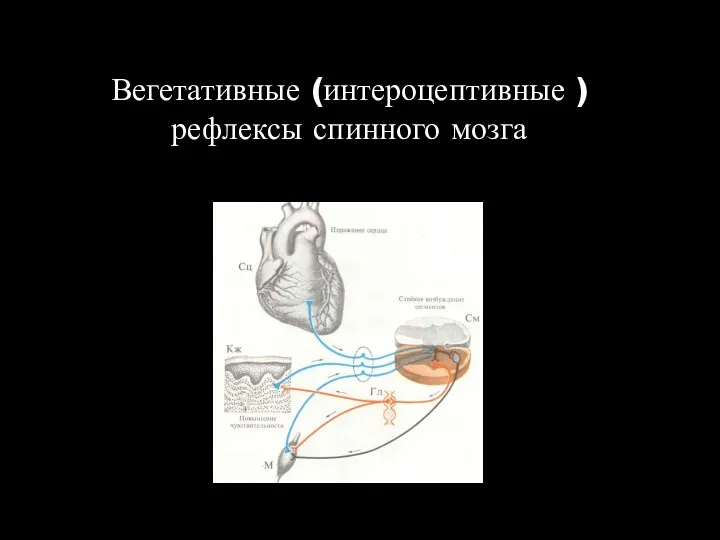
Вегетативные (интероцептивные ) рефлексы спинного мозга
Вегетативные (интероцептивные ) рефлексы спинного мозга

Висцеромоторные рефлексы
Проявляются в двигательных реакциях мышц грудной клетки и брюшной стенки,
Висцеромоторные рефлексы Проявляются в двигательных реакциях мышц грудной клетки и брюшной стенки,
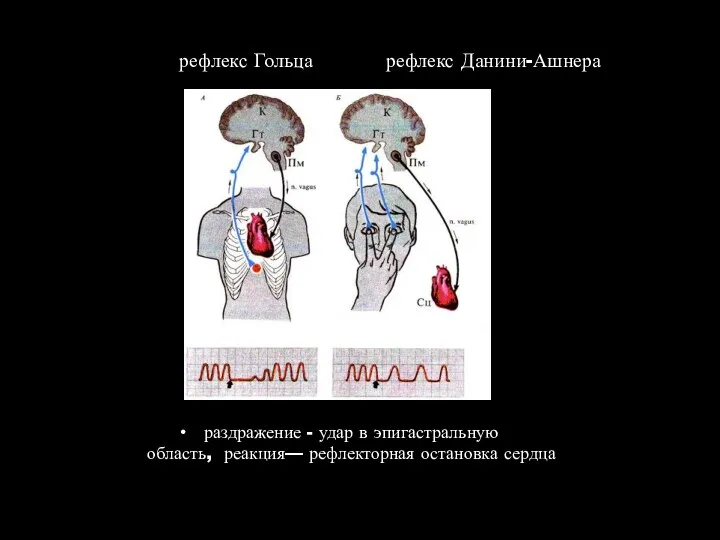
рефлекс Гольца рефлекс Данини-Ашнера
раздражение - удар в эпигастральную область, реакция— рефлекторная остановка сердца
рефлекс Гольца рефлекс Данини-Ашнера
раздражение - удар в эпигастральную область, реакция— рефлекторная остановка сердца
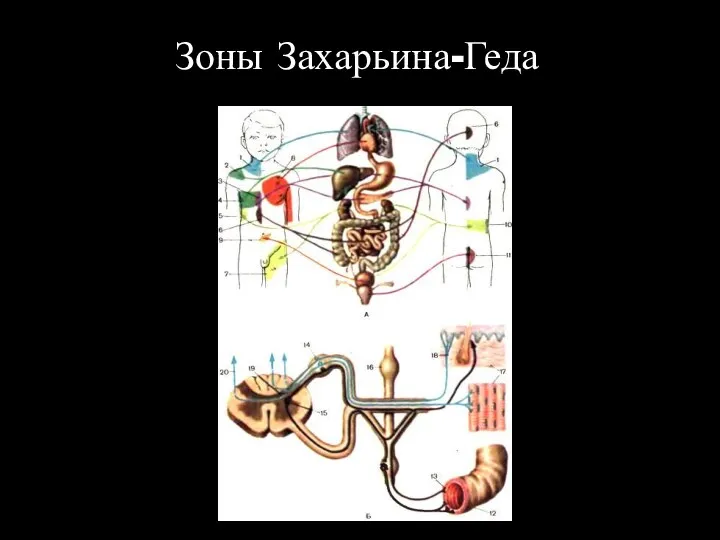
Зоны Захарьина-Геда
Зоны Захарьина-Геда
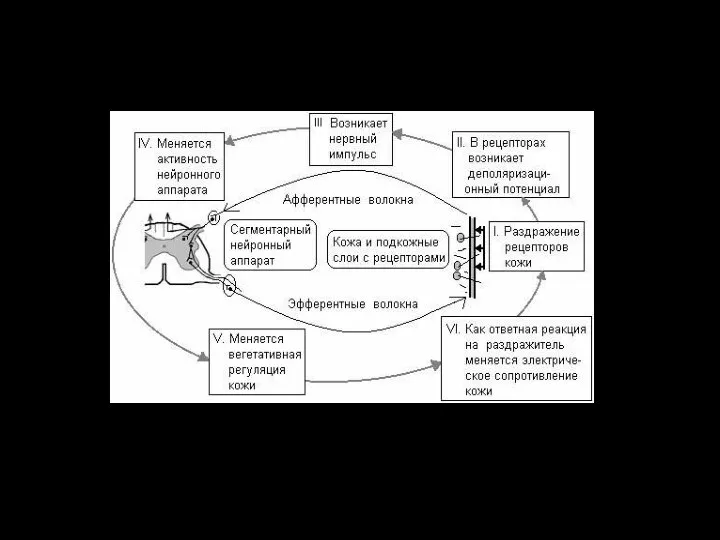

Афферентная функция
Восприятие возбуждений, поступающих от афферентных клеток в ответ на раздражение
Афферентная функция
Восприятие возбуждений, поступающих от афферентных клеток в ответ на раздражение

ПРОВОДЯЩИЕ СИСТЕМЫ
СПИННОГО МОЗГА:
ВОСХОДЯЩИЕ ПУТИ (ЭКСТЕРО- ПРОПРИО-ИНТЕРОЦЕПТИВНАЯ ЧУВСТВИТЕЛЬНОСТЬ)
НИСХОДЯЩИЕ ПУТИ (ЭФФЕКТОРНЫЕ, ДВИГАТЕЛЬНЫЕ)
СОБСТВЕННЫЕ
ПРОВОДЯЩИЕ СИСТЕМЫ
СПИННОГО МОЗГА:
ВОСХОДЯЩИЕ ПУТИ (ЭКСТЕРО- ПРОПРИО-ИНТЕРОЦЕПТИВНАЯ ЧУВСТВИТЕЛЬНОСТЬ)
НИСХОДЯЩИЕ ПУТИ (ЭФФЕКТОРНЫЕ, ДВИГАТЕЛЬНЫЕ)
СОБСТВЕННЫЕ

Тонкий пучок Голля (fasciculus gracilis) - от нижней части тела -
Тонкий пучок Голля (fasciculus gracilis) - от нижней части тела -

Латеральный кортикоспинальный пирамидный тракт - двигательные зоны коры - перекрест в
Латеральный кортикоспинальный пирамидный тракт - двигательные зоны коры - перекрест в
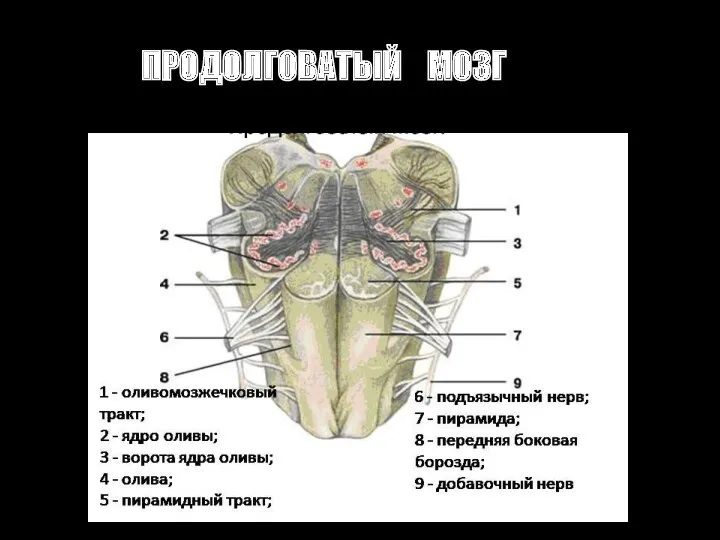
ПРОДОЛГОВАТЫЙ МОЗГ
ПРОДОЛГОВАТЫЙ МОЗГ

Функции продолговатого мозга
Рефлекторная
Проводниковая
Тоническая
Функции продолговатого мозга
Рефлекторная
Проводниковая
Тоническая

Ядра продолговатого мозга
1. ЯДРА ЧЕРЕПНОМОЗГОВЫХ НЕРВОВ
XII пара - ПОДЪЯЗЫЧНОГО НЕРВА -
Ядра продолговатого мозга
1. ЯДРА ЧЕРЕПНОМОЗГОВЫХ НЕРВОВ
XII пара - ПОДЪЯЗЫЧНОГО НЕРВА -

IX пара - ЯЗЫКОГЛОТОЧНОГО НЕРВА:
1) двигательное ядро - рот и
IX пара - ЯЗЫКОГЛОТОЧНОГО НЕРВА:
1) двигательное ядро - рот и

ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ
ЧЕРЕПНО-МОЗГОВЫЕ НЕРВЫ

Ядра продолговатого мозга - 2
ПЕРЕКЛЮЧАЮЩИЕ ЯДРА:
- Голля и Бурдаха -
Ядра продолговатого мозга - 2
ПЕРЕКЛЮЧАЮЩИЕ ЯДРА:
- Голля и Бурдаха -

РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА
Жизненно-важные рефлексы
Защитные рефлексы
Рефлексы поддержания позы
Вегетативные рефлексы
Вестибуло-вегетативные рефлексы
РЕФЛЕКСЫ ПРОДОЛГОВАТОГО МОЗГА
Жизненно-важные рефлексы
Защитные рефлексы
Рефлексы поддержания позы
Вегетативные рефлексы
Вестибуло-вегетативные рефлексы

Центры продолговатого мозга
ЖИЗНЕННОВАЖНЫЕ: ЗАЩИТНЫЕ:
Слезоотделения
1) Дыхательный Чиха тельный
Кашлевой
Центры продолговатого мозга
ЖИЗНЕННОВАЖНЫЕ: ЗАЩИТНЫЕ:
Слезоотделения
1) Дыхательный Чиха тельный
Кашлевой

Постуральные рефлексы (поддержания позы)
СТАТИЧЕСКИЕ - от рецепторов преддверия
- рефлексы положения
Постуральные рефлексы (поддержания позы)
СТАТИЧЕСКИЕ - от рецепторов преддверия
- рефлексы положения

Постуральные (статические) рефлексы (Р. Магнус):
1. шейные тонические - запускаются при возбуждении
Постуральные (статические) рефлексы (Р. Магнус):
1. шейные тонические - запускаются при возбуждении

2. вестибулярные тонические рефлексы связаны с возбуждением рецепторов преддверия перепончатого лабиринта,
2. вестибулярные тонические рефлексы связаны с возбуждением рецепторов преддверия перепончатого лабиринта,

Мост
В мосту расположены ядра ЧМН:
V пара - тройничный нерв,
VI
Мост
В мосту расположены ядра ЧМН:
V пара - тройничный нерв,
VI
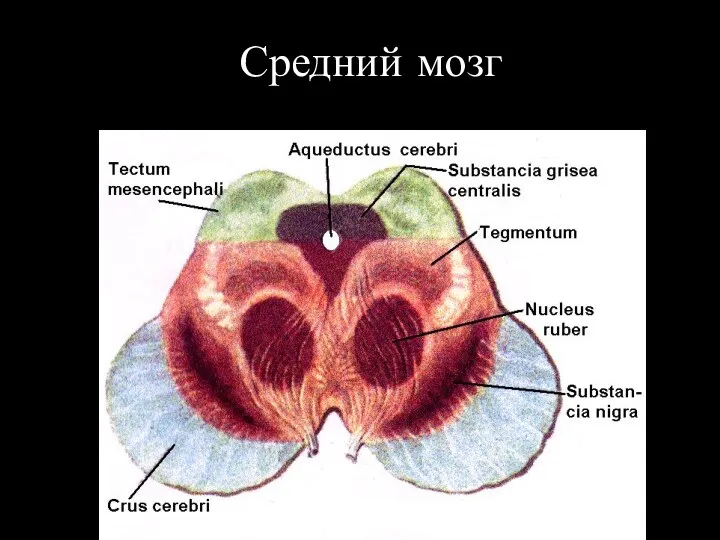
Средний мозг
Средний мозг

Основные ядра среднего мозга
Ядра черепномозговых нервов:
--- III пара -
Основные ядра среднего мозга
Ядра черепномозговых нервов:
--- III пара -
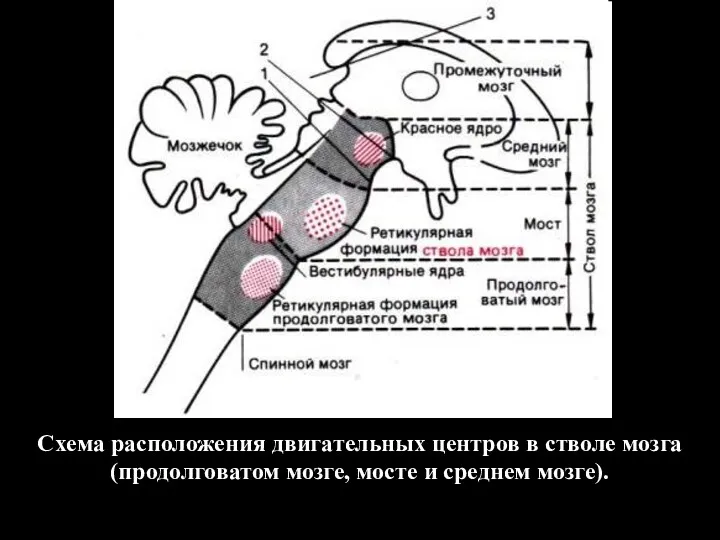
Схема расположения двигательных центров в стволе мозга (продолговатом мозге, мосте и
Схема расположения двигательных центров в стволе мозга (продолговатом мозге, мосте и

Двигательные рефлексы среднего мозга:
СТАТИЧЕСКИЕ - от рецепторов преддверия - рефлексы выпрямления
Двигательные рефлексы среднего мозга:
СТАТИЧЕСКИЕ - от рецепторов преддверия - рефлексы выпрямления

СТАТОКИНЕТИЧЕСКИЕ –
от рецепторов полукружных каналов
- рефлексы прямолинейного ускорения
- рефлексы углового
СТАТОКИНЕТИЧЕСКИЕ –
от рецепторов полукружных каналов
- рефлексы прямолинейного ускорения
- рефлексы углового

Красное ядро
Участвует в поддержании мышечного тонуса. Тесно связано с мозжечком, вестибулярными
Красное ядро
Участвует в поддержании мышечного тонуса. Тесно связано с мозжечком, вестибулярными

децеребрационная ригидность
В случае перерезки головного мозга ниже красного ядра возникает децеребрационная
децеребрационная ригидность
В случае перерезки головного мозга ниже красного ядра возникает децеребрационная
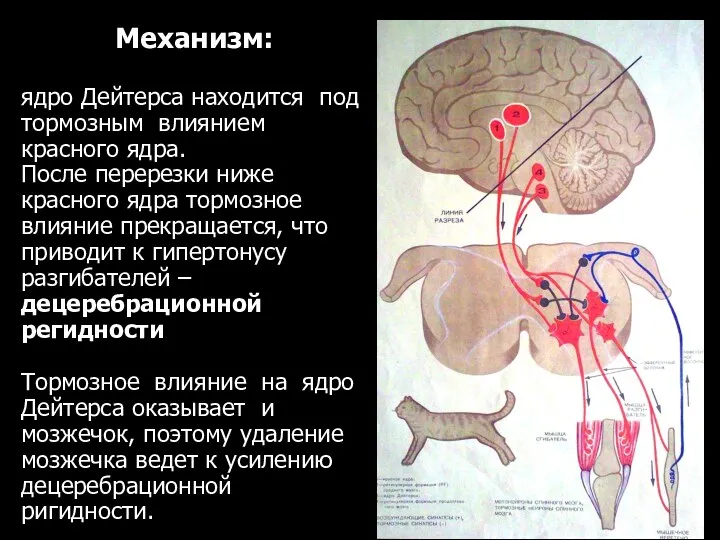
Механизм:
ядро Дейтерса находится под тормозным влиянием красного ядра.
После перерезки ниже
Механизм:
ядро Дейтерса находится под тормозным влиянием красного ядра.
После перерезки ниже

Чёрная субстанция
Тесно связана с четверохолмием, красным ядром, ретикулярной формацией ствола мозга,
Чёрная субстанция
Тесно связана с четверохолмием, красным ядром, ретикулярной формацией ствола мозга,

медиальный продольный пучок
В среднем мозге проходит медиальный продольный пучок , который
медиальный продольный пучок
В среднем мозге проходит медиальный продольный пучок , который

водопровод среднего мозга
Водопровод среднего мозга окружает центральное серое вещество, относящееся к
водопровод среднего мозга
Водопровод среднего мозга окружает центральное серое вещество, относящееся к

четверохолмие
Верхние бугры являются подкорковыми центрами зрительного анализатора.
Нижние бугры – слухового анализатора.
четверохолмие
Верхние бугры являются подкорковыми центрами зрительного анализатора.
Нижние бугры – слухового анализатора.
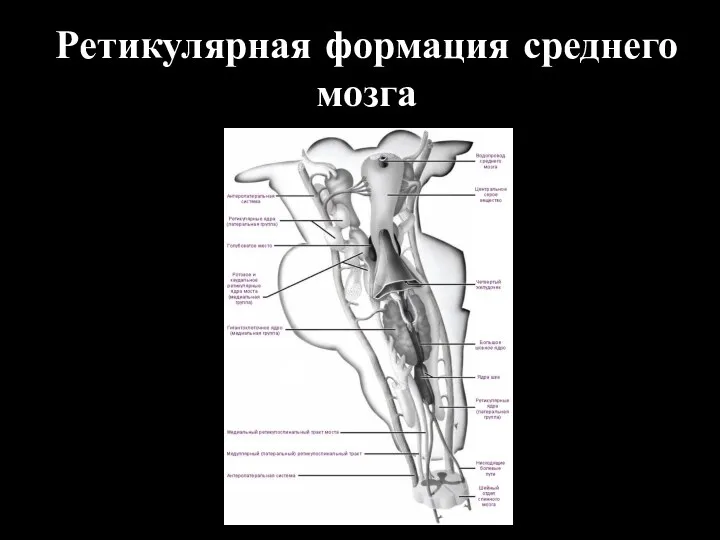
Ретикулярная формация среднего мозга
Ретикулярная формация среднего мозга

Ретикулярная формация
Филогенетически древняя сердцевина мозга — занимает медиовентральную область продолговатого мозга,
Ретикулярная формация
Филогенетически древняя сердцевина мозга — занимает медиовентральную область продолговатого мозга,

Неспецифичность системы
Ретикулярная формация — комплекс полисинаптических путей. Аксоны входят в неё не
Неспецифичность системы
Ретикулярная формация — комплекс полисинаптических путей. Аксоны входят в неё не
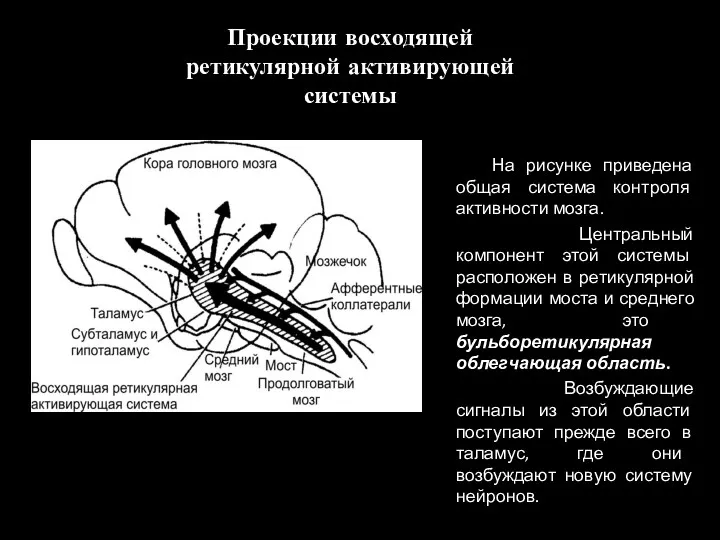
Проекции восходящей ретикулярной активирующей системы
На рисунке приведена общая система контроля
Проекции восходящей ретикулярной активирующей системы
На рисунке приведена общая система контроля
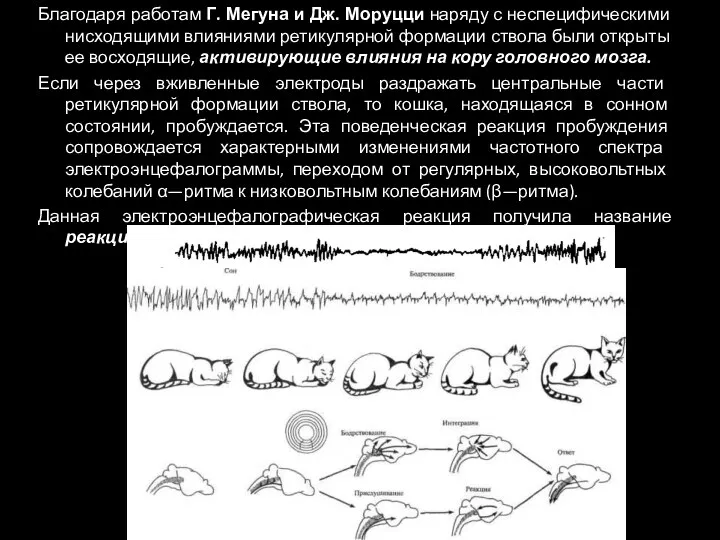
Благодаря работам Г. Мегуна и Дж. Моруцци наряду с неспецифическими нисходящими
Благодаря работам Г. Мегуна и Дж. Моруцци наряду с неспецифическими нисходящими

Уровень активности ретикулярной формации и, следовательно, уровень активности большого мозга в
Уровень активности ретикулярной формации и, следовательно, уровень активности большого мозга в
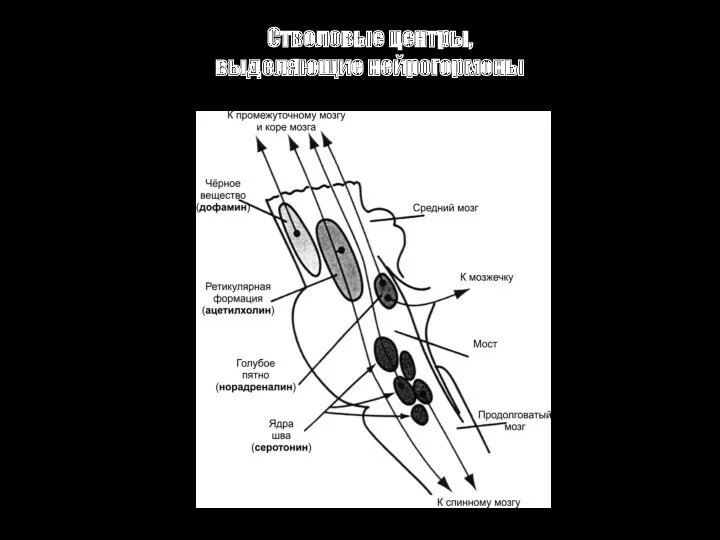
Стволовые центры, выделяющие нейрогормоны
Стволовые центры, выделяющие нейрогормоны

Голубое пятно и норадреналиновая система. Нервные волокна из этой области распространяются
Голубое пятно и норадреналиновая система. Нервные волокна из этой области распространяются

Ядра шва и серотонинергическая система.
Нервные волокна идут к промежуточному
Ядра шва и серотонинергическая система.
Нервные волокна идут к промежуточному

Основные системы ретикулярной формации
АФФЕРЕНТНЫЕ СИСТЕМЫ:
от спинного мозга, от мозжечка, от четверохолмия,
Основные системы ретикулярной формации
АФФЕРЕНТНЫЕ СИСТЕМЫ:
от спинного мозга, от мозжечка, от четверохолмия,
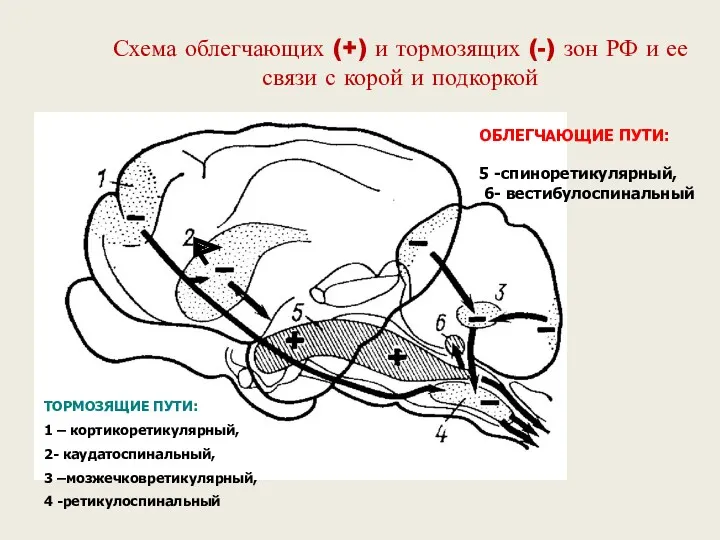
Схема облегчающих (+) и тормозящих (-) зон РФ и ее связи
Схема облегчающих (+) и тормозящих (-) зон РФ и ее связи
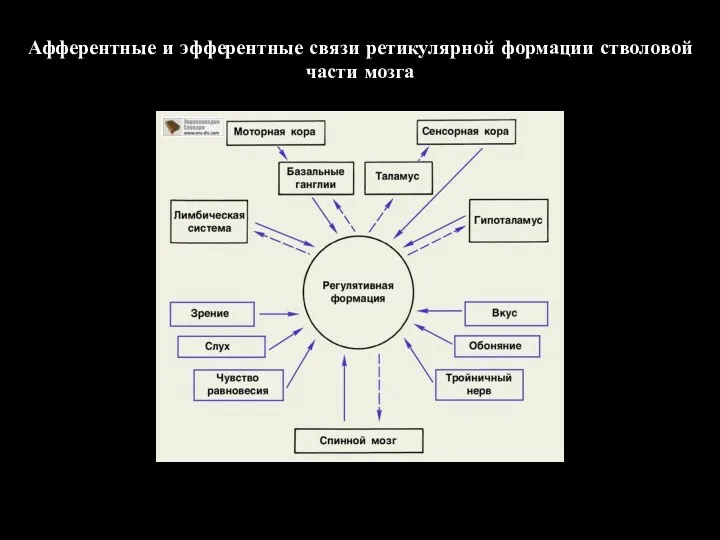
Афферентные и эфферентные связи ретикулярной формации стволовой части мозга
Афферентные и эфферентные связи ретикулярной формации стволовой части мозга
 Клиническая генетика
Клиническая генетика Аяқ қол хирургиялық аурулары,туа біткен ақаулары және жарақаттары
Аяқ қол хирургиялық аурулары,туа біткен ақаулары және жарақаттары Зоонозды инфекция (сарып, ирсиниоз және листериоз )
Зоонозды инфекция (сарып, ирсиниоз және листериоз ) Модификации сухожильных швов
Модификации сухожильных швов Заболевания мочевыделительной системы у беременных
Заболевания мочевыделительной системы у беременных Консервативное лечение заболеваний щитовидной железы
Консервативное лечение заболеваний щитовидной железы Лекарственные препараты при беременности, возможный вред при их применении
Лекарственные препараты при беременности, возможный вред при их применении Повреждения острыми предметами. Огнестрельная травма
Повреждения острыми предметами. Огнестрельная травма Лимфатическая система ( systema lymphaticum)
Лимфатическая система ( systema lymphaticum) Dental clinics
Dental clinics Некротизирующий энтероколит и спонтанная перфорация кишки. MHO
Некротизирующий энтероколит и спонтанная перфорация кишки. MHO Профили суточной динамики АД и их клиническое значение
Профили суточной динамики АД и их клиническое значение Пиелонефрит. Классификация
Пиелонефрит. Классификация Гомеопатия
Гомеопатия Техника омоложения кожи
Техника омоложения кожи Риккетсии. Риккетсиозы
Риккетсии. Риккетсиозы Лептоспироз. История изучения. Экономический ущерб. Распространение. Возбудитель
Лептоспироз. История изучения. Экономический ущерб. Распространение. Возбудитель Медико-санитарное обеспечение при ликвидации последствий чрезвычайных ситуаций природного характера
Медико-санитарное обеспечение при ликвидации последствий чрезвычайных ситуаций природного характера Иммунитет. Виды иммунитета
Иммунитет. Виды иммунитета Кишечная инфекция – эшерихии
Кишечная инфекция – эшерихии Краснуха
Краснуха Антиоксиданты, витамины, минералы
Антиоксиданты, витамины, минералы Местное обезболивание в стоматологии
Местное обезболивание в стоматологии Узкий таз. Определение узкого таза у женщин
Узкий таз. Определение узкого таза у женщин Нормальная анатомия позвоночного столба
Нормальная анатомия позвоночного столба Реализация регионального проекта Укрепление общественного здоровья
Реализация регионального проекта Укрепление общественного здоровья Реализация региональных проектов национального проекта Здравоохранение в Липецкой области
Реализация региональных проектов национального проекта Здравоохранение в Липецкой области Μυοκαρδίτιδες. Ορισμός, επιδημιολογία
Μυοκαρδίτιδες. Ορισμός, επιδημιολογία