- Трансдукция температурного сигнала. TRP ионные каналы

Содержание
- 2. Dorsal horn Ventral horn Efferent (motor) Low-threshold mechanoreceptor Afferent (sensory ) Temperature and pain receptors Proprioceptors
- 3. Терморецепторы – немиелинизированные, неинкапсулированные свободные терминали аксона, содержащие многочисленные митохондрии. Hensel, 1973
- 4. Повышение температуры Внутренняя поверхность мембраны Диффузия K+/Na+ насос Понижение температуры
- 5. Мембрана быстро заморожена до температуры жидкого азота и вскрыта сколом. Плоскость скола проходит через середину липидного
- 6. TRP (Transient Receptor Potential) каналы, как и другие ионные каналы, служат для осуществления реакции клетки на
- 8. TRPС (canonical) TRPP (polycystin) TRPV (vanilloid) TRPML (mucolipin) TRPM (melastatin) TRPA (ankyrin transmembrane protein) Селективные для
- 9. TRP каналы различны по способу их активации Напряжение Концентрация внутриклеточного Са2+, Температура, рН, Осмолярность, Механический стимул,
- 10. Fig. 1 Diagram portraying 3 different molecular roles of TRP channels in the digestive system: (1)
- 11. Сигнальная система G-белка в мембране клетки Скорость движения G-белка в мембране 0.1 - 0.001 мкм/сек.
- 12. Термочувствительные TRP ионные каналы – молекулярная основа терморецепции.
- 13. Термочувствительные TRP ионные каналы Шесть идентифицированных термочувствительных TRР каналов характеризуются необычно высокой температурной чувствительностью (Q10>10-20). Все
- 14. Мята, ментол – TRPM8 Жгучий перец - TRPV1 Камфора - TRPV3 Чеснок, горчичное масло - TRPA1
- 15. Термочувствительный TRP- канал, образует пору между 5 и 6 доменом Температура модулирует работу многих ионных каналов.
- 16. Образование ионного канала
- 17. Встраивание ионных каналов в мембрану клетки
- 18. Анандомин горчичное масло эвкалипт Природные агонисты термочувствительных TRP ионных каналов
- 19. Имеет температурный порог активации 28°С с увеличивающимся током вплоть до 8°С. Преимущественно Са2+ канал. Биофизические свойства
- 20. TRPM8 активируется ментолом, эвкалиптом, ицилином. TRPM8 угнетается капсазепином, снижением рН Мята Терпеновый спирт
- 21. Структура TRPM8
- 22. TRPM8 экспрессируется на нейронах заднекорешковых ганглиев у 15% сенсорных нейронов малого диаметра, что сходно с %
- 23. McKemy et al., 2002 Холод и ментол имеют сходный механизм действия, через активацию Са(2+) канала TRPM8,
- 24. Single channel recordings of TRPM8 channels
- 25. Активация TRPM8 требует присутствия PIP2 PIP2 – phosphatidylinositol-4,5biphosphate Liu, Qin, 2005 PIP2 PIP2
- 26. Активация рецептора ведет через G-белок связывающую систему к активации Са(2+)-зависимой фосфолипазы С (или протеинкиназы С), которая
- 27. TRPА1 активируется холодом при более низкой температуре Неселективный катионный канал, не чувствителен к ментолу. Кроме температуры
- 28. Figure 2.Topology diagram of functional domains of TRPA1. Topology diagram of TRPA1, highlighting domains implicated in
- 29. Cold sensitive Warm sensitive TRPA1
- 30. The divergent thermal sensitivity of TRPA1 channels. TRPA1 is heat-sensitive in invertebrate and ancestral vertebrate (e.g.,
- 31. Активируется температурой >42°С. Известен, как ваниллоидный рецептор. Высоко чувствителен к температуре, Q10>20.6. Неселективный катионный канал, проницаем
- 32. TRPM8 ментол TRPA1 горчица, корица TRPV1 жгучий перец
- 33. Активируется: капсаицином, низким рН (в противоположность TRPM8). Сенситизируется также факторами, участвующими в воспалении: NGF (фактор роста
- 34. Лидокаин блокирует боль и тактильную чувствительность через закрытие натриевых каналов. Если активировать TRP каналы капсаицином, то
- 35. Участие TRPV1 в температурной гипералгезии У мышей с дефицитом TRPV1 наблюдается уменьшение температурной гипералгезии при воспалении.
- 36. реагирует на температуру >52°С не активируется капсаицином и низкой рН активируется NGF (у мышей) блокируется рутением
- 37. Реагирует при температуре 31-39°С. Кроме температуры активируется камфорой. Обнаруживает сильную сенситизацию при повторяющихся тепловых стимулах, увеличивая
- 38. Активируется при 27-34°С. В диапазоне до 42°С обогрев вызывает увеличение тока. Кроме температуры активируется анандомином (метаболит
- 39. Известно, что существуют 2 типа терморецепторов – холодовые и тепловые. Детектор температуры встроен в структуру ионного
- 40. В 2003 году Нобелевская премия по химии была присуждена американскому учёному Родерику Маккиннону (Roderick McKinnon) за
- 42. Созвездие компонентов холодочувствительного нейрона
- 43. С открытием TRP каналов стало возможным понимание молекулярной основы термочувствительности и ее кодирования. Понимание, физиологического значения
- 44. Влияет ли агонист ионного канала TRPM8 ментол на активность и количество функционирующих холодовых афферентов?
- 45. А = f (T°, dT°/dt) * N (ρ, S) Т° - temperature dT°/dt - rate of
- 46. Schafer, Braun, Isenberg, 1986 Влияние ментола на импульсную активность холодового рецептора кожи
- 47. Влияние 1% ментола на количество холодовых точек у человека Влияние охлаждения на количество холодовых точек у
- 48. * Синонимичная замена гуанина (G119662G) на цитозин (G119662С) в 6 экзоне гена, кодирующего ионный канал TRPM8,
- 49. Группа G119662C реагирует снижением минутного объема дыхания (МОД) на ментол и охлаждение, тогда как группа G119662G
- 50. Экспрессия генов разных TRP ионных каналов в структурах мозга в норме и при длительной адаптации организма
- 51. 39°C Адаптация к холоду снижает экспрессию гена ионного канала TRPV3 в гипоталамусе. Изменение экспрессии генов термочувствительных
- 53. Каково влияние активации холодочувствительного ионного канала TRPM8 в коже на терморегуляторные и иммунные показатели?
- 54. В термонейтральных условиях активация ионного канала TRPM8 приводит к повышению потребления кислорода и снижению дыхательного коэффициента,
- 55. Активация ионного канала TRPM8 ментолом изменяет параметры иммунного ответа в термонейтральных условиях.
- 56. Активация ионного канала TRPM8 ментолом в термонейтральных условиях, как и холод, увеличивает уровень IL-6 и L-1β
- 57. Под влиянием холода у гипертензивных животных не увеличивается концентрация провоспалительных цитокинов в крови, тогда как у
- 58. У гипертензивных животных изменена экспрессия генов термочувствительных каналов в гипоталамусе и селезенке, что может являться молекулярной
- 59. Активация ионного канала TRPM8 ментолом не приводит к повышению в крови провоспалительных цитокинов у гипертензивных животных,
- 60. Как активация холодо-чувствительного ионного канала TRPM8 влияет на терморегуляторные и иммунные показатели при действии холода на
- 61. Активация ионного канала TRPM8 приводит к снижению температурных порогов терморегуляторных реакций и улучшению поддержания глубокой температуры
- 62. Активация ионного канала TRPM8 ментолом ослабляет угнетающий эффект глубокого охлаждения на иммунный ответ Антигенсвязывание Антителообразование
- 63. Каково влияние острого охлаждения и активации холодочувствительного ионного канала TRPM8 в коже на экспрессию генов TRP
- 64. Итак, активация холодочувствительного ионного каналаTRPМ8 оказывает существенное влияние на регуляцию жирового обмена, терморегуляторных и иммунных показателей
- 65. N=-25.9+26.2PCa r = 0.62 Козырева ,Тихонова, Ткаченко, Синдаровская, 1987 Влияние ионофореза Ca(2+) в кожу на количество
- 66. Длительная адаптация организма к холоду приводит к снижению концентрации Са(2+) в крови и уменьшению количества чувствительных
- 67. Прослеживается сходство в изменении параметров иммунного ответа под влиянием активации TRPM8, ионов Са(2+) и охлаждения кожи
- 68. Агонист TRPV1 (капсаицин) увеличивает импульсную активность теплового рецептора кожи Capsaicin 10 -7 Kozyreva, Pierau, Neurophysiology, 1999
- 69. Агонист TRPV1 (капсаицин) угнетает импульсную активность холодового рецептора кожи Kozyreva, Pierau, Neurophysiology, 1999
- 70. Под влиянием капсаицина – агониста TRPV1, количество холодовых точек уменьшалось, тогда как количество тепловых точек оставалось
- 71. Козырева, Ткаченко, Елисеева, Симонова, Козарук, Гонсалес, Ломакина, 2004
- 72. Ионные каналы в эволюции TRP ионные каналы обнаружены у тетрапод, рыб, млекопитающих.
- 74. Температура модулирует работу многих ионных каналов. ТермоTRP- каналы активируются непосредственно температурой с высоким Q10. Отдельные TRP-
- 75. Kozyreva, 1984,2007 Dependence of the number of cold spots on the skin temperature
- 78. Figure 1.Temperature Sensitive (TRP) Ion Channels. (A) The cartoon illustrates the different ion-selectivity and current directionality
- 79. Figure 3 Erika Yamamoto-Kasai, advance online publication 05 April 2012. doi:10.1038/jid.2012.97 Lower nerve response to acetone
- 80. The divergent thermal sensitivity of TRPA1 channels. TRPA1 is heat-sensitive in invertebrate and ancestral vertebrate (e.g.,
- 82. Скачать презентацию
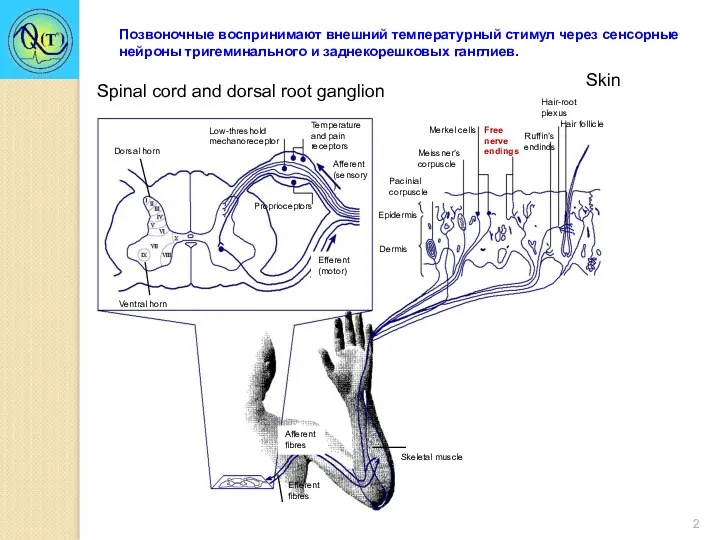
Dorsal horn
Ventral horn
Efferent (motor)
Low-threshold mechanoreceptor
Afferent (sensory )
Temperature and pain receptors
Proprioceptors
Epidermis
Spinal cord
Dorsal horn
Ventral horn
Efferent (motor)
Low-threshold mechanoreceptor
Afferent (sensory )
Temperature and pain receptors
Proprioceptors
Epidermis
Spinal cord
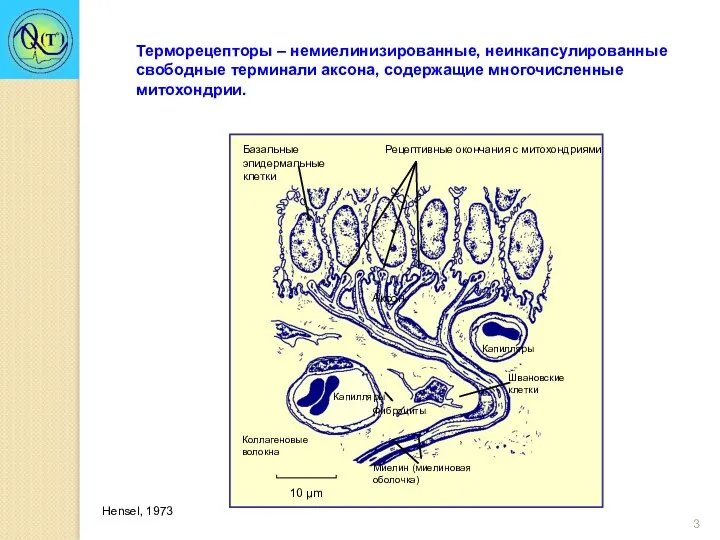
Терморецепторы – немиелинизированные, неинкапсулированные свободные терминали аксона, содержащие многочисленные митохондрии.
Hensel, 1973
Терморецепторы – немиелинизированные, неинкапсулированные свободные терминали аксона, содержащие многочисленные митохондрии.
Hensel, 1973
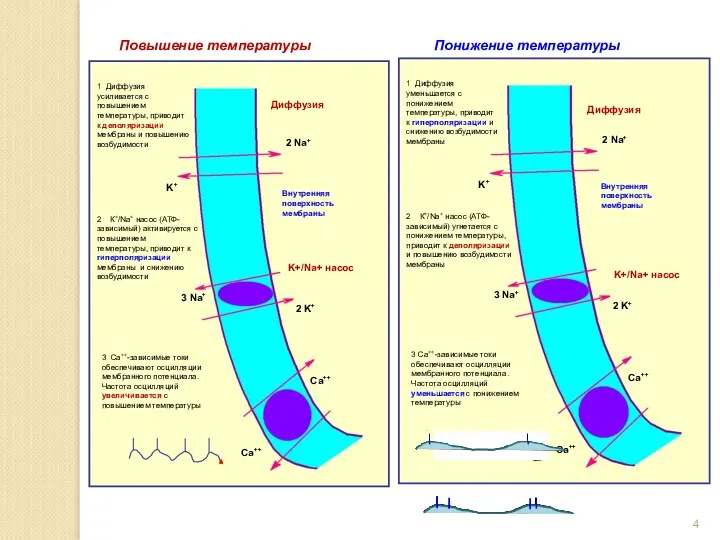
Повышение температуры
Внутренняя поверхность мембраны
Диффузия
K+/Na+ насос
Понижение температуры
Повышение температуры
Внутренняя поверхность мембраны
Диффузия
K+/Na+ насос
Понижение температуры
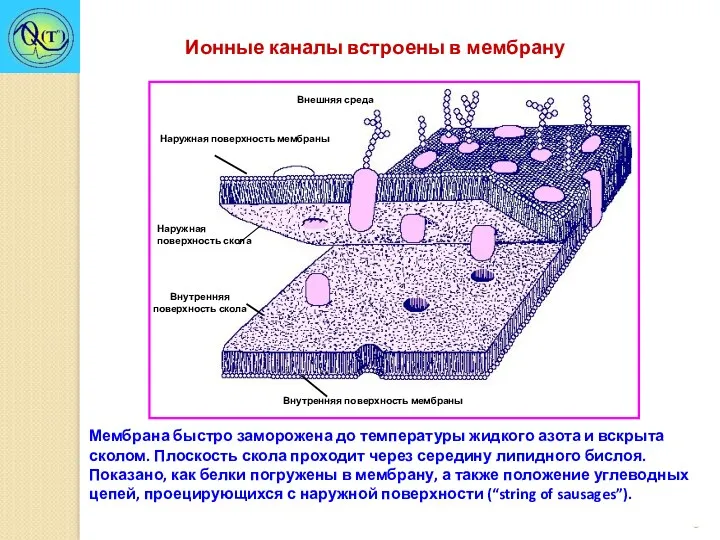
Мембрана быстро заморожена до температуры жидкого азота и вскрыта сколом. Плоскость
Мембрана быстро заморожена до температуры жидкого азота и вскрыта сколом. Плоскость

TRP (Transient Receptor Potential) каналы, как и другие ионные каналы, служат
TRP (Transient Receptor Potential) каналы, как и другие ионные каналы, служат


TRPС (canonical) TRPP (polycystin)
TRPV (vanilloid) TRPML (mucolipin)
TRPM (melastatin) TRPA (ankyrin transmembrane
TRPС (canonical) TRPP (polycystin)
TRPV (vanilloid) TRPML (mucolipin)
TRPM (melastatin) TRPA (ankyrin transmembrane

TRP каналы различны по способу их активации
Напряжение
Концентрация внутриклеточного Са2+,
TRP каналы различны по способу их активации
Напряжение
Концентрация внутриклеточного Са2+,
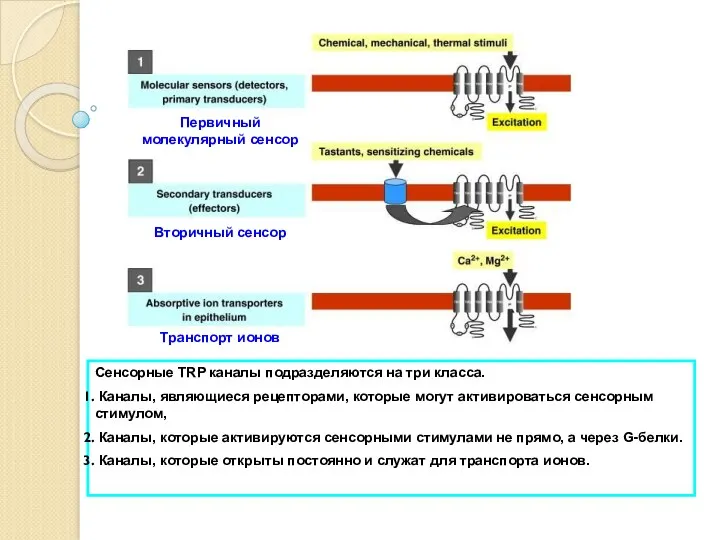
Fig. 1
Diagram portraying 3 different molecular roles of TRP channels in the
Fig. 1
Diagram portraying 3 different molecular roles of TRP channels in the
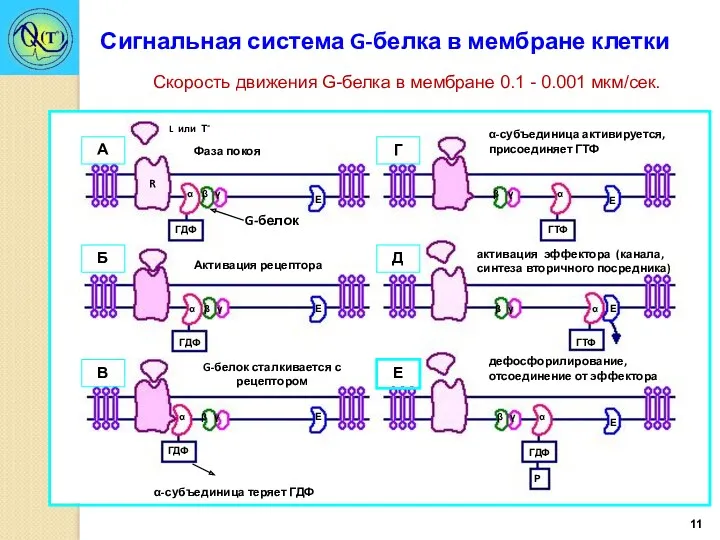
Сигнальная система G-белка в мембране клетки
Скорость движения G-белка в мембране 0.1
Сигнальная система G-белка в мембране клетки
Скорость движения G-белка в мембране 0.1

Термочувствительные TRP ионные каналы
– молекулярная основа терморецепции.
– молекулярная основа терморецепции.
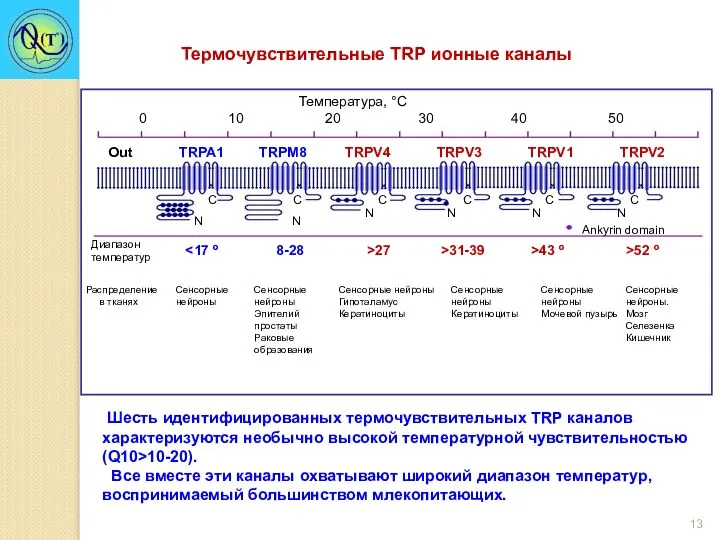
Термочувствительные TRP ионные каналы
Шесть идентифицированных термочувствительных TRР каналов характеризуются необычно
Термочувствительные TRP ионные каналы
Шесть идентифицированных термочувствительных TRР каналов характеризуются необычно

Мята, ментол – TRPM8
Жгучий перец - TRPV1
Камфора - TRPV3
Чеснок, горчичное масло
Мята, ментол – TRPM8
Жгучий перец - TRPV1
Камфора - TRPV3
Чеснок, горчичное масло

Термочувствительный TRP- канал, образует пору между 5 и 6 доменом
Температура модулирует
Термочувствительный TRP- канал, образует пору между 5 и 6 доменом
Температура модулирует
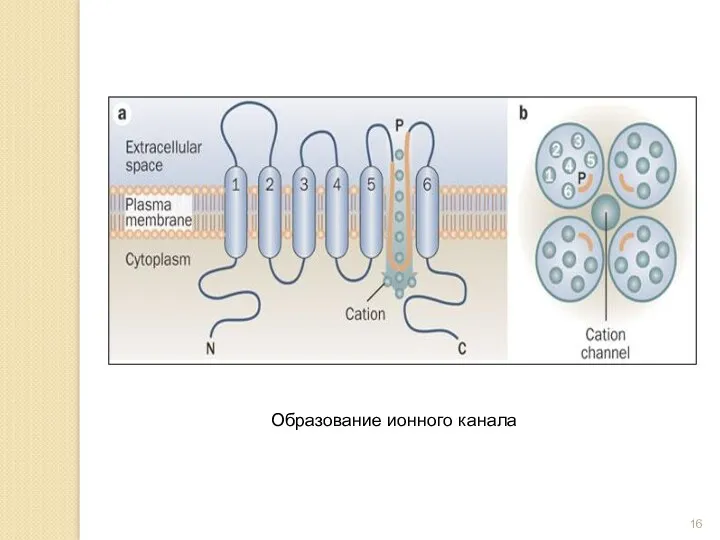
Образование ионного канала
Образование ионного канала

Встраивание ионных каналов в мембрану клетки
Встраивание ионных каналов в мембрану клетки

Анандомин
горчичное масло
эвкалипт
Природные агонисты термочувствительных TRP ионных каналов
Анандомин
горчичное масло
эвкалипт
Природные агонисты термочувствительных TRP ионных каналов

Имеет температурный порог активации 28°С с увеличивающимся током вплоть до 8°С.
Преимущественно
Имеет температурный порог активации 28°С с увеличивающимся током вплоть до 8°С.
Преимущественно
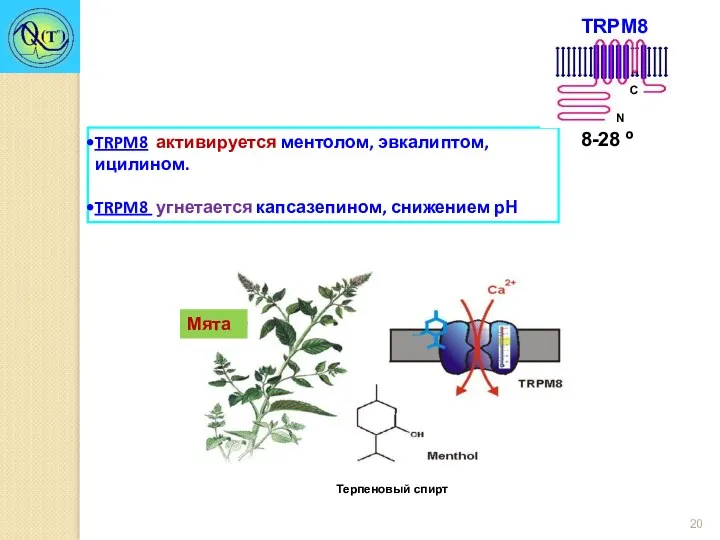
TRPM8 активируется ментолом, эвкалиптом, ицилином.
TRPM8 угнетается капсазепином, снижением рН
Мята
Терпеновый спирт
TRPM8 активируется ментолом, эвкалиптом, ицилином.
TRPM8 угнетается капсазепином, снижением рН
Мята
Терпеновый спирт
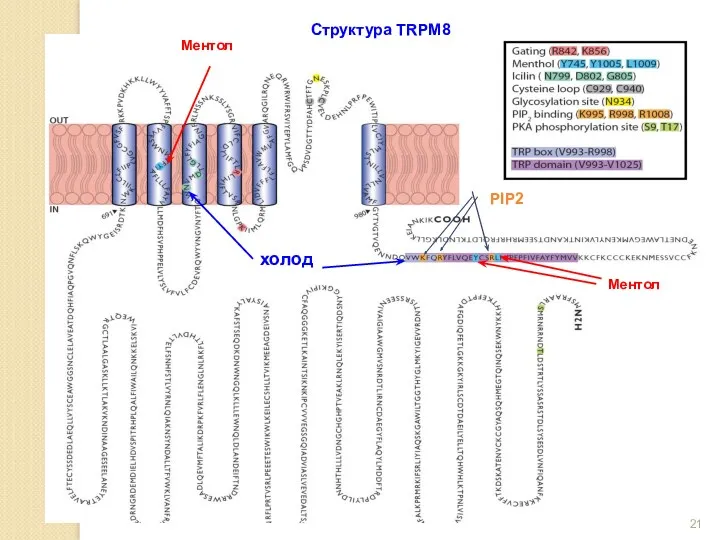
Структура TRPM8
Структура TRPM8
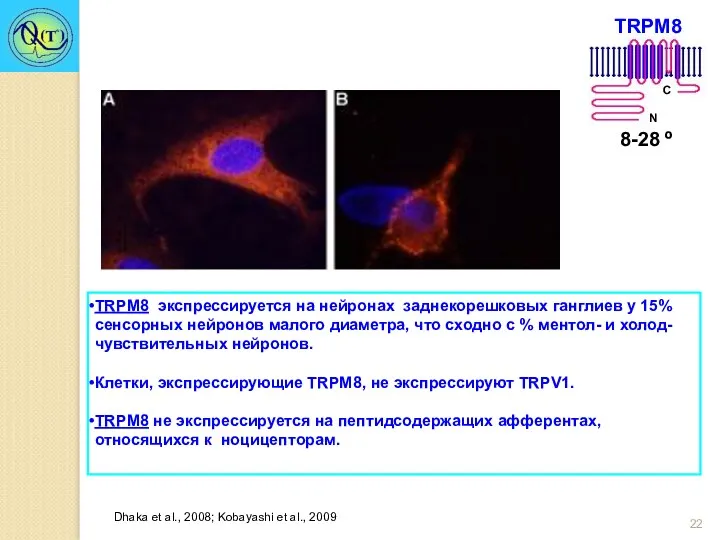
TRPM8 экспрессируется на нейронах заднекорешковых ганглиев у 15% сенсорных нейронов малого
TRPM8 экспрессируется на нейронах заднекорешковых ганглиев у 15% сенсорных нейронов малого

McKemy et al., 2002
Холод и ментол имеют сходный механизм действия, через
McKemy et al., 2002
Холод и ментол имеют сходный механизм действия, через
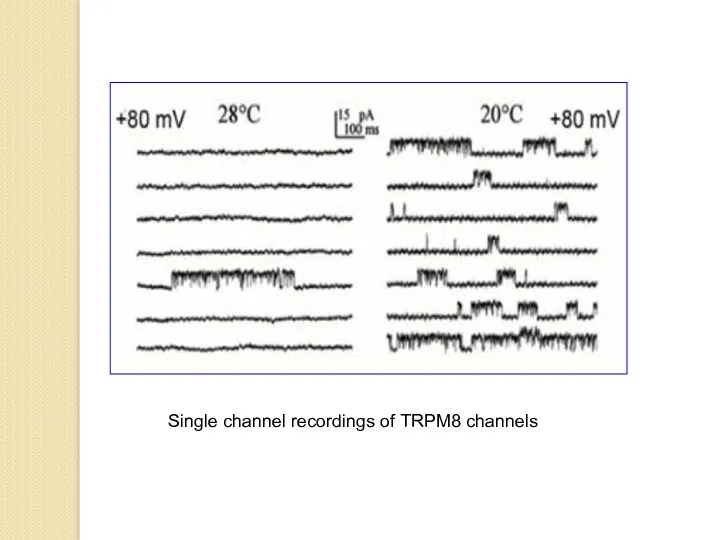
Single channel recordings of TRPM8 channels
Single channel recordings of TRPM8 channels
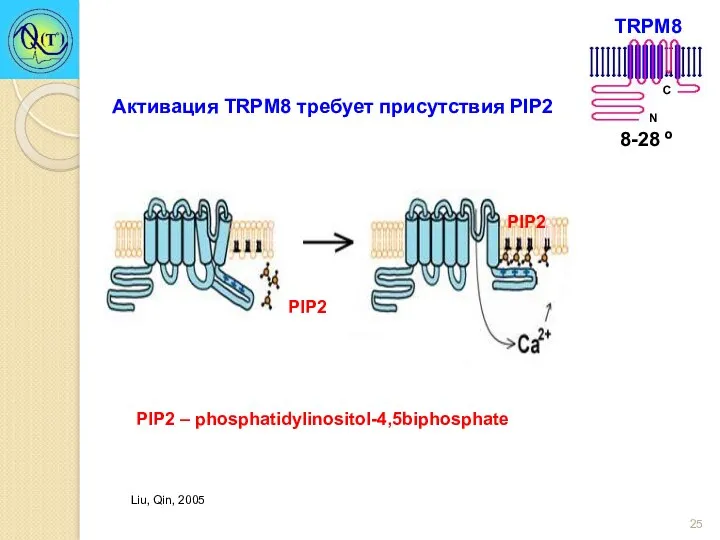
Активация TRPM8 требует присутствия PIP2
PIP2 – phosphatidylinositol-4,5biphosphate
Liu, Qin, 2005
PIP2
PIP2
Активация TRPM8 требует присутствия PIP2
PIP2 – phosphatidylinositol-4,5biphosphate
Liu, Qin, 2005
PIP2
PIP2
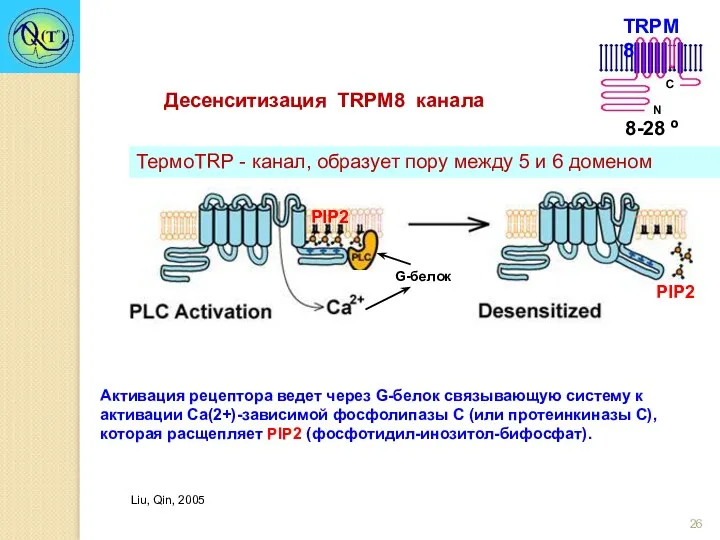
Активация рецептора ведет через G-белок связывающую систему к активации Са(2+)-зависимой фосфолипазы
Активация рецептора ведет через G-белок связывающую систему к активации Са(2+)-зависимой фосфолипазы

TRPА1 активируется холодом при более низкой температуре <17°С. Возможно, является сенсором
TRPА1 активируется холодом при более низкой температуре <17°С. Возможно, является сенсором
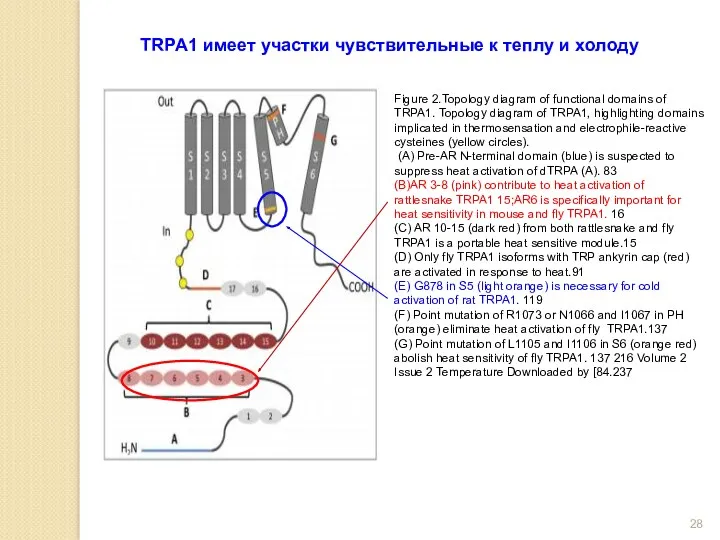
Figure 2.Topology diagram of functional domains of TRPA1. Topology diagram of
Figure 2.Topology diagram of functional domains of TRPA1. Topology diagram of

Cold sensitive
Warm sensitive
TRPA1
Cold sensitive
Warm sensitive
TRPA1
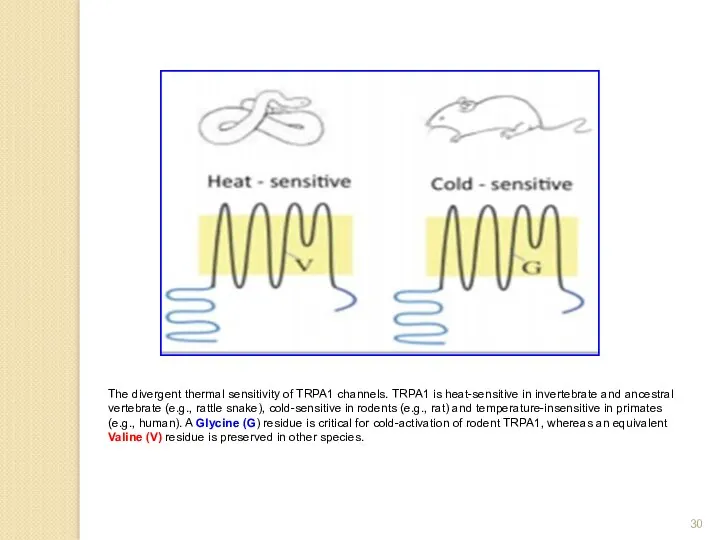
The divergent thermal sensitivity of TRPA1 channels. TRPA1 is heat-sensitive in
The divergent thermal sensitivity of TRPA1 channels. TRPA1 is heat-sensitive in

Активируется температурой >42°С.
Известен, как ваниллоидный рецептор.
Высоко чувствителен к температуре, Q10>20.6.
Активируется температурой >42°С.
Известен, как ваниллоидный рецептор.
Высоко чувствителен к температуре, Q10>20.6.
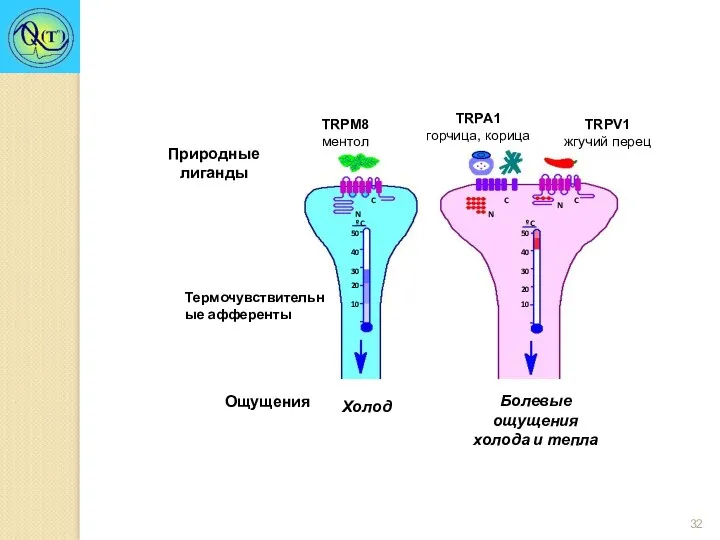
TRPM8
ментол
TRPA1
горчица, корица
TRPV1
жгучий перец
TRPM8
ментол
TRPA1
горчица, корица
TRPV1
жгучий перец

Активируется: капсаицином, низким рН (в противоположность TRPM8).
Сенситизируется также факторами, участвующими в
Активируется: капсаицином, низким рН (в противоположность TRPM8).
Сенситизируется также факторами, участвующими в
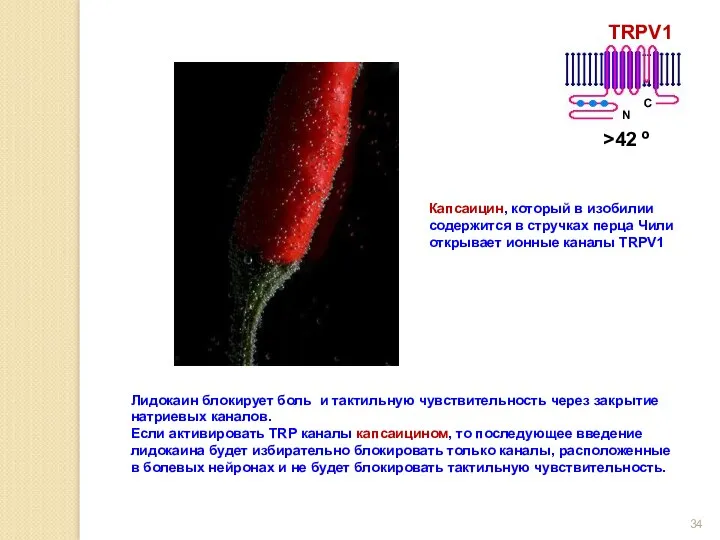
Лидокаин блокирует боль и тактильную чувствительность через закрытие натриевых каналов.
Если
Лидокаин блокирует боль и тактильную чувствительность через закрытие натриевых каналов.
Если
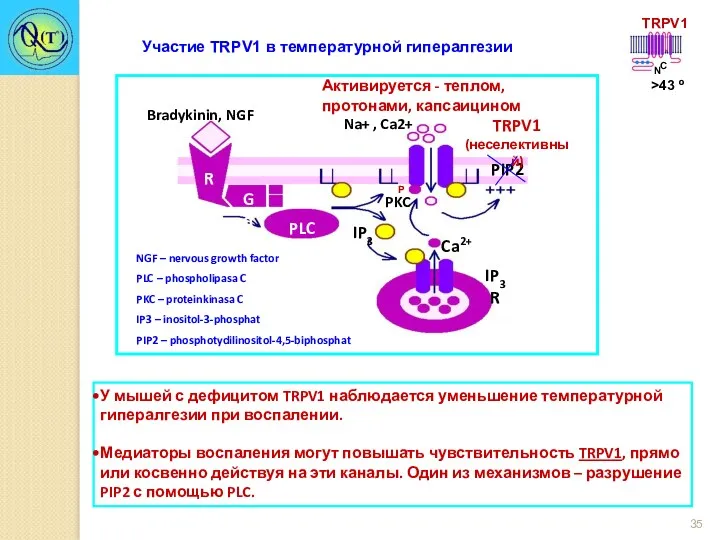
Участие TRPV1 в температурной гипералгезии
У мышей с дефицитом TRPV1 наблюдается уменьшение
Участие TRPV1 в температурной гипералгезии
У мышей с дефицитом TRPV1 наблюдается уменьшение

реагирует на температуру >52°С
не активируется капсаицином и низкой рН
реагирует на температуру >52°С
не активируется капсаицином и низкой рН

Реагирует при температуре 31-39°С.
Кроме температуры активируется камфорой.
Обнаруживает сильную сенситизацию при повторяющихся
Реагирует при температуре 31-39°С.
Кроме температуры активируется камфорой.
Обнаруживает сильную сенситизацию при повторяющихся

Активируется при 27-34°С. В диапазоне до 42°С обогрев вызывает увеличение тока.
Кроме
Активируется при 27-34°С. В диапазоне до 42°С обогрев вызывает увеличение тока.
Кроме

Известно, что существуют 2 типа терморецепторов – холодовые и тепловые.
Детектор температуры
Известно, что существуют 2 типа терморецепторов – холодовые и тепловые.
Детектор температуры

В 2003 году Нобелевская премия по химии была присуждена американскому учёному Родерику Маккиннону
В 2003 году Нобелевская премия по химии была присуждена американскому учёному Родерику Маккиннону

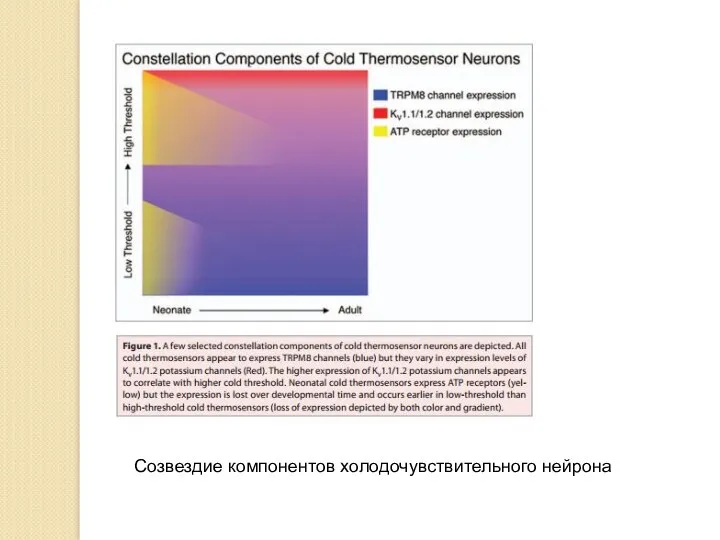
Созвездие компонентов холодочувствительного нейрона
Созвездие компонентов холодочувствительного нейрона

С открытием TRP каналов стало возможным понимание молекулярной основы термочувствительности и
С открытием TRP каналов стало возможным понимание молекулярной основы термочувствительности и

Влияет ли агонист ионного канала TRPM8 ментол на активность и количество
Влияет ли агонист ионного канала TRPM8 ментол на активность и количество
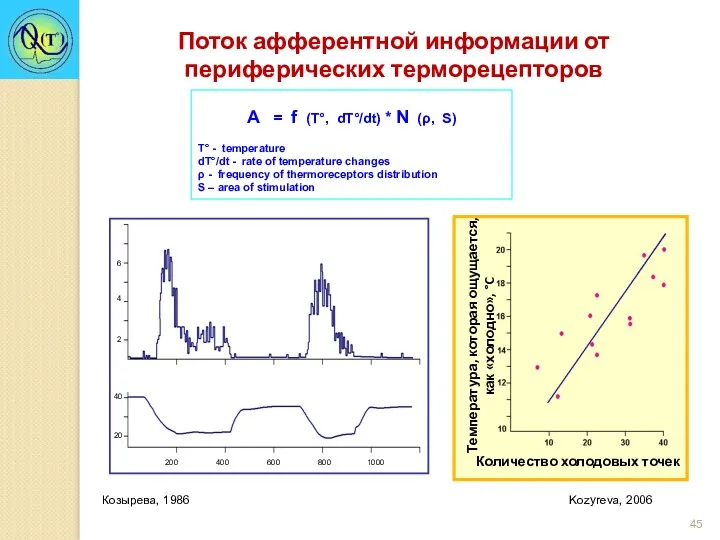
А = f (T°, dT°/dt) * N (ρ, S)
Т° - temperature
dT°/dt
Т° - temperature
dT°/dt

Schafer, Braun, Isenberg, 1986
Влияние ментола на импульсную активность холодового рецептора кожи
Schafer, Braun, Isenberg, 1986
Влияние ментола на импульсную активность холодового рецептора кожи

Влияние 1% ментола на количество холодовых точек у человека
Влияние охлаждения
Влияние 1% ментола на количество холодовых точек у человека
Влияние охлаждения
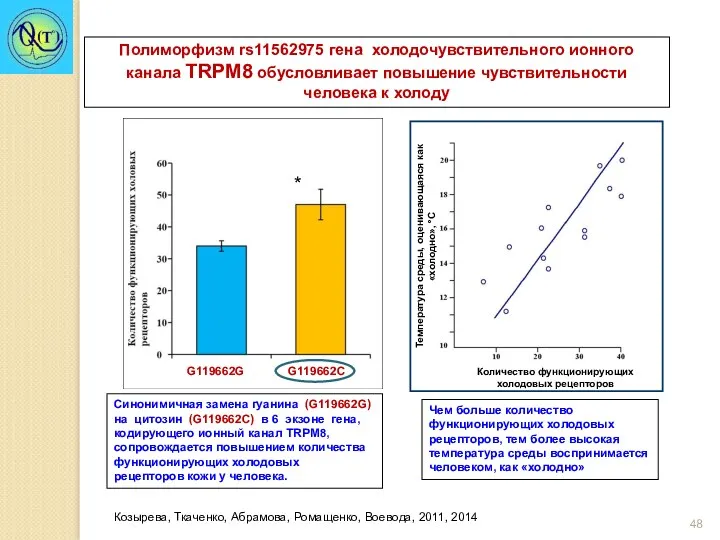
*
Синонимичная замена гуанина (G119662G) на цитозин (G119662С) в 6 экзоне гена,
*
Синонимичная замена гуанина (G119662G) на цитозин (G119662С) в 6 экзоне гена,

Группа G119662C реагирует снижением минутного объема дыхания (МОД) на ментол и
Группа G119662C реагирует снижением минутного объема дыхания (МОД) на ментол и
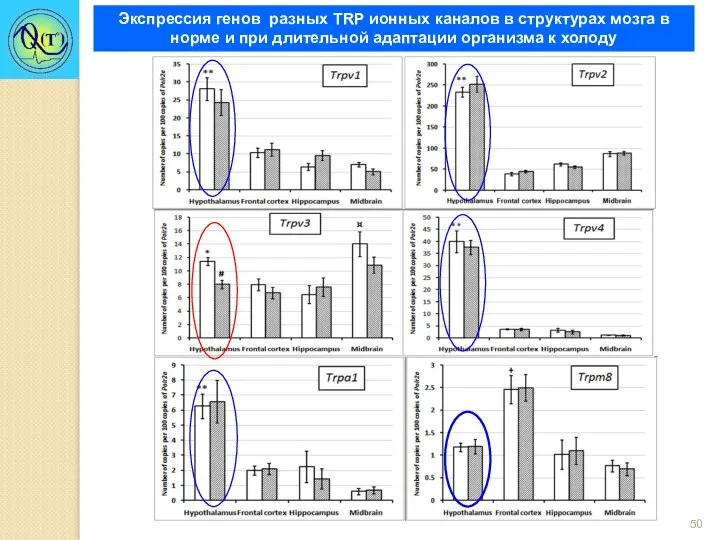
Экспрессия генов разных TRP ионных каналов в структурах мозга в норме
Экспрессия генов разных TRP ионных каналов в структурах мозга в норме

<17°C <25°C 35-39°C >39°C
Адаптация к холоду снижает экспрессию гена ионного
<17°C <25°C 35-39°C >39°C
Адаптация к холоду снижает экспрессию гена ионного

Каково влияние активации холодочувствительного ионного канала TRPM8 в коже на терморегуляторные
Каково влияние активации холодочувствительного ионного канала TRPM8 в коже на терморегуляторные
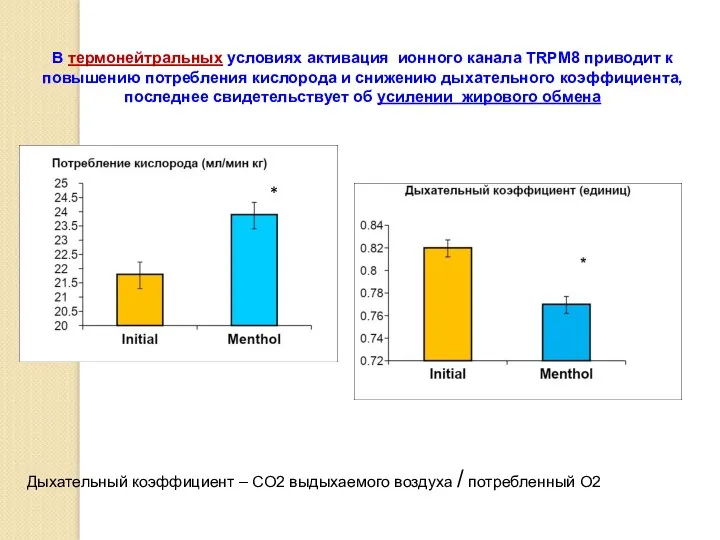
В термонейтральных условиях активация ионного канала TRPM8 приводит к повышению потребления
В термонейтральных условиях активация ионного канала TRPM8 приводит к повышению потребления
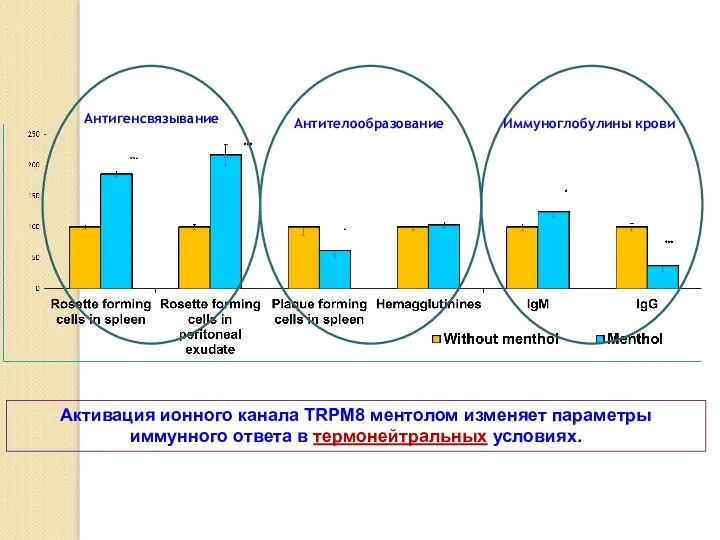
Активация ионного канала TRPM8 ментолом изменяет параметры иммунного ответа в термонейтральных
Активация ионного канала TRPM8 ментолом изменяет параметры иммунного ответа в термонейтральных
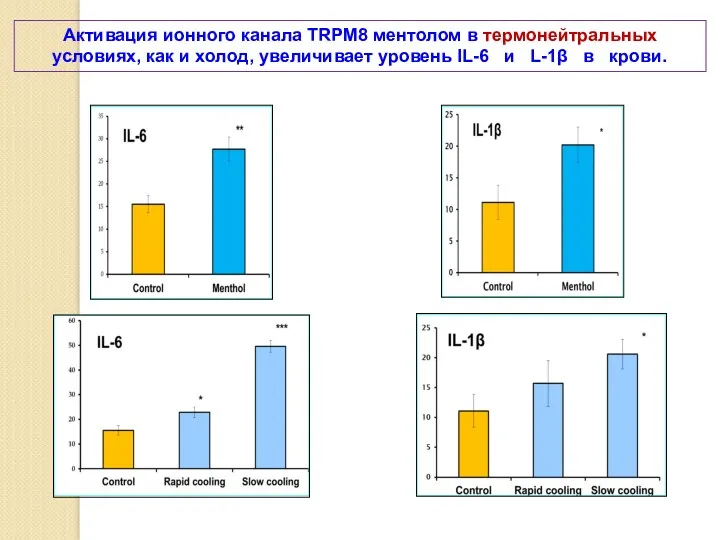
Активация ионного канала TRPM8 ментолом в термонейтральных условиях, как и холод,
Активация ионного канала TRPM8 ментолом в термонейтральных условиях, как и холод,
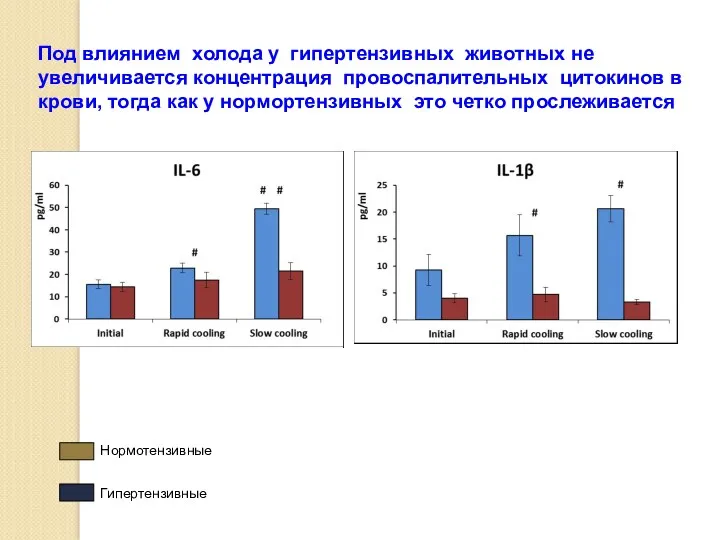
Под влиянием холода у гипертензивных животных не увеличивается концентрация провоспалительных цитокинов
Под влиянием холода у гипертензивных животных не увеличивается концентрация провоспалительных цитокинов
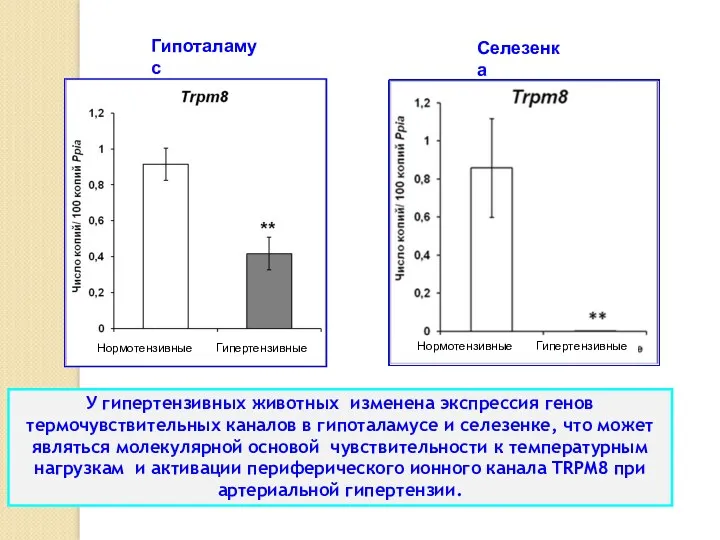
У гипертензивных животных изменена экспрессия генов термочувствительных каналов в гипоталамусе и
У гипертензивных животных изменена экспрессия генов термочувствительных каналов в гипоталамусе и
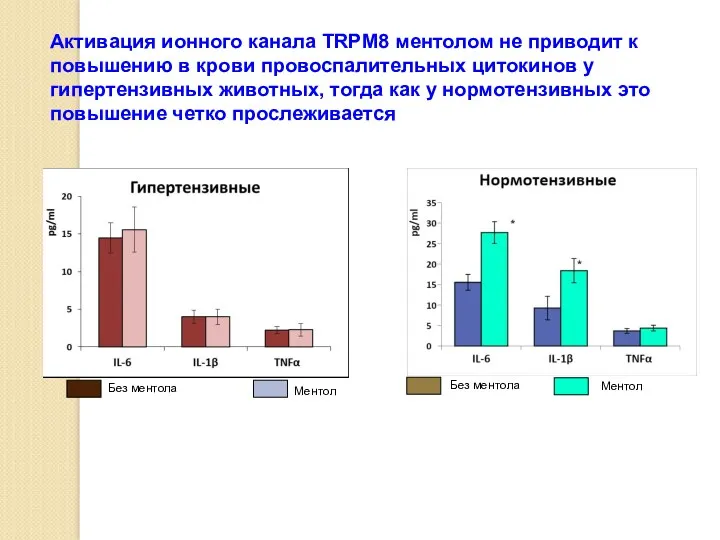
Активация ионного канала TRPM8 ментолом не приводит к повышению в крови
Активация ионного канала TRPM8 ментолом не приводит к повышению в крови

Как активация холодо-чувствительного ионного канала TRPM8 влияет на терморегуляторные и иммунные
Как активация холодо-чувствительного ионного канала TRPM8 влияет на терморегуляторные и иммунные

Активация ионного канала TRPM8 приводит к снижению температурных порогов терморегуляторных реакций
Активация ионного канала TRPM8 приводит к снижению температурных порогов терморегуляторных реакций

Активация ионного канала TRPM8 ментолом ослабляет угнетающий эффект глубокого охлаждения на
Активация ионного канала TRPM8 ментолом ослабляет угнетающий эффект глубокого охлаждения на
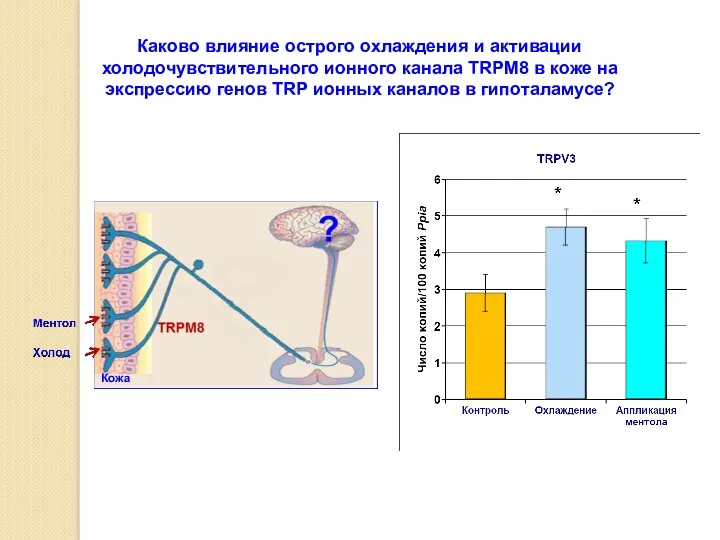
Каково влияние острого охлаждения и активации холодочувствительного ионного канала TRPM8 в
Каково влияние острого охлаждения и активации холодочувствительного ионного канала TRPM8 в

Итак, активация холодочувствительного ионного каналаTRPМ8 оказывает существенное влияние на регуляцию жирового
Итак, активация холодочувствительного ионного каналаTRPМ8 оказывает существенное влияние на регуляцию жирового

N=-25.9+26.2PCa
r = 0.62
Козырева ,Тихонова, Ткаченко, Синдаровская, 1987
Влияние ионофореза Ca(2+) в кожу
N=-25.9+26.2PCa
r = 0.62
Козырева ,Тихонова, Ткаченко, Синдаровская, 1987
Влияние ионофореза Ca(2+) в кожу

Длительная адаптация организма к холоду приводит к снижению концентрации Са(2+) в
Длительная адаптация организма к холоду приводит к снижению концентрации Са(2+) в
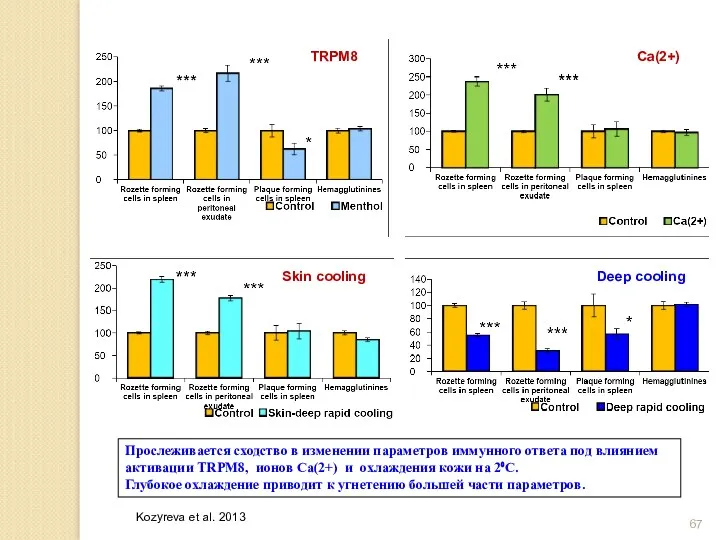
Прослеживается сходство в изменении параметров иммунного ответа под влиянием активации TRPM8,
Прослеживается сходство в изменении параметров иммунного ответа под влиянием активации TRPM8,
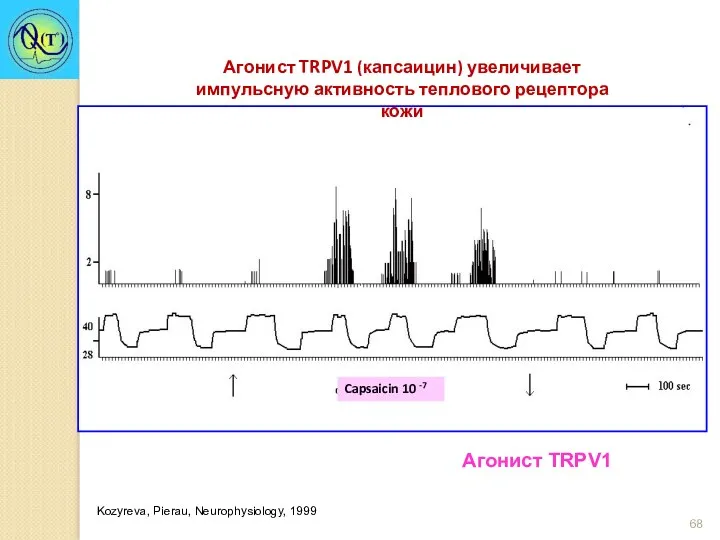
Агонист TRPV1 (капсаицин) увеличивает импульсную активность теплового рецептора кожи
Capsaicin 10 -7
Kozyreva,
Агонист TRPV1 (капсаицин) увеличивает импульсную активность теплового рецептора кожи
Capsaicin 10 -7
Kozyreva,
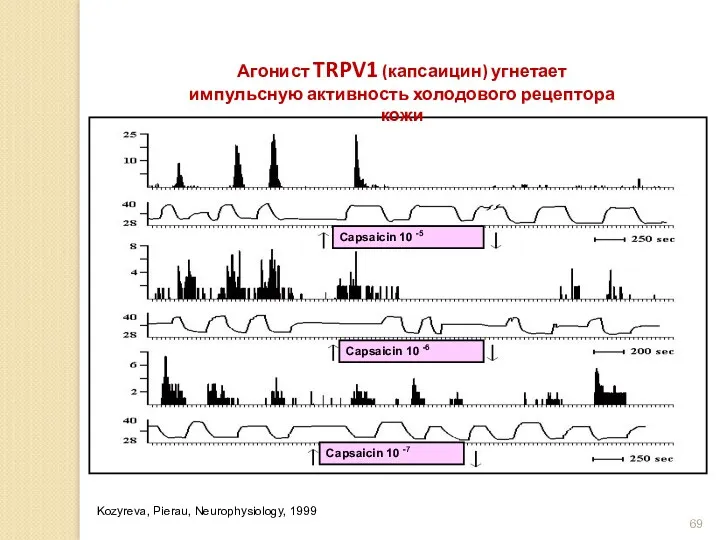
Агонист TRPV1 (капсаицин) угнетает импульсную активность холодового рецептора кожи
Kozyreva, Pierau, Neurophysiology,
Агонист TRPV1 (капсаицин) угнетает импульсную активность холодового рецептора кожи
Kozyreva, Pierau, Neurophysiology,
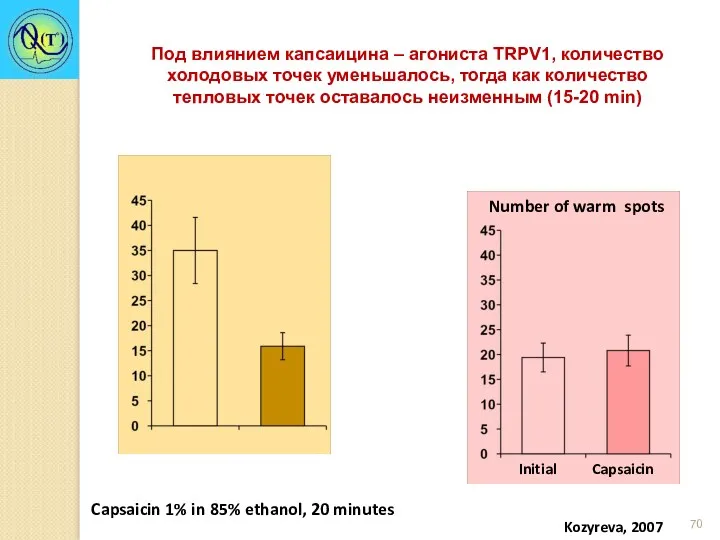
Под влиянием капсаицина – агониста TRPV1, количество холодовых точек уменьшалось, тогда
Под влиянием капсаицина – агониста TRPV1, количество холодовых точек уменьшалось, тогда
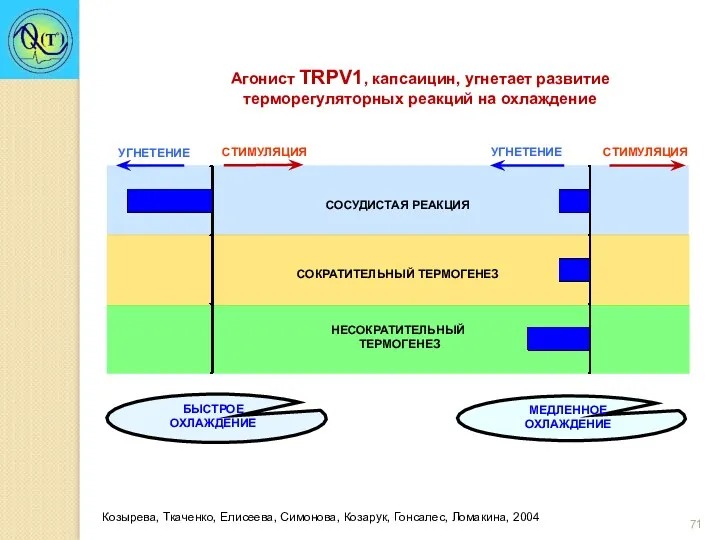
Козырева, Ткаченко, Елисеева, Симонова, Козарук, Гонсалес, Ломакина, 2004
Козырева, Ткаченко, Елисеева, Симонова, Козарук, Гонсалес, Ломакина, 2004
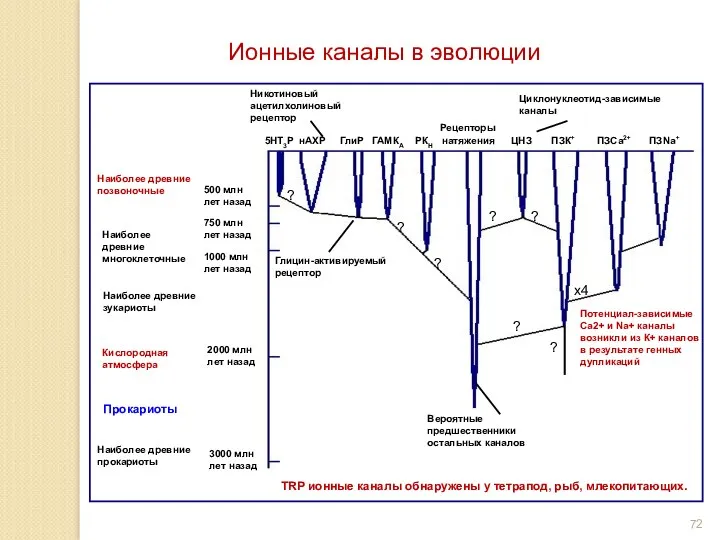
Ионные каналы в эволюции
TRP ионные каналы обнаружены у тетрапод, рыб, млекопитающих.
Ионные каналы в эволюции
TRP ионные каналы обнаружены у тетрапод, рыб, млекопитающих.


Температура модулирует работу многих ионных каналов.
ТермоTRP- каналы активируются непосредственно температурой
Температура модулирует работу многих ионных каналов.
ТермоTRP- каналы активируются непосредственно температурой
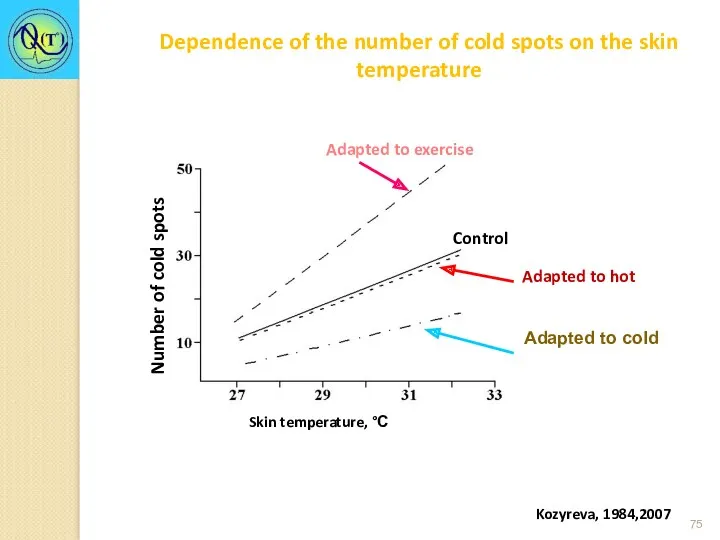
Kozyreva, 1984,2007
Dependence of the number of cold spots on the
Kozyreva, 1984,2007
Dependence of the number of cold spots on the


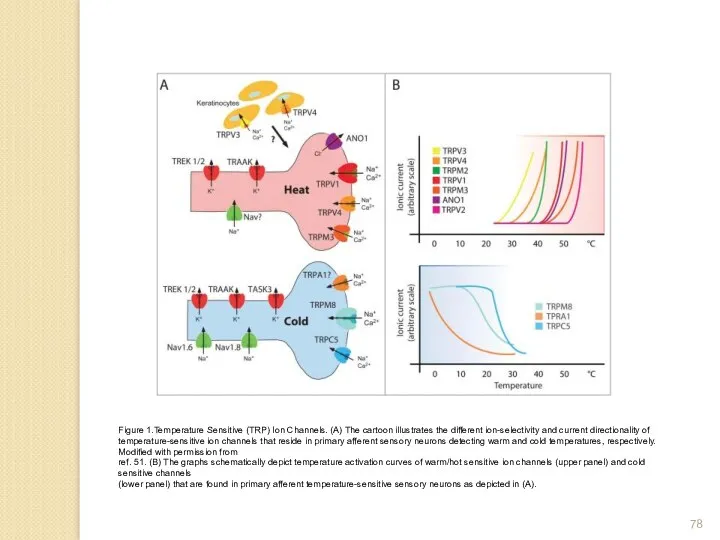
Figure 1.Temperature Sensitive (TRP) Ion Channels. (A) The cartoon illustrates the
Figure 1.Temperature Sensitive (TRP) Ion Channels. (A) The cartoon illustrates the
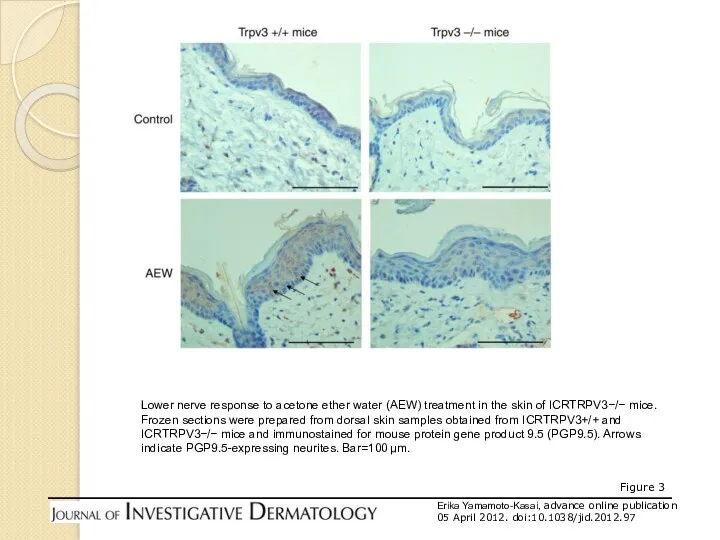
Figure 3
Erika Yamamoto-Kasai, advance online publication
05 April 2012. doi:10.1038/jid.2012.97
Lower nerve
Figure 3
Erika Yamamoto-Kasai, advance online publication
05 April 2012. doi:10.1038/jid.2012.97
Lower nerve
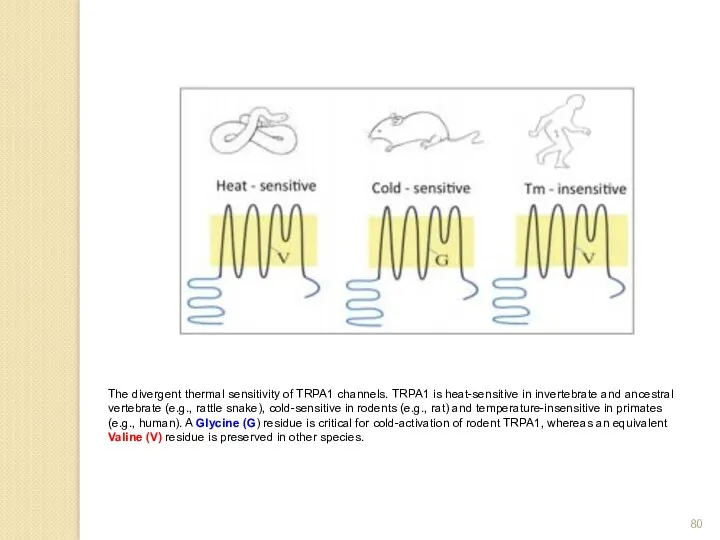
The divergent thermal sensitivity of TRPA1 channels. TRPA1 is heat-sensitive in
The divergent thermal sensitivity of TRPA1 channels. TRPA1 is heat-sensitive in
 Нормальная рентгенанатомия опорно-двигательной системы
Нормальная рентгенанатомия опорно-двигательной системы Роль пациентской организации в повышении качества жизни больных с хроническими заболеваниями почек
Роль пациентской организации в повышении качества жизни больных с хроническими заболеваниями почек Введение в вакцинопрофилактику
Введение в вакцинопрофилактику Сердечно–сосудистая система человека
Сердечно–сосудистая система человека СПИД и его профилактика
СПИД и его профилактика Реабилитация. Реабилитацияның түрлері
Реабилитация. Реабилитацияның түрлері Эмоциональный стресс: его вегетативные и моторные компоненты
Эмоциональный стресс: его вегетативные и моторные компоненты Иммунитет
Иммунитет Эпидемиология. Причинно-следственные связи
Эпидемиология. Причинно-следственные связи Захворювання щитовидної залози у дітей
Захворювання щитовидної залози у дітей Микробиология холеры
Микробиология холеры Чума. Морфологические свойства
Чума. Морфологические свойства Врачебная помощь при воздействии электрического тока на человека. (Лекция 6)
Врачебная помощь при воздействии электрического тока на человека. (Лекция 6) Медико-генетикалық кеңестің нәрестенің туа біткен ақаулықтарын алдын-алудағы рөлі және диагностикасы
Медико-генетикалық кеңестің нәрестенің туа біткен ақаулықтарын алдын-алудағы рөлі және диагностикасы Рентгенография легких
Рентгенография легких Физиологическая и патологическая реактивность
Физиологическая и патологическая реактивность Анатомо-физиологические особенности периодонта у детей различного возраста
Анатомо-физиологические особенности периодонта у детей различного возраста Клініко-діагностичний центр Фармбіотест. Науково-дослідний медичний центр європейського типу. Рубіжне, Україна
Клініко-діагностичний центр Фармбіотест. Науково-дослідний медичний центр європейського типу. Рубіжне, Україна Құтыру
Құтыру Аллергический ринит и бронхиальная астма - актуальная проблема
Аллергический ринит и бронхиальная астма - актуальная проблема Руки персонала как основа инфекционной безопасности
Руки персонала как основа инфекционной безопасности Ultrasound is
Ultrasound is Ісік жасушаларын зерттеу
Ісік жасушаларын зерттеу Опорно-двигательный аппарат
Опорно-двигательный аппарат Легочная гипертензия
Легочная гипертензия Creators and discovery of vaccines against COVID‐19
Creators and discovery of vaccines against COVID‐19 Анатомия системы головного мозга
Анатомия системы головного мозга Инфекционная безопасность
Инфекционная безопасность