- Защитные функции антител

Содержание
- 2. Внешняя среда Макрофаг Дендритная клетка Фибробласт Лимфатический узел (миндалина, Пейерова бляшка) Бактерии
- 3. В-клетка, у которой сработали рецепторы, секретирует огромное количество молекул, подобных рецепторам Black box
- 4. При попадании в организм чужеродных веществ, например, компонент микроба, происходит селективное размножение лимфоцитов, рецепторы которых способны
- 5. В-лимфоциты, «схватившие» антиген , не только размножаются (~ 1 деление в сутки), но и сильно изменяются
- 6. В клетка Плазматическая клетка
- 7. Антиген Антитело В клетка
- 8. Антитела находят соответствующие антигены и связываются с ними. Растворимые антигены переводятся в нерастворимые комплексы [антиген +
- 9. Антиген Антитело Иммунный комплекс
- 10. Наличие двух «хваталок» на молекуле антитела и хотя бы двух эпитопов на молекуле антигена являются необходимым
- 11. Антитело 1 Антитело 2 Эпитоп 1 Эпитоп 2
- 12. Мультимолекулярный комплекс [Аг + Ат] Условия образования мультимолекулярных комплексов: Антитело должно иметь два Fab-фрагмента, чтобы хвататься
- 13. Антиген Антитело АГ АТ Комплекс [Антиген+Антитело]
- 18. Как и от чего можно защититься с помощью антител? Римский додекаэдр – не известно, зачем он
- 19. ЗАЩИТНЫЕ ФУНКЦИИ АНТИТЕЛ Нейтрализация токсина Удаление токсина в виде иммунных комплексов Нейтрализация вируса Препятствие адгезии бактерий
- 20. Клетка Клетка Антитело Токсин Рецептор токсина
- 21. Зачастую вред наносит не бактерия, а ее токсины. В таких случаях, если обезвредить токсины, то присутствие
- 23. Токсичность ядов бактерий Яд змеи тайпан (самая ядовитая змея) LD50 = 10 мкг/кг, 1 грамма достаточно
- 24. Нейтрализация опасного зверя
- 25. Антитела препятствуют адгезии бактерий
- 26. Антитела нейтрализуют вирусы
- 27. Антитела опсонизируют поверхность микробов
- 28. Fc-фрагмент Ig Fc-рецептор Микроб Фагоцит
- 29. Fc-рецепторы фагоцитов облегчают фагоцитоз частиц, покрытых антителами
- 31. Fc-рецептор 108 молекулярных инструментов, чтобы найти «чужое», и всего 1 молекулярный инструмент, чтобы удалить эти 108
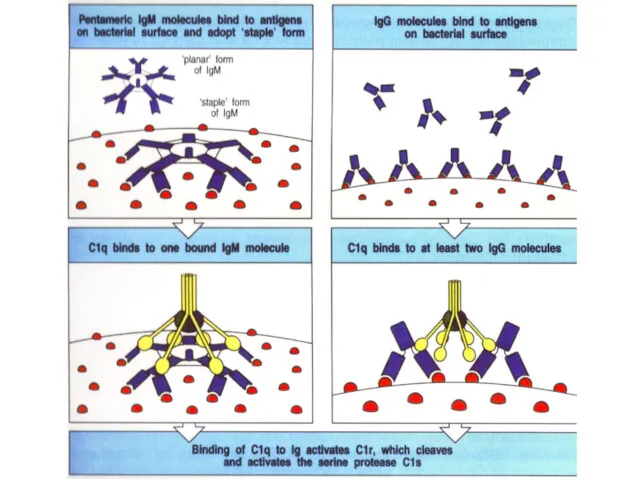
- 32. Комплексы [антиген + антитело] активируют комплемент
- 33. Антитело класса IgM (пентамер, вырабатывается в первые дни) Антитело класса IgG (мономер, вырабатывается, начиная со 2-й
- 35. Маннан-связывающий лектин Концевая манноза Alternative pathway Lectin pathway C4
- 36. Маннан-связывающий лектин Концевая манноза C1q Антиген Антитело
- 40. Рентгеновская структура C1q
- 41. Сайт, с которым связывается C1q CH3-domain CH2-domain CH1-domain VH-domain
- 42. Посадку C1q на две молекулы Ig можно реконструировать с учетом рентгеновской структуры молекул (см. следующие два
- 43. Модель взаимодействия C1q c Fc-фрагментом антитела (IgG) C1q Ig
- 44. Ig Ig Поверхность C1q
- 45. Figure 6 Structural Basis of Complement Activation in the Lectin Pathway (A) Superimposition of the structures
- 46. C9 C1q C1r, C1s C2 ? C2a +C2b C4 ? C4a + C4b C2aC4bC3b (конвертаза С5)
- 47. С1q Mannan-binding protein (lectin) Alternative pathway Lectin pathway Classical pathway
- 48. С3 С1q С8 С2 С1r С6 С4 С1s С3b С2b С4b С5 С5a С5b С9 С7
- 50. ЗАЩИТНЫЕ ФУНКЦИИ АНТИТЕЛ Нейтрализация токсина Удаление токсина в виде иммунных комплексов Нейтрализация вируса Препятствие адгезии бактерий
- 51. Антитела и комплемент синергичны при опсонизации микроорганизмов
- 53. Антитела и комплемент повреждают даже многоклеточных паразитов
- 55. Антитела в роли «черной» метки, по которой клетки-киллеры атакуют паразитов
- 56. Паразитические черви, «меченые» антителами IgG и IgE, подвергаются атаке эозинофильными гранулоцитами
- 57. ЭОЗИНОФИЛ
- 58. Eosinophil granule major basic protein //en.wikipedia.org/wiki/Major_basic_protein Образует кристаллическое ядро эозинофильной гранулы, участвует в защите от паразитов
- 62. Антитела защищают нас не только внутри, но и снаружи, на границе с внешним миром
- 63. Многослойный реснитчатый эпителий Схема Эпителий носовых ходов Гистологический препарат x270 Схема
- 64. Трахея Эпителий дыхательных путей Трахея Бронх Терминальный бронх
- 65. ILEUM villus L - lumen V – villus CL – crypt of Lieberkuhn GC - goblet
- 66. Эпителий Фибробласты Бактерии Вирусы Грибы Простейшие Плазматические клетки Антитело
- 67. Антитела транспортируются на поверхности, служащие границей между нашим организмом и внешней средой.
- 68. Через эпителии секретируются антитела изотипа IgA
- 69. Poly-Ig receptor переносит IgA или IgM внутри эпителиальной клетки на ее внешнюю (апикальную) мембрану. Здесь ферменты
- 70. Особенно эффективно poly-Ig-рецептор переносит антитела через эпителий молочной железы и печени. В случае молочной железы это
- 71. Что они делают? Они очищаются от паразитов
- 72. Антитела сообщают собаке (лошади): О том, что на теле поселился паразит Точно указывают место, где этот
- 73. Антитела изотипа IgE синтезируются в очень малых количествах в ответ на вторжение малых количеств чужих антигенов,
- 74. Клетка, секретирующая антитела класса IgE IgE Ка ~1010 [Fc-рецептор + IgE] Паразит Антигены паразита Тучная клетка,
- 75. Клетка, секретирующая антитела класса IgE IgE Ка ~1010 [Fc-рецептор + IgE] Паразит Антигены паразита Тучная клетка,
- 76. Антитела IgE служат рецепторами антигенов в мембране тучных клеток. Это позволяет тучным клеткам запускать следующие защитные
- 77. Изотипы антител имеют разные защитные функции Мономеры IgD и IgM служат рецепторами на поверхности B-клеток Пентамеры
- 78. Полезные сведения о изотипах антител
- 80. Скачать презентацию
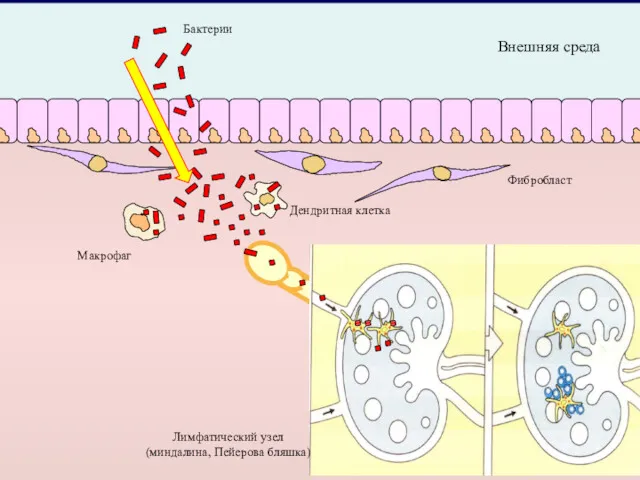
Внешняя среда
Макрофаг
Дендритная клетка
Фибробласт
Лимфатический узел
(миндалина, Пейерова бляшка)
Бактерии
Внешняя среда
Макрофаг
Дендритная клетка
Фибробласт
Лимфатический узел
(миндалина, Пейерова бляшка)
Бактерии
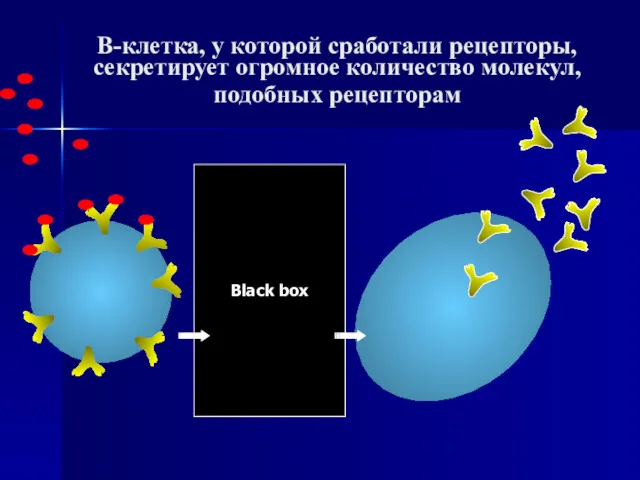
В-клетка, у которой сработали рецепторы, секретирует огромное количество молекул, подобных рецепторам
В-клетка, у которой сработали рецепторы, секретирует огромное количество молекул, подобных рецепторам
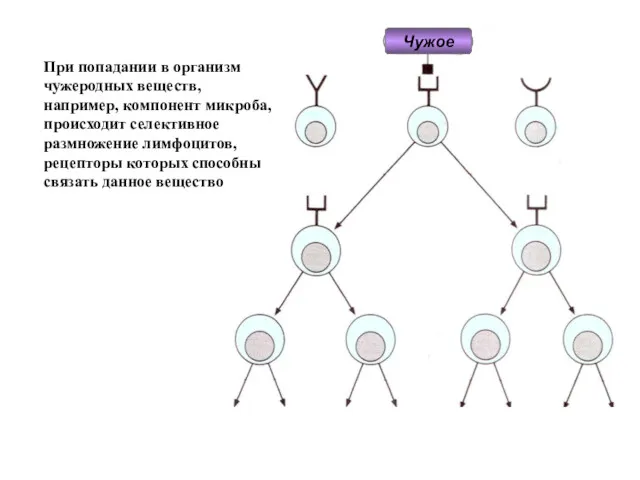
При попадании в организм
чужеродных веществ,
например, компонент микроба,
происходит селективное размножение
При попадании в организм
чужеродных веществ,
например, компонент микроба,
происходит селективное размножение

В-лимфоциты, «схватившие» антиген ,
не только размножаются (~ 1 деление в
В-лимфоциты, «схватившие» антиген ,
не только размножаются (~ 1 деление в
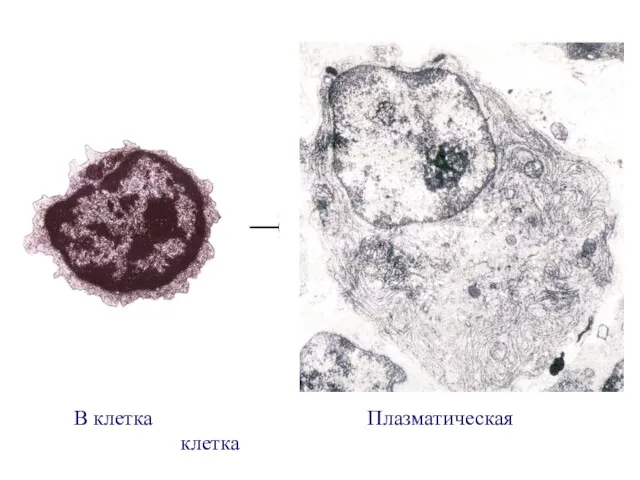
В клетка Плазматическая клетка
В клетка Плазматическая клетка
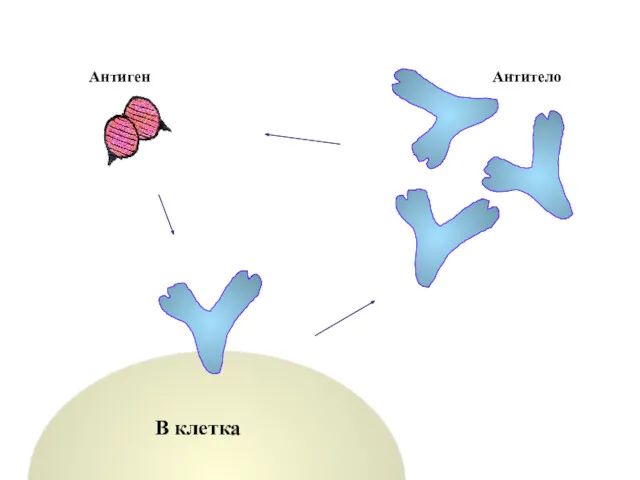
Антиген
Антитело
В клетка
Антиген
Антитело
В клетка

Антитела находят соответствующие антигены
и связываются с ними.
Растворимые антигены переводятся в
Антитела находят соответствующие антигены
и связываются с ними.
Растворимые антигены переводятся в
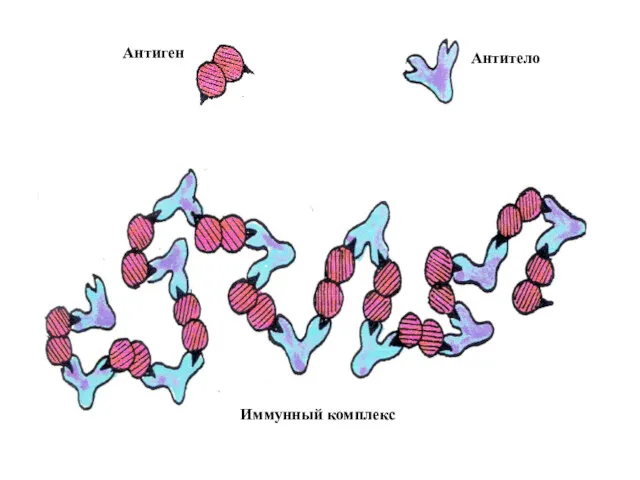
Антиген
Антитело
Иммунный комплекс
Антиген
Антитело
Иммунный комплекс
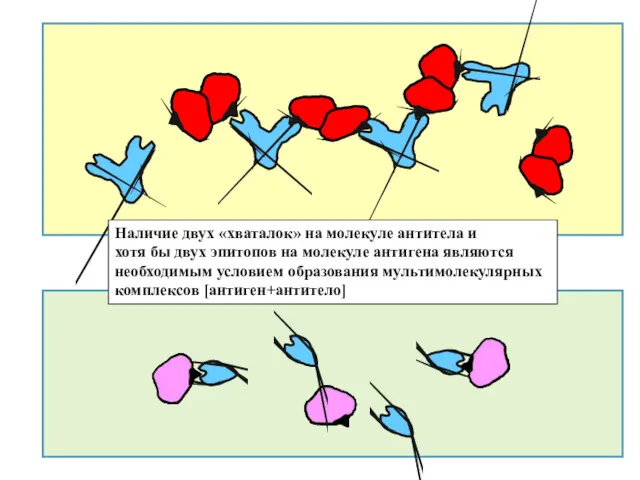
Наличие двух «хваталок» на молекуле антитела и
хотя бы двух эпитопов
Наличие двух «хваталок» на молекуле антитела и
хотя бы двух эпитопов
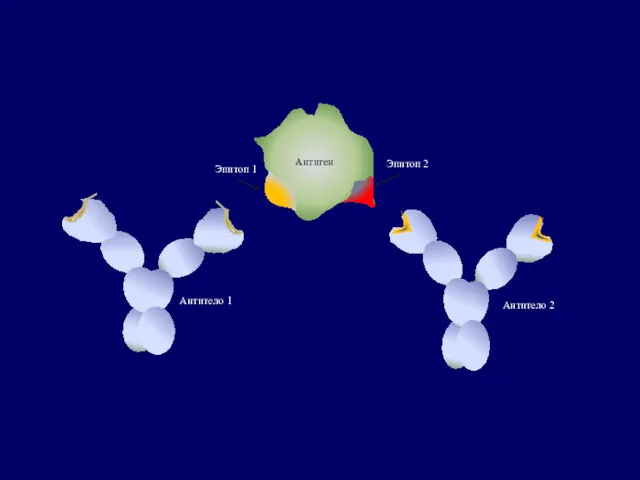
Антитело 1
Антитело 2
Эпитоп 1
Эпитоп 2
Антитело 1
Антитело 2
Эпитоп 1
Эпитоп 2
![Мультимолекулярный комплекс [Аг + Ат] Условия образования мультимолекулярных комплексов: Антитело](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/125202/slide-11.jpg)
Мультимолекулярный комплекс [Аг + Ат]
Условия образования мультимолекулярных комплексов:
Антитело должно иметь два
Мультимолекулярный комплекс [Аг + Ат]
Условия образования мультимолекулярных комплексов:
Антитело должно иметь два
![Антиген Антитело АГ АТ Комплекс [Антиген+Антитело]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/125202/slide-12.jpg)
Антиген
Антитело
АГ
АТ
Комплекс [Антиген+Антитело]
Антиген
Антитело
АГ
АТ
Комплекс [Антиген+Антитело]
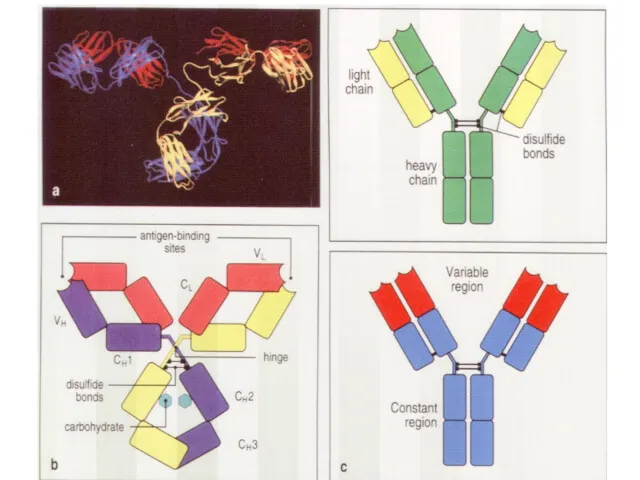
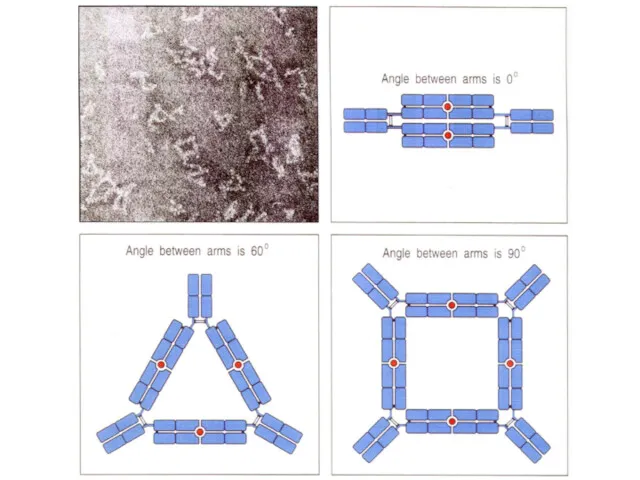

Как и от чего можно защититься с помощью антител?
Римский додекаэдр –
Как и от чего можно защититься с помощью антител?
Римский додекаэдр –

ЗАЩИТНЫЕ ФУНКЦИИ АНТИТЕЛ
Нейтрализация токсина
Удаление токсина в виде иммунных комплексов
Нейтрализация вируса
Препятствие адгезии
ЗАЩИТНЫЕ ФУНКЦИИ АНТИТЕЛ
Нейтрализация токсина
Удаление токсина в виде иммунных комплексов
Нейтрализация вируса
Препятствие адгезии
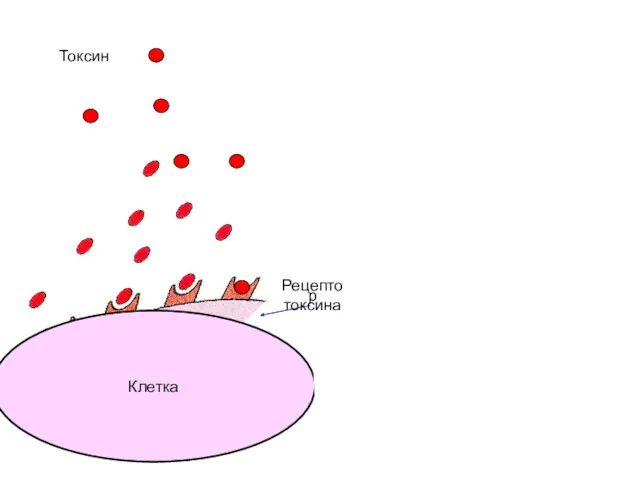
Клетка
Клетка
Антитело
Токсин
Рецептор
токсина
Клетка
Клетка
Антитело
Токсин
Рецептор
токсина

Зачастую вред наносит не бактерия, а ее токсины.
В таких случаях, если
Зачастую вред наносит не бактерия, а ее токсины.
В таких случаях, если
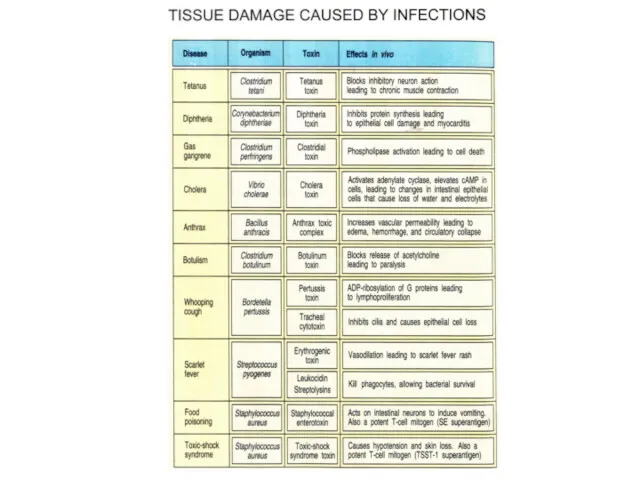
Токсичность ядов бактерий
Яд змеи тайпан (самая ядовитая змея) LD50 = 10
Токсичность ядов бактерий
Яд змеи тайпан (самая ядовитая змея) LD50 = 10
Нейтрализация опасного зверя
Нейтрализация опасного зверя
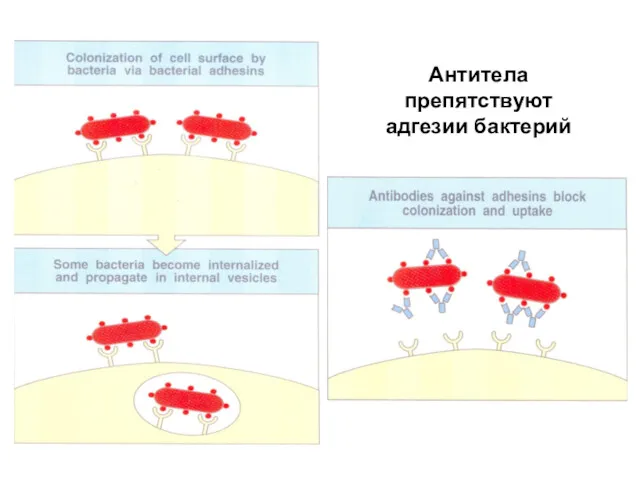
Антитела препятствуют
адгезии бактерий
Антитела препятствуют
адгезии бактерий
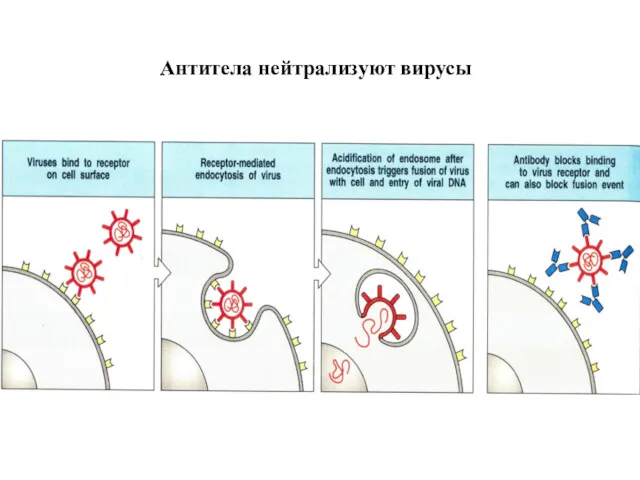
Антитела нейтрализуют вирусы
Антитела нейтрализуют вирусы

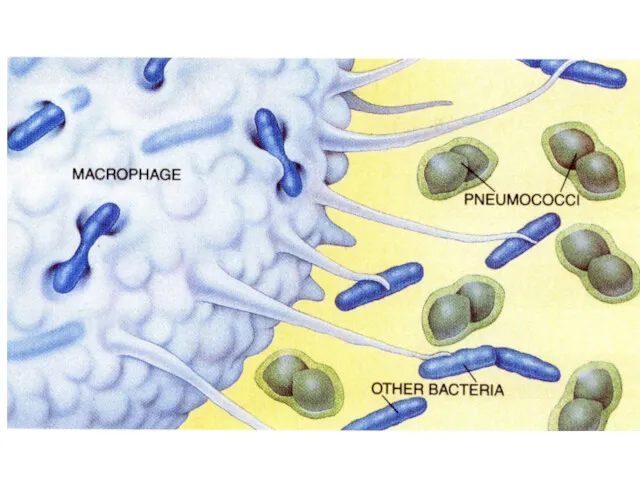
Антитела опсонизируют поверхность микробов
Антитела опсонизируют поверхность микробов
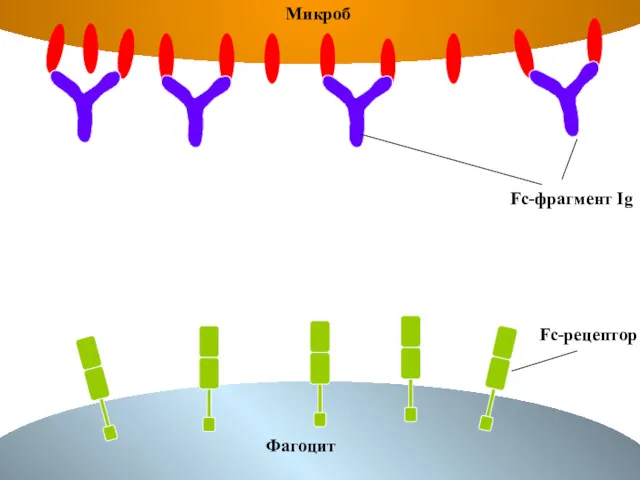
Fc-фрагмент Ig
Fc-рецептор
Микроб
Фагоцит
Fc-фрагмент Ig
Fc-рецептор
Микроб
Фагоцит
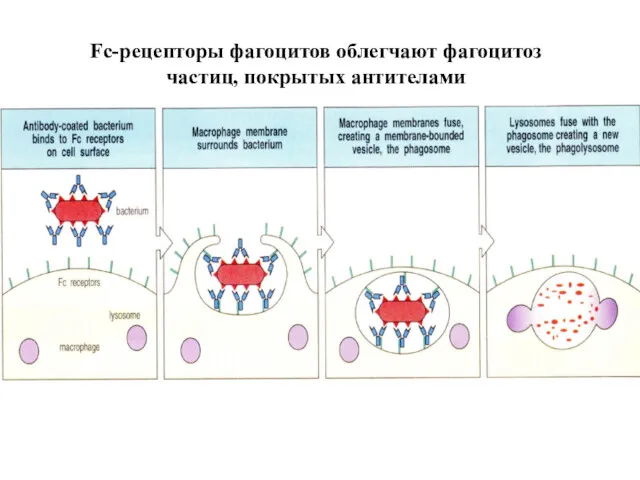
Fc-рецепторы фагоцитов облегчают фагоцитоз
частиц, покрытых антителами
Fc-рецепторы фагоцитов облегчают фагоцитоз
частиц, покрытых антителами
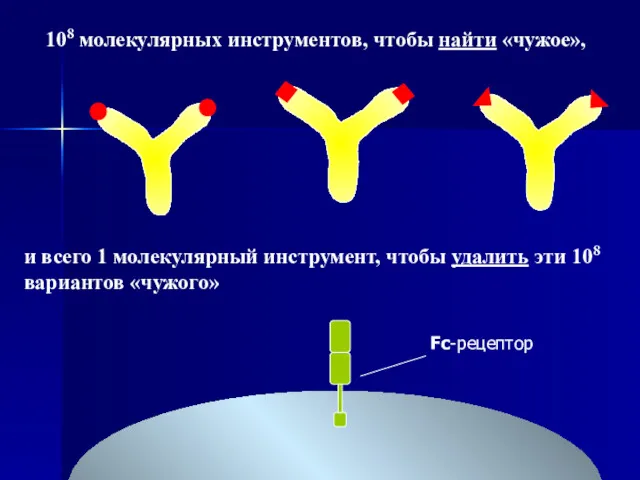
Fc-рецептор
108 молекулярных инструментов, чтобы найти «чужое»,
и всего 1 молекулярный инструмент,
Fc-рецептор
108 молекулярных инструментов, чтобы найти «чужое»,
и всего 1 молекулярный инструмент,
![Комплексы [антиген + антитело] активируют комплемент](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/125202/slide-31.jpg)
Комплексы [антиген + антитело] активируют комплемент
Комплексы [антиген + антитело] активируют комплемент
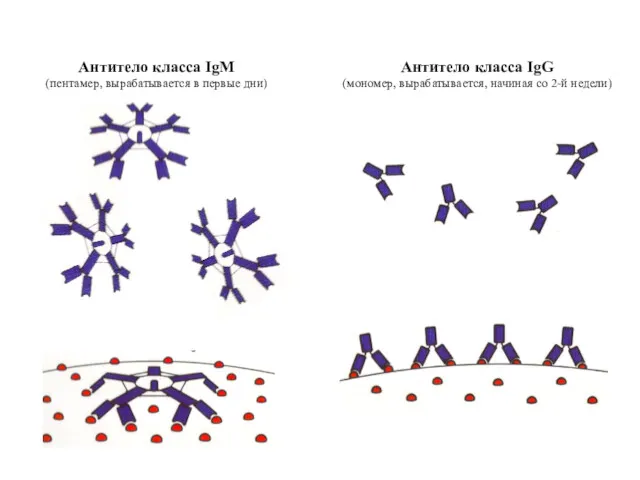
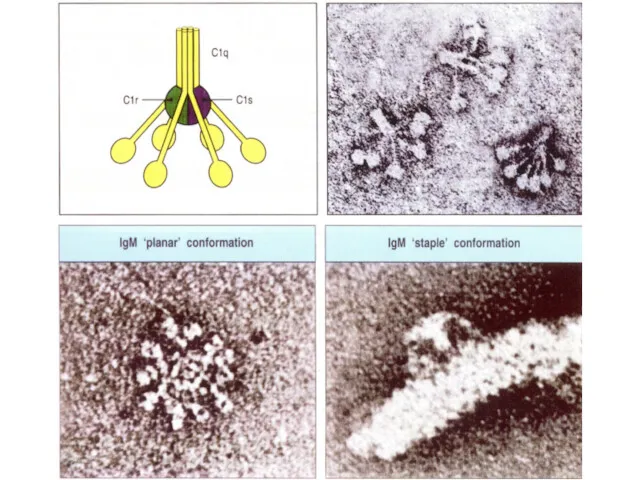
Антитело класса IgM
(пентамер, вырабатывается в первые дни)
Антитело класса IgG
(мономер, вырабатывается, начиная
Антитело класса IgM
(пентамер, вырабатывается в первые дни)
Антитело класса IgG
(мономер, вырабатывается, начиная
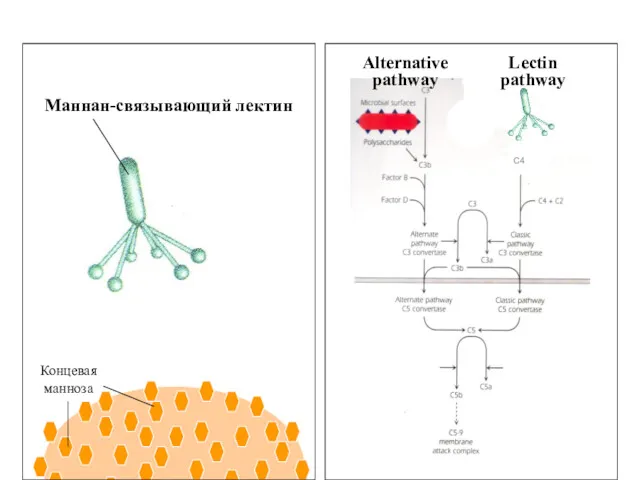
Маннан-связывающий лектин
Концевая
манноза
Alternative
pathway
Lectin
pathway
C4
Маннан-связывающий лектин
Концевая
манноза
Alternative
pathway
Lectin
pathway
C4
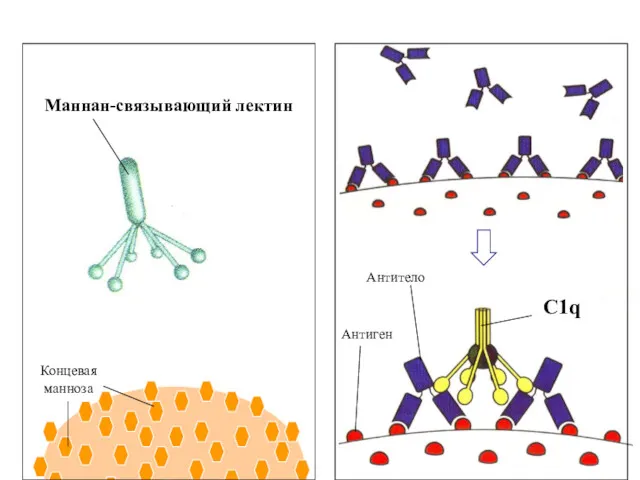
Маннан-связывающий лектин
Концевая
манноза
C1q
Антиген
Антитело
Маннан-связывающий лектин
Концевая
манноза
C1q
Антиген
Антитело
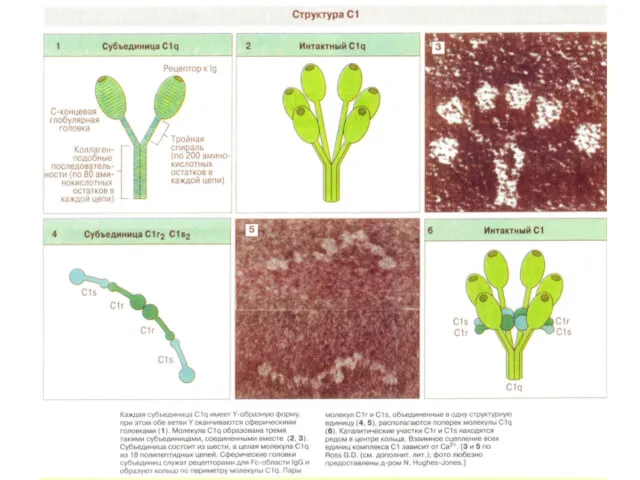
Рентгеновская структура C1q
Рентгеновская структура C1q
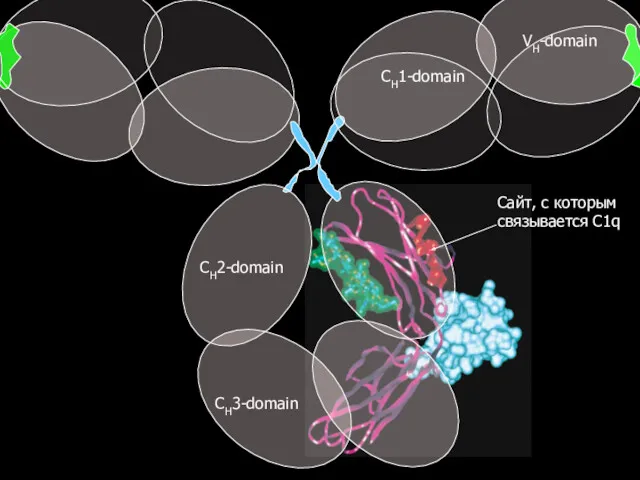
Сайт, с которым связывается C1q
CH3-domain
CH2-domain
CH1-domain
VH-domain
Сайт, с которым связывается C1q
CH3-domain
CH2-domain
CH1-domain
VH-domain
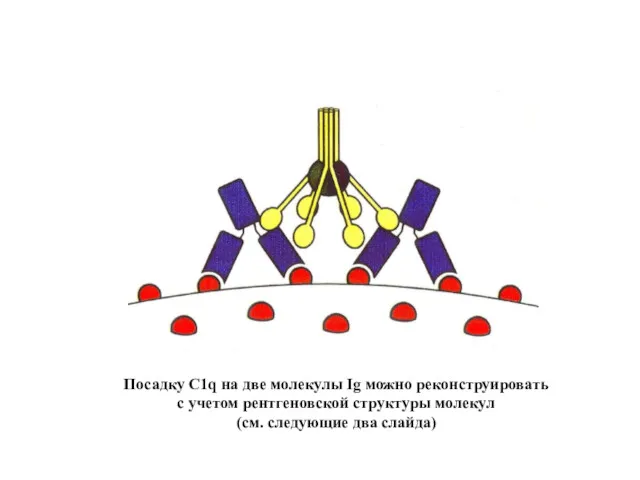
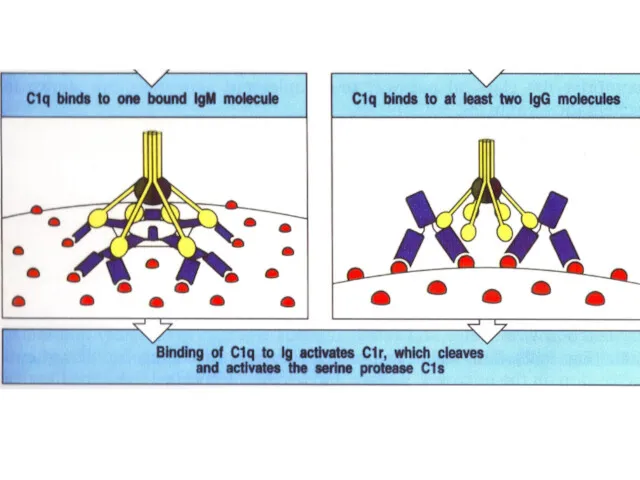
Посадку C1q на две молекулы Ig можно реконструировать
с учетом рентгеновской
Посадку C1q на две молекулы Ig можно реконструировать
с учетом рентгеновской
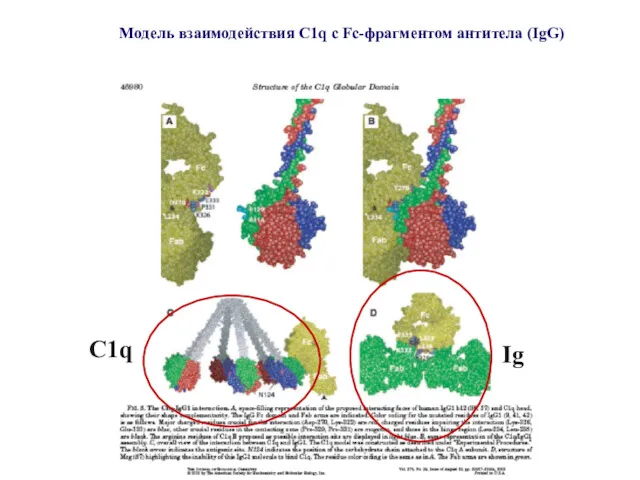
Модель взаимодействия C1q c Fc-фрагментом антитела (IgG)
C1q
Ig
Модель взаимодействия C1q c Fc-фрагментом антитела (IgG)
C1q
Ig
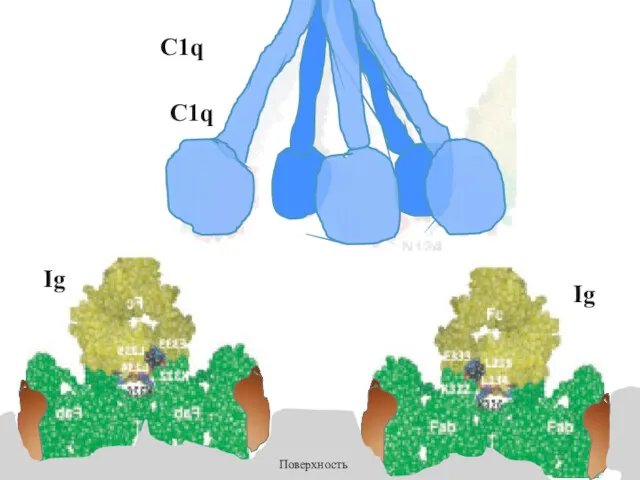
Ig
Ig
Поверхность
C1q
Ig
Ig
Поверхность
C1q
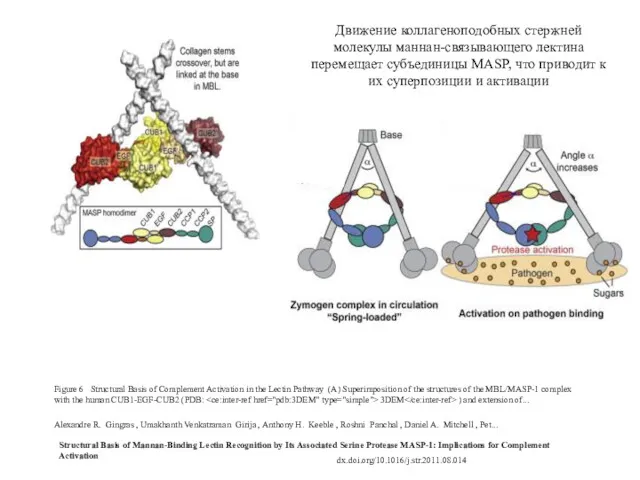
Figure 6 Structural Basis of Complement Activation in the Lectin Pathway (A)
Figure 6 Structural Basis of Complement Activation in the Lectin Pathway (A)
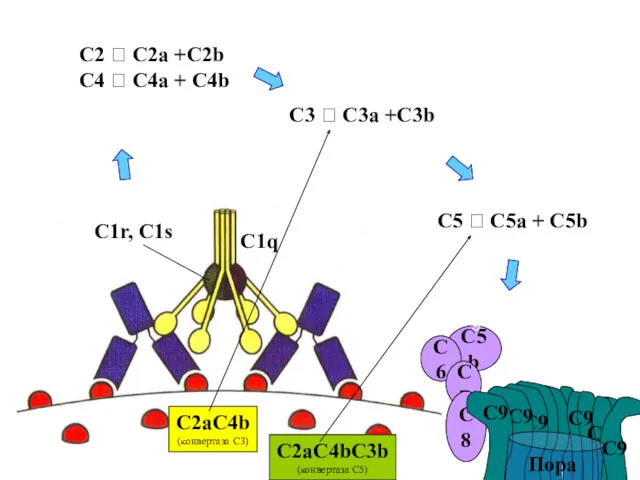
C9
C1q
C1r, C1s
C2 ? C2a +C2b
C4 ? C4a + C4b
C2aC4bC3b
(конвертаза С5)
C2aC4b
(конвертаза С3)
C3
C9
C1q
C1r, C1s
C2 ? C2a +C2b
C4 ? C4a + C4b
C2aC4bC3b
(конвертаза С5)
C2aC4b
(конвертаза С3)
C3
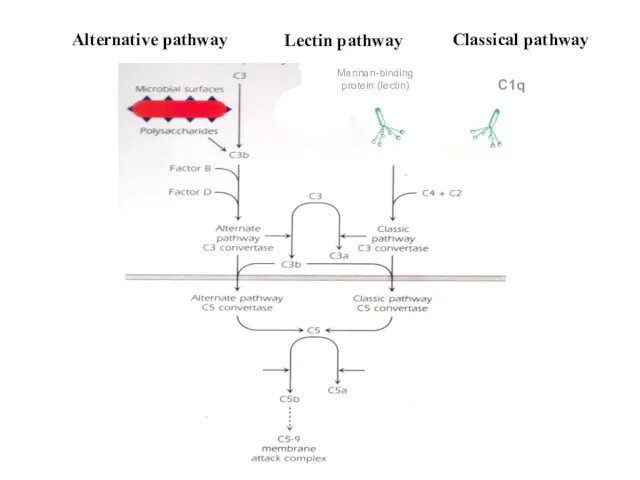
С1q
Mannan-binding
protein (lectin)
Alternative pathway
Lectin pathway
Classical pathway
С1q
Mannan-binding
protein (lectin)
Alternative pathway
Lectin pathway
Classical pathway
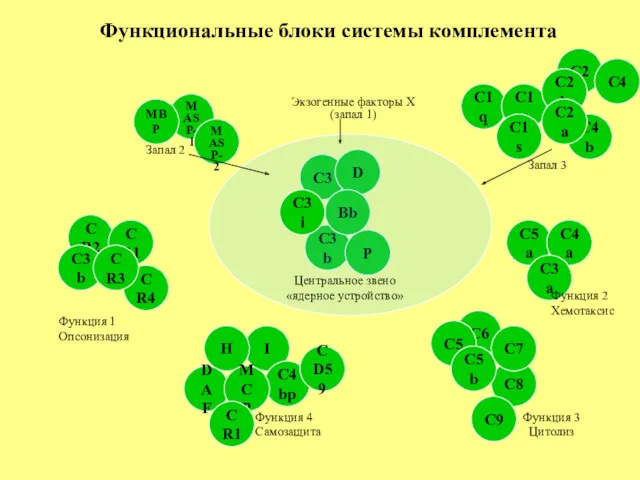
С3
С1q
С8
С2
С1r
С6
С4
С1s
С3b
С2b
С4b
С5
С5a
С5b
С9
С7
I
С4bp
H
P
DAF
D
Bb
MCP
СD59
MASP-
1
MASP-
2
CR1
CR2
CR4
H
Функциональные блоки системы комплемента
С2a
MBP
CR1
С4a
С3a
С3i
Центральное звено
«ядерное устройство»
Функция 2
Хемотаксис
Запал 3
Функция 3
Цитолиз
Функция
С3
С1q
С8
С2
С1r
С6
С4
С1s
С3b
С2b
С4b
С5
С5a
С5b
С9
С7
I
С4bp
H
P
DAF
D
Bb
MCP
СD59
MASP-
1
MASP-
2
CR1
CR2
CR4
H
Функциональные блоки системы комплемента
С2a
MBP
CR1
С4a
С3a
С3i
Центральное звено
«ядерное устройство»
Функция 2
Хемотаксис
Запал 3
Функция 3
Цитолиз
Функция


ЗАЩИТНЫЕ ФУНКЦИИ АНТИТЕЛ
Нейтрализация токсина
Удаление токсина в виде иммунных комплексов
Нейтрализация вируса
Препятствие адгезии
ЗАЩИТНЫЕ ФУНКЦИИ АНТИТЕЛ
Нейтрализация токсина
Удаление токсина в виде иммунных комплексов
Нейтрализация вируса
Препятствие адгезии
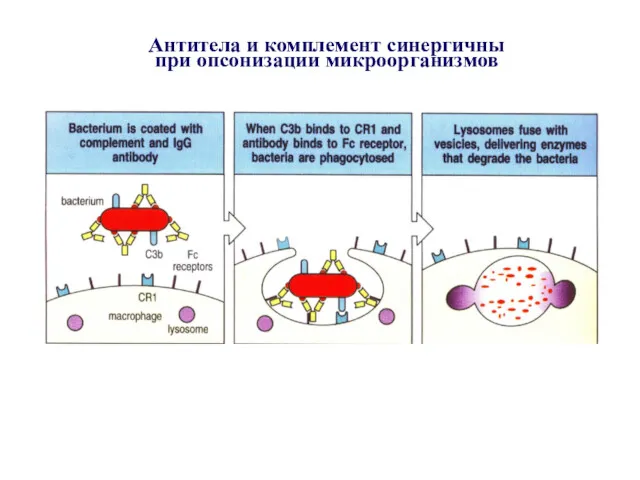
Антитела и комплемент синергичны
при опсонизации микроорганизмов
Антитела и комплемент синергичны
при опсонизации микроорганизмов
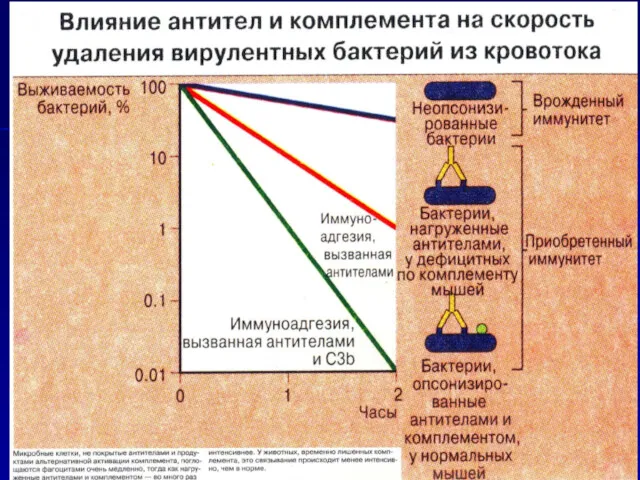

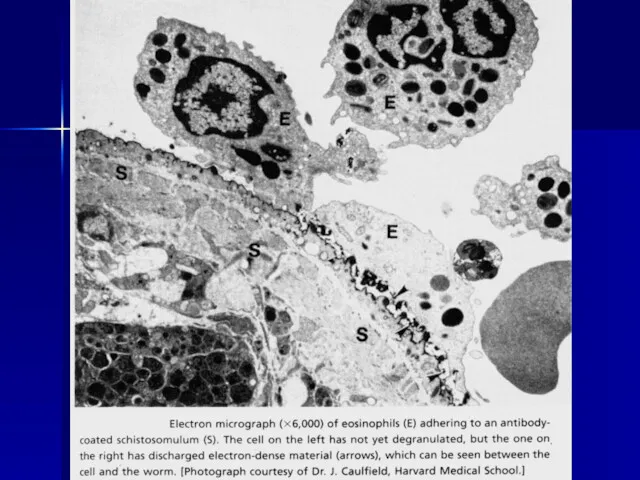
Антитела и комплемент повреждают даже многоклеточных паразитов
Антитела и комплемент повреждают даже многоклеточных паразитов
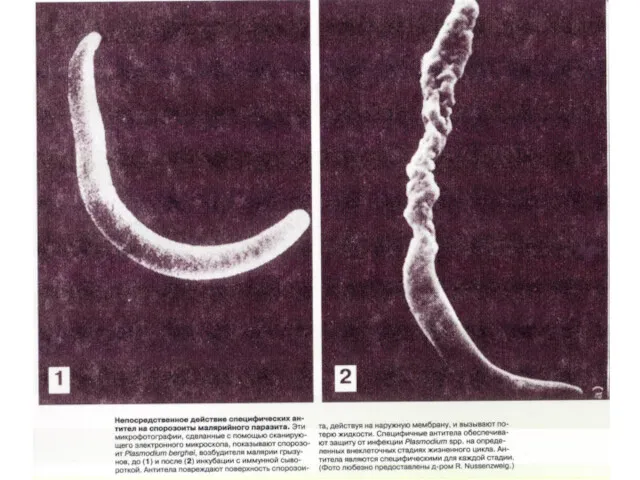

Антитела в роли «черной» метки,
по которой клетки-киллеры атакуют паразитов
Антитела в роли «черной» метки,
по которой клетки-киллеры атакуют паразитов
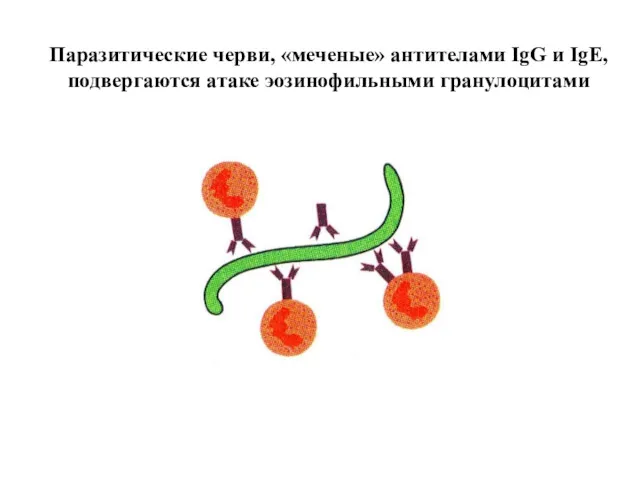
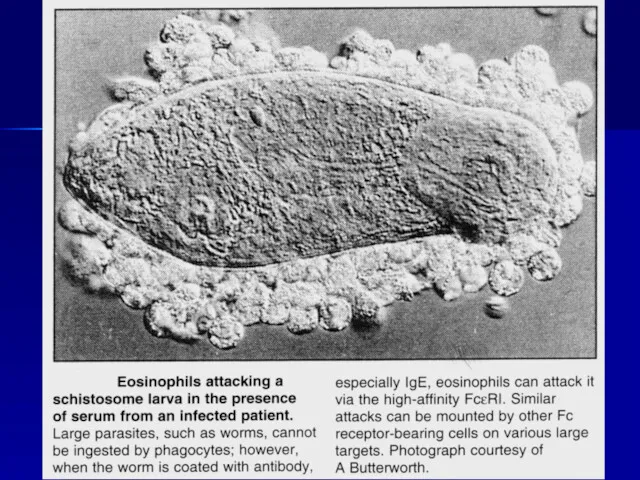
Паразитические черви, «меченые» антителами IgG и IgE,
подвергаются атаке эозинофильными гранулоцитами
Паразитические черви, «меченые» антителами IgG и IgE,
подвергаются атаке эозинофильными гранулоцитами
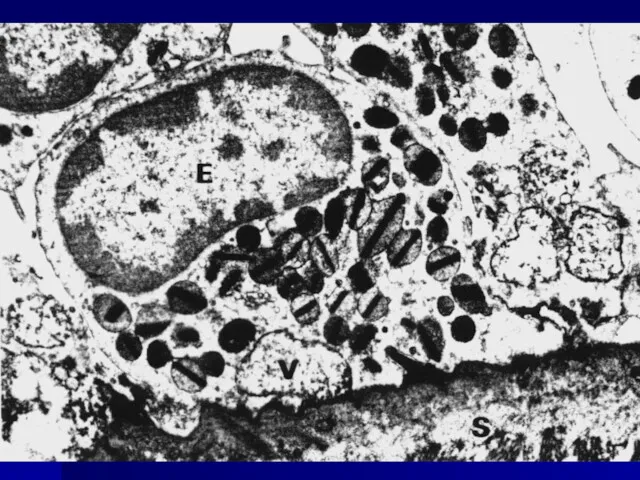
ЭОЗИНОФИЛ
ЭОЗИНОФИЛ

Eosinophil granule major basic protein
//en.wikipedia.org/wiki/Major_basic_protein
Образует кристаллическое ядро эозинофильной гранулы, участвует
Eosinophil granule major basic protein
//en.wikipedia.org/wiki/Major_basic_protein
Образует кристаллическое ядро эозинофильной гранулы, участвует

Антитела защищают нас не только внутри,
но и снаружи,
на границе
Антитела защищают нас не только внутри,
но и снаружи,
на границе
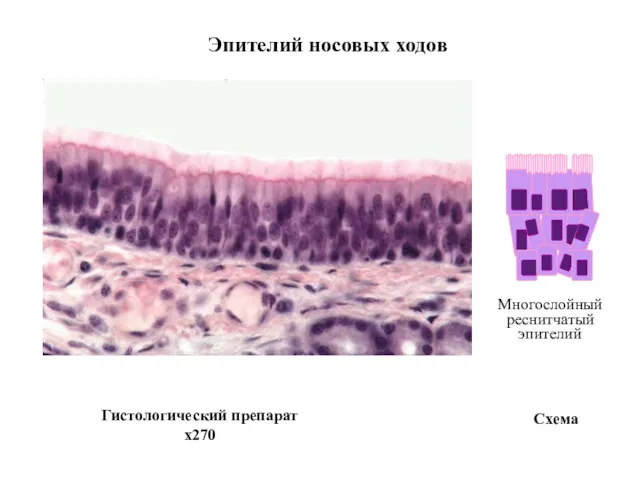
Многослойный
реснитчатый
эпителий
Схема
Эпителий носовых ходов
Гистологический препарат
x270
Схема
Многослойный
реснитчатый
эпителий
Схема
Эпителий носовых ходов
Гистологический препарат
x270
Схема
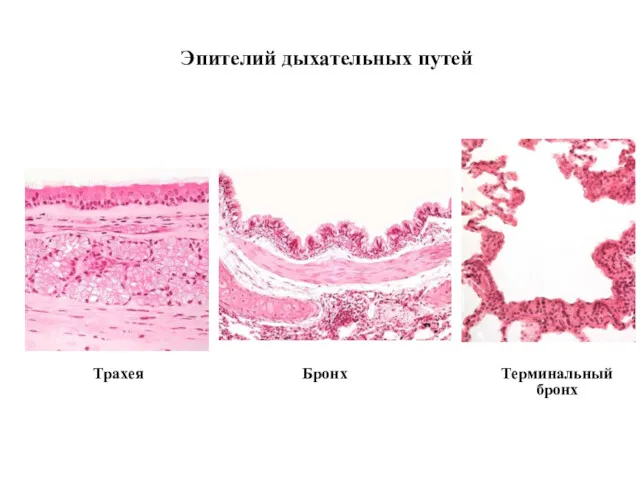
Трахея
Эпителий дыхательных путей
Трахея
Бронх
Терминальный
бронх
Трахея
Эпителий дыхательных путей
Трахея
Бронх
Терминальный
бронх
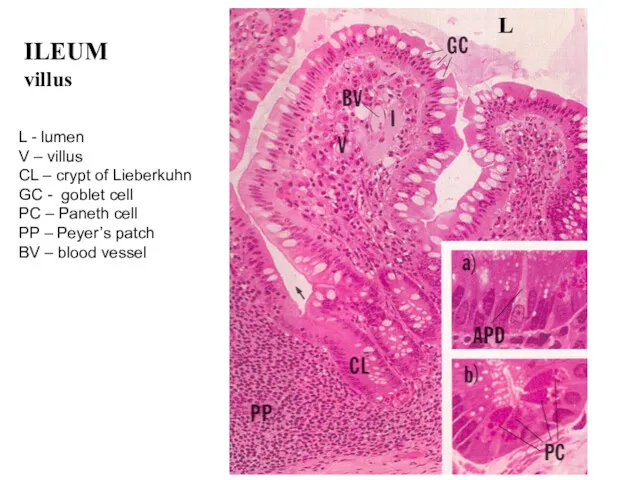
ILEUM
villus
L - lumen
V – villus
CL – crypt of Lieberkuhn
GC - goblet
ILEUM
villus
L - lumen
V – villus
CL – crypt of Lieberkuhn
GC - goblet
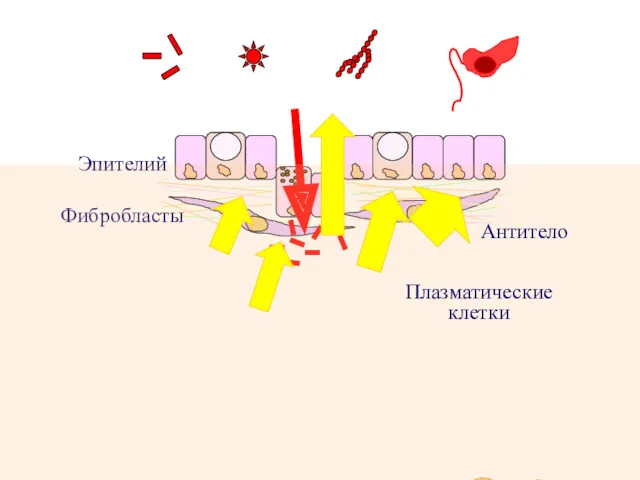
Эпителий
Фибробласты
Бактерии Вирусы Грибы Простейшие
Плазматические
клетки
Антитело
Эпителий
Фибробласты
Бактерии Вирусы Грибы Простейшие
Плазматические
клетки
Антитело
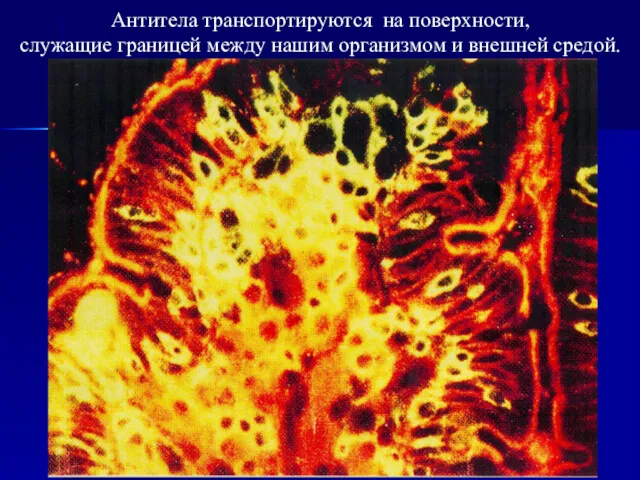
Антитела транспортируются на поверхности,
служащие границей между нашим организмом и внешней
Антитела транспортируются на поверхности,
служащие границей между нашим организмом и внешней
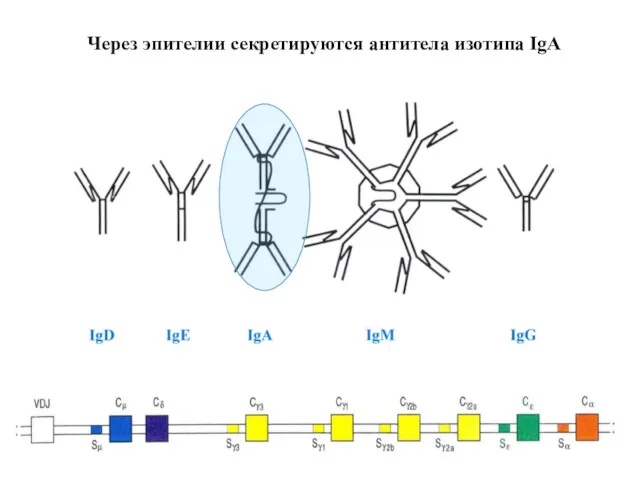
Через эпителии секретируются антитела изотипа IgA
Через эпителии секретируются антитела изотипа IgA
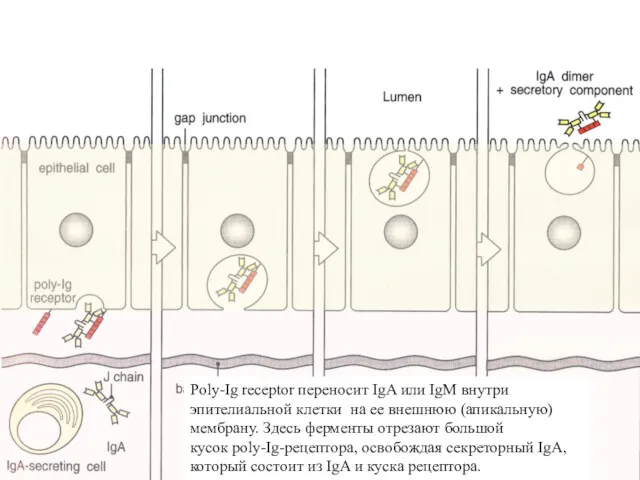
Poly-Ig receptor переносит IgA или IgM внутри эпителиальной клетки на ее
Poly-Ig receptor переносит IgA или IgM внутри эпителиальной клетки на ее

Особенно эффективно poly-Ig-рецептор переносит антитела через эпителий молочной железы и печени.
В
Особенно эффективно poly-Ig-рецептор переносит антитела через эпителий молочной железы и печени.
В

Что они делают?
Они очищаются от паразитов
Что они делают?
Они очищаются от паразитов

Антитела сообщают собаке (лошади):
О том, что на теле поселился паразит
Точно указывают
Антитела сообщают собаке (лошади):
О том, что на теле поселился паразит
Точно указывают
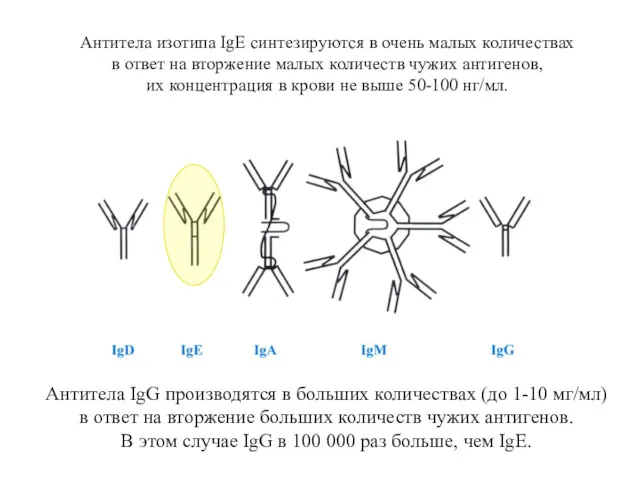
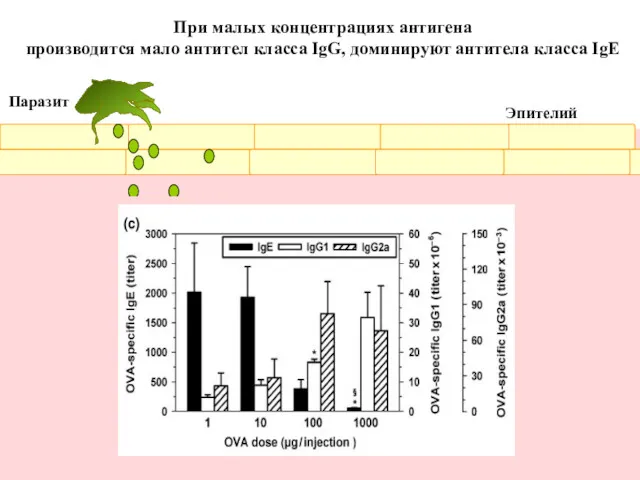
Антитела изотипа IgE синтезируются в очень малых количествах
в ответ на
Антитела изотипа IgE синтезируются в очень малых количествах
в ответ на
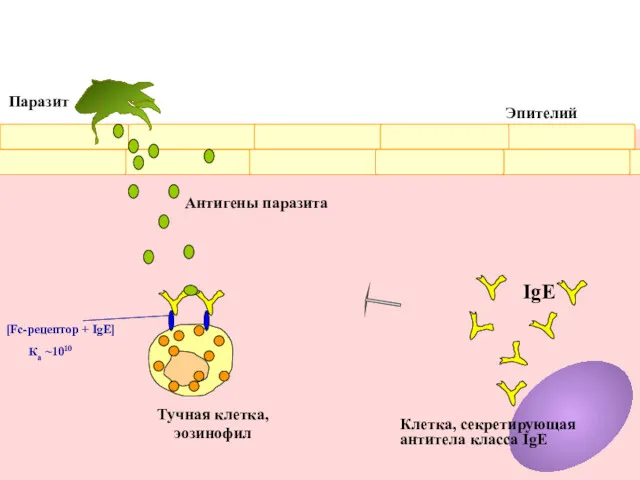
Клетка, секретирующая
антитела класса IgE
IgE
Ка ~1010
[Fc-рецептор + IgE]
Паразит
Антигены паразита
Тучная клетка,
эозинофил
Эпителий
Зуд и
Клетка, секретирующая
антитела класса IgE
IgE
Ка ~1010
[Fc-рецептор + IgE]
Паразит
Антигены паразита
Тучная клетка,
эозинофил
Эпителий
Зуд и
Клетка, секретирующая
антитела класса IgE
IgE
Ка ~1010
[Fc-рецептор + IgE]
Паразит
Антигены паразита
Тучная клетка,
эозинофил
Эпителий
Зуд и
Клетка, секретирующая
антитела класса IgE
IgE
Ка ~1010
[Fc-рецептор + IgE]
Паразит
Антигены паразита
Тучная клетка,
эозинофил
Эпителий
Зуд и

Антитела IgE служат рецепторами антигенов в мембране тучных клеток.
Это позволяет тучным
Антитела IgE служат рецепторами антигенов в мембране тучных клеток.
Это позволяет тучным

Изотипы антител имеют разные защитные функции
Мономеры IgD и IgM служат рецепторами
Изотипы антител имеют разные защитные функции
Мономеры IgD и IgM служат рецепторами
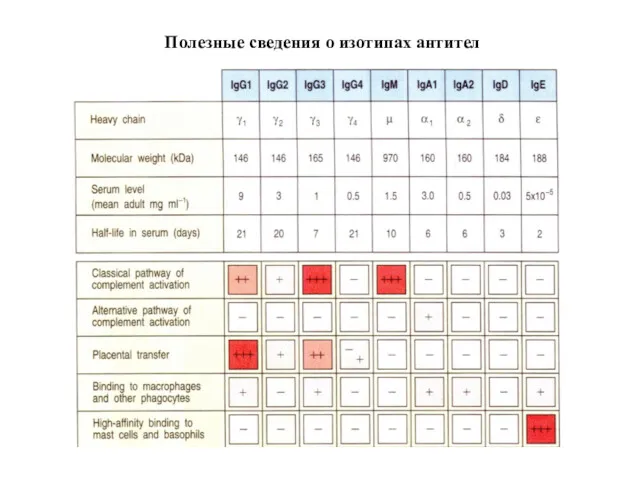
Полезные сведения о изотипах антител
Полезные сведения о изотипах антител
 Медикаментозное лечение глаукомы
Медикаментозное лечение глаукомы Терінің саңырауқұлақты аурулары. Трихофития
Терінің саңырауқұлақты аурулары. Трихофития Оказание первой помощи при укусах змей и насекомых
Оказание первой помощи при укусах змей и насекомых Эктопротезы, сравнительная характеристика отечественных и зарубежных аналогов
Эктопротезы, сравнительная характеристика отечественных и зарубежных аналогов Некроз. Атрофия
Некроз. Атрофия ВКР: Изготовление цельнолитого комбинированного металлопластмассового мостовидного протеза
ВКР: Изготовление цельнолитого комбинированного металлопластмассового мостовидного протеза Организация oнкологической службы в России
Организация oнкологической службы в России Желудочковые аритмии
Желудочковые аритмии Здоровье. Здоровый образ жизни
Здоровье. Здоровый образ жизни SW : The weakness of labor
SW : The weakness of labor Бронхиальная астма
Бронхиальная астма Уход за пациентом. ГОСТ Р 56819-2015. Профилактика пролежней
Уход за пациентом. ГОСТ Р 56819-2015. Профилактика пролежней Симптоматические язвы желудка и ДПК
Симптоматические язвы желудка и ДПК Мұрынның алдыңғы және артқы тампонадасын жасау техникасы
Мұрынның алдыңғы және артқы тампонадасын жасау техникасы Рак предстательной железы. Диагностика
Рак предстательной железы. Диагностика Клинические формы шизофрении и расстройств шизофренического спектра
Клинические формы шизофрении и расстройств шизофренического спектра Бедеуліктің дамуында қауіпқатер болып табылатын әйелдер арасындағы темекі тарту жайында ақпараттарды табыңыз
Бедеуліктің дамуында қауіпқатер болып табылатын әйелдер арасындағы темекі тарту жайында ақпараттарды табыңыз Болезни слюнных желез
Болезни слюнных желез Гемобластозы
Гемобластозы ДВС - синдром у детей
ДВС - синдром у детей Одонтогенные кисты
Одонтогенные кисты Травматические повреждения (часть 2)
Травматические повреждения (часть 2) Работа хирурга в условиях поликлиники и СВА
Работа хирурга в условиях поликлиники и СВА Воспаление. Этиология. Медиаторы. Виды. Острое воспаление
Воспаление. Этиология. Медиаторы. Виды. Острое воспаление Физическая работоспособность женщин
Физическая работоспособность женщин Мутагенез. Что такое ДНК
Мутагенез. Что такое ДНК Вагітність та інфекційні захворювання
Вагітність та інфекційні захворювання Причины и особенности употребления ПАВ (1)
Причины и особенности употребления ПАВ (1)