- Биохимия и молекулярная биология. Биоэнергетика (лекция 12)

Содержание
- 2. План лекции Окислительное фосфорилирование в дыхательной цепи Хемиосмотическая теория П. Митчелла. Мембранные транслоказы. Ингибиторы ЭТЦ. Разобщение
- 3. Дыхательная цепь Биоэнергетика Часть 2 Электронпереносящие комплексы митохондрий
- 4. Дыхательная цепь Биоэнергетика Часть 2 Окислительно-восстановительные реакции в электронтранспортной цепи В дыхательной цепи все реакции направлены
- 5. Дыхательная цепь Биоэнергетика Часть 2 Суммарная реакция, катализируемая дыхательной цепью, состоит в окислении NADH кислородом, приводящим
- 6. Дыхательная цепь Биоэнергетика Часть 2 Следовательно, при переносе одной пары электронов от NADH к кислороду через
- 7. Дыхательная цепь Биоэнергетика Часть 2 Расчет изменения ΔG0′ на каждом участке переноса электронов в ЭТЦ показывает,
- 8. Дыхательная цепь митохондрий Биоэнергетика Часть 2 Стандартная свободная энергия, выделяемая в окислительных реакциях, катализируемых комплексами ЭТЦ
- 9. Хемиосмотическая теория Биоэнергетика Часть 2 Сопряжение окисления и синтеза АТР в митохондриях
- 10. Хемиосмотическая теория Биоэнергетика Часть 2 Коэффициент Р/О Коэффициентом окислительного фосфорилирования (Р/О) называют отношение количества фосфорной кислоты
- 11. Хемиосмотическая теория Биоэнергетика Часть 2 Окислительное фосфорилирование Окислительным фосфорилированием называют синтез АТР из АDP и Н3РО4
- 12. Хемиосмотическая теория Биоэнергетика Часть 2 Хемиосмотическая теория сопряжения окисления и фосфорилирования Эта теория предложена в 1961
- 13. Хемиосмотическая теория Биоэнергетика Часть 2 При этом предполагается, что мембрана непроницаема для ионов, особенно протонов, их
- 14. Хемиосмотическая теория Биоэнергетика Часть 2 Создание электрохимического градиента ионов Н+ (протонного потенциала) ΔμН+ = ΔрН +
- 15. Хемиосмотическая теория Биоэнергетика Часть 2 Создание протонного градиента комплексами I, III и IV I, III и
- 16. Дыхательная цепь Биоэнергетика Часть 2 Окислительно-восстановительные реакции в электронтранспортной цепи Комплекс I. NADH + H+ +
- 17. Дыхательная цепь Биоэнергетика Часть 2 Окислительно-восстановительные реакции в электронтранспортной цепи Комплекс III. CoQH2 + 2Fe3+-цит.b →
- 18. Дыхательная цепь Биоэнергетика Часть 2 Окислительно-восстановительные реакции в электронтранспортной цепи Комплекс IV. 2Fe2+-цит.с + 2Fe3+-цит.а →
- 19. Хемиосмотическая теория Биоэнергетика Часть 2
- 20. Хемиосмотическая теория Биоэнергетика Часть 2 Cинтез АТР Градиент протонов, создающий разность химических и электрических потенциалов, является
- 21. Хемиосмотическая теория Биоэнергетика Часть 2 АТР-синтаза
- 22. Хемиосмотическая теория Биоэнергетика Часть 2 Cинтез АТР
- 23. Хемиосмотическая теория Биоэнергетика Часть 2 Cинтез АТР
- 24. Дыхательная цепь митохондрий Биоэнергетика Часть 2 Трансмембранный перенос веществ в митохондриях
- 25. Дыхательная цепь митохондрий Биоэнергетика Часть 2 Разобщение дыхания и окислительного фосфорилирования
- 26. Дыхательная цепь митохондрий Биоэнергетика Часть 2 Ингибиторы ЭТЦ
- 27. Дыхательная цепь митохондрий Биоэнергетика Часть 2 Ингибиторы ЭТЦ
- 28. Биологическое окисление Биоэнергетика Часть 2 Внемитохондриальные источники восстановительных эквивалентов
- 29. Биологическое окисление Биоэнергетика Часть 2 Глицерофосфатный челночный механизм, обеспечивающий окисление цитозольного NADH 1 – глцеральдегид-3-фосфатдегидрогеназа; 2
- 30. Биологическое окисление Биоэнергетика Часть 2 Малат-аспартатный челнок, обеспечивающий окисление цитозольного NADH
- 31. Биологическое окисление Биоэнергетика Часть 2
- 32. Биологическое окисление Биоэнергетика Часть 2 6 АТР 6 АТР 18 АТР 4 АТР Всего 38 АТР:
- 33. Биологическое окисление Биоэнергетика Часть 2 Энергетический баланс аэробного окисления глюкозы Аэробное окисление глюкозы до СО2 и
- 34. Биологическое окисление Биоэнергетика Часть 2 Общее уравнение дыхания, баланс АТР
- 35. Биологическое окисление Биоэнергетика Часть 2 Пути энергопродукции в клетках животных
- 37. Скачать презентацию

План лекции
Окислительное фосфорилирование в дыхательной цепи
Хемиосмотическая теория П. Митчелла.
Мембранные транслоказы.
Ингибиторы ЭТЦ.
Разобщение
План лекции
Окислительное фосфорилирование в дыхательной цепи
Хемиосмотическая теория П. Митчелла.
Мембранные транслоказы.
Ингибиторы ЭТЦ.
Разобщение
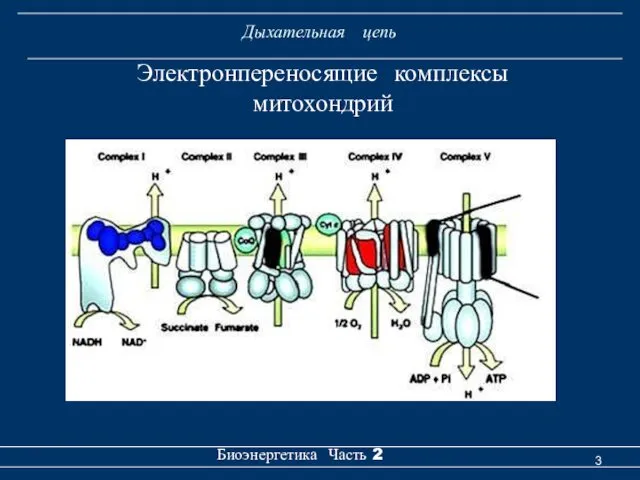
Дыхательная цепь
Биоэнергетика Часть 2
Электронпереносящие комплексы
митохондрий
Дыхательная цепь
Биоэнергетика Часть 2
Электронпереносящие комплексы
митохондрий
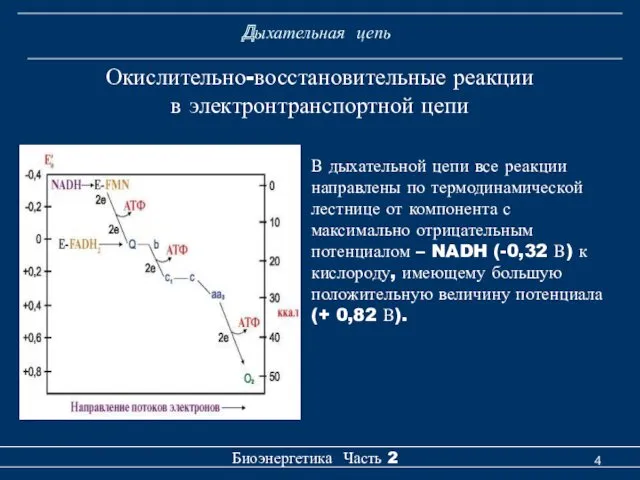
Дыхательная цепь
Биоэнергетика Часть 2
Окислительно-восстановительные реакции
в электронтранспортной цепи
В дыхательной цепи все
Дыхательная цепь
Биоэнергетика Часть 2
Окислительно-восстановительные реакции
в электронтранспортной цепи
В дыхательной цепи все

Дыхательная цепь
Биоэнергетика Часть 2
Суммарная реакция, катализируемая дыхательной цепью, состоит в
Дыхательная цепь
Биоэнергетика Часть 2
Суммарная реакция, катализируемая дыхательной цепью, состоит в

Дыхательная цепь
Биоэнергетика Часть 2
Следовательно, при переносе одной пары электронов от
Дыхательная цепь
Биоэнергетика Часть 2
Следовательно, при переносе одной пары электронов от

Дыхательная цепь
Биоэнергетика Часть 2
Расчет изменения ΔG0′ на каждом участке переноса
Дыхательная цепь
Биоэнергетика Часть 2
Расчет изменения ΔG0′ на каждом участке переноса
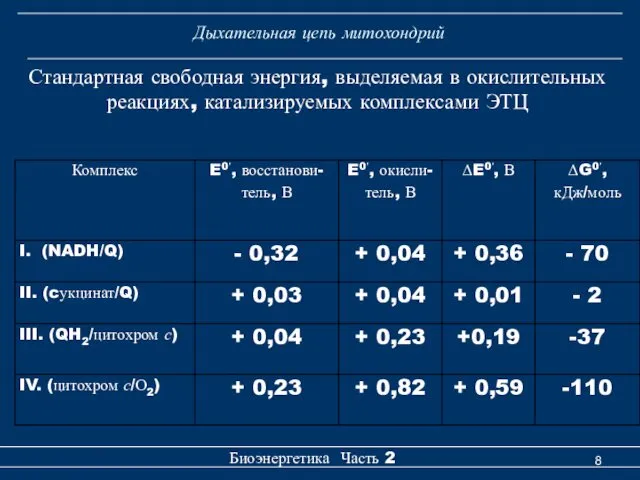
Дыхательная цепь митохондрий
Биоэнергетика Часть 2
Стандартная свободная энергия, выделяемая в окислительных
Дыхательная цепь митохондрий
Биоэнергетика Часть 2
Стандартная свободная энергия, выделяемая в окислительных
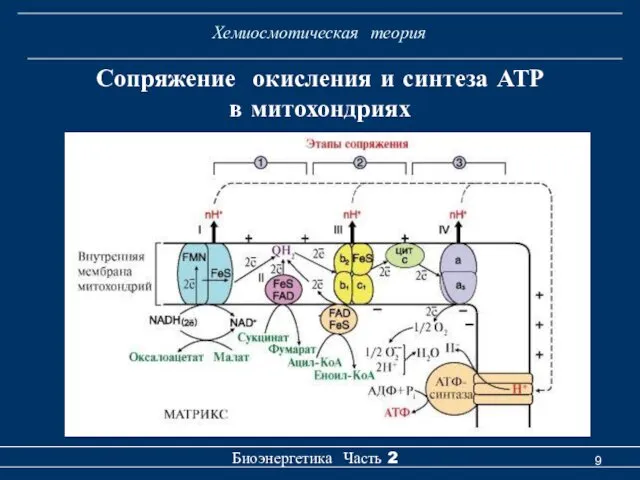
Хемиосмотическая теория
Биоэнергетика Часть 2
Сопряжение окисления и синтеза АТР
в
Хемиосмотическая теория
Биоэнергетика Часть 2
Сопряжение окисления и синтеза АТР
в

Хемиосмотическая теория
Биоэнергетика Часть 2
Коэффициент Р/О
Коэффициентом окислительного фосфорилирования (Р/О) называют
Хемиосмотическая теория
Биоэнергетика Часть 2
Коэффициент Р/О
Коэффициентом окислительного фосфорилирования (Р/О) называют

Хемиосмотическая теория
Биоэнергетика Часть 2
Окислительное фосфорилирование
Окислительным фосфорилированием называют синтез АТР
Хемиосмотическая теория
Биоэнергетика Часть 2
Окислительное фосфорилирование
Окислительным фосфорилированием называют синтез АТР

Хемиосмотическая теория
Биоэнергетика Часть 2
Хемиосмотическая теория сопряжения окисления и фосфорилирования
Эта теория
Хемиосмотическая теория
Биоэнергетика Часть 2
Хемиосмотическая теория сопряжения окисления и фосфорилирования
Эта теория

Хемиосмотическая теория
Биоэнергетика Часть 2
При этом предполагается, что мембрана непроницаема для
Хемиосмотическая теория
Биоэнергетика Часть 2
При этом предполагается, что мембрана непроницаема для

Хемиосмотическая теория
Биоэнергетика Часть 2
Создание электрохимического градиента ионов Н+ (протонного
Хемиосмотическая теория
Биоэнергетика Часть 2
Создание электрохимического градиента ионов Н+ (протонного
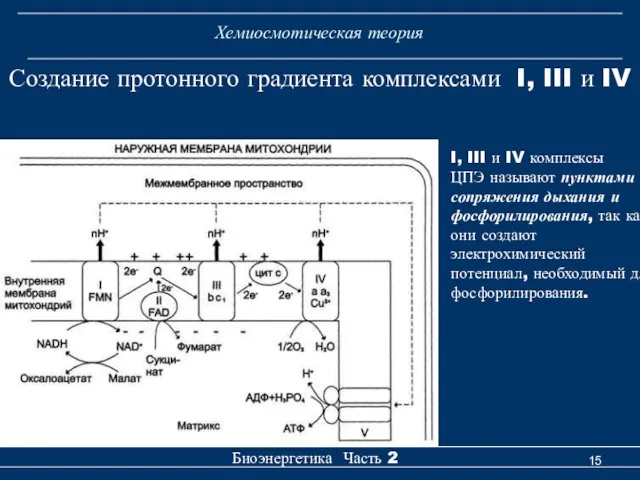
Хемиосмотическая теория
Биоэнергетика Часть 2
Создание протонного градиента комплексами I, III
Хемиосмотическая теория
Биоэнергетика Часть 2
Создание протонного градиента комплексами I, III

Дыхательная цепь
Биоэнергетика Часть 2
Окислительно-восстановительные реакции
в электронтранспортной цепи
Комплекс I.
NADH + H+
Дыхательная цепь
Биоэнергетика Часть 2
Окислительно-восстановительные реакции
в электронтранспортной цепи
Комплекс I.
NADH + H+

Дыхательная цепь
Биоэнергетика Часть 2
Окислительно-восстановительные реакции
в электронтранспортной цепи
Комплекс III.
CoQH2 + 2Fe3+-цит.b
Дыхательная цепь
Биоэнергетика Часть 2
Окислительно-восстановительные реакции
в электронтранспортной цепи
Комплекс III.
CoQH2 + 2Fe3+-цит.b

Дыхательная цепь
Биоэнергетика Часть 2
Окислительно-восстановительные реакции
в электронтранспортной цепи
Комплекс IV.
2Fe2+-цит.с + 2Fe3+-цит.а
Дыхательная цепь
Биоэнергетика Часть 2
Окислительно-восстановительные реакции
в электронтранспортной цепи
Комплекс IV.
2Fe2+-цит.с + 2Fe3+-цит.а
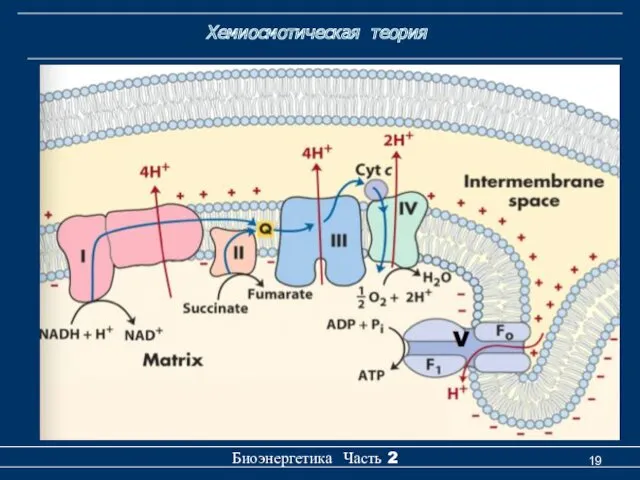
Хемиосмотическая теория
Биоэнергетика Часть 2
Хемиосмотическая теория
Биоэнергетика Часть 2

Хемиосмотическая теория
Биоэнергетика Часть 2
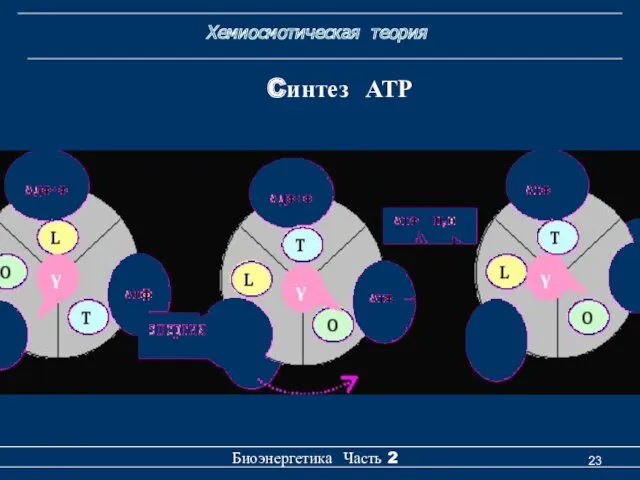
Cинтез АТР
Градиент протонов, создающий разность химических и
Хемиосмотическая теория
Биоэнергетика Часть 2
Cинтез АТР
Градиент протонов, создающий разность химических и
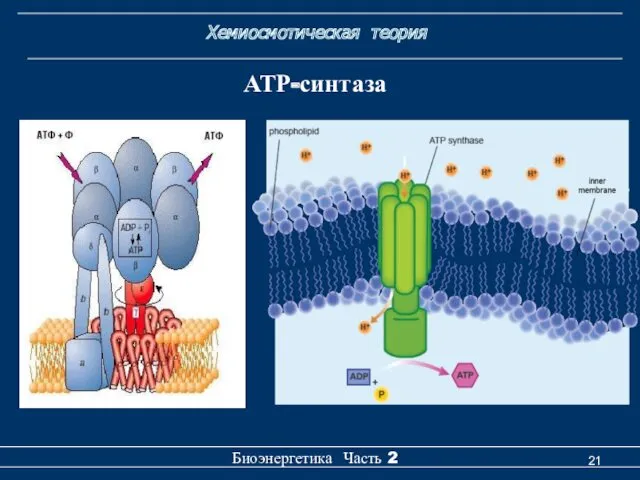
Хемиосмотическая теория
Биоэнергетика Часть 2
АТР-синтаза
Хемиосмотическая теория
Биоэнергетика Часть 2
АТР-синтаза
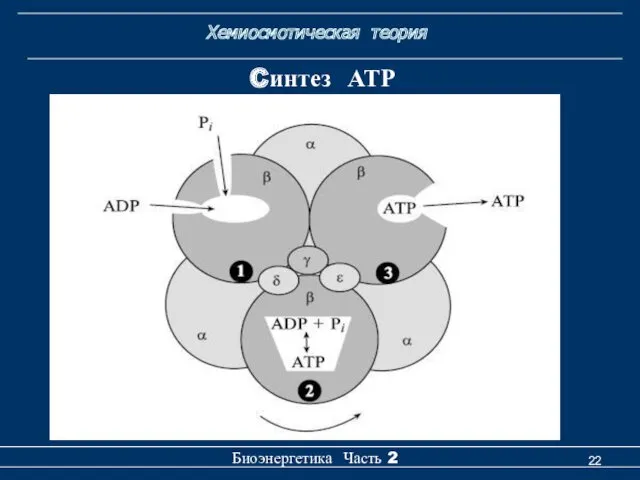
Хемиосмотическая теория
Биоэнергетика Часть 2
Cинтез АТР
Хемиосмотическая теория
Биоэнергетика Часть 2
Cинтез АТР
Хемиосмотическая теория
Биоэнергетика Часть 2
Cинтез АТР
Хемиосмотическая теория
Биоэнергетика Часть 2
Cинтез АТР
Дыхательная цепь митохондрий
Биоэнергетика Часть 2
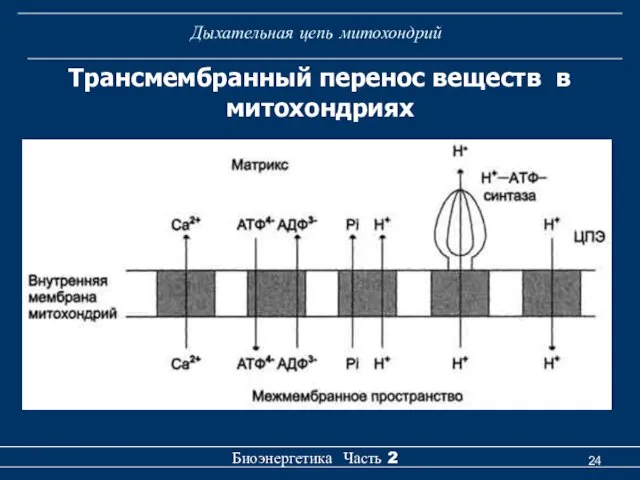
Трансмембранный перенос веществ в митохондриях
Дыхательная цепь митохондрий
Биоэнергетика Часть 2
Трансмембранный перенос веществ в митохондриях
Дыхательная цепь митохондрий
Биоэнергетика Часть 2
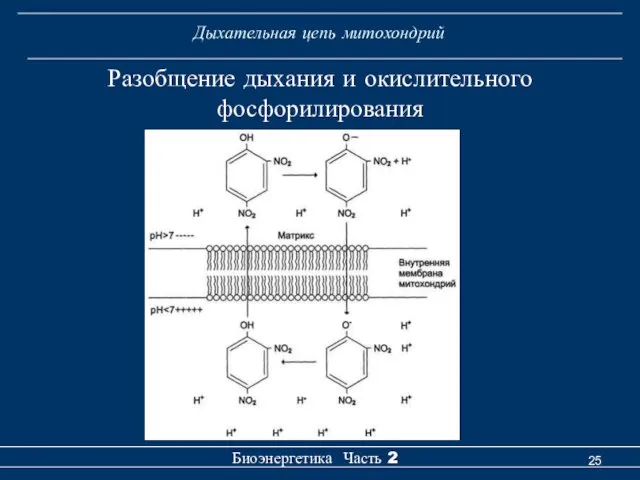
Разобщение дыхания и окислительного
фосфорилирования
Дыхательная цепь митохондрий
Биоэнергетика Часть 2
Разобщение дыхания и окислительного
фосфорилирования
Дыхательная цепь митохондрий
Биоэнергетика Часть 2
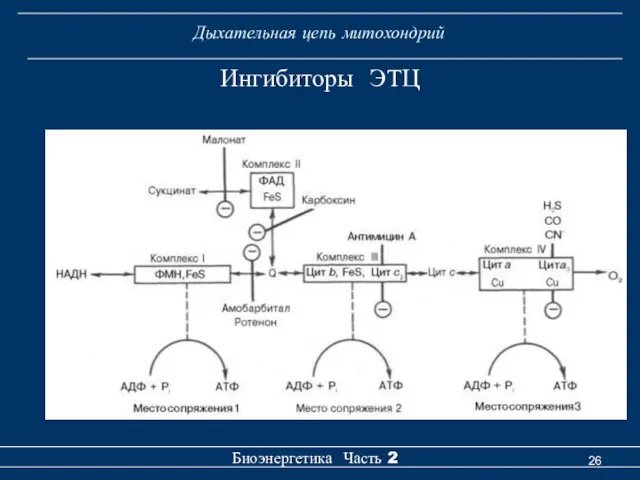
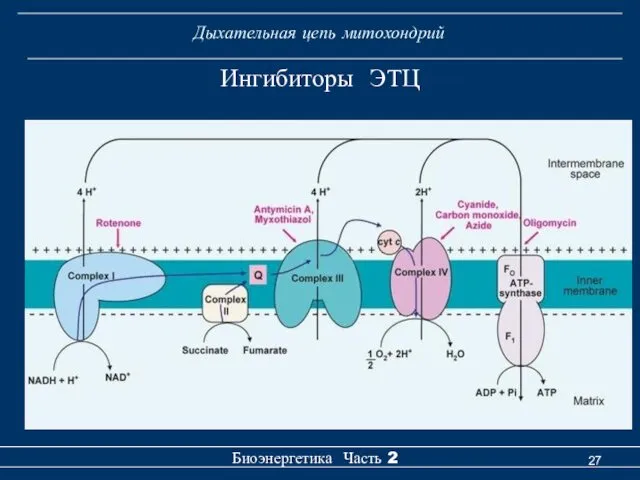
Ингибиторы ЭТЦ
Дыхательная цепь митохондрий
Биоэнергетика Часть 2
Ингибиторы ЭТЦ
Дыхательная цепь митохондрий
Биоэнергетика Часть 2
Ингибиторы ЭТЦ
Дыхательная цепь митохондрий
Биоэнергетика Часть 2
Ингибиторы ЭТЦ
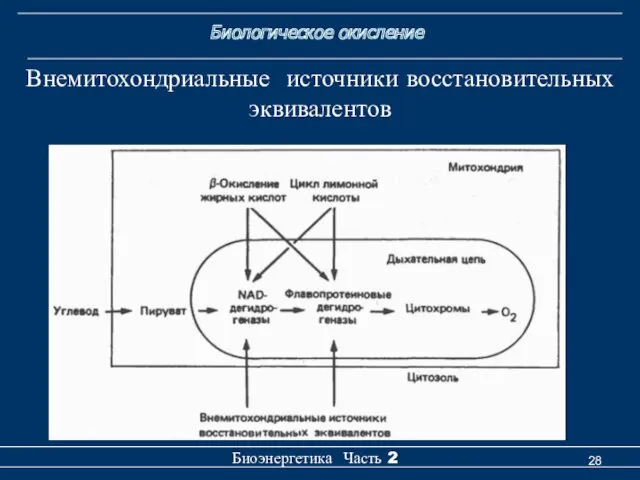
Биологическое окисление
Биоэнергетика Часть 2
Внемитохондриальные источники восстановительных эквивалентов
Биологическое окисление
Биоэнергетика Часть 2
Внемитохондриальные источники восстановительных эквивалентов
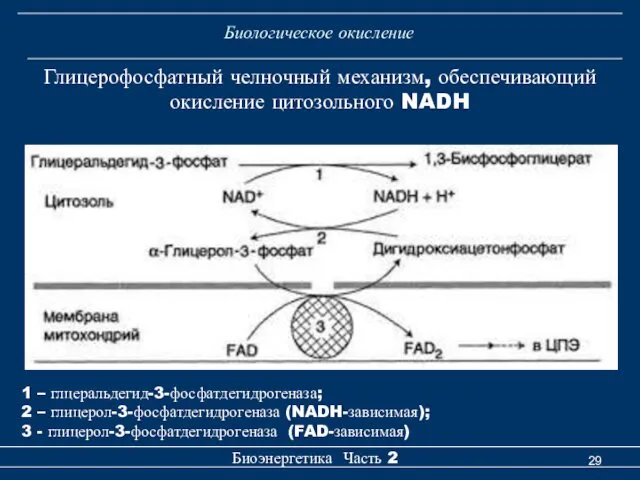
Биологическое окисление
Биоэнергетика Часть 2
Глицерофосфатный челночный механизм, обеспечивающий окисление цитозольного
Биологическое окисление
Биоэнергетика Часть 2
Глицерофосфатный челночный механизм, обеспечивающий окисление цитозольного
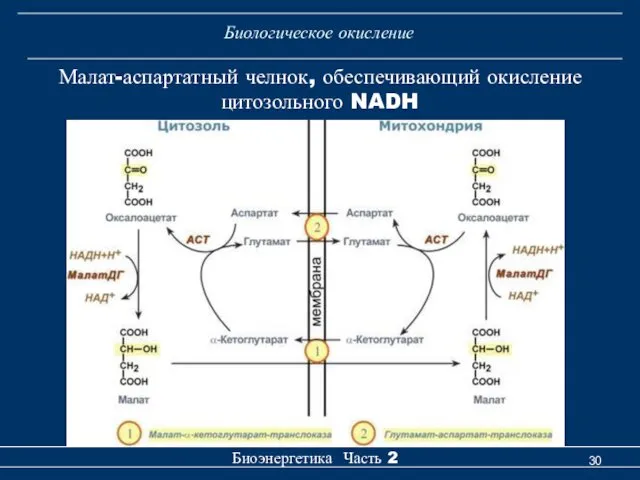
Биологическое окисление
Биоэнергетика Часть 2
Малат-аспартатный челнок, обеспечивающий окисление цитозольного NADH
Биологическое окисление
Биоэнергетика Часть 2
Малат-аспартатный челнок, обеспечивающий окисление цитозольного NADH
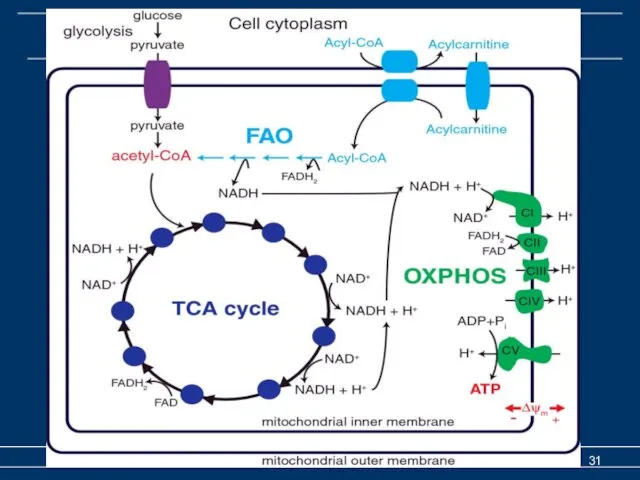
Биологическое окисление
Биоэнергетика Часть 2
Биологическое окисление
Биоэнергетика Часть 2
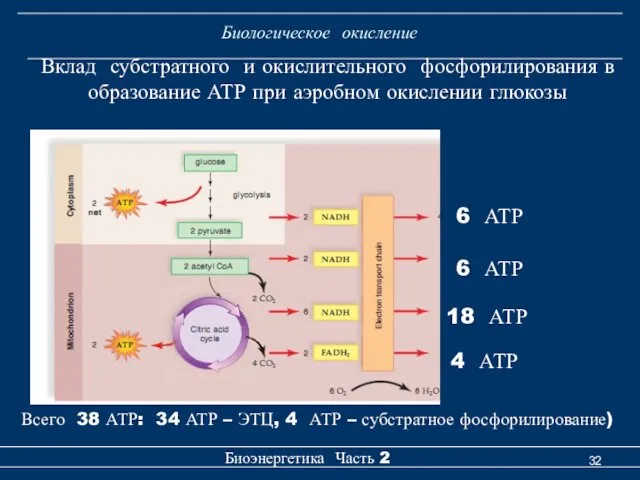
Биологическое окисление
Биоэнергетика Часть 2
6 АТР
6 АТР
18 АТР
4 АТР
Всего 38 АТР:
Биологическое окисление
Биоэнергетика Часть 2
6 АТР
6 АТР
18 АТР
4 АТР
Всего 38 АТР:

Биологическое окисление
Биоэнергетика Часть 2
Энергетический баланс аэробного окисления глюкозы
Аэробное окисление
Биологическое окисление
Биоэнергетика Часть 2
Энергетический баланс аэробного окисления глюкозы
Аэробное окисление
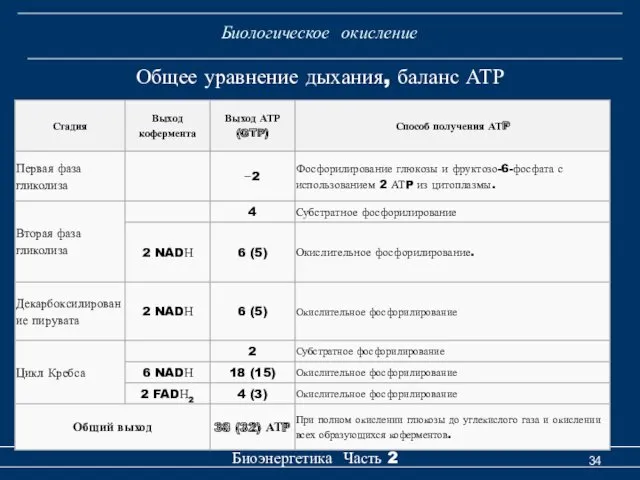
Биологическое окисление
Биоэнергетика Часть 2
Общее уравнение дыхания, баланс АТР
Биологическое окисление
Биоэнергетика Часть 2
Общее уравнение дыхания, баланс АТР
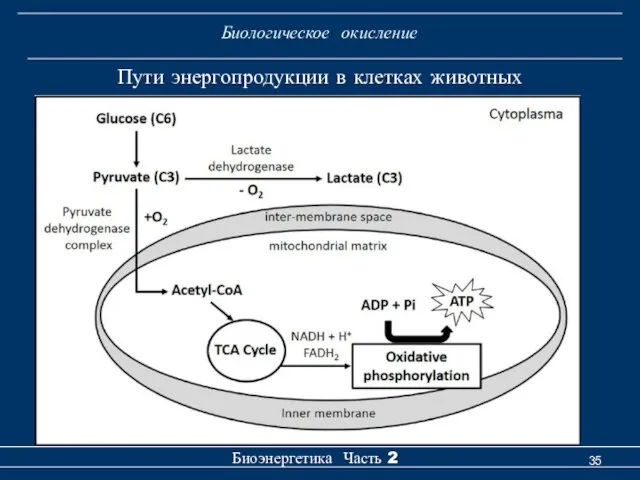
Биологическое окисление
Биоэнергетика Часть 2
Пути энергопродукции в клетках животных
Биологическое окисление
Биоэнергетика Часть 2
Пути энергопродукции в клетках животных
 Электронно-дырочный переход. Транзистор
Электронно-дырочный переход. Транзистор Сканеры. Устройства ввода информации в компьютер
Сканеры. Устройства ввода информации в компьютер Santa Claus or Saint Nicolas
Santa Claus or Saint Nicolas Отвод поверхностных вод. Водосборно-водоотводные сооружения (водоотводы)
Отвод поверхностных вод. Водосборно-водоотводные сооружения (водоотводы) Презентация Как помочь ребенку хорошо учиться
Презентация Как помочь ребенку хорошо учиться Курганский филиал РАНХиГС. Основы проекта
Курганский филиал РАНХиГС. Основы проекта Формирование соединений при контактной точечной сварке. (Лекция 4)
Формирование соединений при контактной точечной сварке. (Лекция 4) Информационные технологии в менеджменте качества
Информационные технологии в менеджменте качества Италия. Рим
Италия. Рим Мотивация дошкольников
Мотивация дошкольников Скользящая опалубка
Скользящая опалубка Свидетель эпохи. Жак-Луи Давид
Свидетель эпохи. Жак-Луи Давид Арифметические операции в позиционных системах счисления
Арифметические операции в позиционных системах счисления Роговица. Свойства и функции роговицы
Роговица. Свойства и функции роговицы Вторая фаза метаболизма ксенобиотиков и эндогенных соединений
Вторая фаза метаболизма ксенобиотиков и эндогенных соединений Сухопутные войска, их состав и предназначение
Сухопутные войска, их состав и предназначение Покров Пресвятой Богородицы
Покров Пресвятой Богородицы Проект планировки территории и проект межевания территории
Проект планировки территории и проект межевания территории Понятие, система, задачи, принципы уголовного права. Уголовная политика. Наука уголовного права
Понятие, система, задачи, принципы уголовного права. Уголовная политика. Наука уголовного права Интегрированный урок по химии и географии Почва 8 класс
Интегрированный урок по химии и географии Почва 8 класс 20231029_uroki_stariny_glubokoy
20231029_uroki_stariny_glubokoy Социализация детей с расстройствами аутистического спектра в сюжетно-ролевой игре
Социализация детей с расстройствами аутистического спектра в сюжетно-ролевой игре Бизнес идея создания сувенирной лавки РАЙские сувениры
Бизнес идея создания сувенирной лавки РАЙские сувениры Атеросклероз. Факторы риска
Атеросклероз. Факторы риска Суицид как форма девиантного поведения подростков
Суицид как форма девиантного поведения подростков Выбор марки насосов
Выбор марки насосов Обработка персональных данных
Обработка персональных данных Размещение рекламы в фильме Лицо со шрамом
Размещение рекламы в фильме Лицо со шрамом