- Биосинтез белка

Содержание
- 2. Реализация генетической информации
- 3. Генетический код Генетическая информация – сумма признаков. В основе - информация о строении (аа-последовательности) белков. Язык,
- 4. Генетический код – это запись только о последовательности аминокислот в белках. Кроме этого, в ДНК записана
- 5. История Уотсон-Крик 1953 – двойная спираль ДНК 1954 – Георгий Гамов. Предложил в качестве механизма кодирования
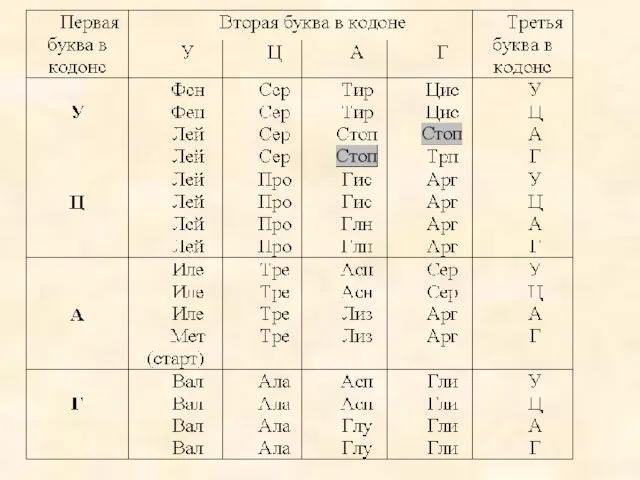
- 6. Свойства генетического кода: Триплетность (почему именно – объяснить). 2) Однозначность (исключения – fMet/Met – AUG, Sec
- 7. 5) Компактность – отсутствие знаков препинания внутри генов, знаки препинания только между генами – 3 стоп-кодона
- 8. Рамки считывания Одна и та же нуклеотидная последовательность может кодировать совершенно различные последовательности аминокислот. Мутации со
- 9. Пример – в 1974 г. была секвенирована ДНК фага φX174. Перекрывание генов в геноме фага φX174:
- 11. Что содержится в ДНК Длина ДНК человека ок. 3.2 х 10 пар нуклеотидов. В ней ок.
- 12. антикодон Акцепторый (ССА)-конец Строение тРНК 3’ 5’ 3’ 5’ Пространственная структура тРНК впервые установлена с помощью
- 13. Вторичная структура тРНК. Консервативные (красного цвета) и полуконсервативные нуклеотиды (синего цвета; R – пурин, Y –
- 14. Структурные элементы, отличающие инициаторную тРНК от элонгаторных.
- 15. Особенностью тРНК является присутствие минорных нуклеозидов. Структура митохондриальных тРНК млекопитающих сильно отличается от канонической из-за необычных
- 16. Типы минорных нуклеозидов. 1. Стандартные – встречаются почти во всех тРНК Дигидроуридин - D Риботимидин -
- 17. 2. Метилированные основания – встречаются почти во всех тРНК. 3. Гипермодифицированные основания (обычно с 3’- стороны
- 18. Рекогниция Присоединение аминокислотного остатка к тРНК Аминоацил-тРНК-синтетазы Изоакцепторные тРНК
- 19. (1) aaRS + аа + АТР → aaRS•аа-АМР + РРi. (2) aaRS•аа-АМР + тРНК → аа-тРНК
- 20. Нестандартные ситуации 1. Во многих бактериях и органеллах эукариот отсутствуют GlnRS и АsnRS, а в ряде
- 21. 2. Синтез 21-й аминокислоты селеноцистеина (Sec). PLP - пиридоксальфосфат Спец. киназа В обоих случаях донором селена
- 22. 3. Другая нестандартная аминокислота, кодируемая генами некоторых метаногенных архебактерий, – производное лизина пирролизин (Pyl) – включается
- 23. АРСазы Мол. массы - в диапазоне 37–250 кДа, четвертичная структура - от мономеров до гомо- и
- 24. Ферменты разных классов отличаются устройством активного центра и способами связывания акцепторной ветви тРНК. Все ферменты класса
- 25. Структура активных центров аминоацил-тРНК-синтетаз, принадлежащих классу I (GlnRS) и классу II (AspRS). Характерные структурные мотивы выделены
- 26. Для большинства синтетаз (за исключением 4-х ферментов класса I – ArgRS, GlnRS, GluRS и LysRS1 )
- 27. Узнавание тРНК АРСазами Методы исследования - генетические in vivo и кинетических экспериментов in vitro с мутантными
- 28. Узнавание боковых групп аминокислот обеспечивается изначальной комплементарностью многих ферментов субстрату по известной модели «замка и ключа».
- 29. Селективность отбора аминокислот некоторыми ферментами (CysRS, MetRS, HisRS и ProRS), обеспечивается индуцированным соответствием («рука-перчатка»): связывание специфичной
- 30. Установлены элементы узнавания тРНК для всех 20-ти синтетаз из E. сoli, а также некоторых АРС ряда
- 31. Распределение элементов узнавания тРНК E. coli аминоацил-тРНК-синтетазами класса I (а) и класса II (б) Выделенная серым
- 32. Антикодон не важен для узнавания тРНК E. сoli, специфичных к Leu, Ser и Ala. Это неудивительно
- 34. Рибосома - сложнейшая молекулярная машина клетки, ее функция состоит в том, чтобы переводить (транслировать) генетическую информацию,
- 35. мРНК Общие черты строения: односпиральность, направление чтения и написания 5’-3’, НТП, старт-кодон, кодирующая последовательность, стоп-кодон, 3’-НТП
- 36. Особенности мРНК эукариот : Одноцистронные Нет ШД-послед-тей. Кэп на 5’-конце. Поли(А) – послед-ть на 3’-конце. Полицистронными
- 37. Структура кэпа
- 38. тРНК – сходство у всех организмов Инициаторные тРНК аминоацилируются только остатками Met и «работают» только на
- 39. Факторы трансляции - одно- или многосубъединичные белки, подразделяющиеся на факторы инициации (IF), факторы элонгации (EF) и
- 40. Инициация – процесс, приводящий к образованию комплекса, в котором старт-кодон AUG находится в пептидильном (Р) –
- 41. Структурные элементы, отличающие инициаторную тРНК от элонгаторных.
- 42. Схема инициации у прокариот IF2 взаимод. с тРНК IF1 стимул IF2 и IF3 IF3 помогает ШД
- 43. Схема инициации у эукариот IF1 IF3(функц. аналог) IF2 eIF4F – кэп-связывающий комплекс факторов eIF4E – кэп-узнающий
- 44. eIF4A – АТР-завис. геликаза eIF4B, Н – ассистируют ей E eIF4E-Кэп-связывающий белок
- 46. Образование тройного комплекса и замена GDP на GTP в eIF2 eIF2 GDP
- 47. Активируется в стрессовых условиях (тепловой шок, голодание и т.д.) Образуется прочный комплекс, где оказывается связанным весь
- 48. Факторы инициации эукариот Фактор Гомологи Бакт Археи Субъединицы Функции eIF1 1 (13 kDa) Предотвращает инициацию на
- 49. Фактор Субъединицы Функции eIF4E нет 1 (25 kDa) Связывается с кэпом eIF4F нет Комплекс eIF4E, A
- 50. Цикл элонгации. eEF1 – EF-Tu eEF2 – EF-G Аналогичны по функции, но не работают в гетерологичных
- 51. 2. Цикл элонгации полипептидной цепи.
- 52. Р-участок А-участок Р-участок А-участок Схема образования пептидной связи
- 53. 3. Терминация трансляции наступает, когда в аминоацильном (А)-участке оказывается один из 3-х стоп кодонов – UGA,
- 54. Молекулярная мимикрия – сходство структуры RF и тРНК. Связываясь с рибосомой, RF одним своим фрагментом, напоминающим
- 55. >100 Å anticodon CCA-3’end 76Å тРНК eRF1
- 56. Субчастицы готовы к реинициации на этой же мРНК или к инициации трансляции новой мРНК. Общая схема
- 57. Характерные особенности терминации у прокариот: 1) освобождение синтезированного полипептида происходит до гидролиза GTP и без участия
- 58. У эукариот оба фактора терминации действуют кооперативно и скоодинированно: 1) eRF1 имеет высокое сродство к eRF3,
- 59. Гидролиз GTP приводит к изменению конформации терминационного комплекса таким образом, что универсальная для всех организмов последовательность
- 60. Рециклинг - диссоциация мРНК и деацилированной тРНК и последующая диссоциация рибосом на субчастицы, которые затем снова
- 61. У эукариот и архей специализированного фактора рециклинга нет. Диссоциация 80S рибосом эукариот на субчастицы после завершения
- 62. Отклонения от канонических правил Неканоническая инициация – трансляционные энхансеры в мРНК и IRES – элементы некоторых
- 63. >100 Å anticodon CCA-3’end 76Å тРНК eRF1
- 64. Аминокислота селеноцистеин - биологическая форма селена. Ее кодирует триплет UGA, являющийся обычно стоп-кодоном. Специальный механизм, благодаря
- 65. К человеческим селенопротеинам относят: Иодтирониндеиодиназы 1—3: DIO1Иодтирониндеиодиназы 1—3: DIO1, DIO2Иодтирониндеиодиназы 1—3: DIO1, DIO2, DIO3 Глутанионпероксидазы: GPX1Глутанионпероксидазы:
- 66. IRES-элемент РНК вируса гепатита С- IRES (Internal Ribosome Entry Site) – очень своеобразный высокоструктурированный элемент вирусной
- 67. Последовательность событий при инициации трансляции РНК вируса гепатита С. Бинарный комплекс Рибосома, готовая трансли- ровать РНК
- 68. Вход мРНК Выход мРНК Участки связывания IRES и мРНК на рибосоме S3a
- 70. Выход синтезированного полипептида из рибосомы Сигнал-узнающая частица SRP распознает специальную сигнальную последовательность в синтезируемом полипептиде, как
- 71. Крио-электронное изображение комплекса транслирующей эукариотической рибосомы с SRP . Обозначены субчастицы, тРНК (tRNA), SRP и белок-рецептор
- 72. Трансмембранный канал для транслокации полипептида в ЭР Образован кольцеобразной олигомерной белковой структурой, состоящей из трех или
- 73. Сравнительная характеристика рибосом про- и эукариот Субчастицы: малая – 40S (1.4 мДа) большая - 60S (ок.
- 74. Рибосома высших организмов Рибосома бактерий 28S рРНК 3500-5000 нт, 5.8S рРНК, 5S рРНК, 47 белков 18S
- 75. Гомология между структурными элементами 70S и 80S рибосом Гомология между рРНК прокариот и эукариот Степень гомологии
- 76. Консервативные положения и сегменты экспансии во вторичной стуктуре рРНК, изображенные на структуре 16S E.coli. Большие кружки
- 77. Гомологичные рибосомные белки: Невысокая степень гомологии аминокислотных последовательностей и большое сходство пространственных структур гомологичных белков в
- 78. Качественные различия между рибосомами про- и эукариот Рибосомы эукариот невозможно собрать in vitro из набора рибосомных
- 79. Сборка субчастиц прокариот
- 81. Методы изучения строения рибосомы и ее функциональных центров Строение малой субчастицы рибосом Thermus thermophilus по данным
- 82. Электронная микроскопия Футпринтинг Аффинная модификация
- 83. Типы аналогов мРНК
- 84. Функциональные центры рибосомы и принципы их организации 1) участки связывания каждого из участников процесса трансляции (для
- 85. Строение ПТЦ у прокариот
- 86. Динамичность структуры рибосомы При узнавании правильного кодон-антикодонового дуплекса аа-тРНК с кодоном мРНК в А-участке малая субчастица
- 87. Алкалоид рицин расщепляет одну-единственную фосфодиэфирную связь в 23S рРНК в так называемой «сарцин-рициновой» петле (район GTPаза-активирующего
- 88. Рибосома и антибиотики Аминогликозиды узнают с высокой специфичностью структуру декодирующего центра 16S рРНК (по принципу «ключ-замок»),
- 89. Структуры аминогликозидных антибиотиков неомицина (А), канамицина (В) и стрептомицина (С)
- 90. Макролиды, содержащие 14-16-членное лактоновое кольцо, остатки сахара и боковые заместители, взаимодействуют с нуклеотидами 23S рРНК, образующими
- 92. Скачать презентацию
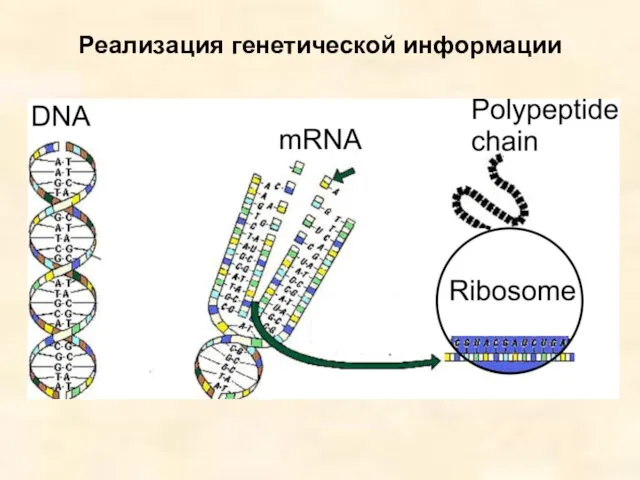
Реализация генетической информации
Реализация генетической информации

Генетический код
Генетическая информация – сумма признаков.
В основе - информация о строении
Генетический код
Генетическая информация – сумма признаков.
В основе - информация о строении

Генетический код – это запись только о последовательности аминокислот в белках.
Кроме
Генетический код – это запись только о последовательности аминокислот в белках.
Кроме

История
Уотсон-Крик 1953 – двойная спираль ДНК
1954 – Георгий Гамов. Предложил в
История
Уотсон-Крик 1953 – двойная спираль ДНК
1954 – Георгий Гамов. Предложил в

Свойства генетического кода:
Триплетность (почему именно – объяснить).
2) Однозначность (исключения – fMet/Met
Свойства генетического кода:
Триплетность (почему именно – объяснить).
2) Однозначность (исключения – fMet/Met

5) Компактность – отсутствие знаков препинания внутри генов, знаки препинания только
5) Компактность – отсутствие знаков препинания внутри генов, знаки препинания только
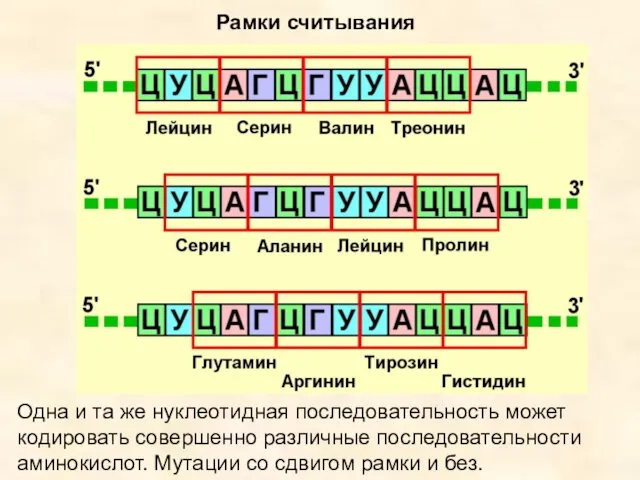
Рамки считывания
Одна и та же нуклеотидная последовательность может кодировать совершенно различные
Рамки считывания
Одна и та же нуклеотидная последовательность может кодировать совершенно различные
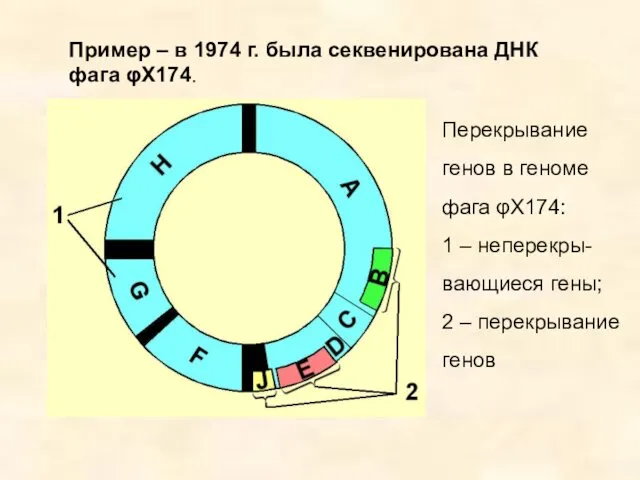
Пример – в 1974 г. была секвенирована ДНК фага φX174.
Перекрывание генов
Пример – в 1974 г. была секвенирована ДНК фага φX174.
Перекрывание генов

Что содержится в ДНК
Длина ДНК человека ок. 3.2 х 10 пар
Что содержится в ДНК
Длина ДНК человека ок. 3.2 х 10 пар
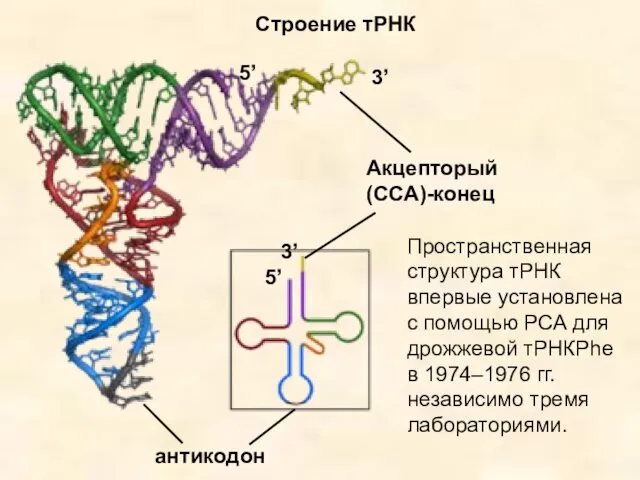
антикодон
Акцепторый
(ССА)-конец
Строение тРНК
3’
5’
3’
5’
Пространственная структура тРНК впервые установлена с помощью РСА для
антикодон
Акцепторый
(ССА)-конец
Строение тРНК
3’
5’
3’
5’
Пространственная структура тРНК впервые установлена с помощью РСА для
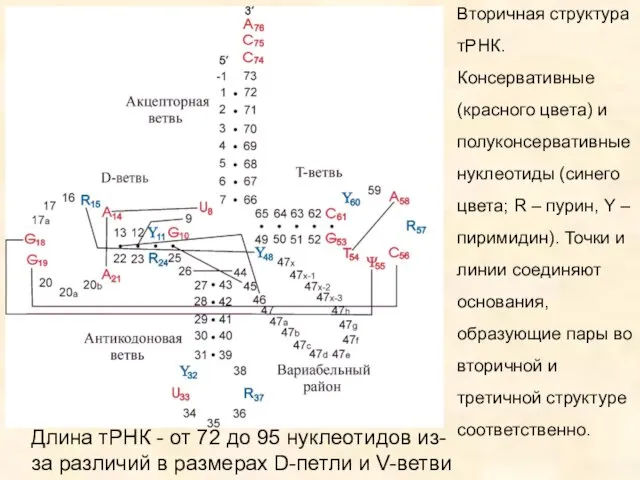
Вторичная структура тРНК. Консервативные (красного цвета) и полуконсервативные нуклеотиды (синего цвета;
Вторичная структура тРНК. Консервативные (красного цвета) и полуконсервативные нуклеотиды (синего цвета;
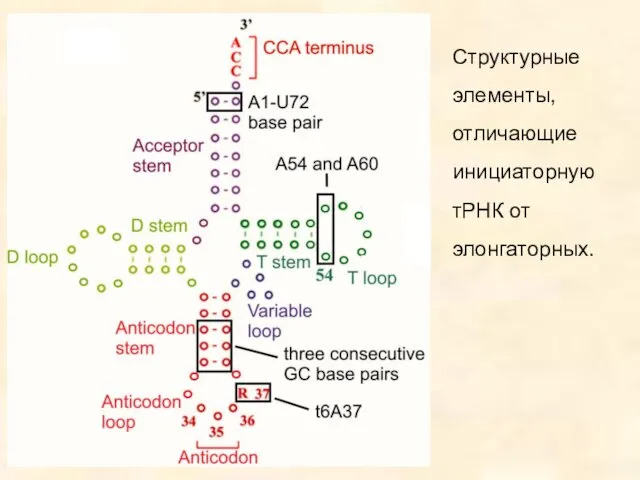
Структурные элементы, отличающие инициаторную тРНК от элонгаторных.
Структурные элементы, отличающие инициаторную тРНК от элонгаторных.

Особенностью тРНК является присутствие минорных нуклеозидов.
Структура митохондриальных тРНК млекопитающих сильно
Особенностью тРНК является присутствие минорных нуклеозидов.
Структура митохондриальных тРНК млекопитающих сильно
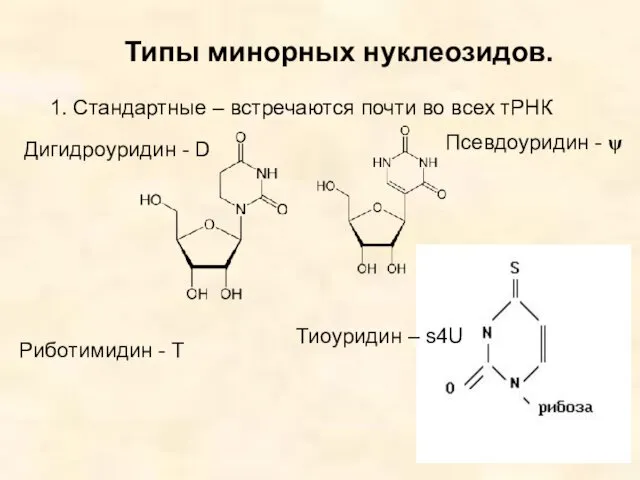
Типы минорных нуклеозидов.
1. Стандартные – встречаются почти во всех тРНК
Дигидроуридин
Типы минорных нуклеозидов.
1. Стандартные – встречаются почти во всех тРНК
Дигидроуридин
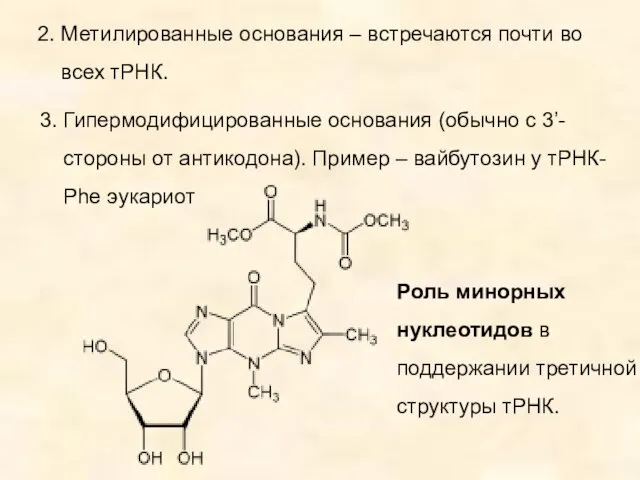
2. Метилированные основания – встречаются почти во
всех тРНК.
3. Гипермодифицированные основания
2. Метилированные основания – встречаются почти во
всех тРНК.
3. Гипермодифицированные основания

Рекогниция
Присоединение аминокислотного остатка к тРНК
Аминоацил-тРНК-синтетазы
Изоакцепторные тРНК
Рекогниция
Присоединение аминокислотного остатка к тРНК
Аминоацил-тРНК-синтетазы
Изоакцепторные тРНК

(1) aaRS + аа + АТР → aaRS•аа-АМР + РРi.
(2) aaRS•аа-АМР
(1) aaRS + аа + АТР → aaRS•аа-АМР + РРi.
(2) aaRS•аа-АМР

Нестандартные ситуации
1. Во многих бактериях и органеллах эукариот отсутствуют GlnRS и
Нестандартные ситуации
1. Во многих бактериях и органеллах эукариот отсутствуют GlnRS и
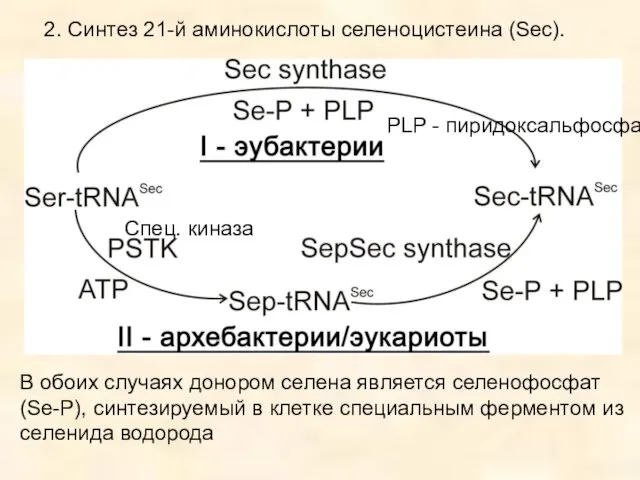
2. Синтез 21-й аминокислоты селеноцистеина (Sec).
PLP - пиридоксальфосфат
Спец. киназа
В обоих
2. Синтез 21-й аминокислоты селеноцистеина (Sec).
PLP - пиридоксальфосфат
Спец. киназа
В обоих
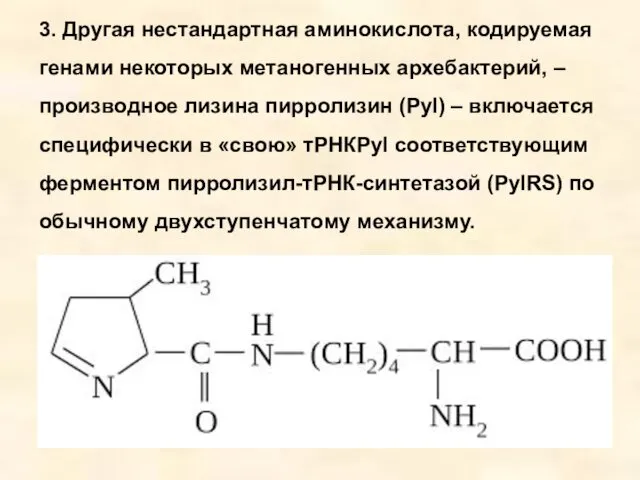
3. Другая нестандартная аминокислота, кодируемая генами некоторых метаногенных архебактерий, – производное
3. Другая нестандартная аминокислота, кодируемая генами некоторых метаногенных архебактерий, – производное
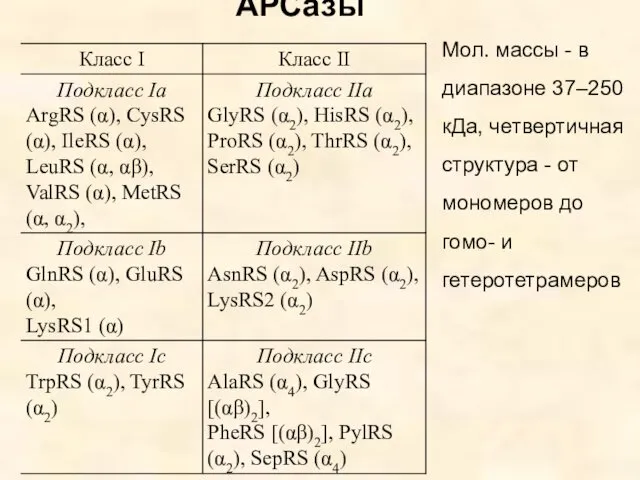
АРСазы
Мол. массы - в диапазоне 37–250 кДа, четвертичная структура - от
АРСазы
Мол. массы - в диапазоне 37–250 кДа, четвертичная структура - от

Ферменты разных классов отличаются устройством активного центра и способами связывания акцепторной
Ферменты разных классов отличаются устройством активного центра и способами связывания акцепторной
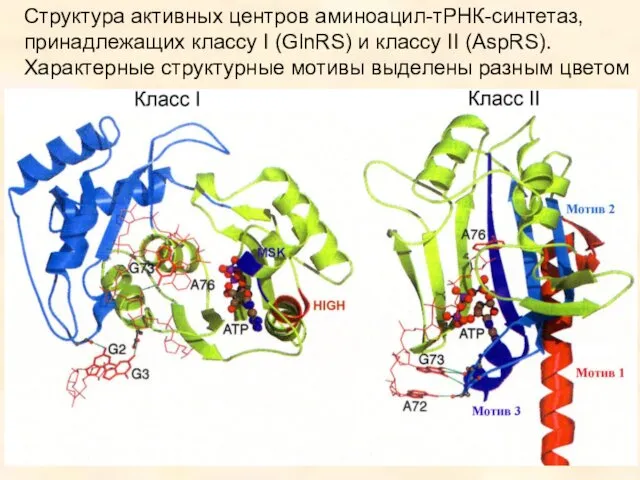
Структура активных центров аминоацил-тРНК-синтетаз, принадлежащих классу I (GlnRS) и классу II
Структура активных центров аминоацил-тРНК-синтетаз, принадлежащих классу I (GlnRS) и классу II

Для большинства синтетаз (за исключением 4-х ферментов класса I – ArgRS,
Для большинства синтетаз (за исключением 4-х ферментов класса I – ArgRS,

Узнавание тРНК АРСазами
Методы исследования - генетические in vivo и кинетических экспериментов
Узнавание тРНК АРСазами
Методы исследования - генетические in vivo и кинетических экспериментов
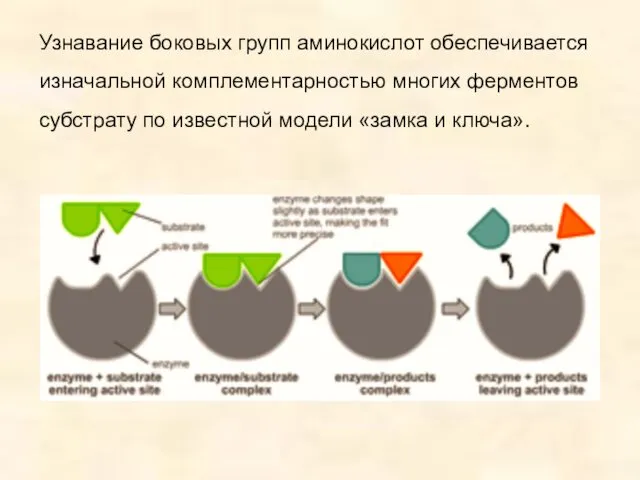
Узнавание боковых групп аминокислот обеспечивается изначальной комплементарностью многих ферментов субстрату по
Узнавание боковых групп аминокислот обеспечивается изначальной комплементарностью многих ферментов субстрату по
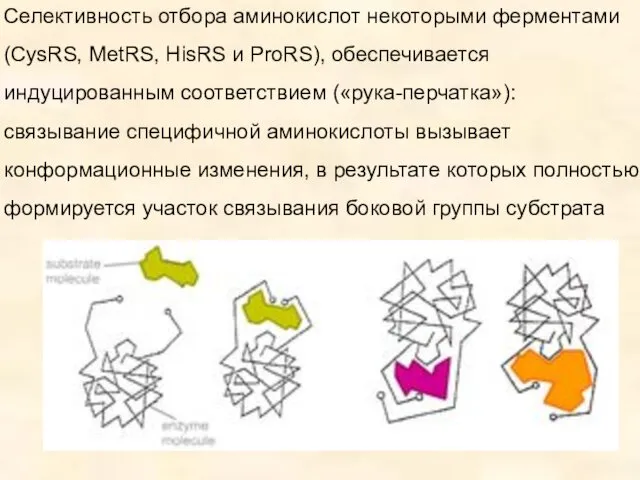
Селективность отбора аминокислот некоторыми ферментами (CysRS, MetRS, HisRS и ProRS), обеспечивается
Селективность отбора аминокислот некоторыми ферментами (CysRS, MetRS, HisRS и ProRS), обеспечивается

Установлены элементы узнавания тРНК для всех 20-ти синтетаз из E. сoli,
Установлены элементы узнавания тРНК для всех 20-ти синтетаз из E. сoli,
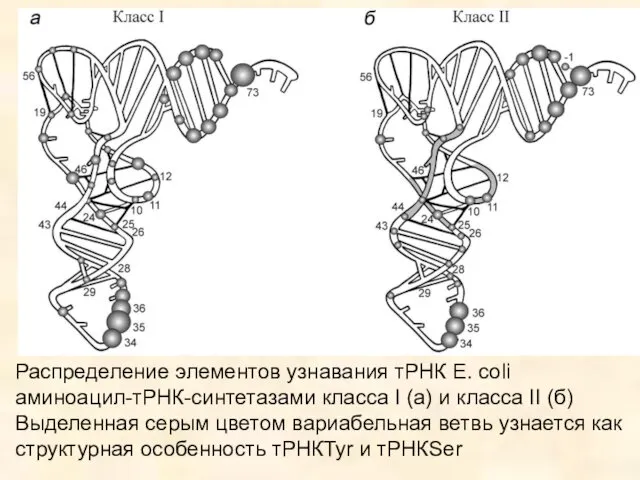
Распределение элементов узнавания тРНК E. coli аминоацил-тРНК-синтетазами класса I (а) и
Распределение элементов узнавания тРНК E. coli аминоацил-тРНК-синтетазами класса I (а) и

Антикодон не важен для узнавания тРНК E. сoli, специфичных к Leu,
Антикодон не важен для узнавания тРНК E. сoli, специфичных к Leu,
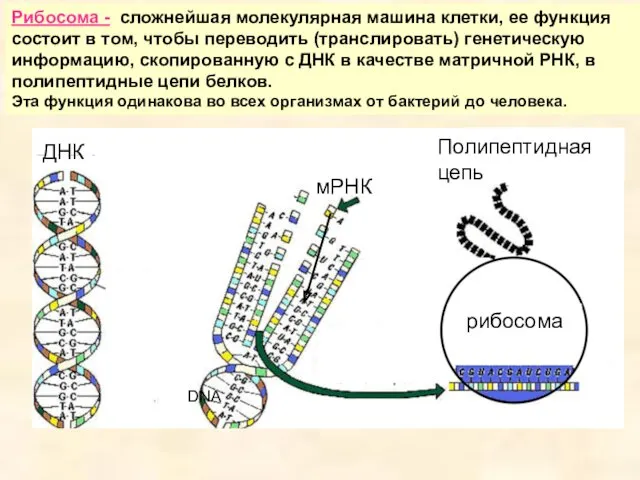
Рибосома - сложнейшая молекулярная машина клетки, ее функция состоит в том,
Рибосома - сложнейшая молекулярная машина клетки, ее функция состоит в том,
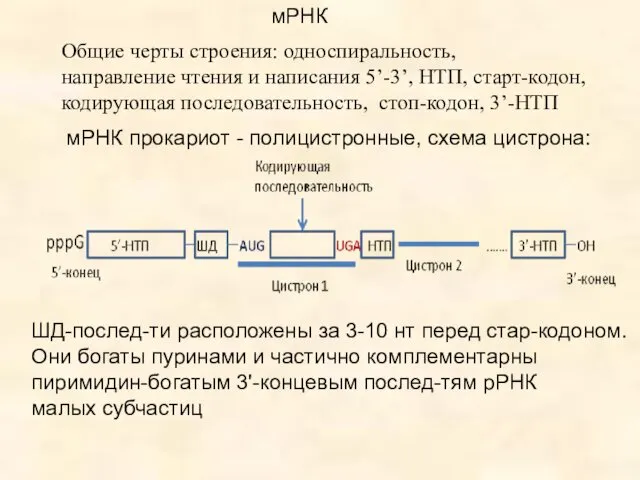
мРНК
Общие черты строения: односпиральность,
направление чтения и написания 5’-3’, НТП, старт-кодон,
кодирующая
мРНК
Общие черты строения: односпиральность,
направление чтения и написания 5’-3’, НТП, старт-кодон,
кодирующая

Особенности мРНК эукариот :
Одноцистронные
Нет ШД-послед-тей.
Кэп на 5’-конце.
Поли(А) – послед-ть на 3’-конце.
Полицистронными
Особенности мРНК эукариот :
Одноцистронные
Нет ШД-послед-тей.
Кэп на 5’-конце.
Поли(А) – послед-ть на 3’-конце.
Полицистронными
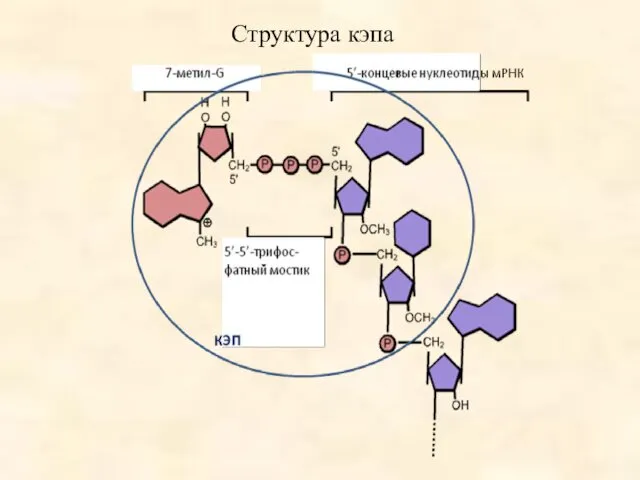
Структура кэпа
Структура кэпа

тРНК – сходство у всех организмов
Инициаторные тРНК аминоацилируются только остатками
тРНК – сходство у всех организмов
Инициаторные тРНК аминоацилируются только остатками

Факторы трансляции - одно- или многосубъединичные белки, подразделяющиеся на
факторы инициации
Факторы трансляции - одно- или многосубъединичные белки, подразделяющиеся на
факторы инициации

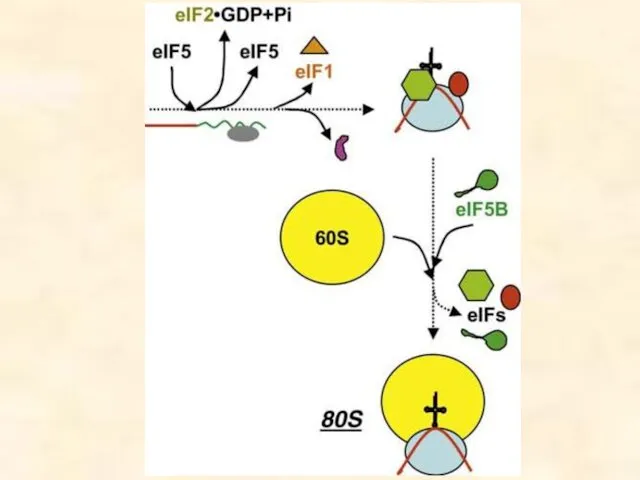
Инициация – процесс, приводящий к образованию комплекса, в котором старт-кодон AUG
Инициация – процесс, приводящий к образованию комплекса, в котором старт-кодон AUG
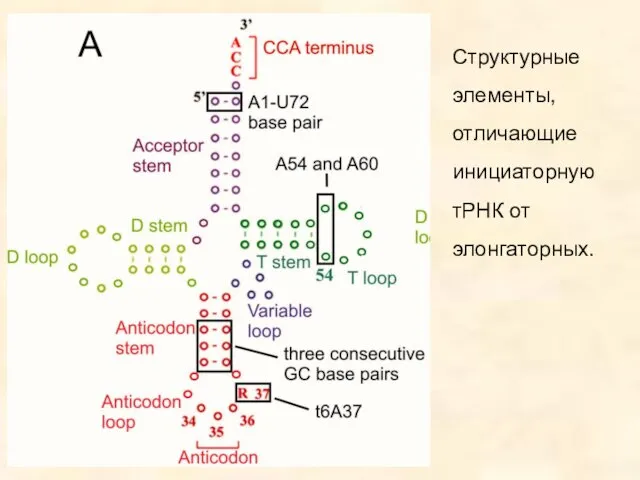
Структурные элементы, отличающие инициаторную тРНК от элонгаторных.
Структурные элементы, отличающие инициаторную тРНК от элонгаторных.
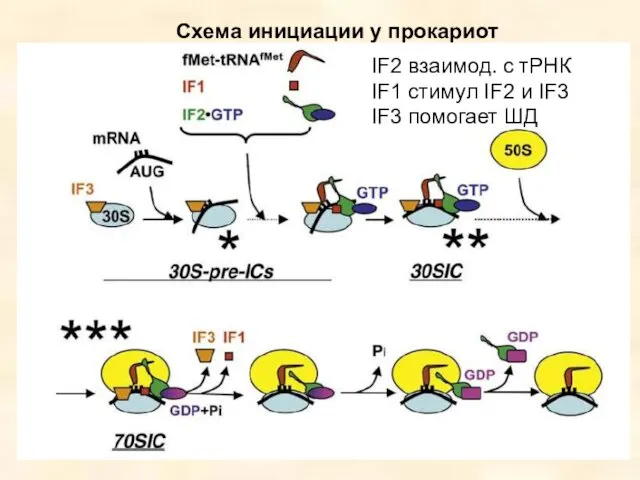
Схема инициации у прокариот
IF2 взаимод. с тРНК
IF1 стимул IF2 и IF3
IF3
Схема инициации у прокариот
IF2 взаимод. с тРНК
IF1 стимул IF2 и IF3
IF3
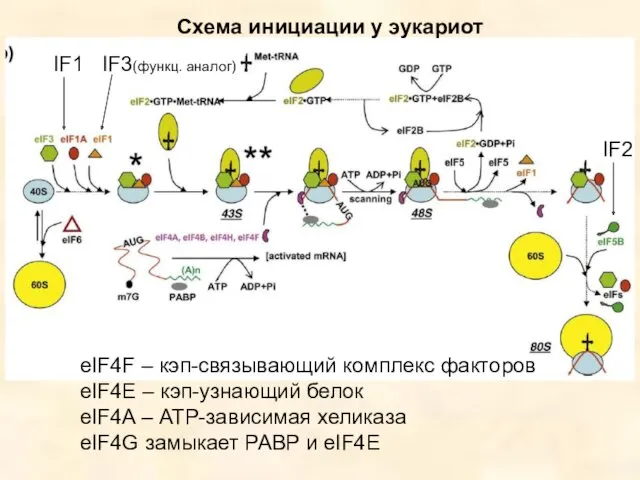
Схема инициации у эукариот
IF1
IF3(функц. аналог)
IF2
eIF4F – кэп-связывающий комплекс факторов
eIF4E – кэп-узнающий
Схема инициации у эукариот
IF1
IF3(функц. аналог)
IF2
eIF4F – кэп-связывающий комплекс факторов
eIF4E – кэп-узнающий
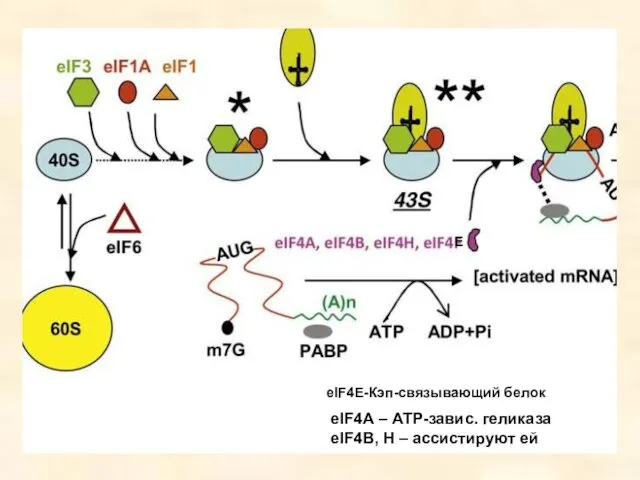
eIF4A – АТР-завис. геликаза
eIF4B, Н – ассистируют ей
E
eIF4E-Кэп-связывающий белок
eIF4A – АТР-завис. геликаза
eIF4B, Н – ассистируют ей
E
eIF4E-Кэп-связывающий белок
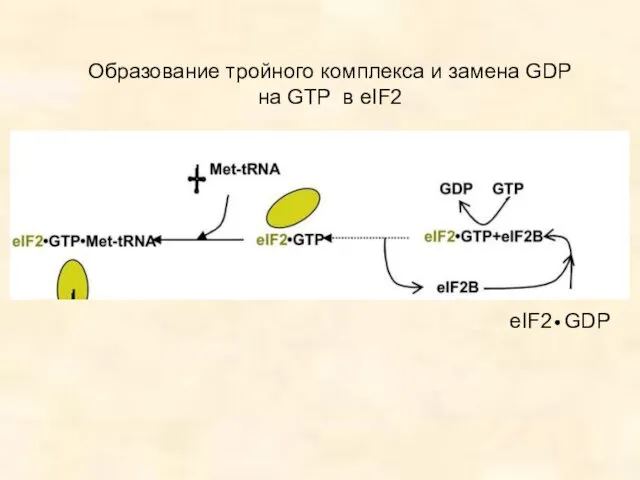
Образование тройного комплекса и замена GDP на GTP в eIF2
eIF2 GDP
Образование тройного комплекса и замена GDP на GTP в eIF2
eIF2 GDP
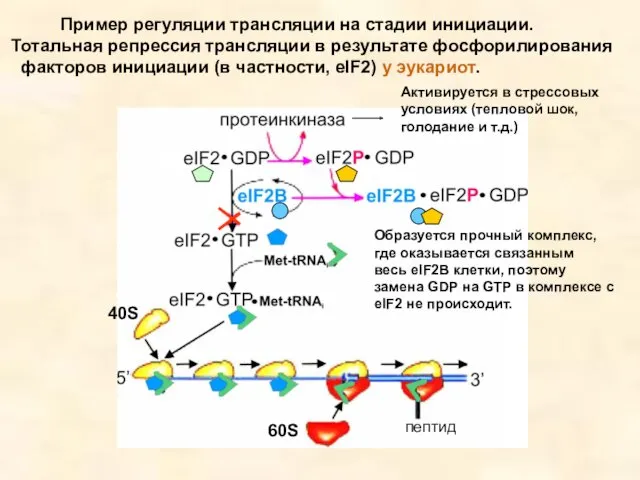
Активируется в стрессовых
условиях (тепловой шок,
голодание и т.д.)
Образуется прочный комплекс,
где
Активируется в стрессовых
условиях (тепловой шок,
голодание и т.д.)
Образуется прочный комплекс,
где
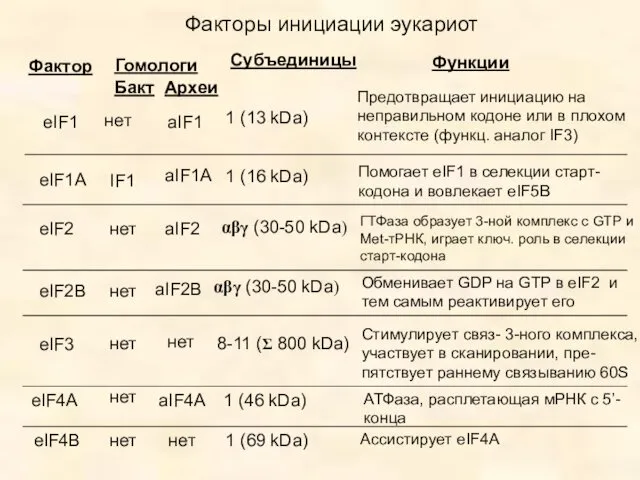
Факторы инициации эукариот
Фактор
Гомологи
Бакт Археи
Субъединицы
Функции
eIF1
1 (13 kDa)
Предотвращает инициацию на неправильном кодоне или
Факторы инициации эукариот
Фактор
Гомологи
Бакт Археи
Субъединицы
Функции
eIF1
1 (13 kDa)
Предотвращает инициацию на неправильном кодоне или

Фактор
Субъединицы
Функции
eIF4E
нет
1 (25 kDa)
Связывается с кэпом
eIF4F
нет
Комплекс
eIF4E, A и G
Расплетает 5’-концевую часть
Фактор
Субъединицы
Функции
eIF4E
нет
1 (25 kDa)
Связывается с кэпом
eIF4F
нет
Комплекс
eIF4E, A и G
Расплетает 5’-концевую часть

Цикл элонгации.
eEF1 – EF-Tu
eEF2 – EF-G
Аналогичны по функции, но
не работают в
Цикл элонгации.
eEF1 – EF-Tu
eEF2 – EF-G
Аналогичны по функции, но
не работают в
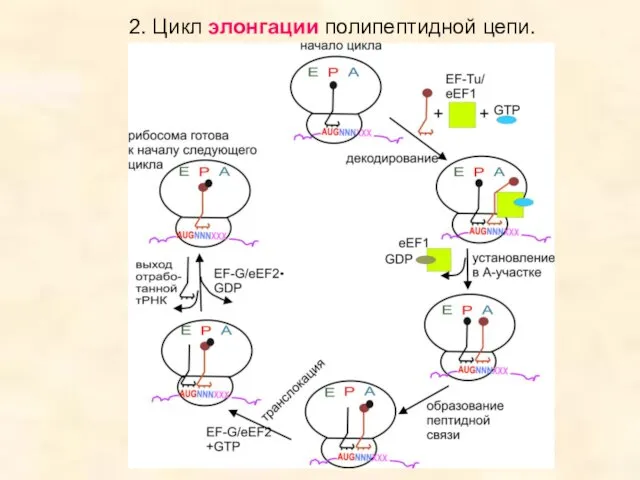
2. Цикл элонгации полипептидной цепи.
2. Цикл элонгации полипептидной цепи.
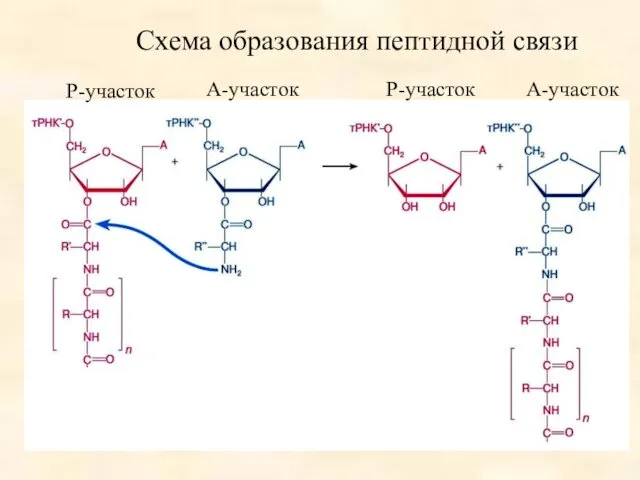
Р-участок
А-участок
Р-участок
А-участок
Схема образования пептидной связи
Р-участок
А-участок
Р-участок
А-участок
Схема образования пептидной связи

3. Терминация трансляции наступает, когда в аминоацильном
(А)-участке оказывается один из 3-х
3. Терминация трансляции наступает, когда в аминоацильном
(А)-участке оказывается один из 3-х

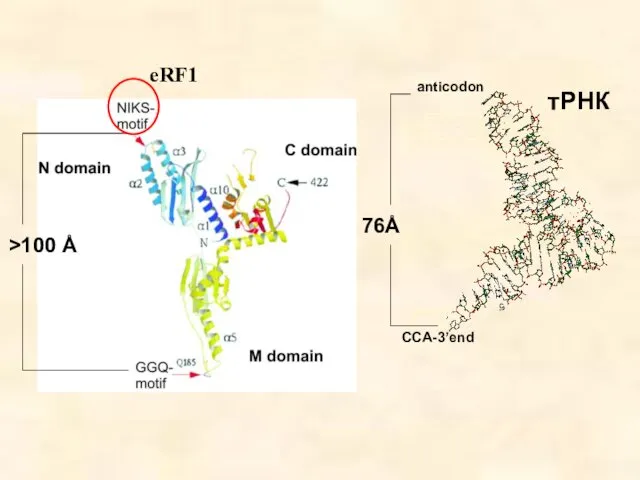
Молекулярная мимикрия – сходство структуры RF и тРНК. Связываясь с рибосомой,
Молекулярная мимикрия – сходство структуры RF и тРНК. Связываясь с рибосомой,
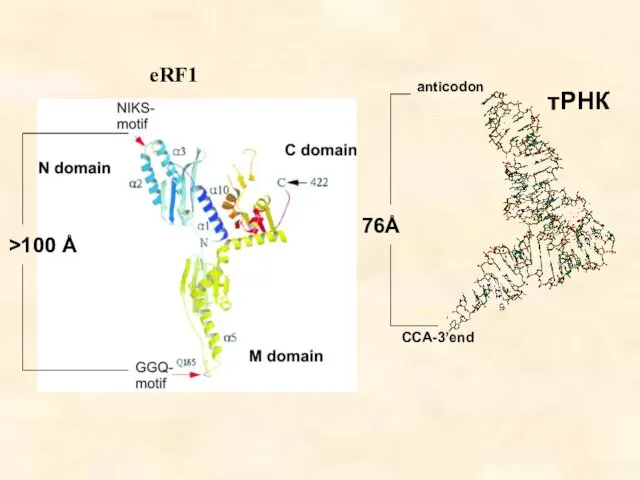
>100 Å
anticodon
CCA-3’end
76Å
тРНК
eRF1
>100 Å
anticodon
CCA-3’end
76Å
тРНК
eRF1
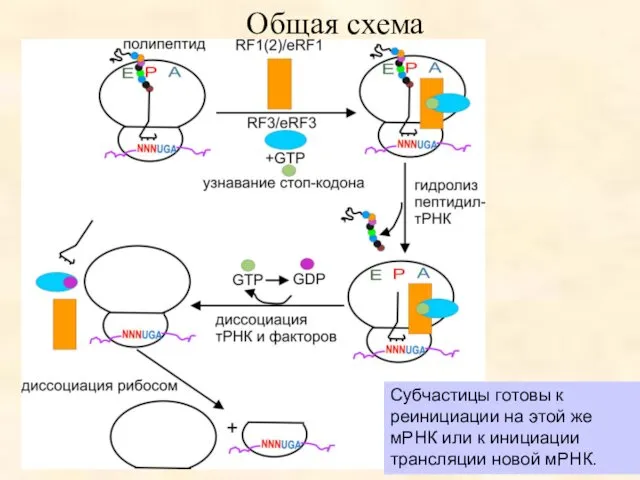
Субчастицы готовы к
реинициации на этой же
мРНК или к инициации
Субчастицы готовы к
реинициации на этой же
мРНК или к инициации

Характерные особенности терминации у прокариот:
1) освобождение синтезированного полипептида происходит до гидролиза
Характерные особенности терминации у прокариот:
1) освобождение синтезированного полипептида происходит до гидролиза

У эукариот
оба фактора терминации действуют кооперативно и скоодинированно:
1) eRF1 имеет
У эукариот
оба фактора терминации действуют кооперативно и скоодинированно:
1) eRF1 имеет

Гидролиз GTP приводит к изменению конформации терминационного комплекса таким образом, что
Гидролиз GTP приводит к изменению конформации терминационного комплекса таким образом, что

Рециклинг - диссоциация мРНК и деацилированной тРНК и последующая диссоциация рибосом
Рециклинг - диссоциация мРНК и деацилированной тРНК и последующая диссоциация рибосом

У эукариот и архей специализированного фактора рециклинга нет.
Диссоциация 80S рибосом
У эукариот и архей специализированного фактора рециклинга нет.
Диссоциация 80S рибосом

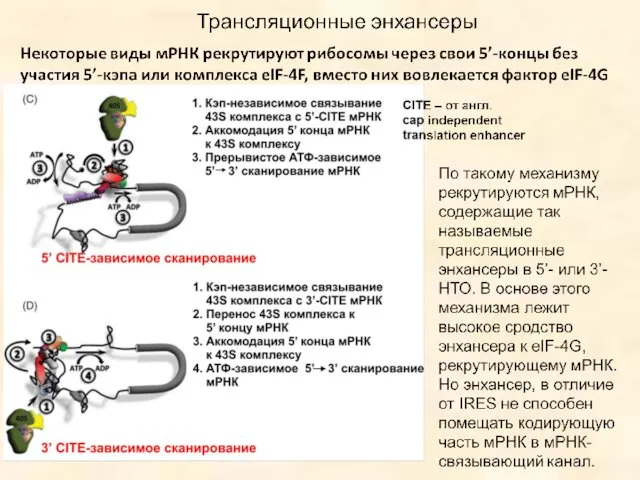
Отклонения от канонических правил
Неканоническая инициация – трансляционные энхансеры в мРНК и
Отклонения от канонических правил
Неканоническая инициация – трансляционные энхансеры в мРНК и
>100 Å
anticodon
CCA-3’end
76Å
тРНК
eRF1
>100 Å
anticodon
CCA-3’end
76Å
тРНК
eRF1
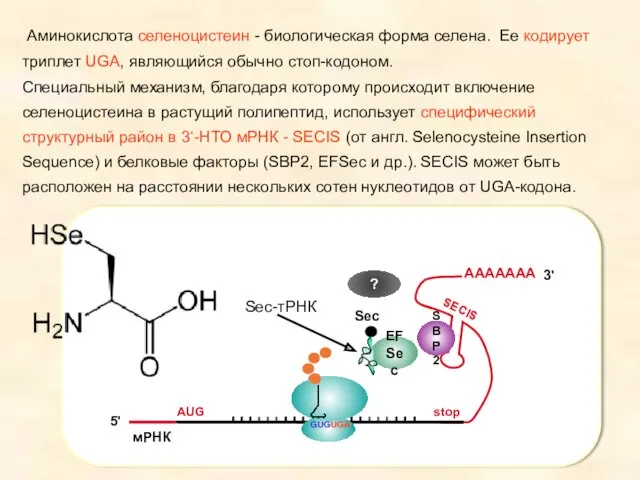
Аминокислота селеноцистеин - биологическая форма селена. Ее кодирует триплет UGA,
Аминокислота селеноцистеин - биологическая форма селена. Ее кодирует триплет UGA,

К человеческим селенопротеинам относят:
Иодтирониндеиодиназы 1—3: DIO1Иодтирониндеиодиназы 1—3: DIO1, DIO2Иодтирониндеиодиназы 1—3: DIO1, DIO2, DIO3
Глутанионпероксидазы: GPX1Глутанионпероксидазы: GPX1, GPX2Глутанионпероксидазы: GPX1, GPX2, GPX3Глутанионпероксидазы: GPX1, GPX2, GPX3, GPX4Глутанионпероксидазы: GPX1, GPX2, GPX3, GPX4, GPX6[4]
Селенопротеины: SelH, SelI, SelK, SelM,
К человеческим селенопротеинам относят:
Иодтирониндеиодиназы 1—3: DIO1Иодтирониндеиодиназы 1—3: DIO1, DIO2Иодтирониндеиодиназы 1—3: DIO1, DIO2, DIO3
Глутанионпероксидазы: GPX1Глутанионпероксидазы: GPX1, GPX2Глутанионпероксидазы: GPX1, GPX2, GPX3Глутанионпероксидазы: GPX1, GPX2, GPX3, GPX4Глутанионпероксидазы: GPX1, GPX2, GPX3, GPX4, GPX6[4]
Селенопротеины: SelH, SelI, SelK, SelM,
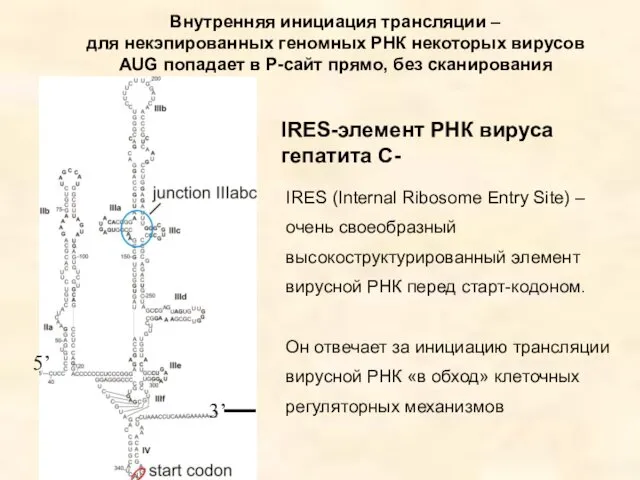
IRES-элемент РНК вируса гепатита С-
IRES (Internal Ribosome Entry Site) –
IRES-элемент РНК вируса гепатита С-
IRES (Internal Ribosome Entry Site) –
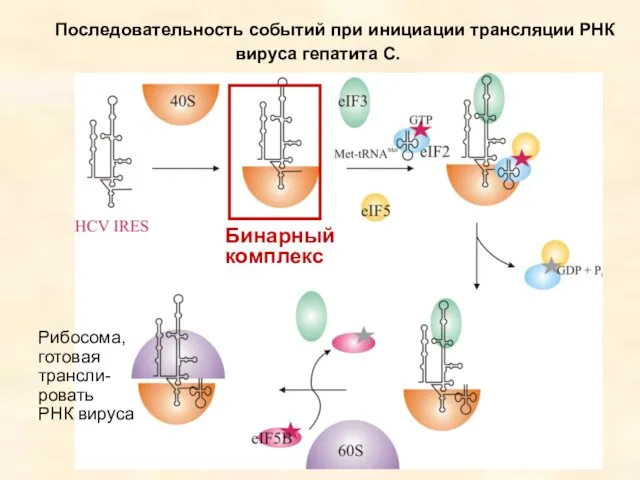
Последовательность событий при инициации трансляции РНК вируса гепатита С.
Бинарный
Последовательность событий при инициации трансляции РНК вируса гепатита С.
Бинарный
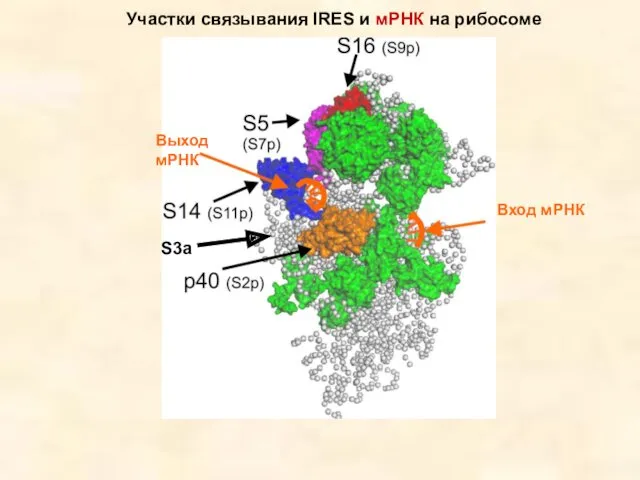
Вход мРНК
Выход
мРНК
Участки связывания IRES и мРНК на рибосоме
S3a
Вход мРНК
Выход
мРНК
Участки связывания IRES и мРНК на рибосоме
S3a

Выход синтезированного полипептида из рибосомы
Сигнал-узнающая частица SRP распознает специальную сигнальную последовательность
Выход синтезированного полипептида из рибосомы
Сигнал-узнающая частица SRP распознает специальную сигнальную последовательность
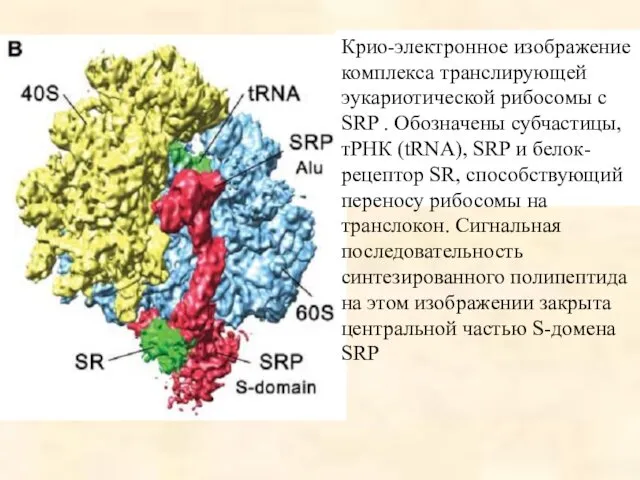
Крио-электронное изображение комплекса транслирующей эукариотической рибосомы с SRP . Обозначены субчастицы,
Крио-электронное изображение комплекса транслирующей эукариотической рибосомы с SRP . Обозначены субчастицы,

Трансмембранный канал для транслокации полипептида в ЭР
Образован кольцеобразной олигомерной белковой структурой,
Трансмембранный канал для транслокации полипептида в ЭР
Образован кольцеобразной олигомерной белковой структурой,

Сравнительная характеристика рибосом про- и эукариот
Субчастицы: малая – 40S (1.4 мДа)
Сравнительная характеристика рибосом про- и эукариот
Субчастицы: малая – 40S (1.4 мДа)
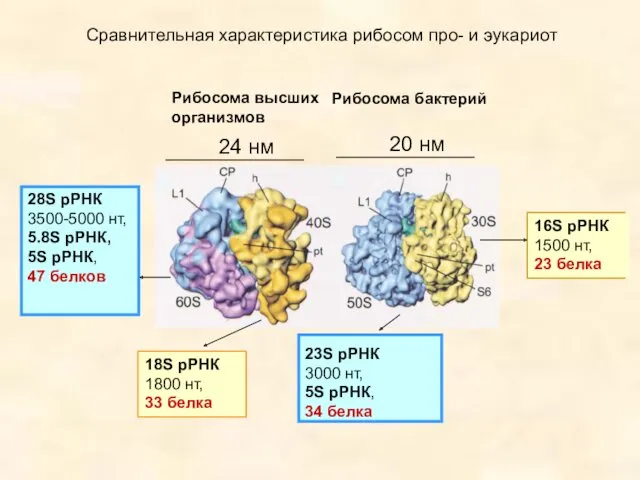
Рибосома высших
организмов
Рибосома бактерий
28S рРНК 3500-5000 нт, 5.8S рРНК,
5S рРНК,
47
Рибосома высших
организмов
Рибосома бактерий
28S рРНК 3500-5000 нт, 5.8S рРНК,
5S рРНК,
47

Гомология между структурными элементами
70S и 80S рибосом
Гомология между рРНК прокариот
Гомология между структурными элементами
70S и 80S рибосом
Гомология между рРНК прокариот
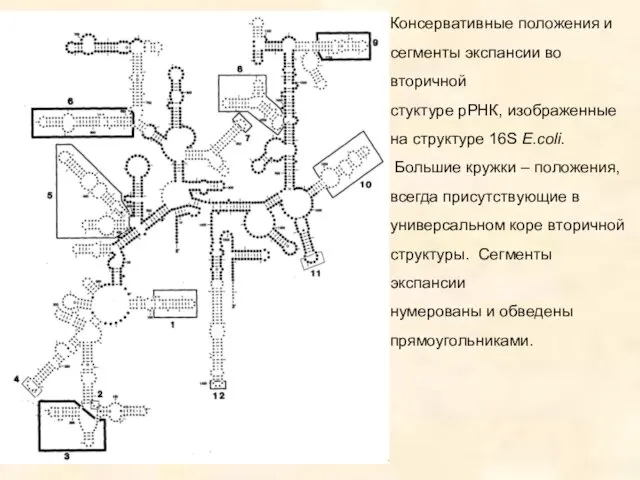
Консервативные положения и
сегменты экспансии во вторичной
стуктуре рРНК, изображенные
на
Консервативные положения и
сегменты экспансии во вторичной
стуктуре рРНК, изображенные
на

Гомологичные рибосомные белки:
Невысокая степень гомологии аминокислотных
последовательностей и
большое сходство пространственных структур
гомологичных
Гомологичные рибосомные белки:
Невысокая степень гомологии аминокислотных
последовательностей и
большое сходство пространственных структур
гомологичных

Качественные различия между рибосомами
про- и эукариот
Рибосомы эукариот невозможно собрать in
Качественные различия между рибосомами
про- и эукариот
Рибосомы эукариот невозможно собрать in
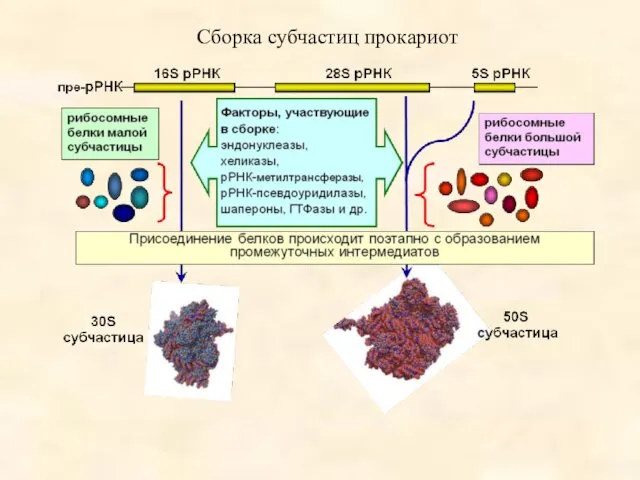
Сборка субчастиц прокариот
Сборка субчастиц прокариот
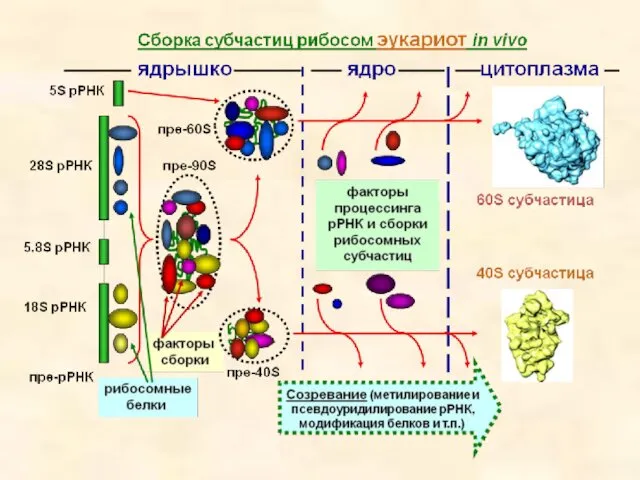
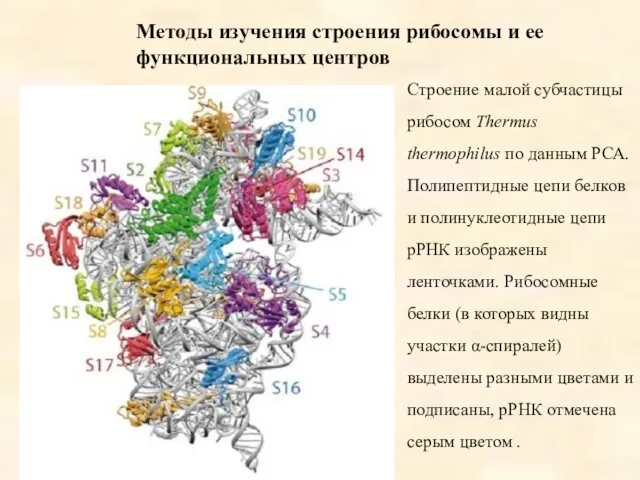
Методы изучения строения рибосомы и ее функциональных центров
Строение малой субчастицы рибосом
Методы изучения строения рибосомы и ее функциональных центров
Строение малой субчастицы рибосом
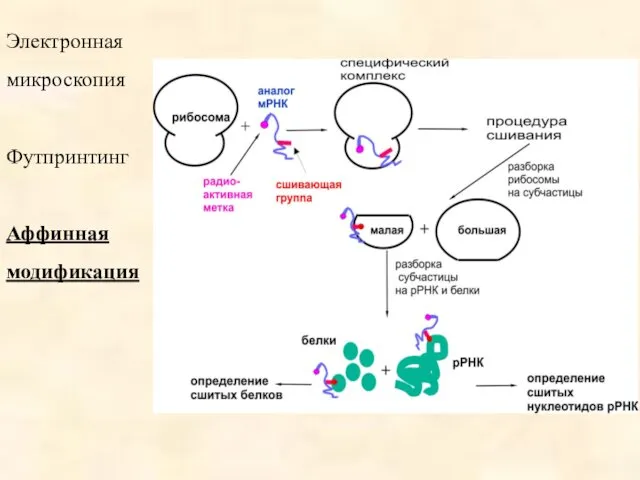
Электронная микроскопия
Футпринтинг
Аффинная
модификация
Электронная микроскопия
Футпринтинг
Аффинная
модификация

Типы аналогов мРНК
Типы аналогов мРНК

Функциональные центры рибосомы и принципы их организации
1) участки связывания каждого из
Функциональные центры рибосомы и принципы их организации
1) участки связывания каждого из
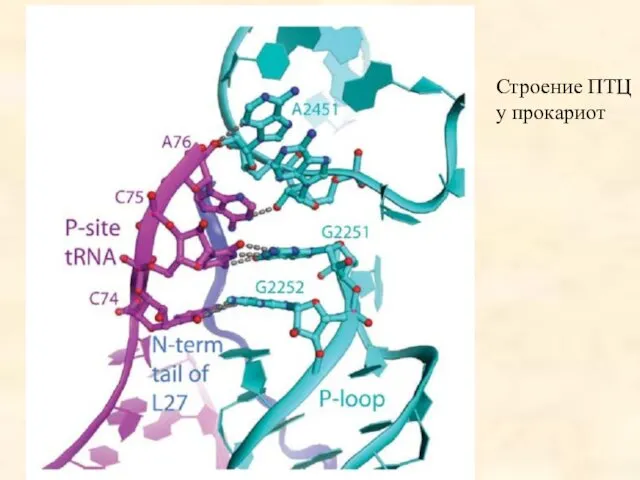
Строение ПТЦ
у прокариот
Строение ПТЦ
у прокариот

Динамичность структуры рибосомы
При узнавании правильного кодон-антикодонового дуплекса аа-тРНК с кодоном мРНК
Динамичность структуры рибосомы
При узнавании правильного кодон-антикодонового дуплекса аа-тРНК с кодоном мРНК

Алкалоид рицин расщепляет одну-единственную фосфодиэфирную связь в 23S рРНК в так
Алкалоид рицин расщепляет одну-единственную фосфодиэфирную связь в 23S рРНК в так

Рибосома и антибиотики
Аминогликозиды узнают с высокой специфичностью структуру декодирующего центра 16S
Рибосома и антибиотики
Аминогликозиды узнают с высокой специфичностью структуру декодирующего центра 16S
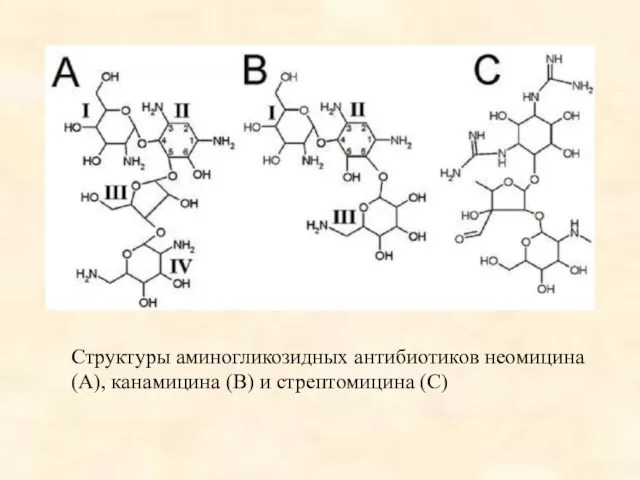
Структуры аминогликозидных антибиотиков неомицина (А), канамицина (В) и стрептомицина (С)
Структуры аминогликозидных антибиотиков неомицина (А), канамицина (В) и стрептомицина (С)
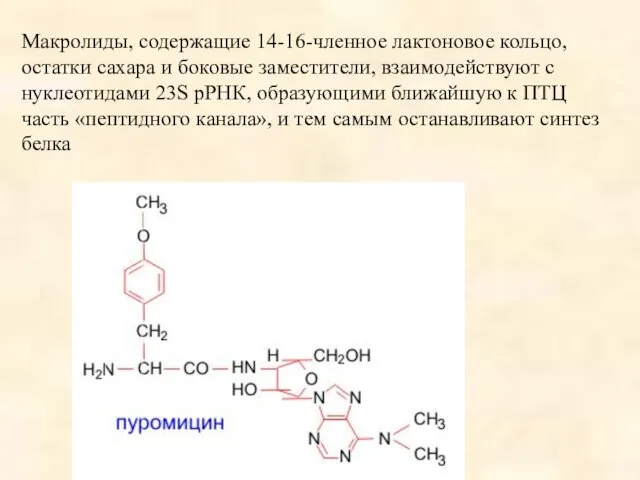
Макролиды, содержащие 14-16-членное лактоновое кольцо, остатки сахара и боковые заместители, взаимодействуют
Макролиды, содержащие 14-16-членное лактоновое кольцо, остатки сахара и боковые заместители, взаимодействуют
 Формирование и совершенствование имиджа менеджера организации ИП Гайсина Ю.Ф
Формирование и совершенствование имиджа менеджера организации ИП Гайсина Ю.Ф Презентация к уроку технология, изделие Торт
Презентация к уроку технология, изделие Торт Сталинградская битва
Сталинградская битва Пищеварение в ротовой полости
Пищеварение в ротовой полости Litania do Ducha Świętego
Litania do Ducha Świętego Деревья
Деревья Пять уроков творчества
Пять уроков творчества Я люблю свой город
Я люблю свой город Класс Земноводные, или амфибии
Класс Земноводные, или амфибии Формы работы с родителями учащихся начальных классов
Формы работы с родителями учащихся начальных классов Фитопатогенные грибы
Фитопатогенные грибы Интерактивный тренажёр. Буквы потерялись. Русский язык, 4 класс
Интерактивный тренажёр. Буквы потерялись. Русский язык, 4 класс Презентация Моя инициатива в образовании.
Презентация Моя инициатива в образовании. Источники противопожарного водоснабжения
Источники противопожарного водоснабжения Интерактивные валюты
Интерактивные валюты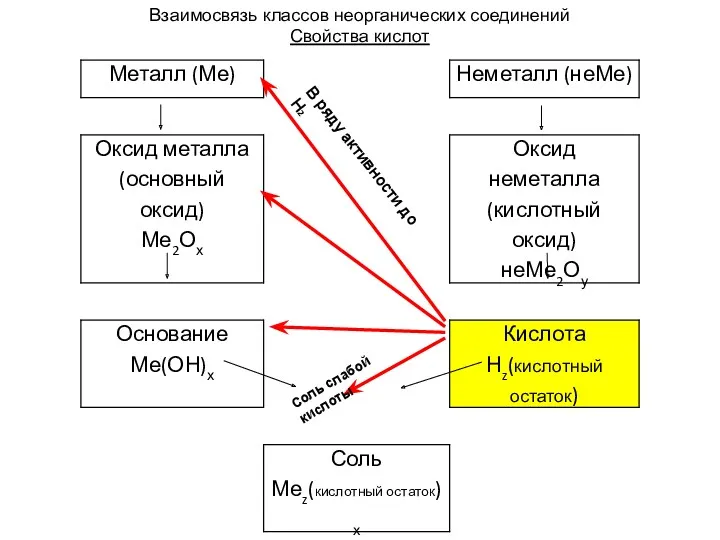 8 класс тема 6 Электролитическая диссоциация
8 класс тема 6 Электролитическая диссоциация Низкопотенциальная геотермальная энергия (тепловые насосы) в энергоэффективном проектировании
Низкопотенциальная геотермальная энергия (тепловые насосы) в энергоэффективном проектировании Мониторинг и прогнозирование чрезвычайных ситуаций
Мониторинг и прогнозирование чрезвычайных ситуаций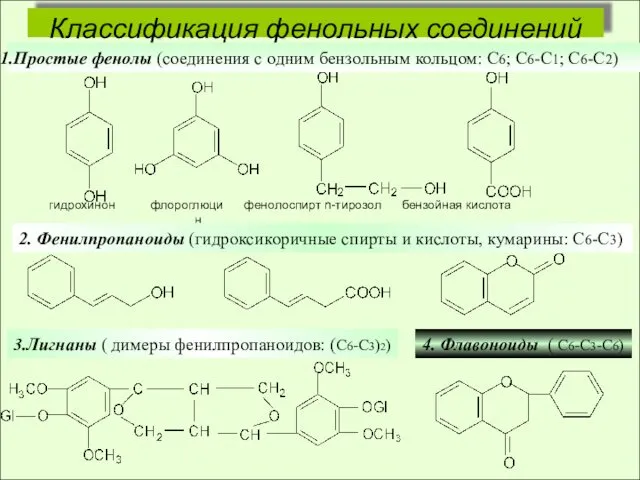 Классификация фенольных соединений
Классификация фенольных соединений Меры безопасности при пользовании предметами бытовой химии
Меры безопасности при пользовании предметами бытовой химии Снятие мерок для построения чертежа фартука
Снятие мерок для построения чертежа фартука Моя Югра
Моя Югра ТВОРЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ОБЪЕДИНЕНИЯ ДЕКОРАТИВНО-ПРИКЛАДНОЕ ТВОРЧЕСТВО
ТВОРЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ ОБЪЕДИНЕНИЯ ДЕКОРАТИВНО-ПРИКЛАДНОЕ ТВОРЧЕСТВО Занятие по экологии в группе продленного дня
Занятие по экологии в группе продленного дня Жүре пайда болған кемақылдық деменция
Жүре пайда болған кемақылдық деменция Водоросли. Многообразие водорослей
Водоросли. Многообразие водорослей ЭОСВ
ЭОСВ Автоматизация звука Ш
Автоматизация звука Ш