- Дипловертеброн

Содержание
- 2. Сеймуриоморфы Сеймурия 303 - 295 млн л н появилась група сеймуриоморф . Их считают промежуточным звеном
- 3. Схема происхождения и эволюции амниот
- 4. Схема родства современных и вымерших амниот
- 5. Эволюция типов черепа амниот
- 6. Котилозавры Котилоза́вры (цельноголовые) - группа вымерших рептилий из подкласса Анапсид. Существовали с середины каменноугольного периода (320
- 7. Анапсиды хилономус одонтохелис - самая древняя протостега архелон - самый крупный эвнотозавр - предок черепах анапсидный
- 8. Диапсиды диапсидный (двухдужный) череп Эозухии Череп имеет две височные впадины, расположенные выше и ниже заглазничной кости.
- 9. Современные Лепидозавры Клювоголовые Чешуйчатые ящерицы змеи гаттерия = туатара
- 10. Текодонты или Псевдозухии Текодонты (ячеистозубые) появились в начале триаса, достигли расцвета в юре и мелу. Имели
- 11. Птицетазовые динозавры Стегозавр Игуанодон Птицетазовые - один из двух отрядов динозавров. Несмотря на своё название, не
- 12. Ящеротазовые динозавры Компсогнатус Бронтозавр Коритозавр Цератозавр и стегозавры Ящерота́зовые - один из двух главных отрядов динозавров.
- 13. Архозавры (высшие ящеры) Большинство архозавров были хищниками, но·растительноядные тоже встречались среди ящеротазовых динозавров (зауроподов, прозавроподов, теризинозавров),птицетазовых
- 14. Пеликозавры Для них характерны отверстия в щёчных костях (боковые височные ямы с двух сторон черепа), служащие
- 15. Териодонты (зверозубые) Циногнатус Териодонты или зверозубые ящеры - вымершая группа тероморф, одна из трёх главных групп
- 16. Терапсиды = зверодужные Пристероогнатус Потомки цинодонтов. Отличались большим разнообразием, некоторые были высокоспе- циализированы. Теменного отверстия нет.
- 17. Покровы наземных позвоночных строение кожи рептилий (с чешуйкой) строение кожи млекопитающих (с волосом) Особенности У рептилий
- 18. чешуя змеи хвост бобра щитки птиц Производные кожи амниот перо шерсть когти птиц когти зверей копыта
- 19. Расположение перьев на теле птицы 1- птерилии (оперённые участки), 2- аптерии (голые участки)
- 20. чехол клюва птиц чехлы рогов зверей китовый ус Производные накладных костей панцирь черепах панцирь броненосца рога
- 21. Водно-солевой обмен и осморегуляция у амниот У рептилий, птиц и млекопитающих сначала образуются нефункционирующие канальцы пронефроса,
- 22. Типы белкового обмена: рептилии, птицы урикотелия - выделение кристаллической мочевой кислоты, не требую-щей воды для растворения,
- 23. Стропние нефрона маммального типа (как у млекопитающих) 1- боуменова капсула, 2- клубочек, 3- проксимальный извитой каналец.
- 24. Почки амниот образуют корковый и мозговой слои. У рептилий нефроны не имеют петель Генле. В почках
- 26. Скачать презентацию

Сеймуриоморфы
Сеймурия
303 - 295 млн л н появилась група
сеймуриоморф .
Их считают
Сеймуриоморфы
Сеймурия
303 - 295 млн л н появилась група
сеймуриоморф .
Их считают
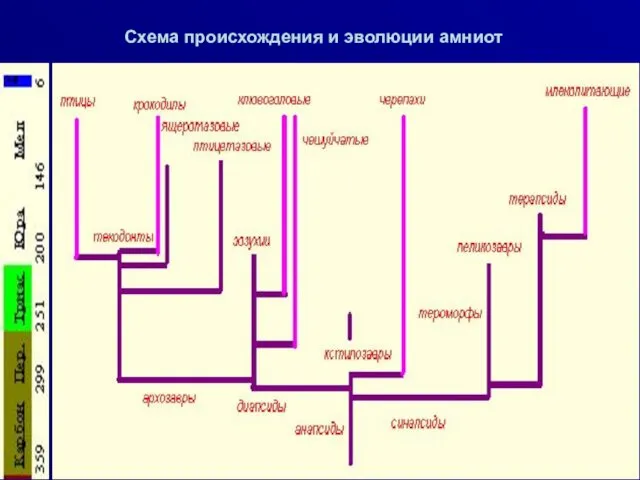
Схема происхождения и эволюции амниот
Схема происхождения и эволюции амниот
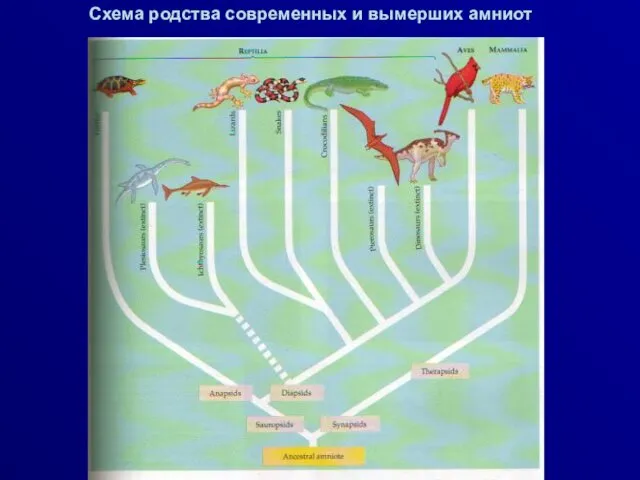
Схема родства современных и вымерших амниот
Схема родства современных и вымерших амниот
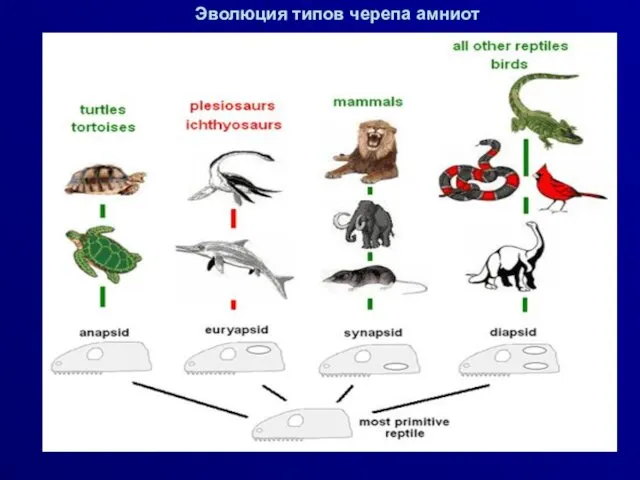
Эволюция типов черепа амниот
Эволюция типов черепа амниот

Котилозавры
Котилоза́вры (цельноголовые) - группа вымерших рептилий из подкласса Анапсид.
Существовали с середины
Котилозавры
Котилоза́вры (цельноголовые) - группа вымерших рептилий из подкласса Анапсид.
Существовали с середины

Анапсиды
хилономус
одонтохелис -
самая древняя
протостега
архелон -
самый крупный
эвнотозавр -
предок черепах
анапсидный (бездужный)
череп
Эвнотозавры имели широкие
Анапсиды
хилономус
одонтохелис -
самая древняя
протостега
архелон -
самый крупный
эвнотозавр -
предок черепах
анапсидный (бездужный)
череп
Эвнотозавры имели широкие

Диапсиды
диапсидный (двухдужный)
череп
Эозухии
Череп имеет две височные впадины, расположенные выше и ниже
Диапсиды
диапсидный (двухдужный)
череп
Эозухии
Череп имеет две височные впадины, расположенные выше и ниже

Современные Лепидозавры
Клювоголовые
Чешуйчатые
ящерицы
змеи
гаттерия = туатара
Современные Лепидозавры
Клювоголовые
Чешуйчатые
ящерицы
змеи
гаттерия = туатара

Текодонты или Псевдозухии
Текодонты (ячеистозубые) появились в начале триаса, достигли расцвета в
Текодонты или Псевдозухии
Текодонты (ячеистозубые) появились в начале триаса, достигли расцвета в

Птицетазовые динозавры
Стегозавр
Игуанодон
Птицетазовые - один из двух отрядов динозавров. Несмотря
Птицетазовые динозавры
Стегозавр
Игуанодон
Птицетазовые - один из двух отрядов динозавров. Несмотря

Ящеротазовые динозавры
Компсогнатус
Бронтозавр
Коритозавр
Цератозавр и стегозавры
Ящерота́зовые - один из двух главных отрядов
Ящеротазовые динозавры
Компсогнатус
Бронтозавр
Коритозавр
Цератозавр и стегозавры
Ящерота́зовые - один из двух главных отрядов
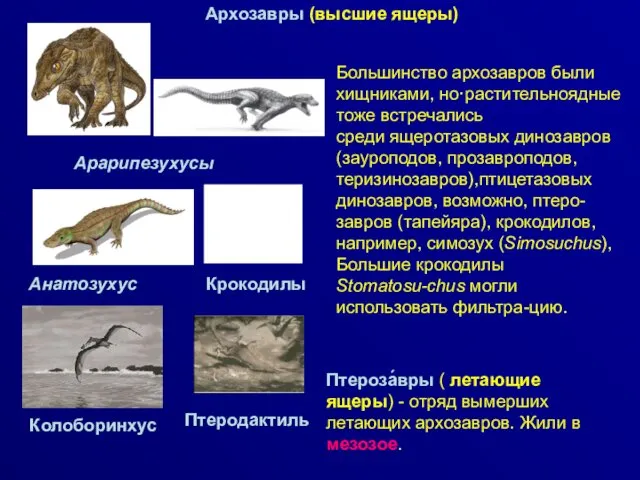
Архозавры (высшие ящеры)
Большинство архозавров были хищниками, но·растительноядные тоже встречались
среди ящеротазовых динозавров
Архозавры (высшие ящеры)
Большинство архозавров были хищниками, но·растительноядные тоже встречались
среди ящеротазовых динозавров

Пеликозавры
Для них характерны отверстия в щёчных костях (боковые височные ямы с
Пеликозавры
Для них характерны отверстия в щёчных костях (боковые височные ямы с

Териодонты (зверозубые)
Циногнатус
Териодонты или зверозубые ящеры - вымершая группа тероморф, одна из
Териодонты (зверозубые)
Циногнатус
Териодонты или зверозубые ящеры - вымершая группа тероморф, одна из

Терапсиды = зверодужные
Пристероогнатус
Потомки цинодонтов. Отличались большим
разнообразием, некоторые были высокоспе-
циализированы.
Теменного отверстия
Терапсиды = зверодужные
Пристероогнатус
Потомки цинодонтов. Отличались большим
разнообразием, некоторые были высокоспе-
циализированы.
Теменного отверстия
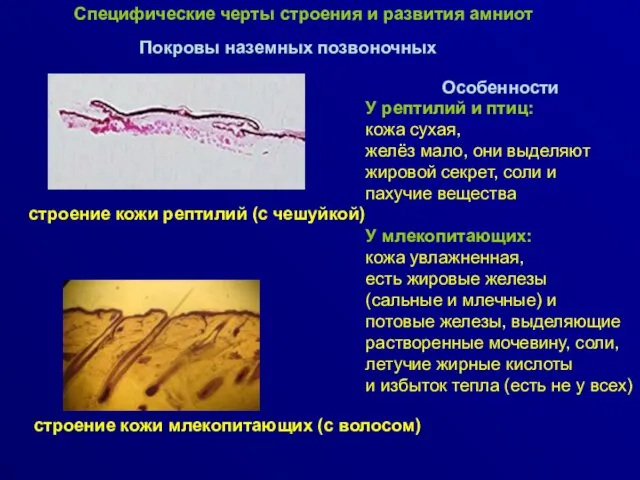
Покровы наземных позвоночных
строение кожи рептилий (с чешуйкой)
строение кожи млекопитающих (с волосом)
Особенности
У
Покровы наземных позвоночных
строение кожи рептилий (с чешуйкой)
строение кожи млекопитающих (с волосом)
Особенности
У

чешуя змеи
хвост бобра
щитки птиц
Производные кожи амниот
перо
шерсть
когти птиц
когти зверей
копыта
чешуя змеи
хвост бобра
щитки птиц
Производные кожи амниот
перо
шерсть
когти птиц
когти зверей
копыта
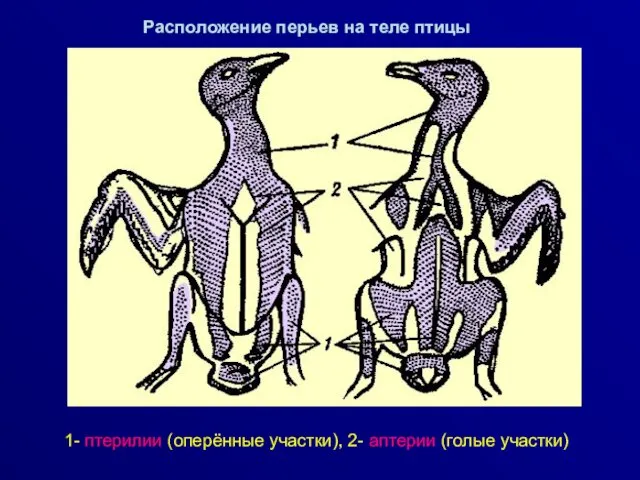
Расположение перьев на теле птицы
1- птерилии (оперённые участки), 2- аптерии (голые
Расположение перьев на теле птицы
1- птерилии (оперённые участки), 2- аптерии (голые

чехол клюва птиц
чехлы рогов зверей
китовый ус
Производные накладных костей
панцирь черепах
панцирь броненосца
рога зверей
чехол клюва птиц
чехлы рогов зверей
китовый ус
Производные накладных костей
панцирь черепах
панцирь броненосца
рога зверей

Водно-солевой обмен и осморегуляция у амниот
У рептилий, птиц и млекопитающих сначала
Водно-солевой обмен и осморегуляция у амниот
У рептилий, птиц и млекопитающих сначала

Типы белкового обмена:
рептилии, птицы
урикотелия - выделение кристаллической мочевой кислоты, не требую-щей
Типы белкового обмена:
рептилии, птицы
урикотелия - выделение кристаллической мочевой кислоты, не требую-щей
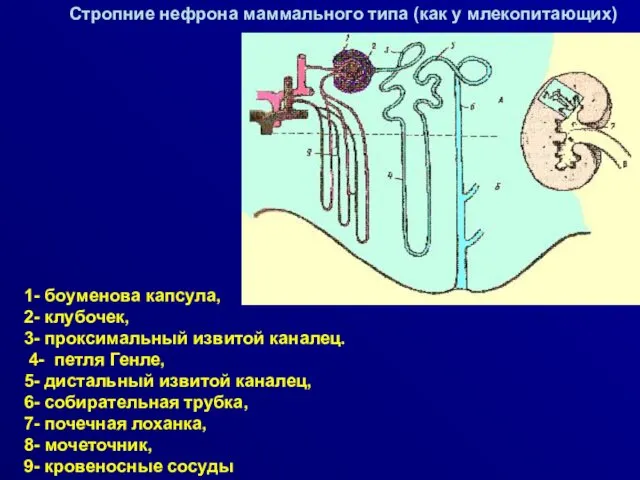
Стропние нефрона маммального типа (как у млекопитающих)
1- боуменова капсула,
2- клубочек,
Стропние нефрона маммального типа (как у млекопитающих)
1- боуменова капсула,
2- клубочек,

Почки амниот образуют корковый и мозговой слои.
У рептилий нефроны не имеют
Почки амниот образуют корковый и мозговой слои.
У рептилий нефроны не имеют
 Тест по информатике
Тест по информатике Применение свай при строительстве в вечной мерзлоте
Применение свай при строительстве в вечной мерзлоте презентация к уроку литературного краеведения по изучению творчества Бехтерева
презентация к уроку литературного краеведения по изучению творчества Бехтерева Применение Вооруженных Сил Российской Федерации в борьбе с терроризмом
Применение Вооруженных Сил Российской Федерации в борьбе с терроризмом Внутреннее устройство компьютера
Внутреннее устройство компьютера Где находится/ находятся …?
Где находится/ находятся …? Көкөністер мен жемістердің пестицидтермен, нитраттармен және ГМО-мен ластануы
Көкөністер мен жемістердің пестицидтермен, нитраттармен және ГМО-мен ластануы Шаблон презентации Лето
Шаблон презентации Лето Рождественский сочельник
Рождественский сочельник Наладка шлифовальных станков
Наладка шлифовальных станков социализация личности и откл поведение
социализация личности и откл поведение Принцип метода ЭКГ. Показания. Отведения. Нормальная ЭКГ. Электрическая ось сердца
Принцип метода ЭКГ. Показания. Отведения. Нормальная ЭКГ. Электрическая ось сердца Картотека загадок по лексической теме Дикие животные
Картотека загадок по лексической теме Дикие животные Декоративный рисунок. 4 класс
Декоративный рисунок. 4 класс Аппликация Павлин.
Аппликация Павлин. урок 13.04
урок 13.04 Возникновение и распад империи Карла Великого
Возникновение и распад империи Карла Великого Электронда есептеуіш машиналардың кезеңдері. ДК сәулетінің замануи даму бағыттары
Электронда есептеуіш машиналардың кезеңдері. ДК сәулетінің замануи даму бағыттары Лувр – палац-музей з світовим ім'ям
Лувр – палац-музей з світовим ім'ям Человек на войне 7 класс ИЗО
Человек на войне 7 класс ИЗО Актуальность. Примеры формулировок
Актуальность. Примеры формулировок Предварительно-напряженные листовые металлические конструкции
Предварительно-напряженные листовые металлические конструкции Презентация к уроку истории Путешествие в средневековый город
Презентация к уроку истории Путешествие в средневековый город Рабочее время
Рабочее время Межкультурная коммуникация. Формы коммуникации при межкультурном взаимодействии
Межкультурная коммуникация. Формы коммуникации при межкультурном взаимодействии Общественный строй Древней Руси
Общественный строй Древней Руси Операторы и выражения
Операторы и выражения Сечение. Фигура сечения на чертеже
Сечение. Фигура сечения на чертеже