- Этилен

Содержание
- 2. Ethylene concentration µL L-1 (ppm) -pea seedlings -grown in darkness for 48h -in the presence of
- 3. Выявление «этиленовых» мутантов (по тройному ответу) воздух этилен Дикий тип этилен Этилен-нечувствительный мутант (ein, etr) Мутант
- 4. Биосинтез этилена
- 5. Тканеспецифичность экспрессии генов ACS A, etiolated seedlings B, light grown seedlings 1, roots 2, leaves 3,
- 6. Регуляция экспрессии генов ACS: А - при поранении В - при обработке ИУК А В
- 7. Фенотипы мутантов eto (ethylene overproducing) и cin (cytokinin insensitive) Мутации cin – результат потери функции ACS
- 8. Уровень этилена в тканях регулируется через стабильность ACS белков А – Деградация ACS белков с помощью
- 9. Регуляция биосинтеза этилена через стабильность ACS белков ETO1 – субъединица убиквитин-лигазного комплекса CUL3-BTB
- 10. ethylene air 1. Мутанты нечувствительные к этилену (etr1, etr2, ers1, ers2, ein4 - доминантные); ein2, ein3,
- 11. Передача сигнала при ответе на этилен Основные компоненты: Рецепторы этилена: ETR1 и 2, ERS1 и 2,
- 12. Белки двухкомпонентных систем передачи сигнала у Arabidopsis A - Гибридные сенсорные гистидин-киназы АНК4/CRE1 (рецептор цитокинина) и
- 13. Пять этиленовых рецепторов Arabidopsis Домены: Трансмембранный (ТМ) домен – связывает этилен в присутствии кофактора (Cu 1+)
- 14. Сравнение систем передачи сигнала цитокинина и этилена Фосфореле Активация каждого последующего компонента предыдущим Нет передачи фосфатных
- 15. Гибридные гистидин-киназы CRE1 и CKI (рецепторы цитокинина), а также ETR1 (рецептор этилена) могут передавать фосфатную группу
- 16. Мутации с потерей функции одного из этиленовых рецепторов не имеют проявления, множественные мутанты характеризуются конститутивным ответом
- 17. В воздухе (без этилена): Рецепторы связывают и активируют протеинкиназу CTR1, которая негативно регулирует ответ на этилен
- 18. Кофакторы этиленовых рецепторов Ионы Cu+ - связывание этилена происходит только при их присоединении к рецептору Трансмембранный
- 19. Важные мотивы лиганд-вязывающих ТМ доменов: 1. мотивы из 7 консервативных аминокислот, необходимых для связывания меди (самая
- 20. Ser/Thr протеинкиназа CTR1 (МАРККК) – негативный регулятор этиленового сигналинга ETR CTR1 Ответ на этилен Мишени CTR1:
- 21. CTR1 образует олигомеры и вызывает олигомеризацию этиленовых рецепторов J. Mol. Biol.: October 2012 CTR1 рецепторы
- 23. Предполагаемая схема действия МАР-киназного каскада в этиленовом сигналинге При отсутствии этилена: Рецепторы активируют МАРККК киназу CTR1
- 24. Белок EIN2 - центральный регулятор этиленового сигналинга Nramp-like CEND (не важен для функции) (важен для функции)
- 25. Взаимодействие EIN2 с CTR1 и рецепторами этилена PNAS: November 2012 ETR CTR1 EIN2 EIN3, EIL1 Ответ
- 26. Этилен-зависимые транскрипционные факторы EIN3 и EIL1 Позитивные регуляторы этиленового сигналлинга Регуляторы транскрипции, локализуются в ядре Сверхэкспрессия
- 27. ТФ EIN3 – координаторы ответа на этилен и его взаимодействия с другими гормонами
- 28. Регуляция стабильности белков EIN3 Стабилизаторы EIN3: MAPK (фосфорилирование Т174), EIN2 (белок-белковые взаимодействия), EIN5 (деградация транскриптов EBF)
- 29. Регуляция стабильности белков EIN3 Белки EIN3 подвергаются протеасомо-зависимой деградации Связывание белков EIN3 с убиквитин-лигазным комплексом SCF
- 30. EIN3 EBF genes, etc. EBF mRNA EBF proteins Взаимодействие EIN3-EBF - негативная обратная связь в этиленовом
- 31. Рибонуклеаза EIN5/XRN4 регулирует уровень транскриптов EBF1/2 Продукт гена EIN5 был идентифицирован как 5’-3’-экзорибонуклеаза XRN4 Участвует в
- 32. Факты: Сверхэкспрессия 3’UTR генов EBF вызывает нечувствительность к этилену (эффект сильнее, чем от «цельных» генов EBF).
- 33. EIN2 репрессирует трансляцию EBF транскриптов полиU-шпильки в 3’UTR транскриптов EBF С-концевой домен EIN2 Участники: Экзорибонуклеаза EIN5
- 34. Полиуридиновые мотивы в 3‘UTR EBF генов
- 35. Опыты по ко-локализации 3‘UTR EBF с белками EIN2 и EIN5
- 36. Транскрипционные факторы ERF Транскрипционные факторы EIN3 и EIL связываются с последовательностью EBS (EIN3-Binding Site) в промоторе
- 37. Филогенетическое древо ERF белков разных видов растений
- 38. ERF-активаторы и ERF-репрессоры Содержат мотивы EDLL (Глн (E), Асп (D), Лей (L) (L)) Рекрутируют в промотор
- 39. ERFs Абиотический стресс (поранение, высокая температура, высыхание) LEA ? дегидрины GST ? глутатион-S-трансферазы Биотический стресс (поражение
- 40. Некоторые ERF - центральные регуляторы защиты от некротрофных патогенов Индуцируются в ответ на разные стимулы (не
- 41. Роль этилена в реакции сверхчувствительности (HR)
- 42. Фитогормоны и системная устойчивость растений 1. ISR (Induced Systemic Resistance) При взаимодействии с непатогенными микроорганизмами Основные
- 43. Газообразные гормоны – этилен и метил-жасмонат могут выступать в роли вторичных медиаторов при передаче стрессовых сигналов
- 44. Местный (HR) и системный (SAR) иммунный ответ у растений 1 2 3 4 5 PAMPs (pathogen-associated
- 45. PAMP-triggered (PTI) and Effector-trigegred (ETI) plant immunity: 2 барьера защиты
- 46. PAMP-triggered (PTI) and Effector-trigegred (ETI) plant immunity: 2 барьера защиты “Gene for gene”
- 47. ТФ NPR1 (nonexpresser of PR genes 1) – Центральный регулятор иммунитета растений (SAR) Может существовать в
- 48. Этилен и созревание плодов
- 49. Климактерические и неклимактерические плоды НЕклимактерические – пик интенсивности дыхания при старении Виноград Климактерические – пик интенсивности
- 50. Роль этилена и АБК в созревании плодов Регулируют одни и те же гены, действуют независимо Созревание
- 51. В процессе созревания климактерических плодов повышается биосинтез этилена ТФ RIN (Ripening Inhibitor) и CNR (Colorless Non-Ripening)
- 52. Роль этилена в старении листьев и созревании плодов Старение: разрушение хлорофилла, прекращение фотосинтеза, распад белков и
- 53. Внешние факторы не вызывают старения Внешние факторы могут вызвать старение Старение независимо от внешних факторов Контроль
- 54. Взаимодействие гормонов и ТФ в регуляции возрастозависимого старения листьев Гормоны «за старение»: Et, ABA, JA, SA
- 55. Взаимодействие гормонов и ТФ в регуляции старения листьев под воздействием внешних факторов Этилен старение в темноте
- 56. SAG (senescence-associated genes): ТФ семейств NAC, WRKY, AP2 Mишени EIN3 – гены OREASA1 и NAP1 –
- 58. Скачать презентацию

Ethylene concentration µL L-1 (ppm)
-pea seedlings
-grown in darkness for 48h
-in the
Ethylene concentration µL L-1 (ppm)
-pea seedlings
-grown in darkness for 48h
-in the
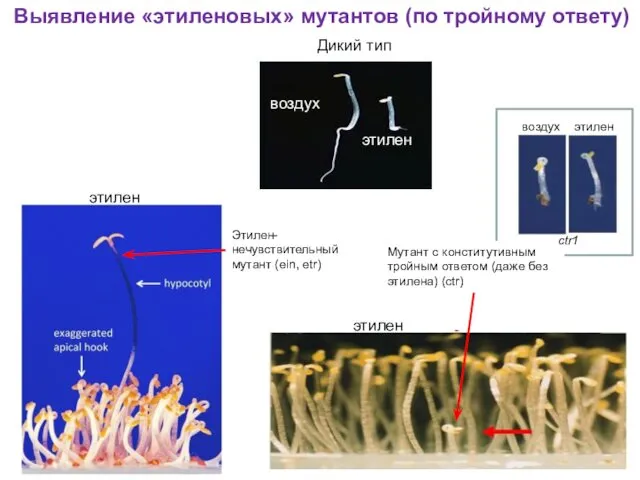
Выявление «этиленовых» мутантов (по тройному ответу)
воздух
этилен
Дикий тип
этилен
Этилен-нечувствительный мутант (ein, etr)
Мутант с
Выявление «этиленовых» мутантов (по тройному ответу)
воздух
этилен
Дикий тип
этилен
Этилен-нечувствительный мутант (ein, etr)
Мутант с
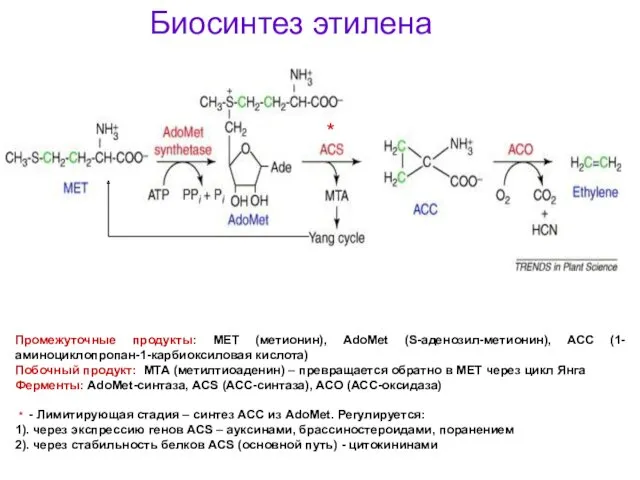
Биосинтез этилена
Биосинтез этилена
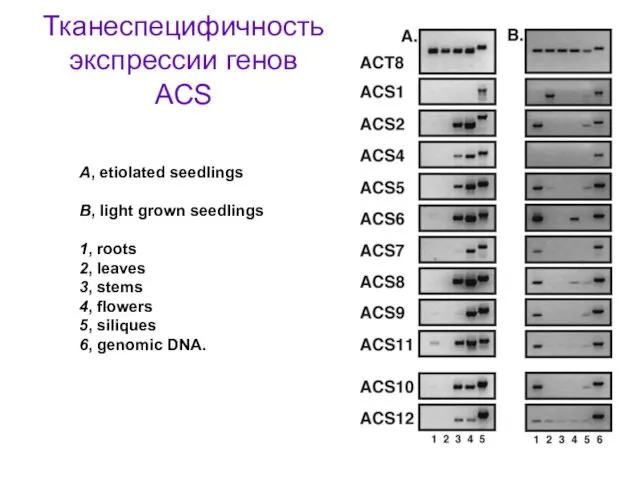
Тканеспецифичность экспрессии генов ACS
A, etiolated seedlings
B, light grown seedlings
1, roots
2,
Тканеспецифичность экспрессии генов ACS
A, etiolated seedlings
B, light grown seedlings
1, roots
2,
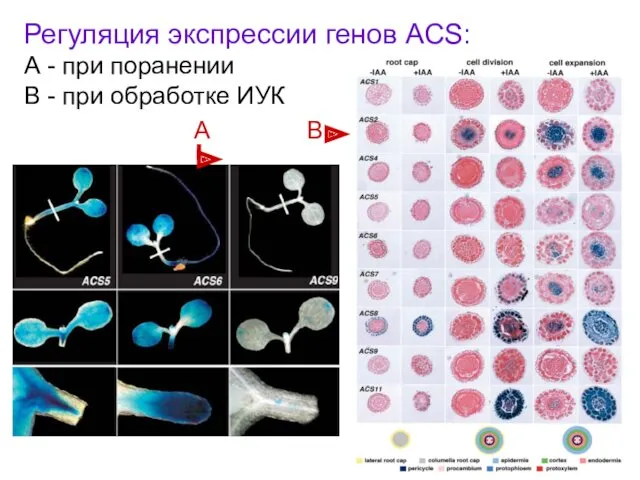
Регуляция экспрессии генов ACS:
А - при поранении
Регуляция экспрессии генов ACS:
А - при поранении
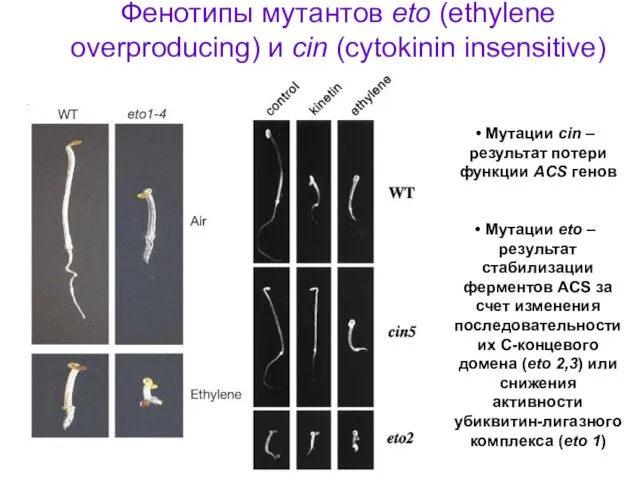
Фенотипы мутантов eto (ethylene overproducing) и cin (cytokinin insensitive)
Мутации cin
Фенотипы мутантов eto (ethylene overproducing) и cin (cytokinin insensitive)
Мутации cin
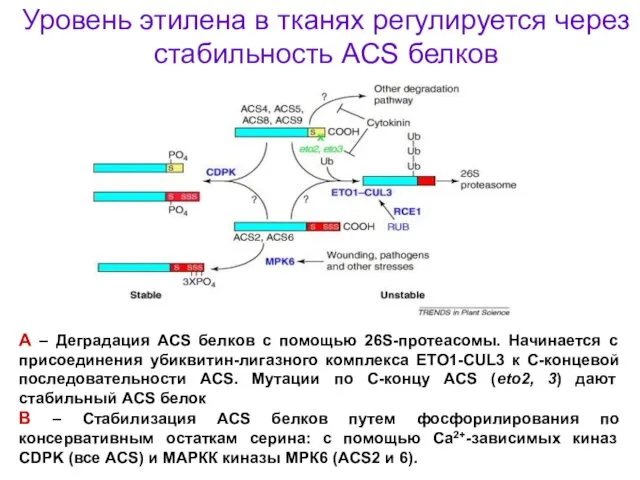
Уровень этилена в тканях регулируется через стабильность ACS белков
А – Деградация
Уровень этилена в тканях регулируется через стабильность ACS белков
А – Деградация
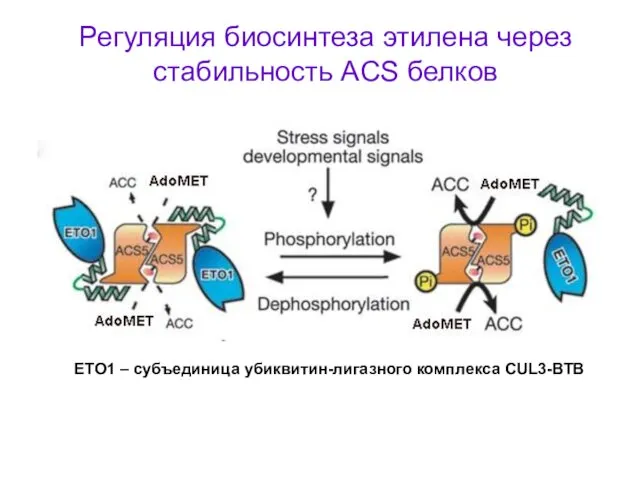
Регуляция биосинтеза этилена через стабильность ACS белков
ETO1 – субъединица убиквитин-лигазного
Регуляция биосинтеза этилена через стабильность ACS белков
ETO1 – субъединица убиквитин-лигазного
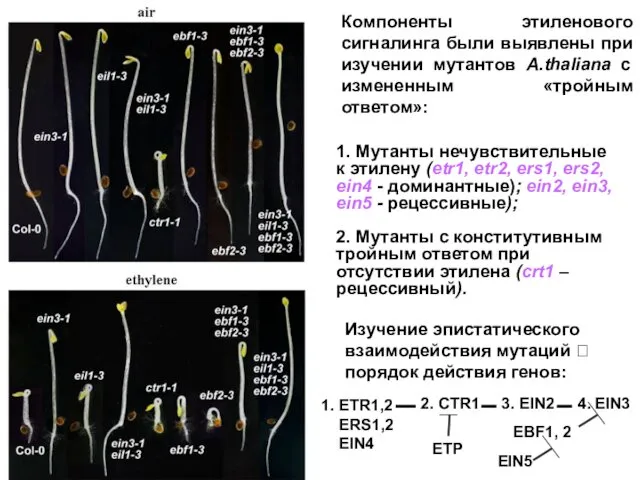
ethylene
air
1. Мутанты нечувствительные к этилену (etr1, etr2, ers1, ers2, ein4
ethylene
air
1. Мутанты нечувствительные к этилену (etr1, etr2, ers1, ers2, ein4
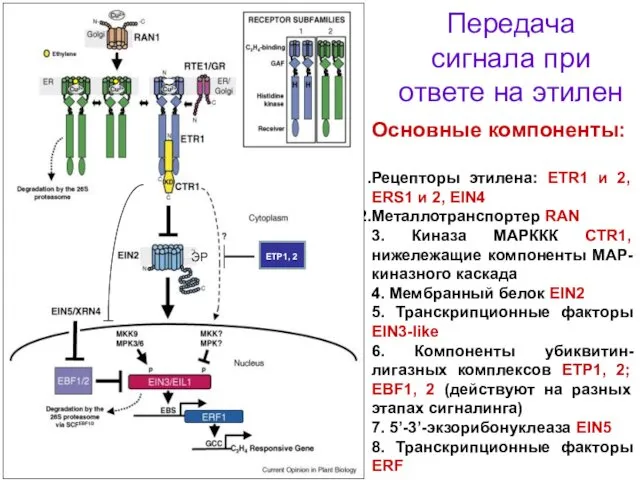
Передача сигнала при ответе на этилен
Основные компоненты:
Рецепторы этилена: ETR1 и 2,
Передача сигнала при ответе на этилен
Основные компоненты:
Рецепторы этилена: ETR1 и 2,
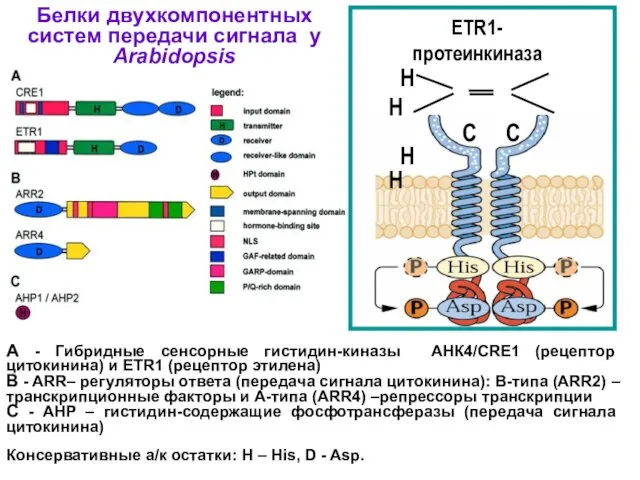
Белки двухкомпонентных систем передачи сигнала у Arabidopsis
A - Гибридные сенсорные гистидин-киназы
Белки двухкомпонентных систем передачи сигнала у Arabidopsis
A - Гибридные сенсорные гистидин-киназы
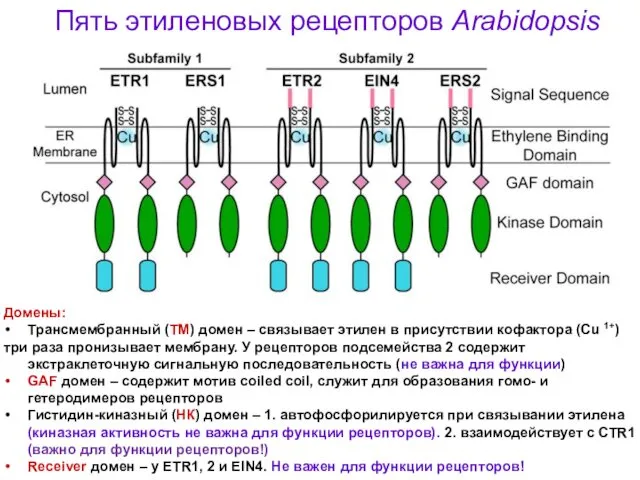
Пять этиленовых рецепторов Arabidopsis
Домены:
Трансмембранный (ТМ) домен – связывает этилен в присутствии
Пять этиленовых рецепторов Arabidopsis
Домены:
Трансмембранный (ТМ) домен – связывает этилен в присутствии
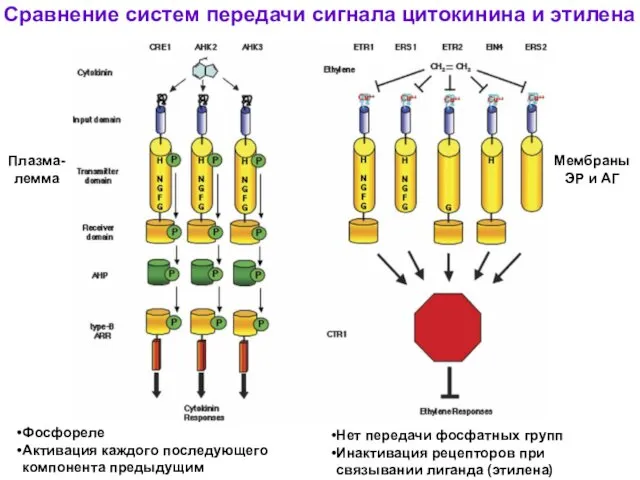
Сравнение систем передачи сигнала цитокинина и этилена
Фосфореле
Активация каждого последующего компонента предыдущим
Нет
Сравнение систем передачи сигнала цитокинина и этилена
Фосфореле
Активация каждого последующего компонента предыдущим
Нет
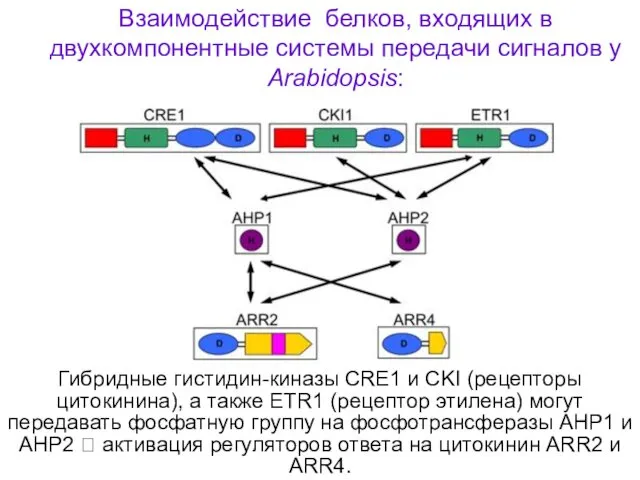
Гибридные гистидин-киназы CRE1 и CKI (рецепторы цитокинина), а также ETR1 (рецептор
Гибридные гистидин-киназы CRE1 и CKI (рецепторы цитокинина), а также ETR1 (рецептор

Мутации с потерей функции одного из этиленовых рецепторов не имеют проявления,
Мутации с потерей функции одного из этиленовых рецепторов не имеют проявления,
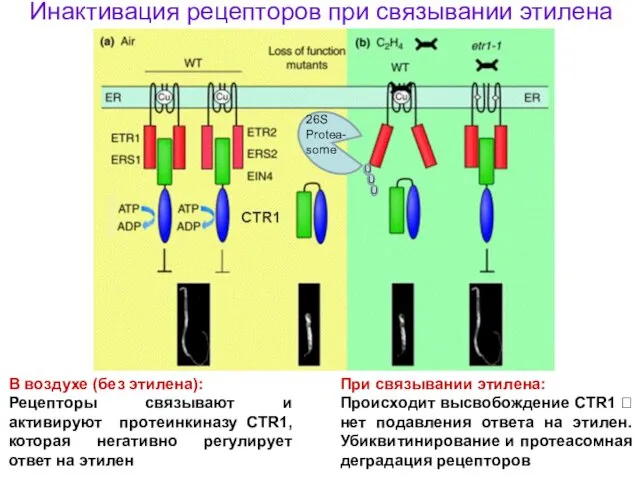
В воздухе (без этилена):
Рецепторы связывают и активируют протеинкиназу CTR1, которая негативно
В воздухе (без этилена):
Рецепторы связывают и активируют протеинкиназу CTR1, которая негативно
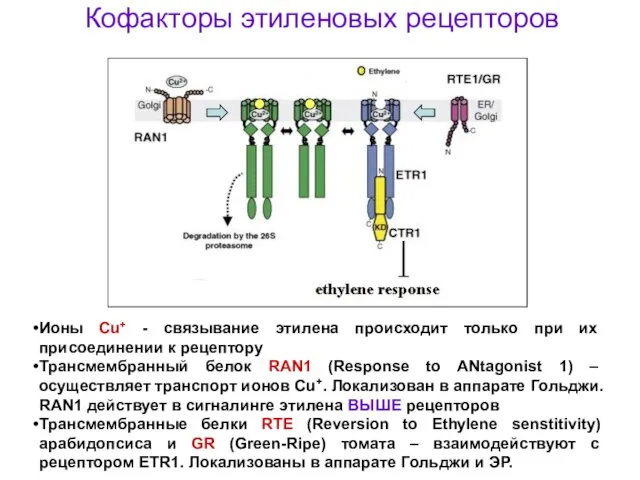
Кофакторы этиленовых рецепторов
Ионы Cu+ - связывание этилена происходит только при их
Кофакторы этиленовых рецепторов
Ионы Cu+ - связывание этилена происходит только при их
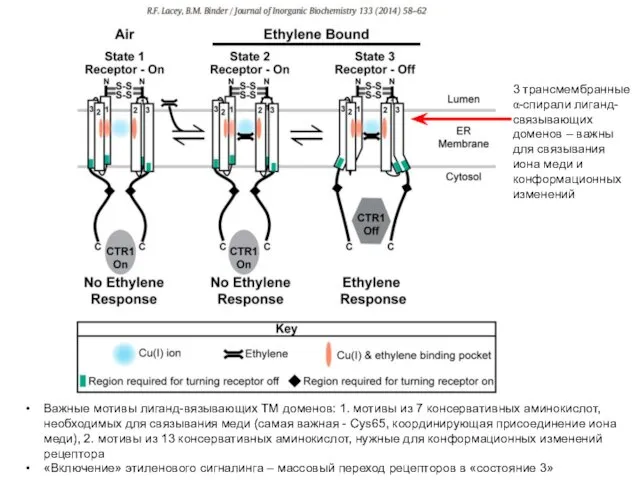
Важные мотивы лиганд-вязывающих ТМ доменов: 1. мотивы из 7 консервативных аминокислот,
Важные мотивы лиганд-вязывающих ТМ доменов: 1. мотивы из 7 консервативных аминокислот,
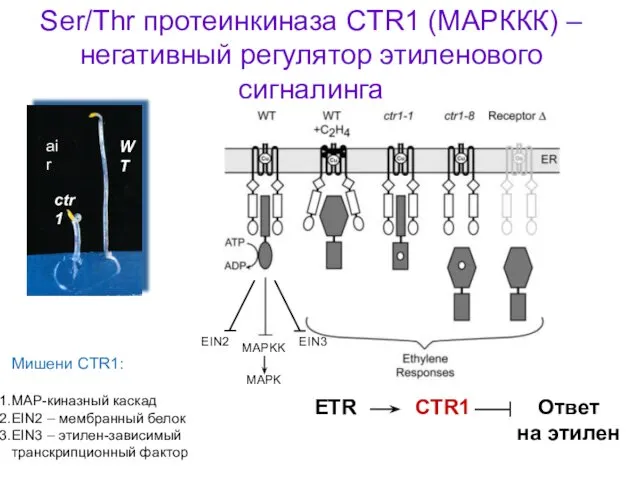
Ser/Thr протеинкиназа CTR1 (МАРККК) – негативный регулятор этиленового сигналинга
ETR
CTR1
Ответ
на этилен
Мишени
Ser/Thr протеинкиназа CTR1 (МАРККК) – негативный регулятор этиленового сигналинга
ETR
CTR1
Ответ
на этилен
Мишени
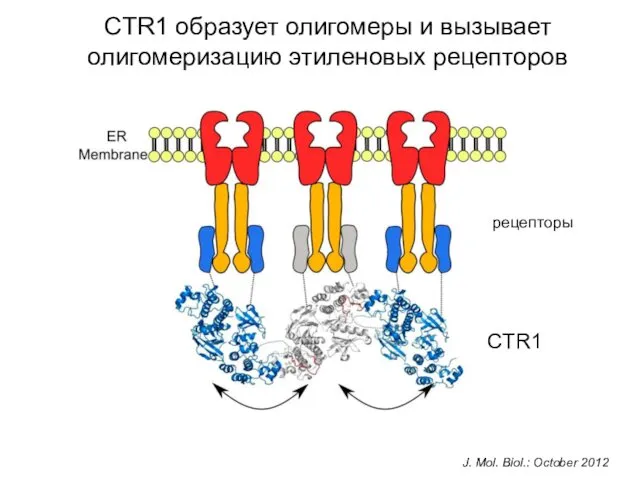
CTR1 образует олигомеры и вызывает олигомеризацию этиленовых рецепторов
J. Mol. Biol.: October
CTR1 образует олигомеры и вызывает олигомеризацию этиленовых рецепторов
J. Mol. Biol.: October
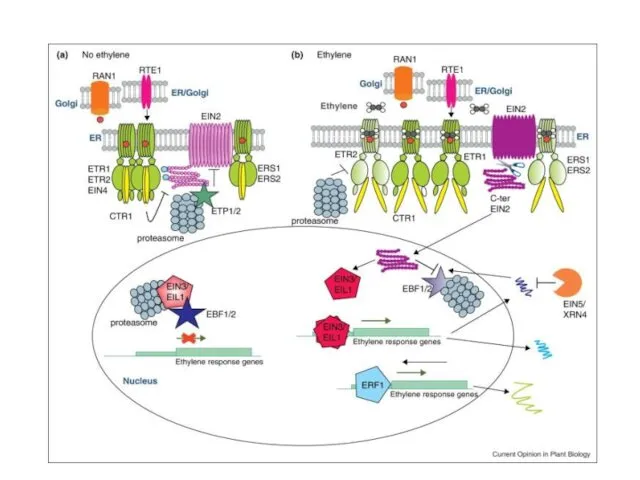
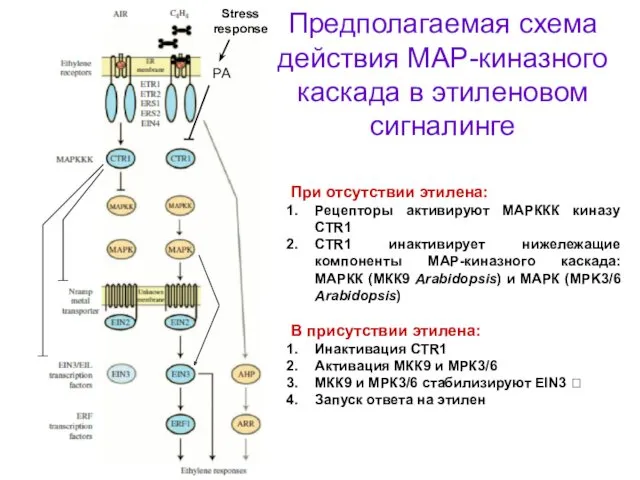
Предполагаемая схема действия МАР-киназного каскада в этиленовом сигналинге
При отсутствии этилена:
Рецепторы активируют
Предполагаемая схема действия МАР-киназного каскада в этиленовом сигналинге
При отсутствии этилена:
Рецепторы активируют
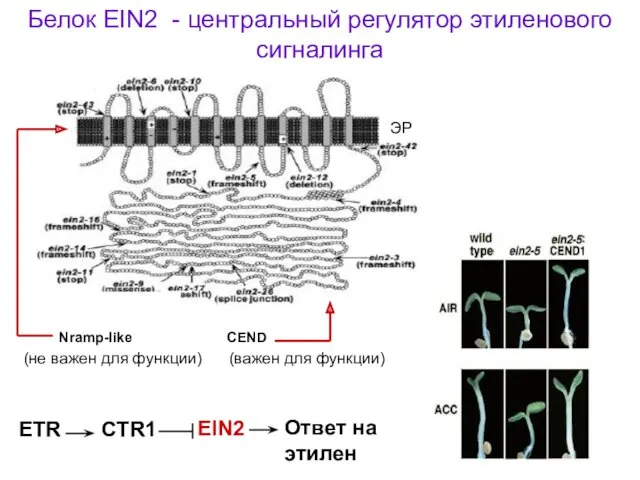
Белок EIN2 - центральный регулятор этиленового сигналинга
Nramp-like CEND
(не важен для функции)
(важен
Белок EIN2 - центральный регулятор этиленового сигналинга
Nramp-like CEND
(не важен для функции)
(важен
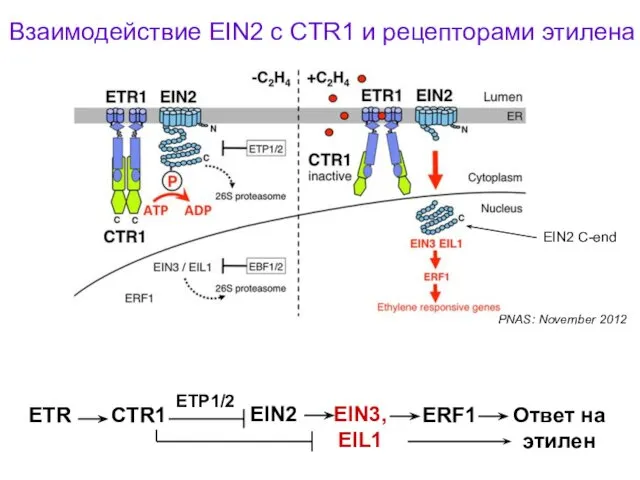
Взаимодействие EIN2 с CTR1 и рецепторами этилена
PNAS: November 2012
ETR
CTR1
EIN2
EIN3,
EIL1
Ответ на этилен
Взаимодействие EIN2 с CTR1 и рецепторами этилена
PNAS: November 2012
ETR
CTR1
EIN2
EIN3,
EIL1
Ответ на этилен
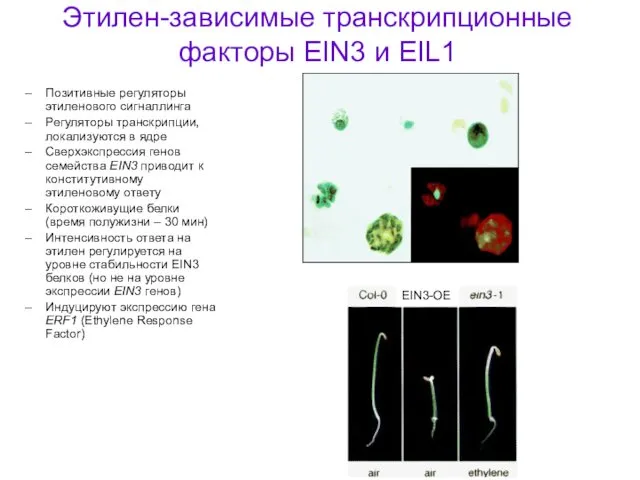
Этилен-зависимые транскрипционные факторы EIN3 и EIL1
Позитивные регуляторы этиленового сигналлинга
Регуляторы транскрипции, локализуются
Этилен-зависимые транскрипционные факторы EIN3 и EIL1
Позитивные регуляторы этиленового сигналлинга
Регуляторы транскрипции, локализуются
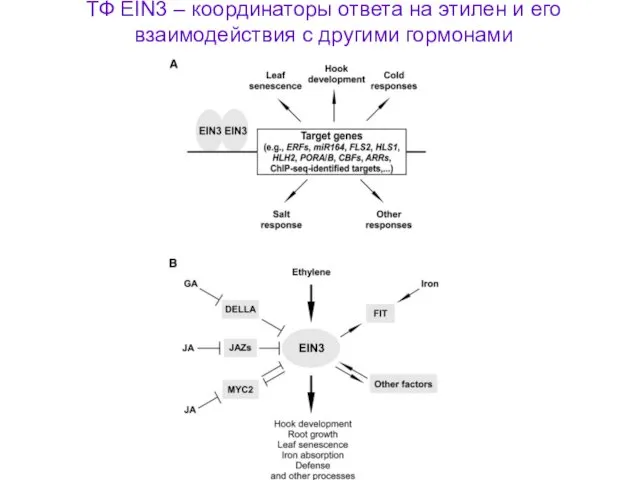
ТФ EIN3 – координаторы ответа на этилен и его взаимодействия с
ТФ EIN3 – координаторы ответа на этилен и его взаимодействия с
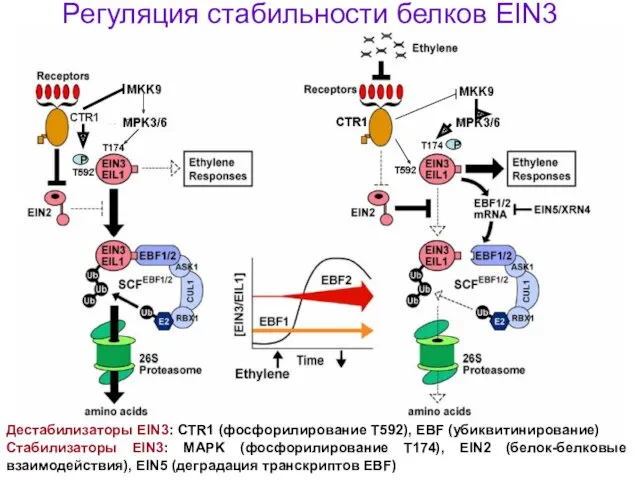
Регуляция стабильности белков EIN3
Стабилизаторы EIN3: MAPK (фосфорилирование Т174), EIN2 (белок-белковые взаимодействия),
Регуляция стабильности белков EIN3
Стабилизаторы EIN3: MAPK (фосфорилирование Т174), EIN2 (белок-белковые взаимодействия),

Регуляция стабильности белков EIN3
Белки EIN3 подвергаются протеасомо-зависимой деградации
Связывание белков EIN3
Регуляция стабильности белков EIN3
Белки EIN3 подвергаются протеасомо-зависимой деградации
Связывание белков EIN3
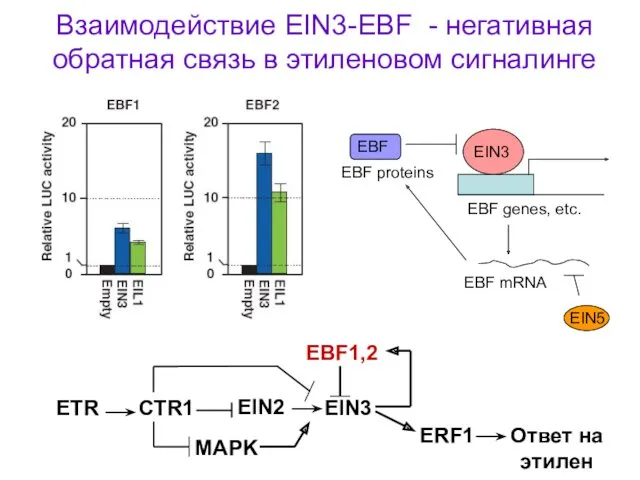
EIN3
EBF genes, etc.
EBF mRNA
EBF proteins
Взаимодействие EIN3-EBF - негативная обратная связь в
EIN3
EBF genes, etc.
EBF mRNA
EBF proteins
Взаимодействие EIN3-EBF - негативная обратная связь в
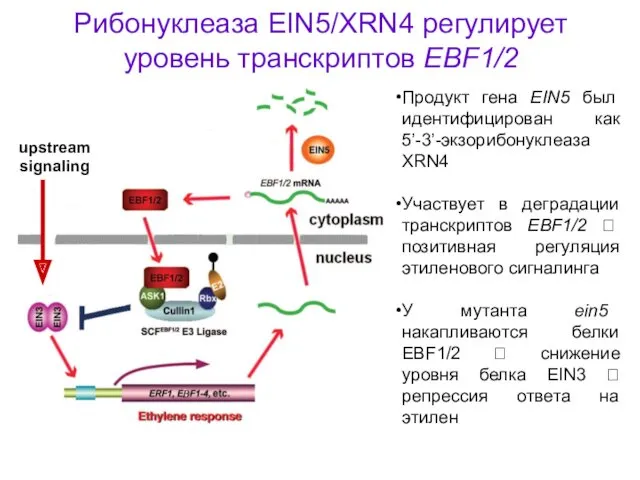
Рибонуклеаза EIN5/XRN4 регулирует уровень транскриптов EBF1/2
Продукт гена EIN5 был идентифицирован как
Рибонуклеаза EIN5/XRN4 регулирует уровень транскриптов EBF1/2
Продукт гена EIN5 был идентифицирован как
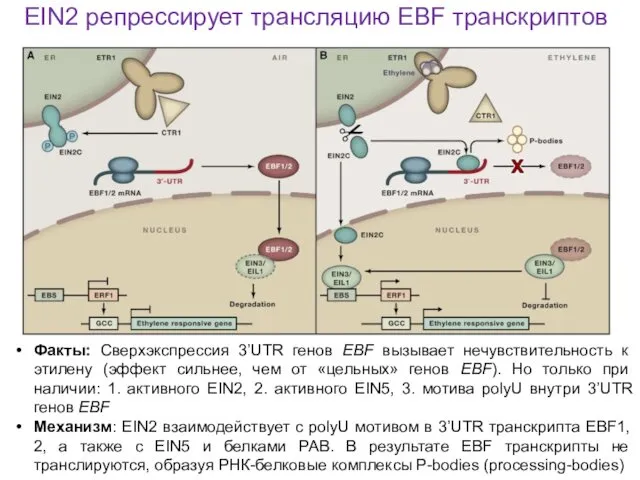
Факты: Сверхэкспрессия 3’UTR генов EBF вызывает нечувствительность к этилену (эффект сильнее,
Факты: Сверхэкспрессия 3’UTR генов EBF вызывает нечувствительность к этилену (эффект сильнее,
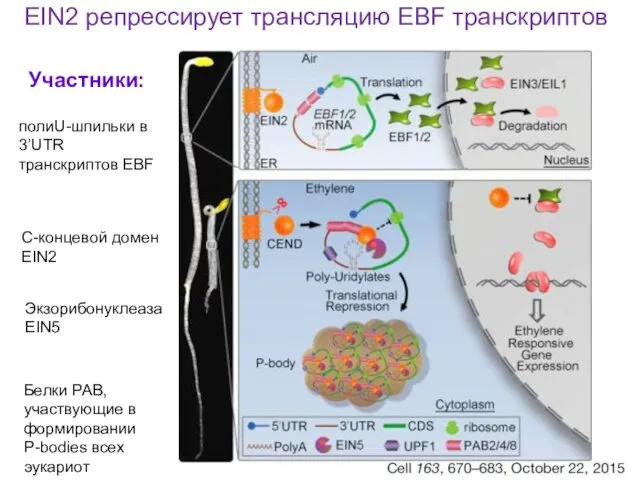
EIN2 репрессирует трансляцию EBF транскриптов
полиU-шпильки в 3’UTR транскриптов EBF
С-концевой домен
EIN2 репрессирует трансляцию EBF транскриптов
полиU-шпильки в 3’UTR транскриптов EBF
С-концевой домен
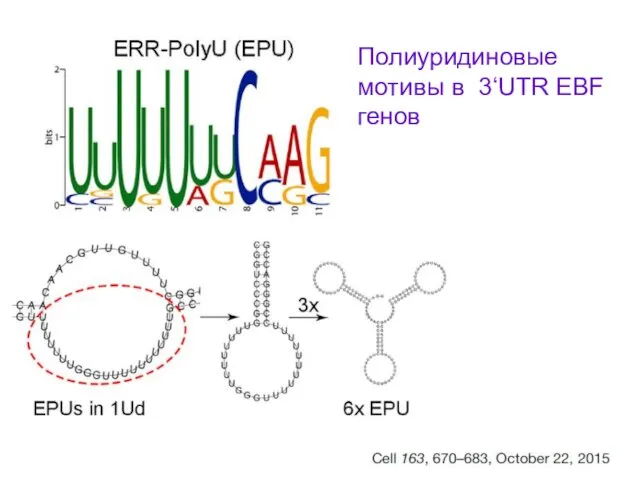
Полиуридиновые мотивы в 3‘UTR EBF генов
Полиуридиновые мотивы в 3‘UTR EBF генов
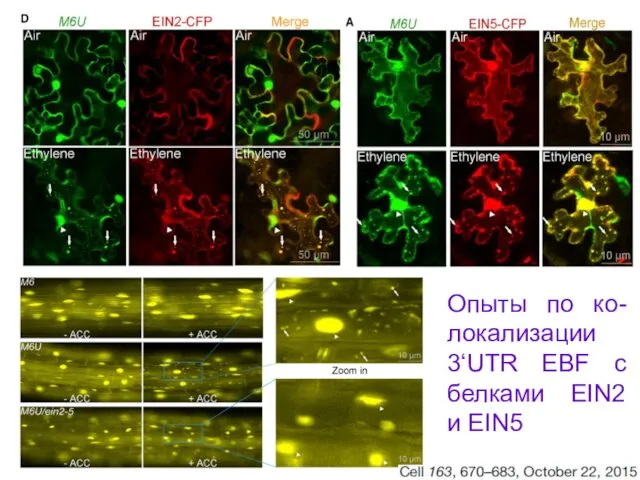
Опыты по ко-локализации 3‘UTR EBF с белками EIN2 и EIN5
Опыты по ко-локализации 3‘UTR EBF с белками EIN2 и EIN5
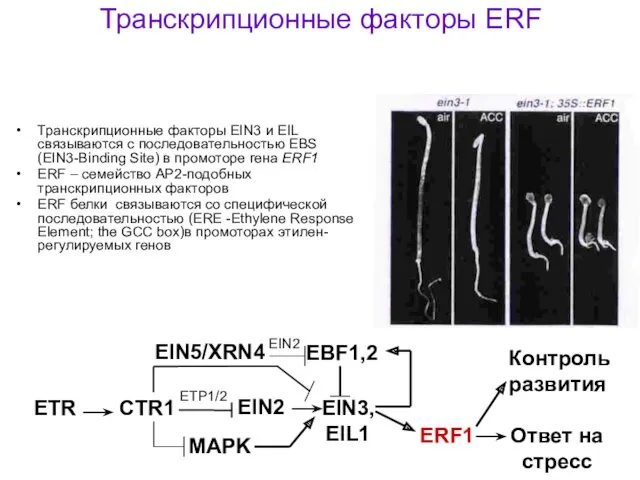
Транскрипционные факторы ERF
Транскрипционные факторы EIN3 и EIL связываются с последовательностью EBS
Транскрипционные факторы ERF
Транскрипционные факторы EIN3 и EIL связываются с последовательностью EBS
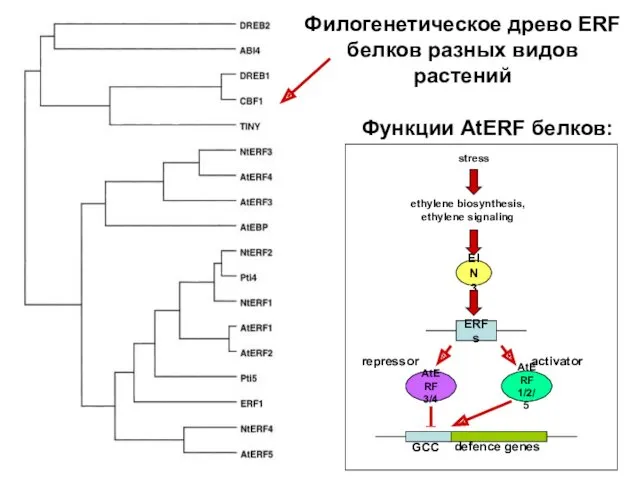
Филогенетическое древо ERF белков разных видов растений
Филогенетическое древо ERF белков разных видов растений
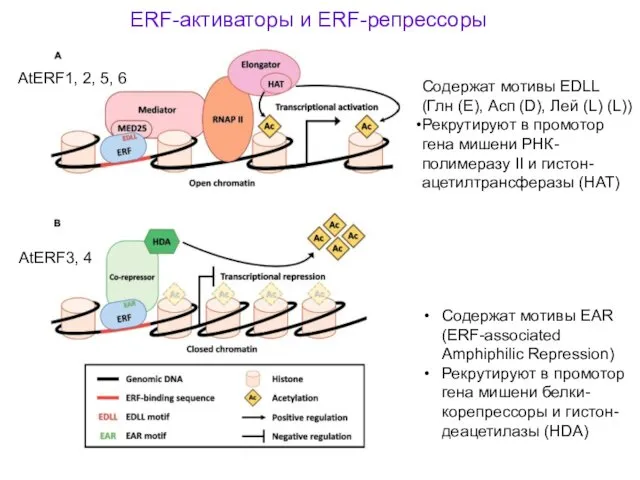
ERF-активаторы и ERF-репрессоры
Содержат мотивы EDLL (Глн (E), Асп (D), Лей (L)
ERF-активаторы и ERF-репрессоры
Содержат мотивы EDLL (Глн (E), Асп (D), Лей (L)
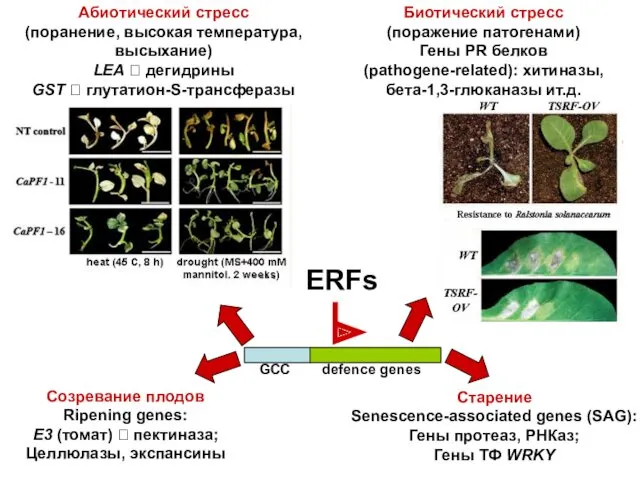
ERFs
Абиотический стресс
(поранение, высокая температура, высыхание)
LEA ? дегидрины
GST ? глутатион-S-трансферазы
Биотический стресс
(поражение
ERFs
Абиотический стресс
(поранение, высокая температура, высыхание)
LEA ? дегидрины
GST ? глутатион-S-трансферазы
Биотический стресс
(поражение
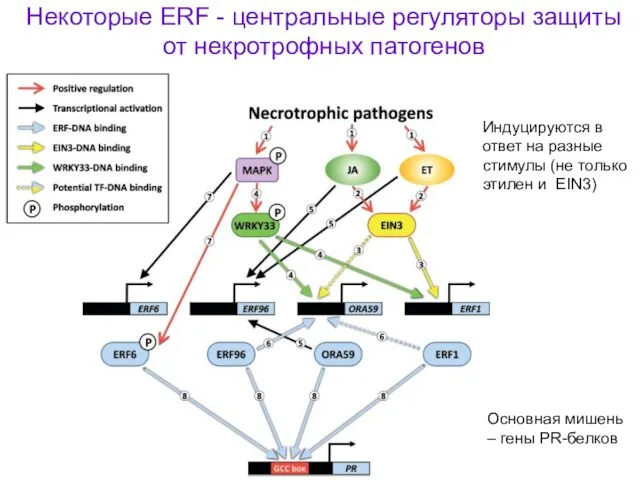
Некоторые ERF - центральные регуляторы защиты от некротрофных патогенов
Индуцируются в ответ
Некоторые ERF - центральные регуляторы защиты от некротрофных патогенов
Индуцируются в ответ
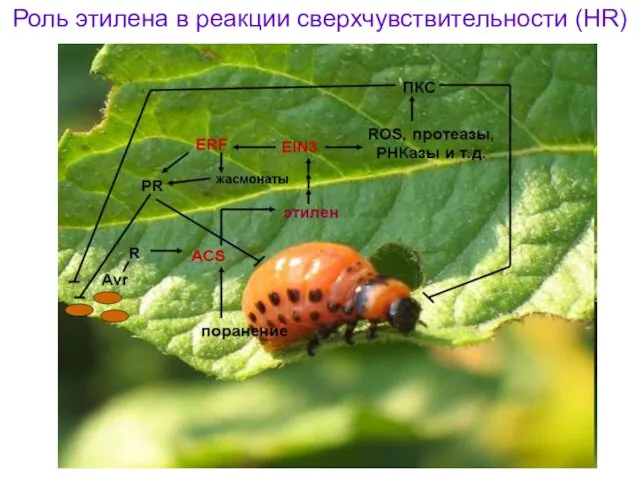
Роль этилена в реакции сверхчувствительности (HR)
Роль этилена в реакции сверхчувствительности (HR)
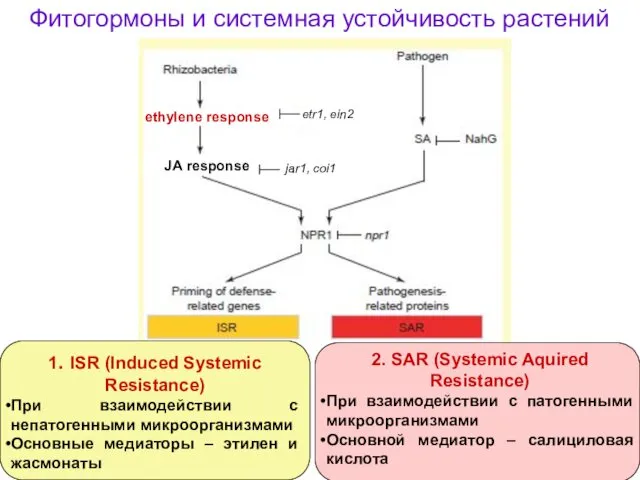
Фитогормоны и системная устойчивость растений
1. ISR (Induced Systemic Resistance)
При взаимодействии
Фитогормоны и системная устойчивость растений
1. ISR (Induced Systemic Resistance)
При взаимодействии
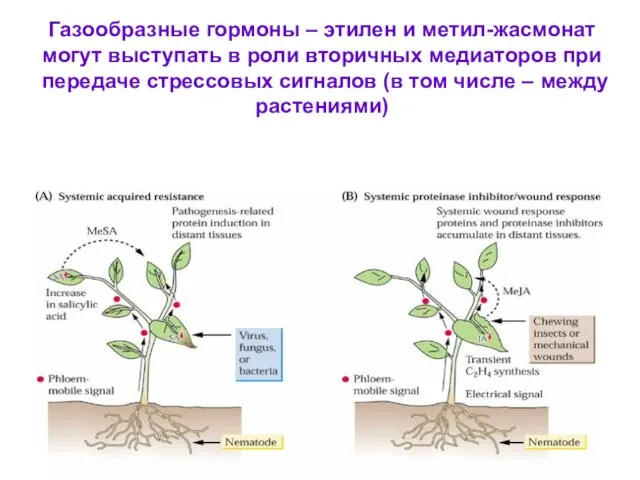
Газообразные гормоны – этилен и метил-жасмонат могут выступать в роли вторичных
Газообразные гормоны – этилен и метил-жасмонат могут выступать в роли вторичных
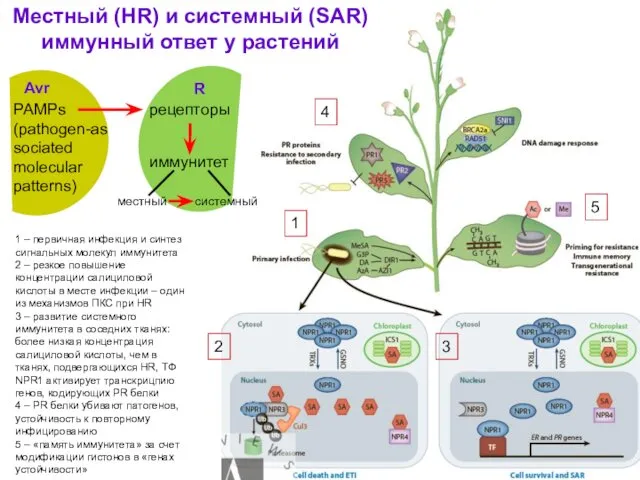
Местный (HR) и системный (SAR) иммунный ответ у растений
1
2
3
4
5
PAMPs (pathogen-associated
Местный (HR) и системный (SAR) иммунный ответ у растений
1
2
3
4
5
PAMPs (pathogen-associated
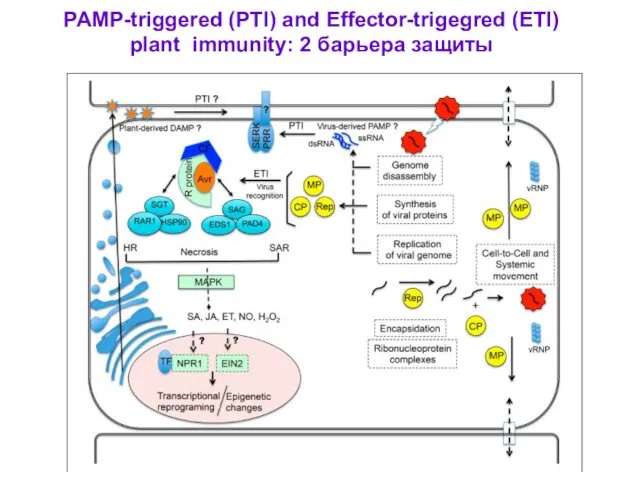
PAMP-triggered (PTI) and Effector-trigegred (ETI)
plant immunity: 2 барьера защиты
PAMP-triggered (PTI) and Effector-trigegred (ETI)
plant immunity: 2 барьера защиты
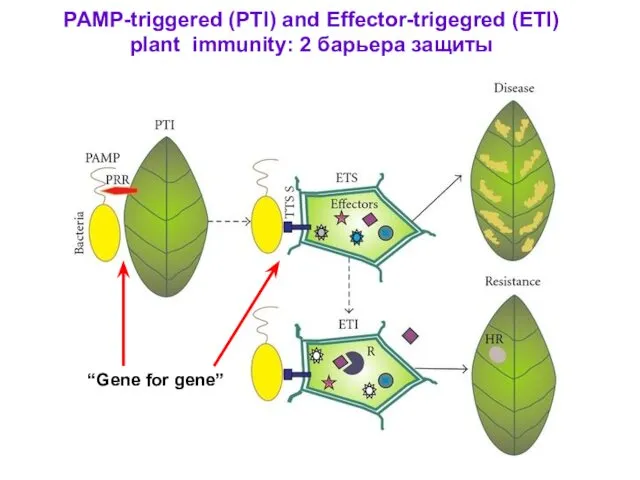
PAMP-triggered (PTI) and Effector-trigegred (ETI)
plant immunity: 2 барьера защиты
“Gene
PAMP-triggered (PTI) and Effector-trigegred (ETI)
plant immunity: 2 барьера защиты
“Gene
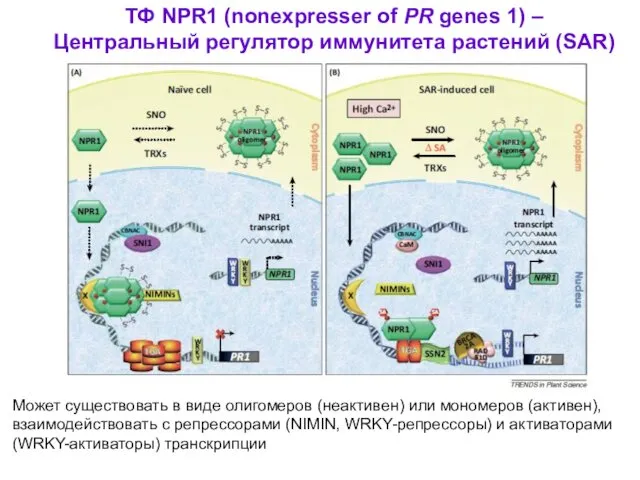
ТФ NPR1 (nonexpresser of PR genes 1) –
Центральный регулятор иммунитета
ТФ NPR1 (nonexpresser of PR genes 1) –
Центральный регулятор иммунитета
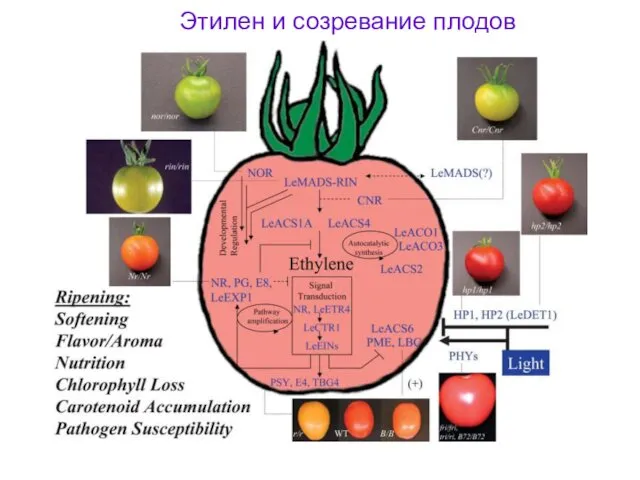
Этилен и созревание плодов
Этилен и созревание плодов
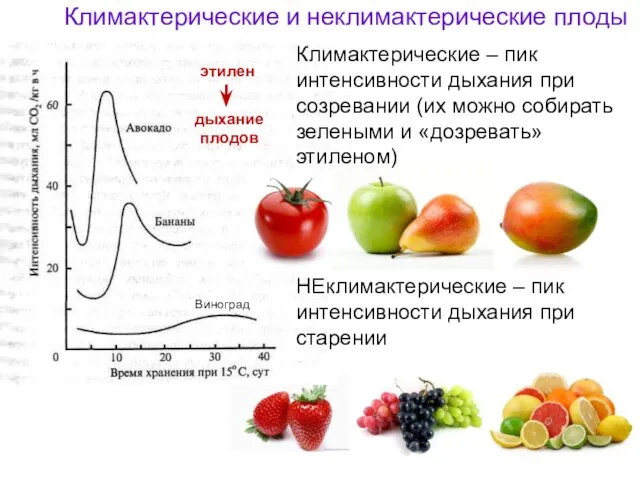
Климактерические и неклимактерические плоды
НЕклимактерические – пик интенсивности дыхания при старении
Виноград
Климактерические –
Климактерические и неклимактерические плоды
НЕклимактерические – пик интенсивности дыхания при старении
Виноград
Климактерические –
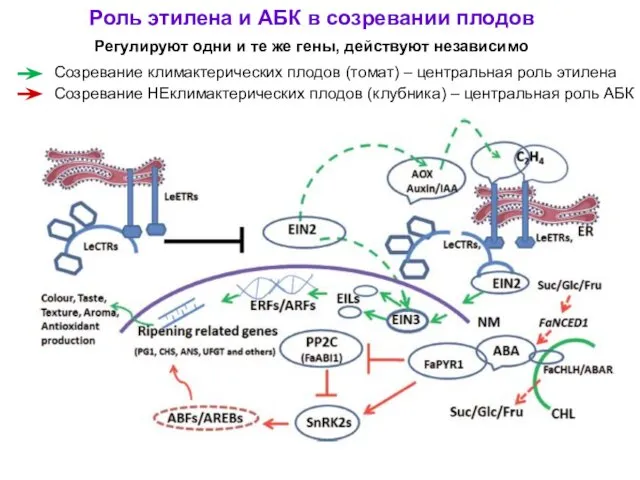
Роль этилена и АБК в созревании плодов
Регулируют одни и те же
Роль этилена и АБК в созревании плодов
Регулируют одни и те же
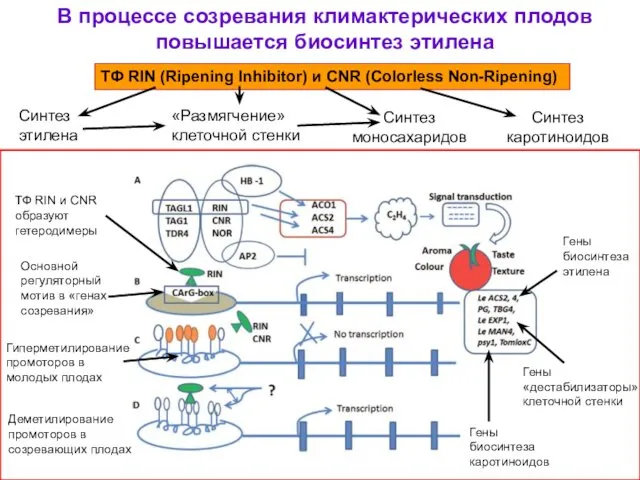
В процессе созревания климактерических плодов повышается биосинтез этилена
ТФ RIN (Ripening Inhibitor)
В процессе созревания климактерических плодов повышается биосинтез этилена
ТФ RIN (Ripening Inhibitor)
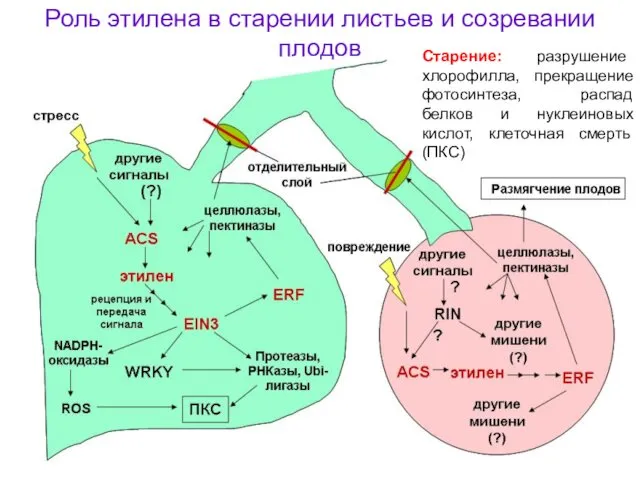
Роль этилена в старении листьев и созревании плодов
Старение: разрушение хлорофилла, прекращение
Роль этилена в старении листьев и созревании плодов
Старение: разрушение хлорофилла, прекращение
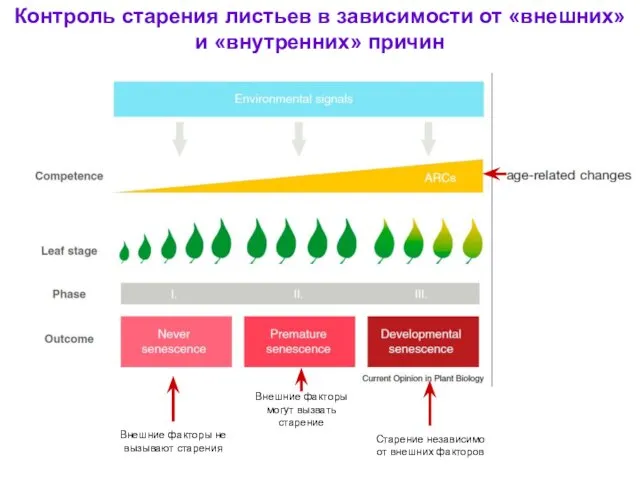
Внешние факторы не вызывают старения
Внешние факторы могут вызвать старение
Старение независимо от
Внешние факторы не вызывают старения
Внешние факторы могут вызвать старение
Старение независимо от
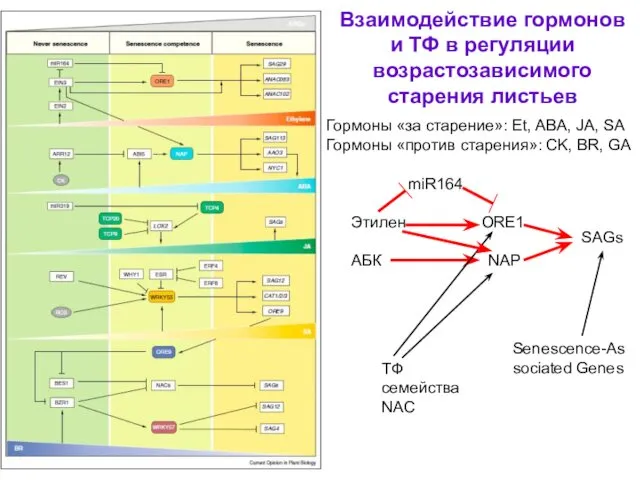
Взаимодействие гормонов и ТФ в регуляции возрастозависимого старения листьев
Гормоны «за старение»:
Взаимодействие гормонов и ТФ в регуляции возрастозависимого старения листьев
Гормоны «за старение»:
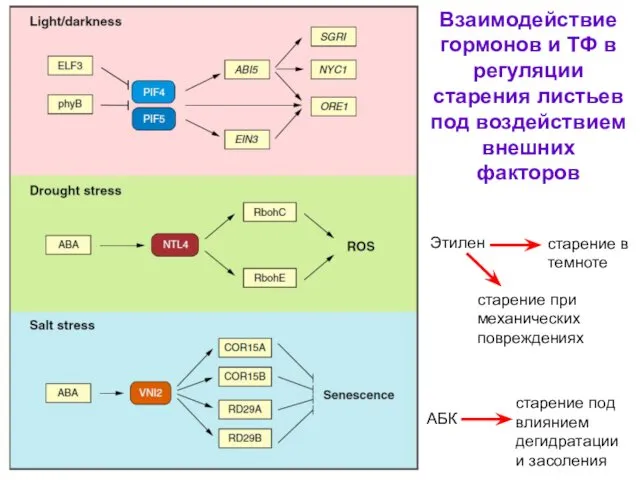
Взаимодействие гормонов и ТФ в регуляции старения листьев под воздействием внешних
Взаимодействие гормонов и ТФ в регуляции старения листьев под воздействием внешних
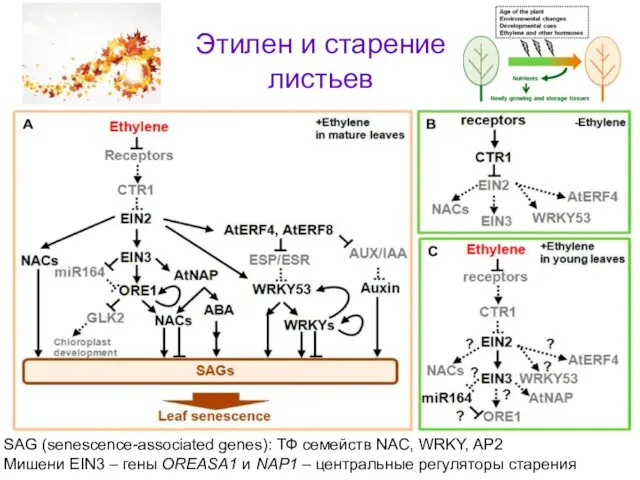
SAG (senescence-associated genes): ТФ семейств NAC, WRKY, AP2
Mишени EIN3 – гены
SAG (senescence-associated genes): ТФ семейств NAC, WRKY, AP2
Mишени EIN3 – гены
 Предметы
Предметы Родительское собрание Семейное чтение
Родительское собрание Семейное чтение История и методология биологии
История и методология биологии викторина Своя игра
викторина Своя игра Государство на берегах Нила
Государство на берегах Нила Производство 19 века
Производство 19 века Презентация № 3 Воздух, которым мы дышим
Презентация № 3 Воздух, которым мы дышим Корзинка на Пасху
Корзинка на Пасху Работа с данными типа запись. Лабораторная работа
Работа с данными типа запись. Лабораторная работа Образы кошки и собаки в русских и английских пословицах и поговорках. Сходства и различия
Образы кошки и собаки в русских и английских пословицах и поговорках. Сходства и различия Баскетбол • История возникновения • Правила соревнований
Баскетбол • История возникновения • Правила соревнований Реакции и революции в Европе 1820-1840-х гг
Реакции и революции в Европе 1820-1840-х гг Управление световым оформлением зданий
Управление световым оформлением зданий Загорский Петр Андреевич
Загорский Петр Андреевич Взаимосвязь эмпатии личности и выбора стратегий поведения в конфликте (на примере общеобразовательной организации)
Взаимосвязь эмпатии личности и выбора стратегий поведения в конфликте (на примере общеобразовательной организации) Мезозойская эра Земли
Мезозойская эра Земли Трудовые споры
Трудовые споры Некариозные поражения, возникающие после прорезывания зуба. Этиология, клиника, диагностика, лечение
Некариозные поражения, возникающие после прорезывания зуба. Этиология, клиника, диагностика, лечение Реализация комплексной образовательной программы Школа 2100 в МБДОУ ЦРР-ДС № 28 Золотой петушок г. Тамбова
Реализация комплексной образовательной программы Школа 2100 в МБДОУ ЦРР-ДС № 28 Золотой петушок г. Тамбова Великий праздник Пасха
Великий праздник Пасха Форма государства
Форма государства Таныш булайыҡ Архангел районы Ҡыҫынды
Таныш булайыҡ Архангел районы Ҡыҫынды Культура и духовная жизнь общества
Культура и духовная жизнь общества Основные направления в психологии
Основные направления в психологии Методология написания научных статей
Методология написания научных статей План воспитательной работы
План воспитательной работы Петербургские традиции 8 класс История и культура Санкт-Петербурга
Петербургские традиции 8 класс История и культура Санкт-Петербурга Презентация. 9 мая. Диск
Презентация. 9 мая. Диск