- Физиология мышц

Содержание
- 2. Проведение ПД по мембране ПД проводится от точки к каждой соседней ранее не возбужденной точке
- 3. Проведение ПД по миелинизированному нервному волокну
- 4. Синапс
- 5. Синаптическая щель Ширина - 20-30 нм Синаптическая щель заполнена синаптической жидкостью, своим составом напоминающей плазму крови.
- 6. Медиатор (химический посредник) Внутри нервного окончания имеется большое количество (до 300.000) синаптических пузырьков (диаметром около 50
- 7. Взаимодействие медиатора с постсинаптической мебраной Медиатор диффундирует по синаптической жидкости и большая часть молекул его достигает
- 8. Нервно-мышечный синапс 1 - пресинаптическая мембрана, 2 - пузырьки с ацетилхолином, 3 - митохондрии, 4 -
- 9. Выброс медиатора обеспечивает взаимодействие его с лигандзависимыми структурами канала
- 10. Явление суммации. Обычно для передачи одного ПД высвобождается до миллиона молекул АХ (200-300 везикул). Обозначения: а,
- 11. Переход ПКП в ВПСП ПД по нерву могут поступать с максимальной частотой до 1000 в с.
- 12. Восстановление медиатора в синаптической бляшке В нервном волокне происходит постоянное пополнение медиатора. Здесь имеется несколько механизмов
- 13. Депо кальция – саркоплазматический ретикулум 1- миофибриллы, 2 – саркоплазматический ретикулум, 3 – цистерны, 4 –
- 14. Схема строения мышечного волокна Саркомер - с двух сторон ограничен Z – линиями. Толстые – миозиновые,
- 15. Взаимодействие актиновых и миозиновых филаментов
- 16. Схема строения актиновых и миозиновых филаментов
- 17. Этапы «шагового» механизма Последовательные этапы: а – расслабление, б – соединение миозиновых головок с активным центром
- 18. Кальмодулин - идентичен тропонину С, имеющемуся в тонких нитях Присоединяя Са2+, кальмодулин способствует активации АТФазы и
- 19. Кальций Деполяризация мембраны цистерн открывает электровозбудимые кальциевые каналы. В связи с тем, что в саркоплазме концентрация
- 20. «Шаговый» механизм
- 21. Головка миозина и «шаг»
- 22. Роль кальция в мышечном сокращении 1 – Выброс медиатора в синаптическую щель. 2 – Освобождение активного
- 23. Различные режимы сокращения мышц А - одиночное сокращение, Б – неполный тетанус, В – полный тетанус.
- 24. Соотношение ПД и рефрактерности 5 – фаза абсолютной рефрактерности, 6 – ф. относительной рефрактерности, 7 -
- 25. Электромиограмма (А – одиночные ДЕ; Б – мышца в целом)
- 26. Роль АТФ в мышечном сокращении а) сокращения (образования мостиков); б) расслабления (разрыва мостиков); в) работы Са-насоса
- 27. Пути ресинтеза АТФ 1) креатинфосфокиназный (КФ): АДФ + КФ АТФ + К 2) гликолитический, З) аэробное
- 28. Максимальная мощность путей ресинтеза АТФ а) фосфагенный (КФ) - 3,6 моль АТФ/мин, б) гликолитический - 1,2
- 29. Двигательные единицы - Единичное нервное волокно мотонейрона и, иннервируемые им мышечные волокна, составляют одну ДЕ 1
- 30. Быстрые и медленные ДЕ Быстрые Большой мотонейрон. Много АТФ. Много КФ. Активный гликолиз. Сильные, но быстро
- 32. Скачать презентацию
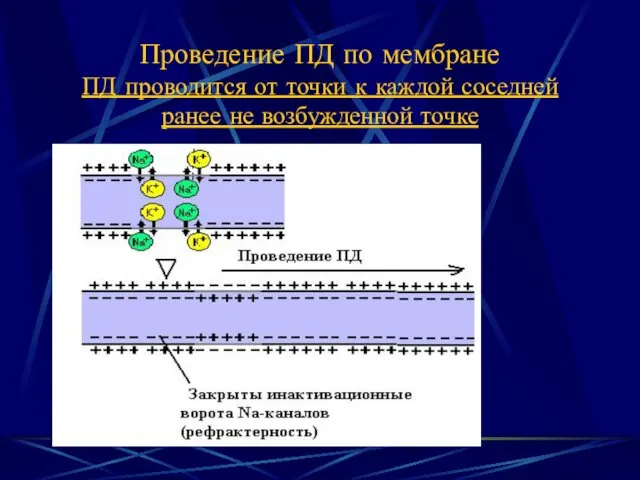
Проведение ПД по мембране
ПД проводится от точки к каждой соседней ранее
Проведение ПД по мембране ПД проводится от точки к каждой соседней ранее
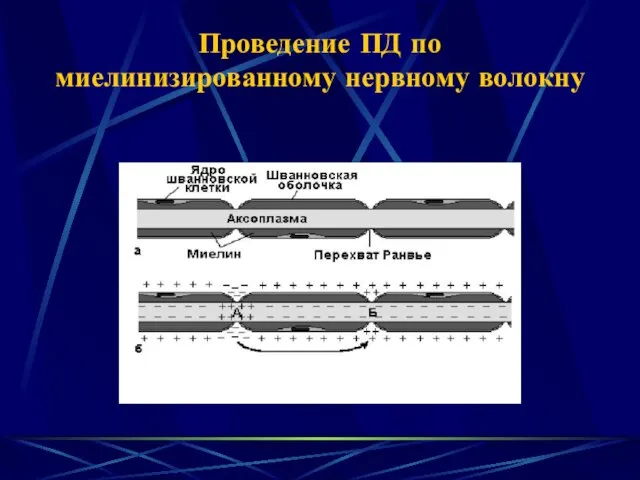
Проведение ПД по миелинизированному нервному волокну
Проведение ПД по миелинизированному нервному волокну
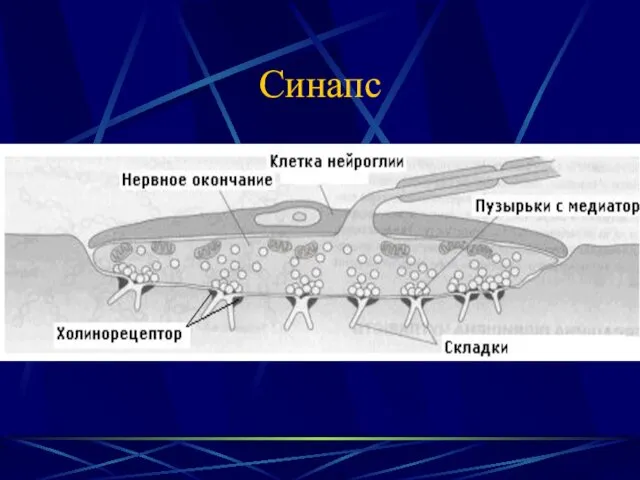
Синапс
Синапс

Синаптическая щель
Ширина - 20-30 нм
Синаптическая щель заполнена синаптической жидкостью, своим
Синаптическая щель
Ширина - 20-30 нм
Синаптическая щель заполнена синаптической жидкостью, своим

Медиатор
(химический посредник)
Внутри нервного окончания имеется большое количество (до 300.000) синаптических пузырьков
Медиатор
(химический посредник)
Внутри нервного окончания имеется большое количество (до 300.000) синаптических пузырьков

Взаимодействие медиатора с постсинаптической мебраной
Медиатор диффундирует по синаптической жидкости и большая
Взаимодействие медиатора с постсинаптической мебраной
Медиатор диффундирует по синаптической жидкости и большая
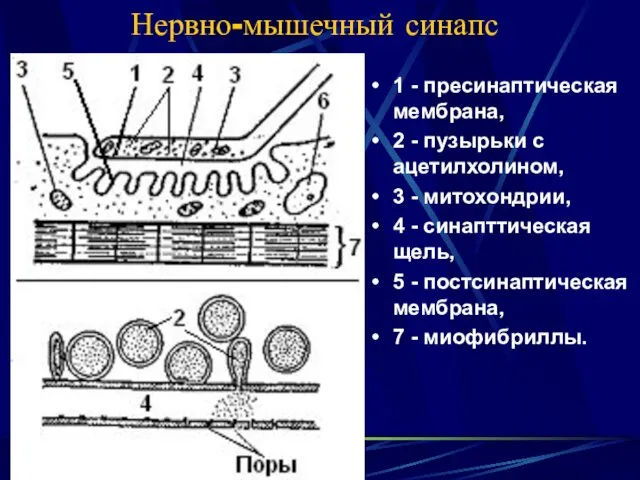
Нервно-мышечный синапс
1 - пресинаптическая мембрана,
2 - пузырьки с ацетилхолином,
3
Нервно-мышечный синапс
1 - пресинаптическая мембрана,
2 - пузырьки с ацетилхолином,
3
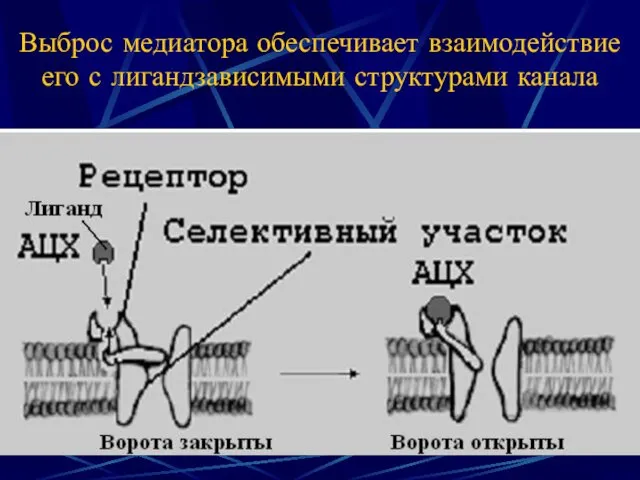
Выброс медиатора обеспечивает взаимодействие его с лигандзависимыми структурами канала
Выброс медиатора обеспечивает взаимодействие его с лигандзависимыми структурами канала
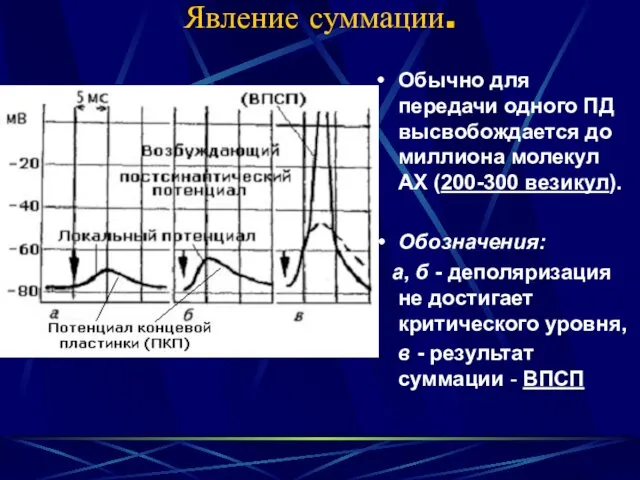
Явление суммации.
Обычно для передачи одного ПД высвобождается до миллиона молекул АХ
Явление суммации.
Обычно для передачи одного ПД высвобождается до миллиона молекул АХ

Переход ПКП в ВПСП
ПД по нерву могут поступать с максимальной частотой
Переход ПКП в ВПСП
ПД по нерву могут поступать с максимальной частотой

Восстановление медиатора в синаптической бляшке
В нервном волокне происходит постоянное пополнение
Восстановление медиатора в синаптической бляшке
В нервном волокне происходит постоянное пополнение
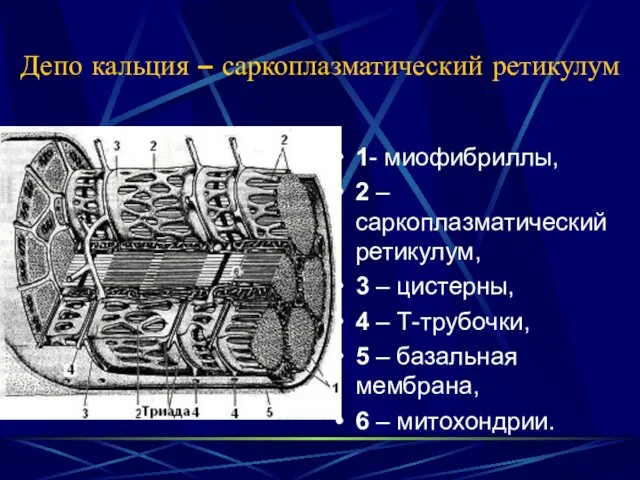
Депо кальция – саркоплазматический ретикулум
1- миофибриллы,
2 – саркоплазматический ретикулум,
Депо кальция – саркоплазматический ретикулум
1- миофибриллы,
2 – саркоплазматический ретикулум,
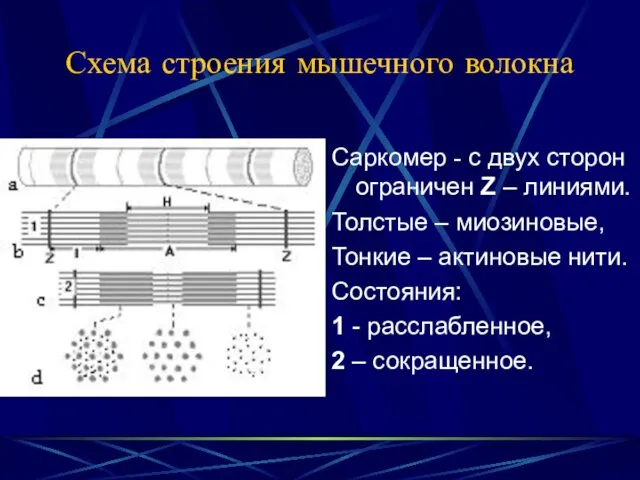
Схема строения мышечного волокна
Саркомер - с двух сторон ограничен Z –
Схема строения мышечного волокна
Саркомер - с двух сторон ограничен Z –
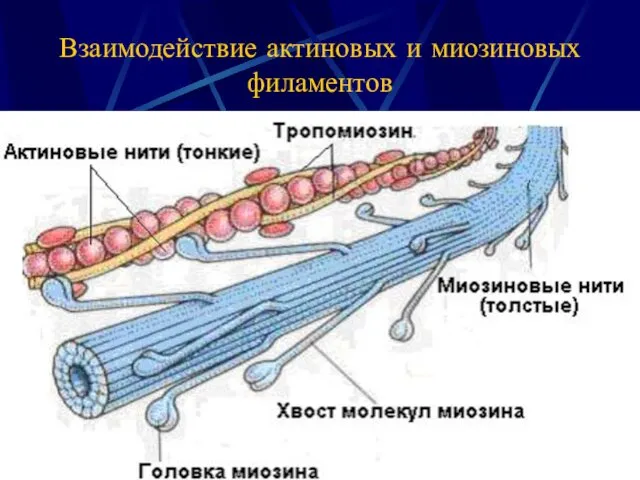
Взаимодействие актиновых и миозиновых филаментов
Взаимодействие актиновых и миозиновых филаментов
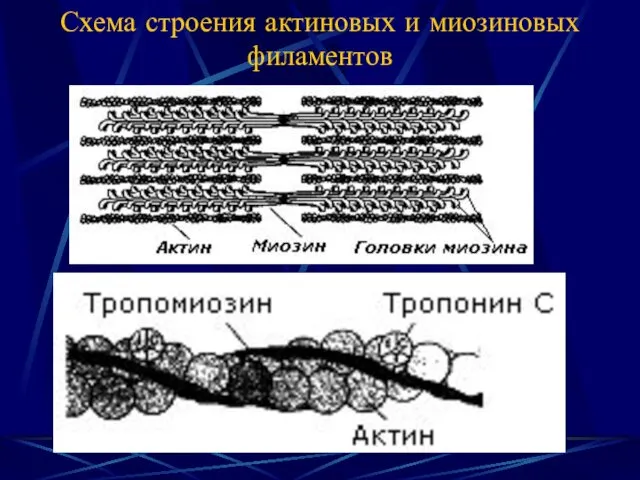
Схема строения актиновых и миозиновых филаментов
Схема строения актиновых и миозиновых филаментов

Этапы «шагового» механизма
Последовательные этапы:
а – расслабление,
б – соединение миозиновых головок с
Этапы «шагового» механизма
Последовательные этапы:
а – расслабление,
б – соединение миозиновых головок с

Кальмодулин
- идентичен тропонину С, имеющемуся в тонких нитях
Присоединяя Са2+,
Кальмодулин
- идентичен тропонину С, имеющемуся в тонких нитях
Присоединяя Са2+,

Кальций
Деполяризация мембраны цистерн открывает электровозбудимые кальциевые каналы.
В связи с тем,
Кальций
Деполяризация мембраны цистерн открывает электровозбудимые кальциевые каналы.
В связи с тем,
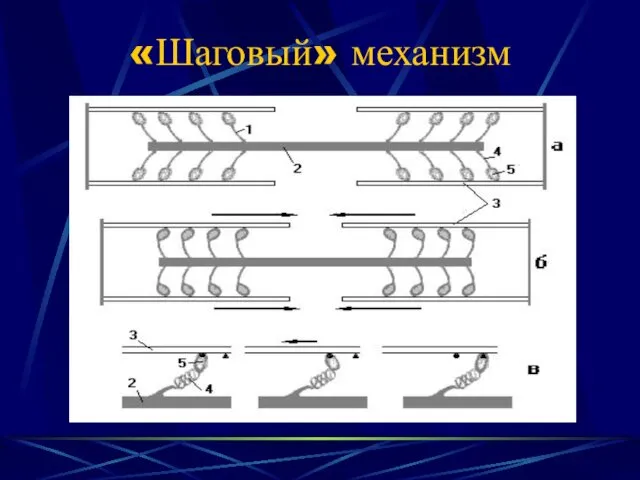
«Шаговый» механизм
«Шаговый» механизм
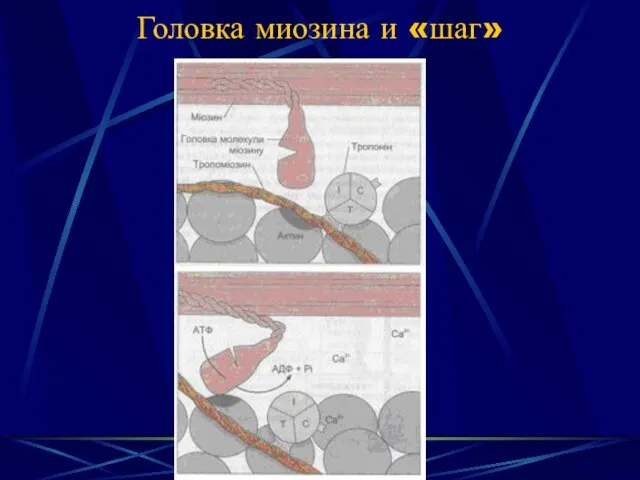
Головка миозина и «шаг»
Головка миозина и «шаг»

Роль кальция в мышечном сокращении
1 – Выброс медиатора в синаптическую щель.
2
Роль кальция в мышечном сокращении
1 – Выброс медиатора в синаптическую щель.
2
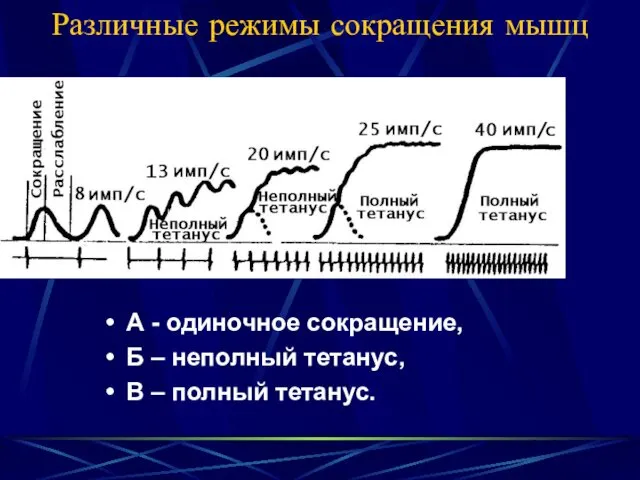
Различные режимы сокращения мышц
А - одиночное сокращение,
Б – неполный тетанус,
Различные режимы сокращения мышц
А - одиночное сокращение,
Б – неполный тетанус,
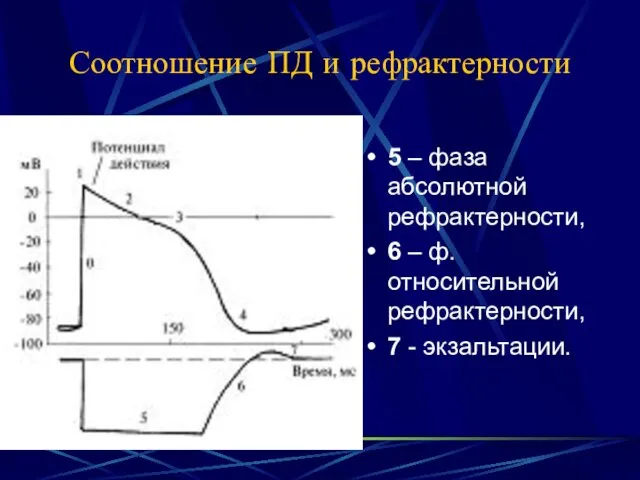
Соотношение ПД и рефрактерности
5 – фаза абсолютной рефрактерности,
6 – ф.
Соотношение ПД и рефрактерности
5 – фаза абсолютной рефрактерности,
6 – ф.

Электромиограмма
(А – одиночные ДЕ; Б – мышца в целом)
Электромиограмма
(А – одиночные ДЕ; Б – мышца в целом)

Роль АТФ в мышечном сокращении
а) сокращения (образования мостиков);
б) расслабления (разрыва
Роль АТФ в мышечном сокращении
а) сокращения (образования мостиков);
б) расслабления (разрыва

Пути ресинтеза АТФ
1) креатинфосфокиназный (КФ):
АДФ + КФ <==> АТФ
Пути ресинтеза АТФ
1) креатинфосфокиназный (КФ):
АДФ + КФ <==> АТФ

Максимальная мощность путей ресинтеза АТФ
а) фосфагенный (КФ) - 3,6 моль
Максимальная мощность путей ресинтеза АТФ
а) фосфагенный (КФ) - 3,6 моль
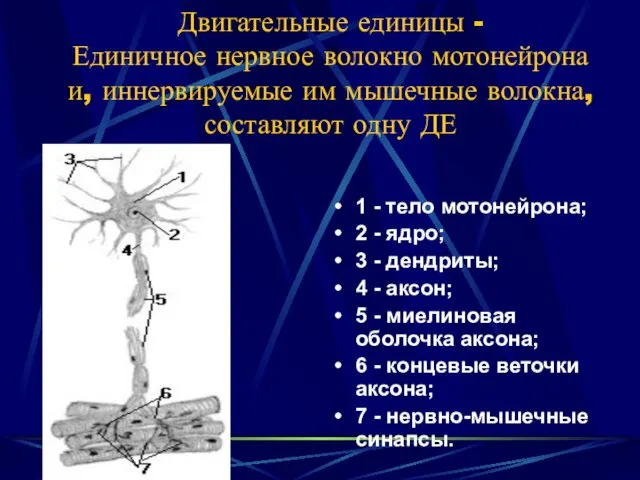
Двигательные единицы -
Единичное нервное волокно мотонейрона и, иннервируемые им мышечные волокна,
Двигательные единицы - Единичное нервное волокно мотонейрона и, иннервируемые им мышечные волокна,

Быстрые и медленные ДЕ
Быстрые
Большой мотонейрон.
Много АТФ.
Много КФ.
Активный гликолиз.
Сильные, но быстро
Быстрые и медленные ДЕ
Быстрые
Большой мотонейрон.
Много АТФ.
Много КФ.
Активный гликолиз.
Сильные, но быстро
 Современные средства поражения
Современные средства поражения Золотая Орда В Поволжье .Увек.
Золотая Орда В Поволжье .Увек. Встреча с интересными людьми.
Встреча с интересными людьми. Unicorn sight word match. Graphics by prettygrafik design
Unicorn sight word match. Graphics by prettygrafik design Роль страховой медицинской организации в повышении доступности медицинской помощи
Роль страховой медицинской организации в повышении доступности медицинской помощи Мы живём среди друзей
Мы живём среди друзей Развитие детского аутизма и аутистические черты личности
Развитие детского аутизма и аутистические черты личности Этапы компьютерного моделирования
Этапы компьютерного моделирования группа раннего развития
группа раннего развития Презентации по зимним видам спорта
Презентации по зимним видам спорта Классный час Научите свое сердце добру
Классный час Научите свое сердце добру Презентация Теоретические основы работы с детьми с задержкой психического развития
Презентация Теоретические основы работы с детьми с задержкой психического развития Учение о биосфере. (Лекция 2)
Учение о биосфере. (Лекция 2) МОЙ ЛУЧШИЙ ДРУГ - ПАПА
МОЙ ЛУЧШИЙ ДРУГ - ПАПА Бабушкин сундук ( Әбиемнең сандыгы)
Бабушкин сундук ( Әбиемнең сандыгы) Жаңадан шыққан техникалар
Жаңадан шыққан техникалар Скакалочка. Игра
Скакалочка. Игра Чтение №104. Сказки А. С. Пушкина
Чтение №104. Сказки А. С. Пушкина Методические рекомендации по организации выполнения и защиты, оформлению выпускной квалификационной работы
Методические рекомендации по организации выполнения и защиты, оформлению выпускной квалификационной работы Классный час Мое Приморье
Классный час Мое Приморье Устная часть ЕГЭ по английскому языку
Устная часть ЕГЭ по английскому языку Праздники народов россии
Праздники народов россии Профилактика инфекционных болезней и эпидемий. Карантинные инфекции и особо опасные инфекции
Профилактика инфекционных болезней и эпидемий. Карантинные инфекции и особо опасные инфекции Лямблиоз у детей
Лямблиоз у детей Работа с ритмом на уроке сольфеджио
Работа с ритмом на уроке сольфеджио Развитие мелкой моторики через различные виды деятельности
Развитие мелкой моторики через различные виды деятельности Дослідження та удосконалення САК температурним режимом зерносушарки на хлібокомбінаті
Дослідження та удосконалення САК температурним режимом зерносушарки на хлібокомбінаті Пожарная безопасность для дошкольников 5-7 лет
Пожарная безопасность для дошкольников 5-7 лет