- Функциональная морфология цитоскелета. Функциональная морфология ядра клетки

Содержание
- 2. Цитоскелет трехмерная, лабильная система, состоящая из: микротрубочек; микрофиламентов; промежуточных филаментов.
- 3. Микротрубочки элементы цитоскелета, представляющие собой полые цилиндрические трубки. Dвнеш. 25 нм. Dвн. 15 нм Стенка состоит
- 4. Лабильность микротрубочек «-» конец связан с белками ЦОМТ. «+» конец свободный, участвует в процессах полимеризации/ деполимеризации.
- 5. Белки, ассоциированные с микротрубочками (Microtubule-associated protein, MAP) белки, постоянно связанные с микротрубочками и служащие для: направления
- 6. Функции микротрубочек Поддержание формы клетки Обеспечение внутриклеточного транспорта Формирование органелл Обеспечение подвижности клетки Формирования веретена деления
- 7. Клеточный центр (центросома) в 1888 г.Теодор Бовери - универсальный немембранный органоид всех эукариотических клеток, органелла клеточного
- 8. Ценросома (животные, некоторые грибы и простейшие) Диплосома Центросфера Центросома сумчатых и базидиевых грибов, высших растений не
- 9. Диплосома Дуплет центриолей располагаются под прямым углом друг к другу. Обе центриоли сближены своими проксимальными концами.
- 10. Центросфера Сателлиты/гало Микротрубочки Фокусы схождения микротрубочек Исчерченный корешок Сателлиты – ЦОМТ, состоящие из белковой головки и
- 11. Строение центриоли Каждый триплет связан с соседним и с центральной фибриллой. Центриоли имеют белковый каркас, в
- 12. Центросомный цикл М-фаза: две диплосомы на полюсах клетки, от них отходят нити ахроматинового веретена деления. Материнская
- 13. Центр организации микротрубочек Центросомные ЦОМТ. ЦОМТ без определенной локализации. Микротрубочки могут образовываться в цитозоле и вне
- 14. Первичные (неподвижные) реснички Все типы клеток (кроме клеток крови, мышечных клеток) в G0-периоде формируют первичные реснички.
- 15. Кинетоцилии (подвижные реснички) У эукариот представляют собой выросты цитоплазмы, окруженные плазмолеммой и содержащие аксонему. Длина реснички=2-10
- 16. Аксонема девять дублетов, связанных между собой динеином; две центральные микротрубочки; центральная белковая муфта; Спицы.
- 17. Базальное тельце (кинетосома) Состоит из 9 триплетов микротрубочек, имеет центральную белковую фибриллу и спицы идущие к
- 18. Микрофиламенты - белковые нити (D 5-7 нм), формирующие в цитоплазме трех мерную сеть и концентрирующиеся под
- 19. Белки микрофиламентов Актин – глобулярный мономерный белок (G-актин), способный к полимеризации (F-актин). Молекула актина имеет вид
- 20. Актин-связывающие белки Белки, ингибирующие полимеризацию актина (профиллин, ДНКазаI); стабилизирующие белки (тропомиозины); кэпактины (гельзолин/вилин,фрагмин/северин) белки, сшивающие актиновые
- 21. Микроворсинки выросты цитоплазмы, окруженные плазмолеммой, каркас которых образован пучком микрофиламентами. У основания микроворсинки пучок микрофиламентов переходит
- 22. Стереоцилии длинные (50 мкм), неподвижные, ветвящиеся микроворсинки, выполняющие рецепторную функцию: в сенсорном эпителии внутреннего уха; в
- 23. Функции микрофиламентов Обеспечение сокращения клеток Обеспечение движения клеток. Обеспечение мембранных функций. Перемещение в цитозоле органелл, макромолекул.
- 24. Промежуточные филаменты элементы цитоскелета, образованные различными фибриллярными белками. D 10 нм. Формируют трехмерную стабильную сеть.
- 25. Строение промежуточных филаментов Димер Тетрамер Протофиламент Волокно
- 26. Классы промежуточных филаментов Тонофиламенты (кератины); Десминовые филаменты (десмин); Виментиновые филаменты (виментин); Нейрофиламенты (NF-L, NF-M, NF-H); Глиальные
- 27. Функции промежуточных филаментов Поддержание формы клетки; Распределение органелл в цитоплазме; Формирование рогового вещества; Формирование остова отростков
- 28. Микротрабекулы система тонких белковых нитей, пересекающих цитоплазму в различных направлениях. D 2 нм. С микротрабекулярной системой
- 29. Клеточное ядро - основной компонент эукариотической клетки, содержащий её генетический материал. Функции ядра: Хранение генетической информации.
- 30. Компоненты ядра Кариолемма Хроматин/хромосомы Ядрышко Кариоплазма
- 31. Кариолемма (кариотека) Оболочка ядра, состоящая из наружной и внутренней мембран, разделенных перинуклеарным пространством (15-40 нм). Образована
- 32. Наружная мембрана кариолеммы На поверхности имеются рибосомы, перинуклеарное пространство сообщается с люменом ЭПР.
- 33. Внутренняя мембрана кариолеммы ядерная ламина (80-300 нм) - структура, образованная белками-ламинами, к которой прикреплены нити хроматина.
- 34. Функции ламины Поддерживает форму ядра. Участвует в формировании порового комплекса. Отвечает за упорядоченное расположение хроматина в
- 35. Хромосомная территория - хроматин каждой хромосомы прикреплен в определенном месте к внутренней мембране кариолеммы.
- 36. области перехода внутренней мембраны кариолеммы в наружную. В поре расположен комплекс ядерной поры: Два параллельных белковых
- 37. Комплекс ядерной поры Образован - нуклеопоринами
- 38. Функция ядерных пор Обеспечение ядерно-цитоплазматического взаимодействия.
- 39. Хроматин комплекс ДНК и белков интерфазного ядра, представляющий деспирализованные хромосомы. Гетерохроматин: а. Облигатный; б. Факультативный; Эухроматин;
- 40. Уровни компактизации хроматина Нуклеосомный Нуклеомерный Хромомерный Хромонемный Хроматидный
- 41. Строение нуклеосомы Гистоны - основные белки, связывающиеся с ДНК (арг, лиз).10-20кD. (Н2А, Н2В, Н3, Н4)*2 образуют
- 42. Нуклеосомная нить Свободная ДНК между смежными нуклеосомами – линкер (≈ 50 п.н.). Более 90% ДНК присутствует
- 43. Хроматиновая фибрилла Спираль из нуклеосомной нити (1 виток содержит 6 нуклеосом). Толщина фибриллы 30 нм.
- 44. Петельные домены Нити хроматина в хромомерах имеют укладку в виде розеток, собранных у основания, от которого
- 45. Негистоновые белки Большое влияние на структуру хроматина и функционирование эукариотических генов оказывают различные негистоновые белки. Белки
- 46. Упаковка внеядерной ДНК
- 47. Метафазная хромосома Центромера Теломера Хроматида Плечи (р и q) В области центромеры расположен кинетохор - мультибелковый
- 48. Сателлит — это округлое или удлинённое тельце, отделённое от основной части хромосомы вторичной перетяжкой.
- 49. Теломерный участок хромосомы - концевой участок хромосомы, препятствующий соединению хромосомы с другими хромосомами, их фрагментами или
- 50. Типы хромосом телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце); акроцентрические (палочковидные хромосомы с очень
- 51. Кариотип Совокупность признаков (числа, величины и морфологии) полного набора хромосом, присущий клеткам данного биологического вида (видовой
- 52. Геном - тотальная ДНК гаплоидного набора хромосом и внехромосомных генетических элементов, содержащаяся в отдельной клетке многоклеточного
- 53. Общие принципы организации генома
- 54. Организация генома ДНК Е. coli 3,8*106 п.н. (≈3000 генов). ДНК Н. sapiens 3,5*109 (≈31000 генов).
- 55. Семейство, тандем и кластер генов Семейство – группа функционально родственных генов, имеющих сходную структуру и общее
- 56. Организация генетического материала у прокариот
- 57. Организация генетического материала у эукариот
- 58. Тельце Барра Скопление гетерохроматина, соответствующее одной Х-хромосоме у осбей женского пола, которая не деконденсируется даже в
- 59. Микроядра - фрагменты хромосом или целые хромосомы, не включенные в состав ядра после митотического деления клетки.
- 60. Ядрышко (нуклеоль) у человека образуются из хроматина 13-15 и 21, 22 хромосом - структура ядра, сформированная
- 61. Размеры и количество ядрышек В интерфазном ядре на светооптическом уровне – плотная гранула (D 1-3 мкм),
- 62. Структура ядрышка Фибриллярный компонент - внутренняя часть ядрышка, состоит из нитей хроматина и первичных транскриптов рРНК.
- 63. Ядерный матрикс Кариоскелет построен из негистоновых белков, формирующих разветвленную сеть, взаимодействующую с ядерной ламиной. В клетке
- 65. Скачать презентацию
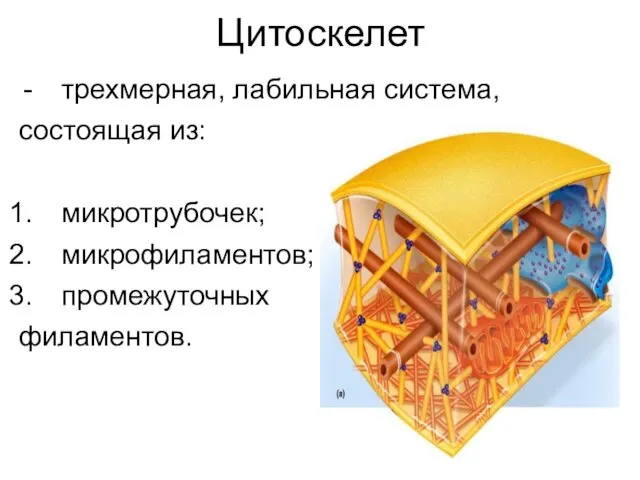
Цитоскелет
трехмерная, лабильная система,
состоящая из:
микротрубочек;
микрофиламентов;
промежуточных
филаментов.
Цитоскелет
трехмерная, лабильная система,
состоящая из:
микротрубочек;
микрофиламентов;
промежуточных
филаментов.
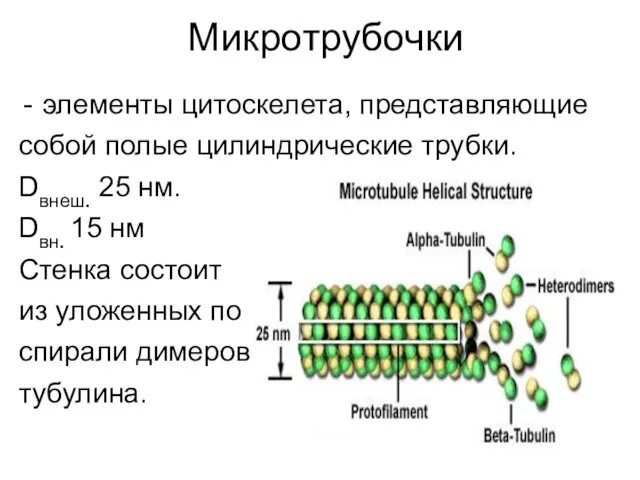
Микротрубочки
элементы цитоскелета, представляющие
собой полые цилиндрические трубки.
Dвнеш. 25 нм.
Dвн. 15 нм
Стенка
Микротрубочки
элементы цитоскелета, представляющие
собой полые цилиндрические трубки.
Dвнеш. 25 нм.
Dвн. 15 нм
Стенка
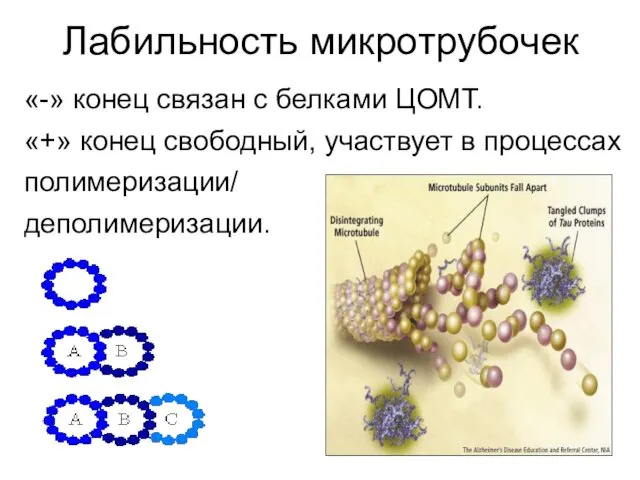
Лабильность микротрубочек
«-» конец связан с белками ЦОМТ.
«+» конец свободный, участвует в
Лабильность микротрубочек
«-» конец связан с белками ЦОМТ.
«+» конец свободный, участвует в

Белки, ассоциированные с микротрубочками
(Microtubule-associated protein, MAP)
белки, постоянно связанные с микротрубочками
Белки, ассоциированные с микротрубочками
(Microtubule-associated protein, MAP)
белки, постоянно связанные с микротрубочками

Функции микротрубочек
Поддержание формы клетки
Обеспечение внутриклеточного транспорта
Формирование органелл
Обеспечение подвижности клетки
Формирования веретена
Функции микротрубочек
Поддержание формы клетки
Обеспечение внутриклеточного транспорта
Формирование органелл
Обеспечение подвижности клетки
Формирования веретена

Клеточный центр (центросома)
в 1888 г.Теодор Бовери
- универсальный немембранный органоид
Клеточный центр (центросома)
в 1888 г.Теодор Бовери
- универсальный немембранный органоид
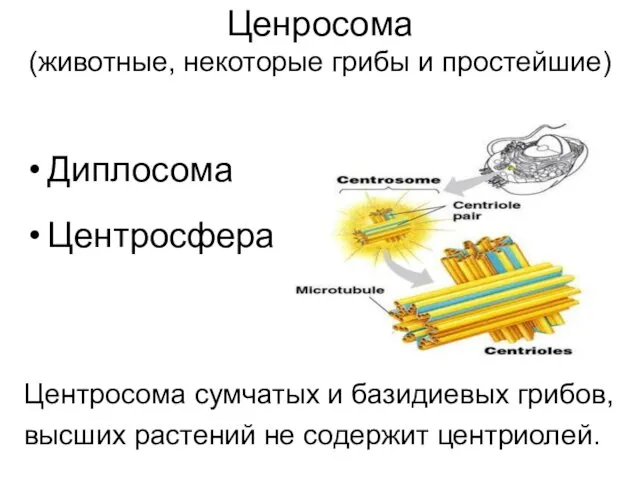
Ценросома
(животные, некоторые грибы и простейшие)
Диплосома
Центросфера
Центросома сумчатых и базидиевых грибов,
Ценросома
(животные, некоторые грибы и простейшие)
Диплосома
Центросфера
Центросома сумчатых и базидиевых грибов,

Диплосома
Дуплет центриолей располагаются под прямым углом друг к другу. Обе центриоли
Диплосома
Дуплет центриолей располагаются под прямым углом друг к другу. Обе центриоли

Центросфера
Сателлиты/гало
Микротрубочки
Фокусы схождения микротрубочек
Исчерченный корешок
Сателлиты – ЦОМТ, состоящие из белковой головки
Центросфера
Сателлиты/гало
Микротрубочки
Фокусы схождения микротрубочек
Исчерченный корешок
Сателлиты – ЦОМТ, состоящие из белковой головки
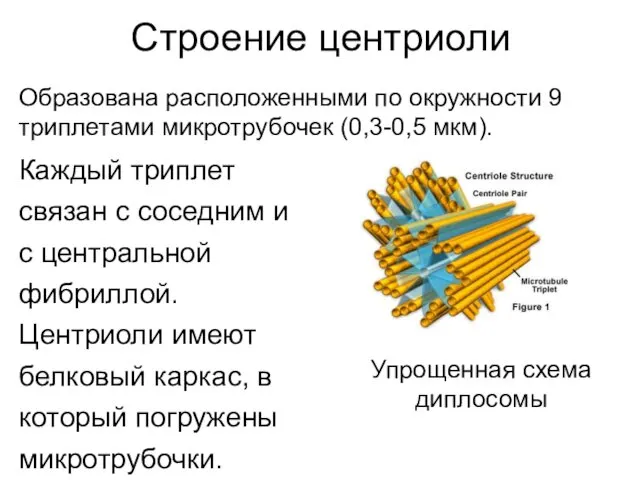
Строение центриоли
Каждый триплет
связан с соседним и
с центральной
фибриллой.
Центриоли имеют
Строение центриоли
Каждый триплет
связан с соседним и
с центральной
фибриллой.
Центриоли имеют

Центросомный цикл
М-фаза: две диплосомы на полюсах клетки, от них отходят нити
Центросомный цикл
М-фаза: две диплосомы на полюсах клетки, от них отходят нити
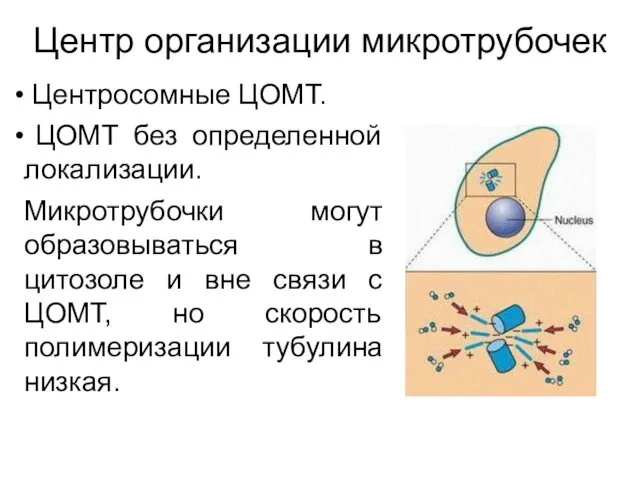
Центр организации микротрубочек
Центросомные ЦОМТ.
ЦОМТ без определенной локализации.
Микротрубочки могут образовываться
Центр организации микротрубочек
Центросомные ЦОМТ.
ЦОМТ без определенной локализации.
Микротрубочки могут образовываться
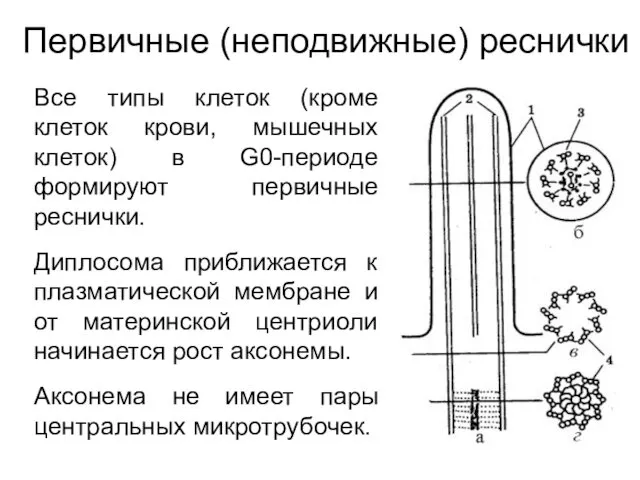
Первичные (неподвижные) реснички
Все типы клеток (кроме клеток крови, мышечных клеток) в
Первичные (неподвижные) реснички
Все типы клеток (кроме клеток крови, мышечных клеток) в
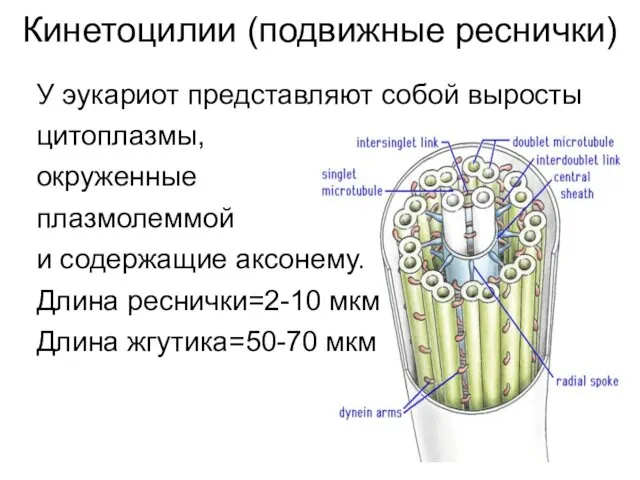
Кинетоцилии (подвижные реснички)
У эукариот представляют собой выросты
цитоплазмы,
окруженные
плазмолеммой
и
Кинетоцилии (подвижные реснички)
У эукариот представляют собой выросты
цитоплазмы,
окруженные
плазмолеммой
и
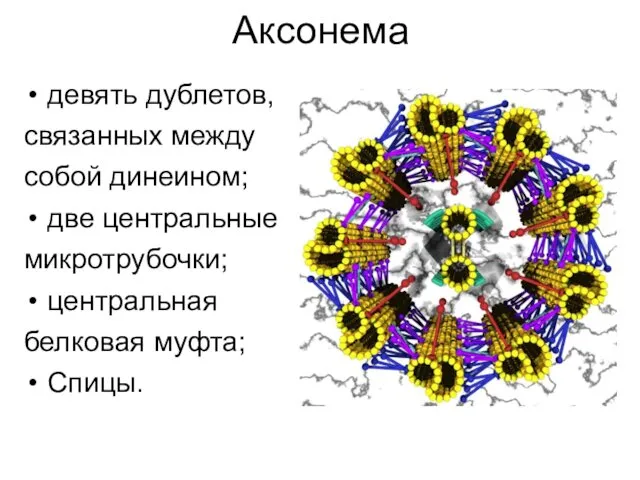
Аксонема
девять дублетов,
связанных между
собой динеином;
две центральные
микротрубочки;
центральная
белковая муфта;
Спицы.
Аксонема
девять дублетов,
связанных между
собой динеином;
две центральные
микротрубочки;
центральная
белковая муфта;
Спицы.

Базальное тельце (кинетосома)
Состоит из 9 триплетов микротрубочек, имеет центральную
белковую фибриллу
Базальное тельце (кинетосома)
Состоит из 9 триплетов микротрубочек, имеет центральную
белковую фибриллу
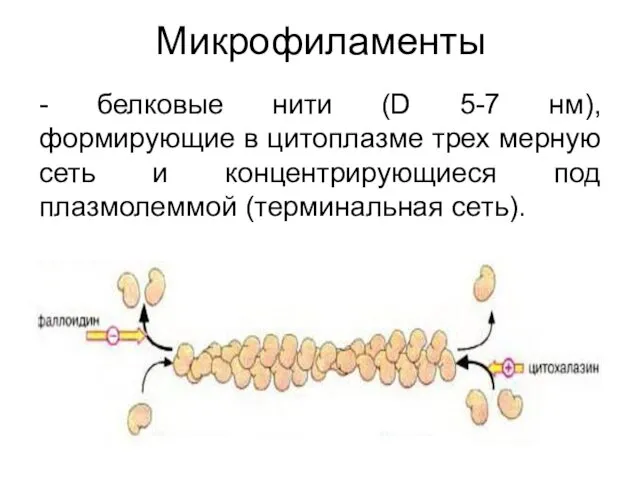
Микрофиламенты
- белковые нити (D 5-7 нм), формирующие в цитоплазме трех мерную
Микрофиламенты
- белковые нити (D 5-7 нм), формирующие в цитоплазме трех мерную
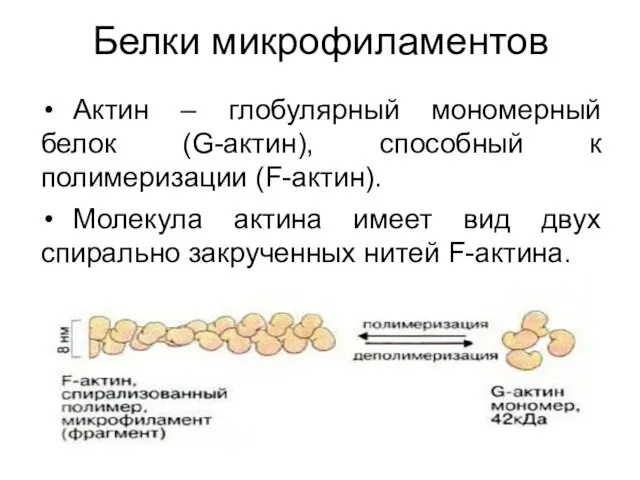
Белки микрофиламентов
Актин – глобулярный мономерный белок (G-актин), способный к полимеризации
Белки микрофиламентов
Актин – глобулярный мономерный белок (G-актин), способный к полимеризации

Актин-связывающие белки
Белки, ингибирующие полимеризацию актина
(профиллин, ДНКазаI);
стабилизирующие белки (тропомиозины);
Актин-связывающие белки
Белки, ингибирующие полимеризацию актина
(профиллин, ДНКазаI);
стабилизирующие белки (тропомиозины);
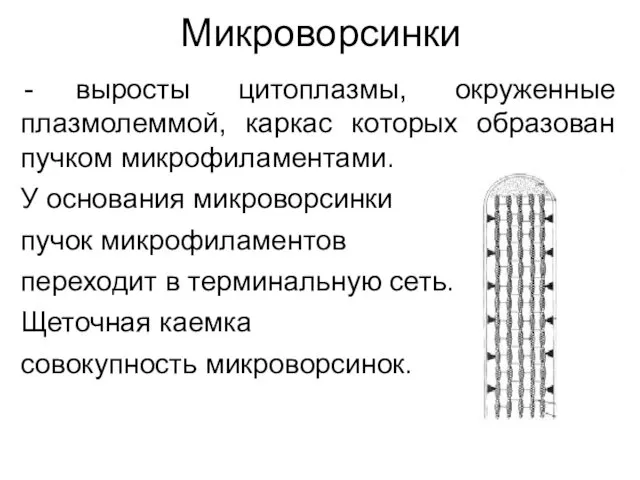
Микроворсинки
выросты цитоплазмы, окруженные плазмолеммой, каркас которых образован пучком микрофиламентами.
У основания
Микроворсинки
выросты цитоплазмы, окруженные плазмолеммой, каркас которых образован пучком микрофиламентами.
У основания
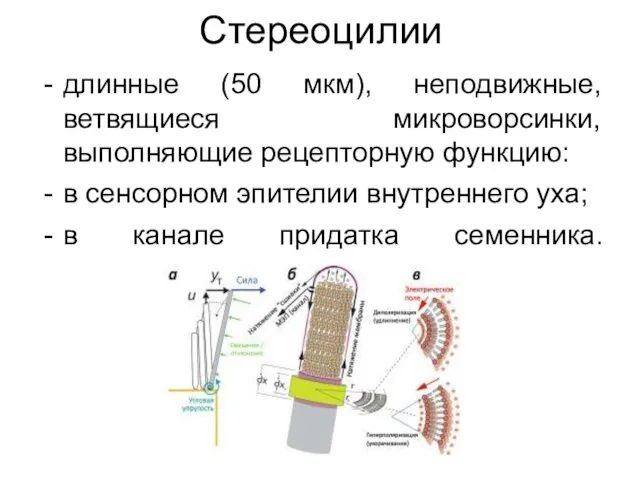
Стереоцилии
длинные (50 мкм), неподвижные, ветвящиеся микроворсинки, выполняющие рецепторную функцию:
в сенсорном эпителии
Стереоцилии
длинные (50 мкм), неподвижные, ветвящиеся микроворсинки, выполняющие рецепторную функцию:
в сенсорном эпителии

Функции микрофиламентов
Обеспечение сокращения клеток
Обеспечение движения клеток.
Обеспечение мембранных функций.
Перемещение в цитозоле органелл,
Функции микрофиламентов
Обеспечение сокращения клеток
Обеспечение движения клеток.
Обеспечение мембранных функций.
Перемещение в цитозоле органелл,
Промежуточные филаменты
элементы цитоскелета, образованные
различными фибриллярными белками.
D 10 нм.
Формируют трехмерную стабильную сеть.
Промежуточные филаменты
элементы цитоскелета, образованные
различными фибриллярными белками.
D 10 нм.
Формируют трехмерную стабильную сеть.
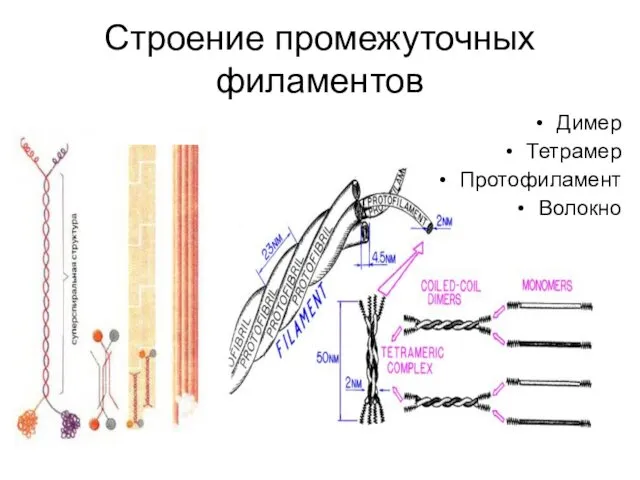
Строение промежуточных филаментов
Димер
Тетрамер
Протофиламент
Волокно
Строение промежуточных филаментов
Димер
Тетрамер
Протофиламент
Волокно

Классы промежуточных филаментов
Тонофиламенты (кератины);
Десминовые филаменты (десмин);
Виментиновые филаменты (виментин);
Нейрофиламенты (NF-L, NF-M, NF-H);
Глиальные
Классы промежуточных филаментов
Тонофиламенты (кератины);
Десминовые филаменты (десмин);
Виментиновые филаменты (виментин);
Нейрофиламенты (NF-L, NF-M, NF-H);
Глиальные

Функции промежуточных филаментов
Поддержание формы клетки;
Распределение органелл в цитоплазме;
Формирование рогового вещества;
Формирование остова
Функции промежуточных филаментов
Поддержание формы клетки;
Распределение органелл в цитоплазме;
Формирование рогового вещества;
Формирование остова
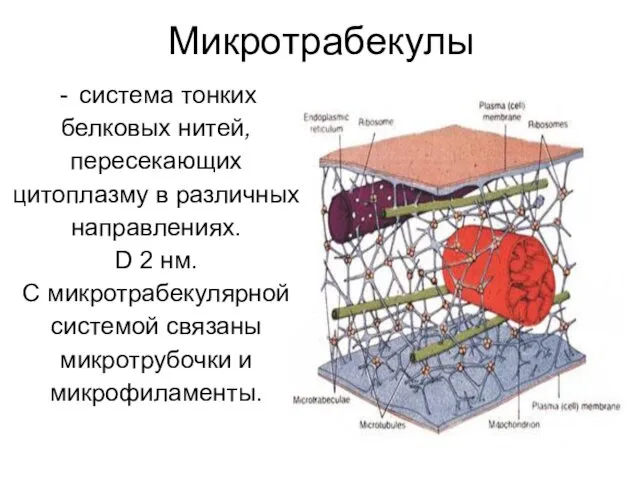
Микротрабекулы
система тонких
белковых нитей,
пересекающих
цитоплазму в различных
направлениях.
D 2
Микротрабекулы
система тонких
белковых нитей,
пересекающих
цитоплазму в различных
направлениях.
D 2
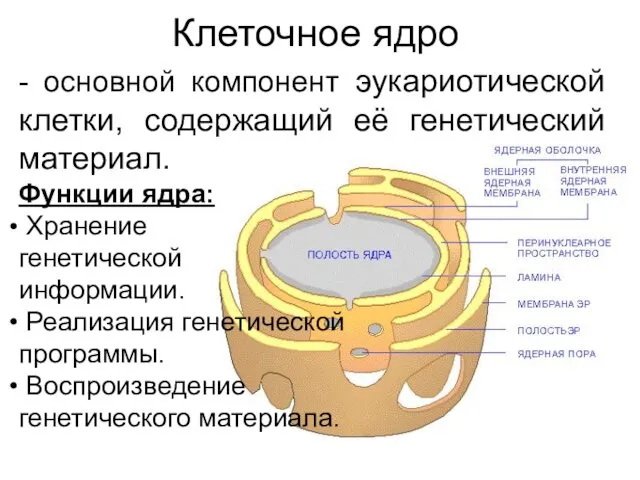
Клеточное ядро
- основной компонент эукариотической клетки, содержащий её генетический материал.
Функции ядра:
Клеточное ядро
- основной компонент эукариотической клетки, содержащий её генетический материал.
Функции ядра:
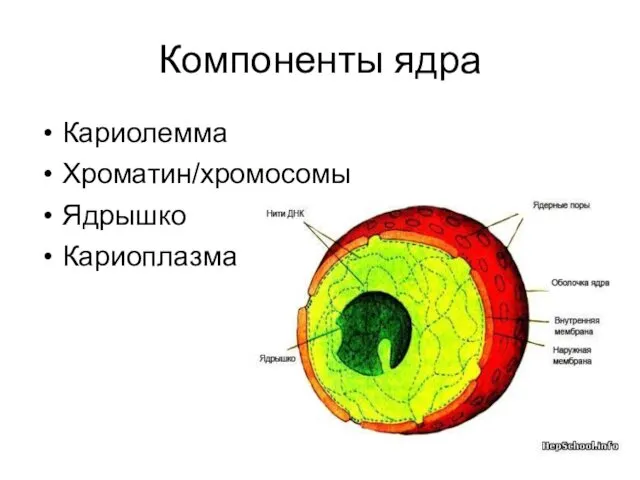
Компоненты ядра
Кариолемма
Хроматин/хромосомы
Ядрышко
Кариоплазма
Компоненты ядра
Кариолемма
Хроматин/хромосомы
Ядрышко
Кариоплазма
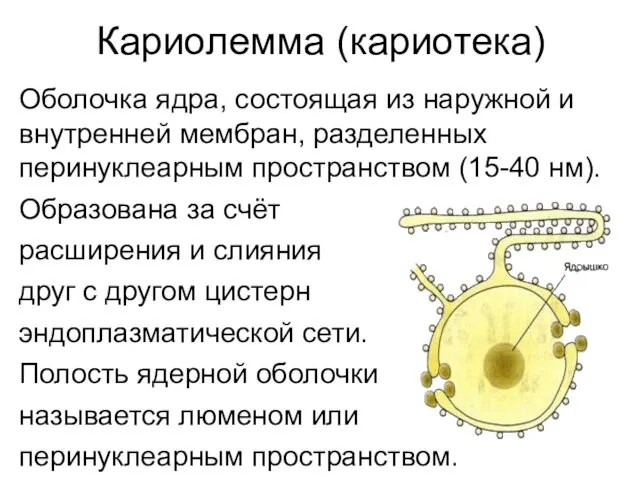
Кариолемма (кариотека)
Оболочка ядра, состоящая из наружной и внутренней мембран, разделенных перинуклеарным
Кариолемма (кариотека)
Оболочка ядра, состоящая из наружной и внутренней мембран, разделенных перинуклеарным
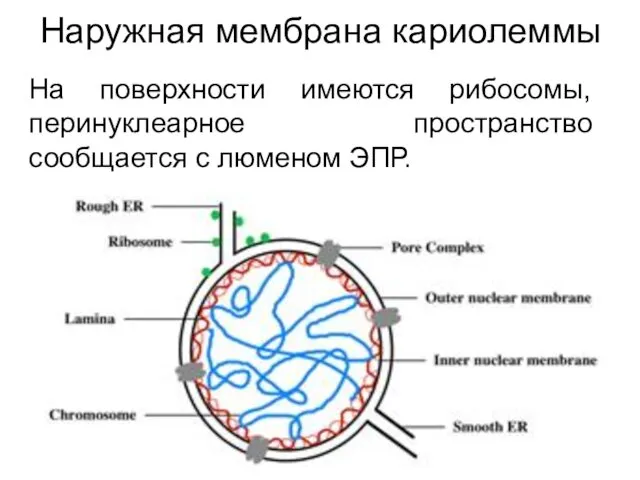
Наружная мембрана кариолеммы
На поверхности имеются рибосомы, перинуклеарное пространство сообщается с люменом
Наружная мембрана кариолеммы
На поверхности имеются рибосомы, перинуклеарное пространство сообщается с люменом
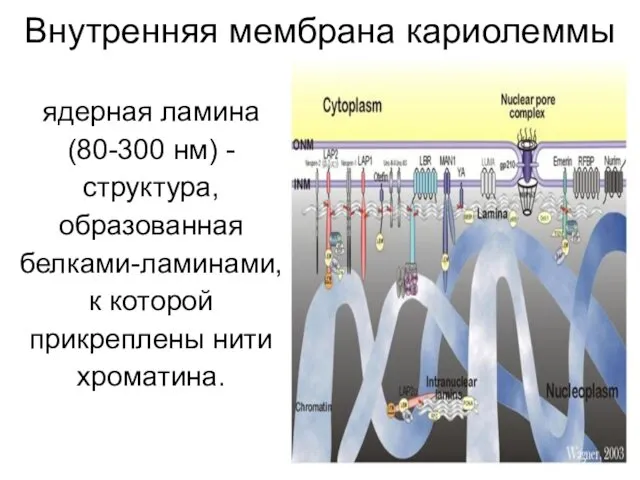
Внутренняя мембрана кариолеммы
ядерная ламина
(80-300 нм) -
структура,
образованная
белками-ламинами,
к которой
Внутренняя мембрана кариолеммы
ядерная ламина
(80-300 нм) -
структура,
образованная
белками-ламинами,
к которой

Функции ламины
Поддерживает форму ядра.
Участвует в формировании порового комплекса.
Отвечает за упорядоченное расположение
Функции ламины
Поддерживает форму ядра.
Участвует в формировании порового комплекса.
Отвечает за упорядоченное расположение
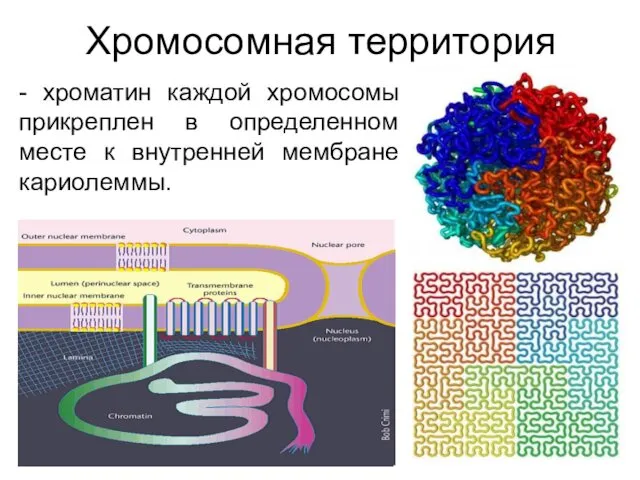
Хромосомная территория
- хроматин каждой хромосомы прикреплен в определенном месте к внутренней
Хромосомная территория
- хроматин каждой хромосомы прикреплен в определенном месте к внутренней

области перехода внутренней мембраны
кариолеммы в наружную.
В поре расположен комплекс ядерной
области перехода внутренней мембраны
кариолеммы в наружную.
В поре расположен комплекс ядерной
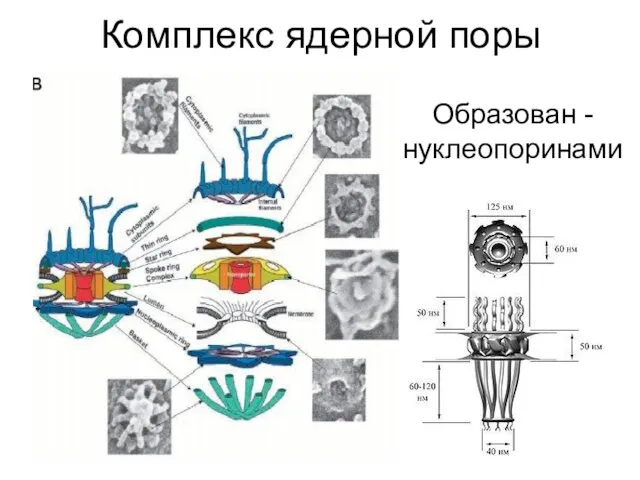
Комплекс ядерной поры
Образован - нуклеопоринами
Комплекс ядерной поры
Образован - нуклеопоринами
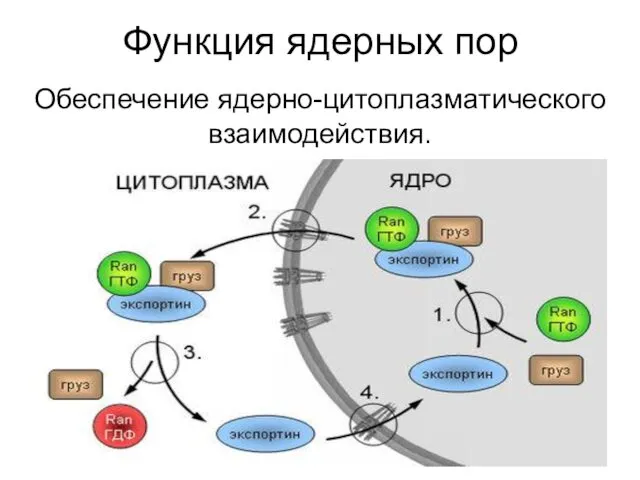
Функция ядерных пор
Обеспечение ядерно-цитоплазматического взаимодействия.
Функция ядерных пор
Обеспечение ядерно-цитоплазматического взаимодействия.
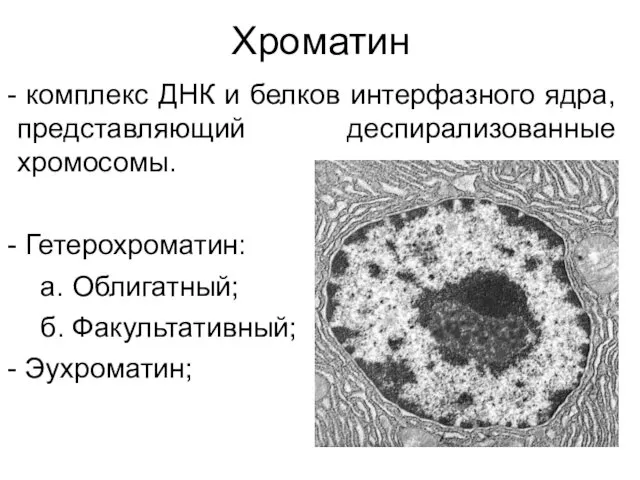
Хроматин
комплекс ДНК и белков интерфазного ядра, представляющий деспирализованные хромосомы.
Гетерохроматин:
Хроматин
комплекс ДНК и белков интерфазного ядра, представляющий деспирализованные хромосомы.
Гетерохроматин:
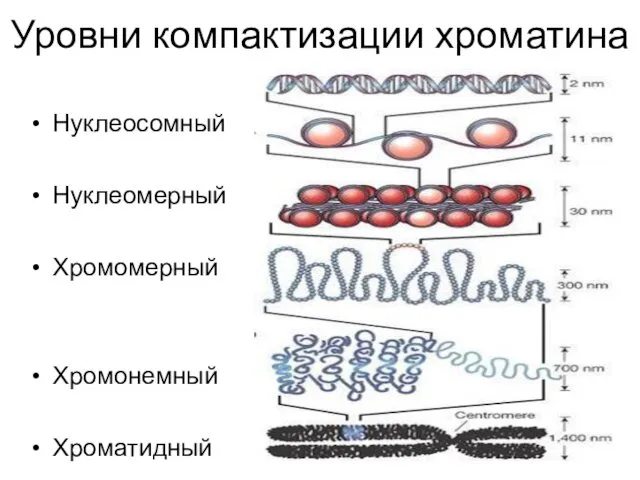
Уровни компактизации хроматина
Нуклеосомный
Нуклеомерный
Хромомерный
Хромонемный
Хроматидный
Уровни компактизации хроматина
Нуклеосомный
Нуклеомерный
Хромомерный
Хромонемный
Хроматидный
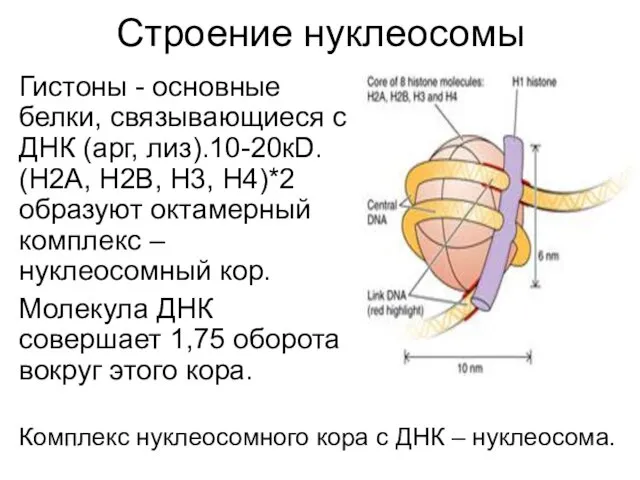
Строение нуклеосомы
Гистоны - основные белки, связывающиеся с ДНК (арг, лиз).10-20кD. (Н2А,
Строение нуклеосомы
Гистоны - основные белки, связывающиеся с ДНК (арг, лиз).10-20кD. (Н2А,
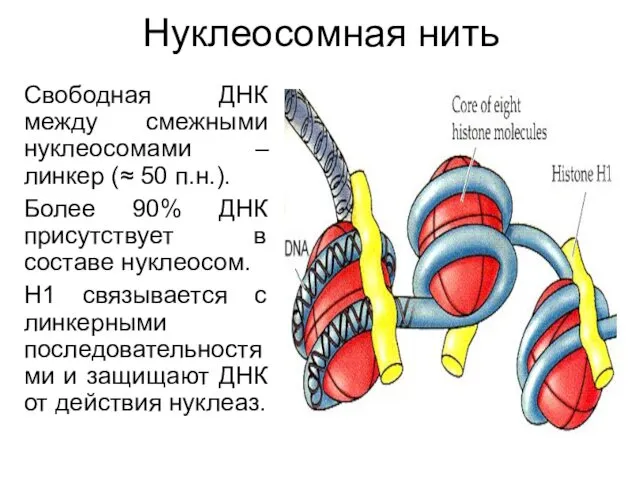
Нуклеосомная нить
Свободная ДНК между смежными нуклеосомами – линкер (≈ 50 п.н.).
Более
Нуклеосомная нить
Свободная ДНК между смежными нуклеосомами – линкер (≈ 50 п.н.).
Более
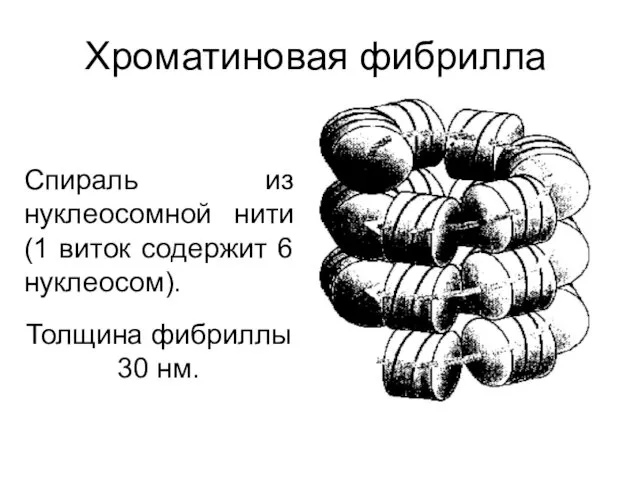
Хроматиновая фибрилла
Спираль из нуклеосомной нити (1 виток содержит 6 нуклеосом).
Толщина фибриллы
Хроматиновая фибрилла
Спираль из нуклеосомной нити (1 виток содержит 6 нуклеосом).
Толщина фибриллы
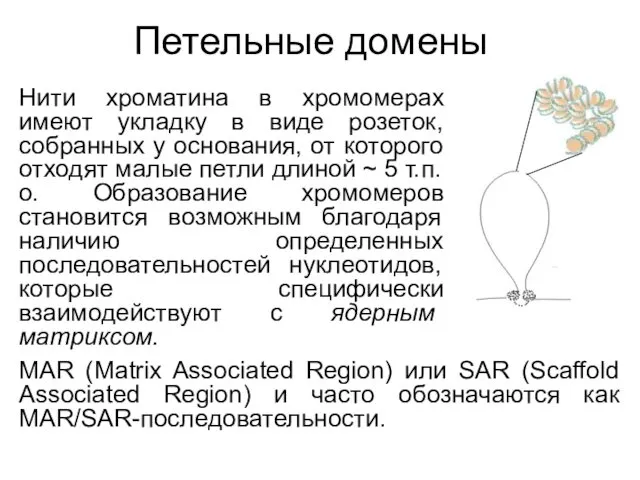
Петельные домены
Нити хроматина в хромомерах имеют укладку в виде розеток, собранных
Петельные домены
Нити хроматина в хромомерах имеют укладку в виде розеток, собранных

Негистоновые белки
Большое влияние на структуру хроматина и функционирование эукариотических генов оказывают
Негистоновые белки
Большое влияние на структуру хроматина и функционирование эукариотических генов оказывают
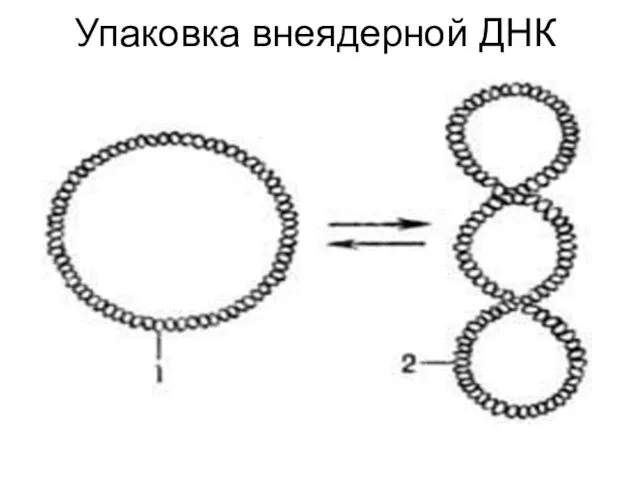
Упаковка внеядерной ДНК
Упаковка внеядерной ДНК
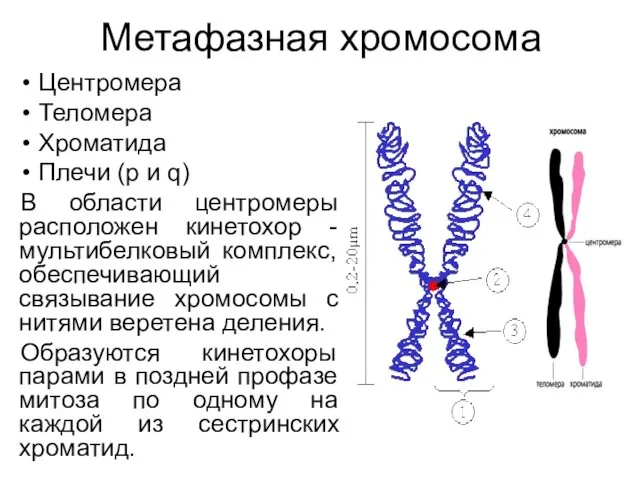
Метафазная хромосома
Центромера
Теломера
Хроматида
Плечи (р и q)
В области центромеры
Метафазная хромосома
Центромера
Теломера
Хроматида
Плечи (р и q)
В области центромеры
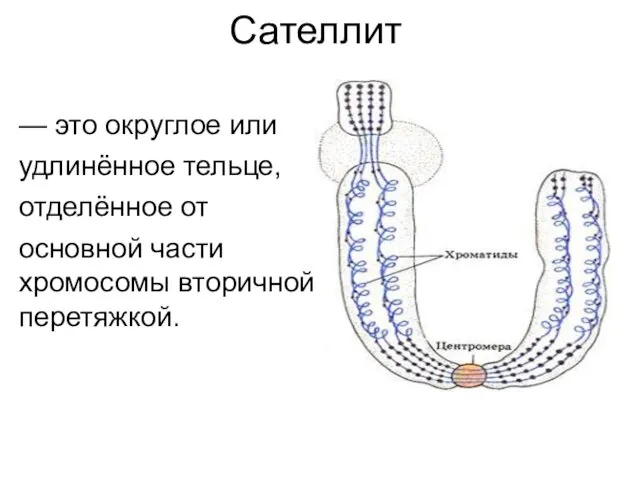
Сателлит
— это округлое или
удлинённое тельце,
отделённое от
основной части хромосомы вторичной
Сателлит
— это округлое или
удлинённое тельце,
отделённое от
основной части хромосомы вторичной
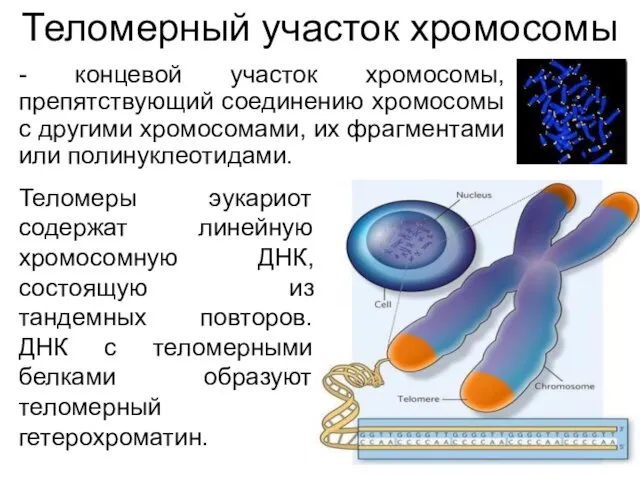
Теломерный участок хромосомы
- концевой участок хромосомы, препятствующий соединению хромосомы с другими
Теломерный участок хромосомы
- концевой участок хромосомы, препятствующий соединению хромосомы с другими

Типы хромосом
телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце);
Типы хромосом
телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце);
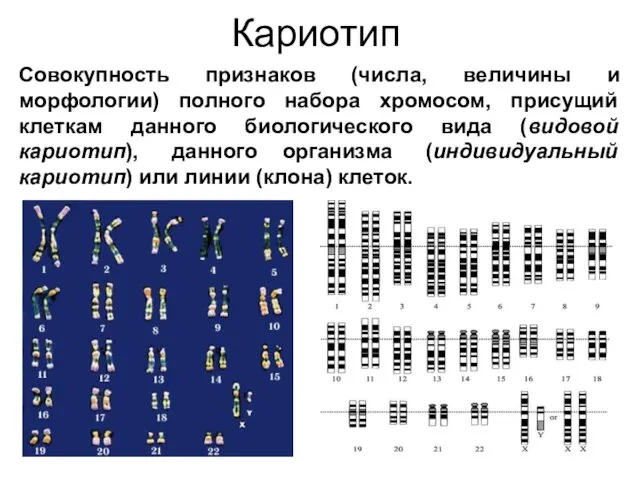
Кариотип
Совокупность признаков (числа, величины и морфологии) полного набора хромосом, присущий клеткам
Кариотип
Совокупность признаков (числа, величины и морфологии) полного набора хромосом, присущий клеткам
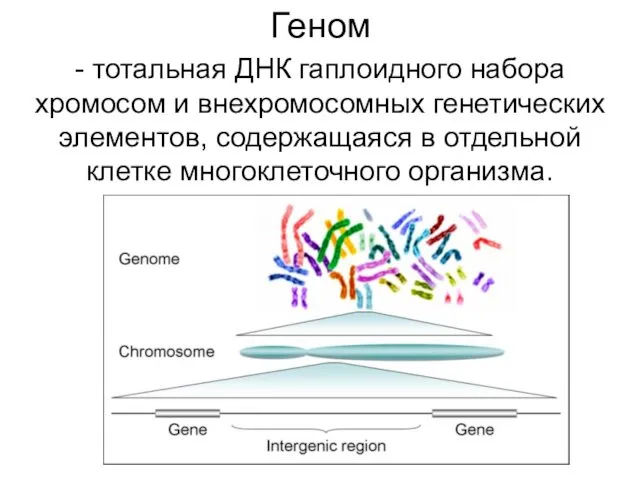
Геном
- тотальная ДНК гаплоидного набора хромосом и внехромосомных генетических элементов, содержащаяся
Геном
- тотальная ДНК гаплоидного набора хромосом и внехромосомных генетических элементов, содержащаяся

Общие принципы организации генома
Общие принципы организации генома
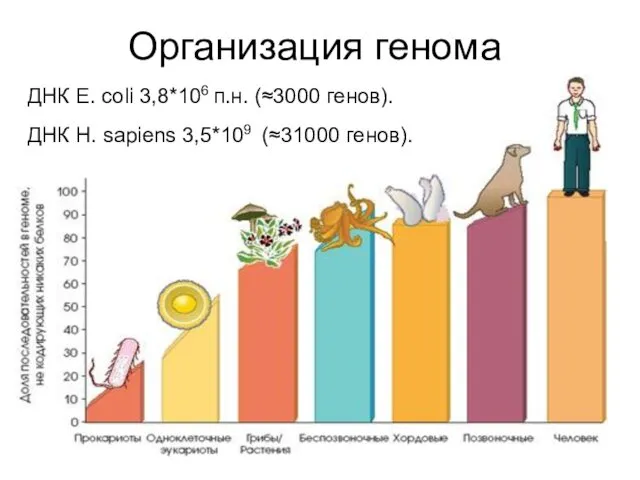
Организация генома
ДНК Е. coli 3,8*106 п.н. (≈3000 генов).
ДНК Н. sapiens 3,5*109
Организация генома
ДНК Е. coli 3,8*106 п.н. (≈3000 генов).
ДНК Н. sapiens 3,5*109

Семейство, тандем и кластер генов
Семейство – группа функционально родственных генов, имеющих
Семейство, тандем и кластер генов
Семейство – группа функционально родственных генов, имеющих

Организация генетического материала у прокариот
Организация генетического материала у прокариот

Организация генетического материала у эукариот
Организация генетического материала у эукариот

Тельце Барра
Скопление гетерохроматина, соответствующее одной Х-хромосоме у осбей женского пола, которая
Тельце Барра
Скопление гетерохроматина, соответствующее одной Х-хромосоме у осбей женского пола, которая

Микроядра
- фрагменты хромосом или целые хромосомы, не включенные в состав ядра
Микроядра
- фрагменты хромосом или целые хромосомы, не включенные в состав ядра
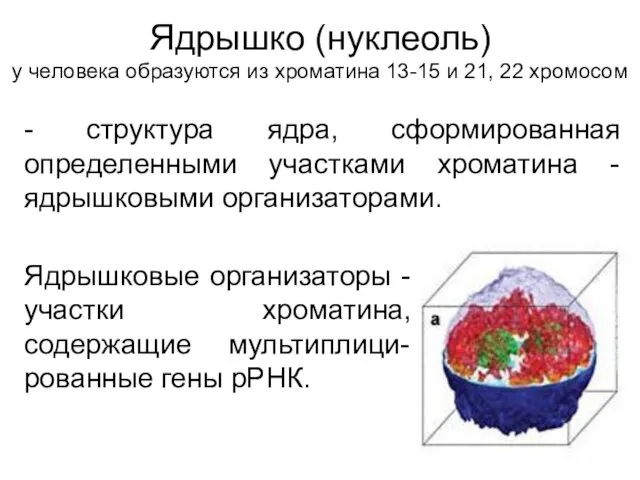
Ядрышко (нуклеоль)
у человека образуются из хроматина 13-15 и 21, 22 хромосом
-
Ядрышко (нуклеоль)
у человека образуются из хроматина 13-15 и 21, 22 хромосом
-

Размеры и количество ядрышек
В интерфазном ядре на светооптическом уровне – плотная
Размеры и количество ядрышек
В интерфазном ядре на светооптическом уровне – плотная

Структура ядрышка
Фибриллярный компонент - внутренняя часть ядрышка, состоит из нитей
Структура ядрышка
Фибриллярный компонент - внутренняя часть ядрышка, состоит из нитей

Ядерный матрикс
Кариоскелет построен из негистоновых белков, формирующих разветвленную сеть, взаимодействующую
Ядерный матрикс
Кариоскелет построен из негистоновых белков, формирующих разветвленную сеть, взаимодействующую
 Образцовый Театр Моды Стиль, коллекция Привет из Простоквашино
Образцовый Театр Моды Стиль, коллекция Привет из Простоквашино Типология научных исследований
Типология научных исследований Игрушки к Новому Году для детей и родителей
Игрушки к Новому Году для детей и родителей Теорема Пифагора. Формулировка и доказательство теоремы
Теорема Пифагора. Формулировка и доказательство теоремы Презентация к уроку по теме Применение распределительного свойства умножения в 6 классе. Диск
Презентация к уроку по теме Применение распределительного свойства умножения в 6 классе. Диск Определения и понятия. Поиски и разведка месторождений полезных ископаемых. Горное дело. Бурение скважин
Определения и понятия. Поиски и разведка месторождений полезных ископаемых. Горное дело. Бурение скважин Визитка учителя математики Диск
Визитка учителя математики Диск Экономика. Предложение на рынке с совершенной конкуренцией
Экономика. Предложение на рынке с совершенной конкуренцией Подборка и анализ поселков численностью 2000 человек
Подборка и анализ поселков численностью 2000 человек Польові телефонні аппарати, комутатори та кабелі
Польові телефонні аппарати, комутатори та кабелі Проблемы долгов Евросоюза. Современные проблемы мировой финансовой системы
Проблемы долгов Евросоюза. Современные проблемы мировой финансовой системы Учебное занятие в системе дополнительного образования
Учебное занятие в системе дополнительного образования Отцы и дети. Аргументы
Отцы и дети. Аргументы Инфузионная терапия
Инфузионная терапия Пам’ятні виставки та стенди
Пам’ятні виставки та стенди المحاضرة السادسة تابع تصنيف الجرائم االلكترونية
المحاضرة السادسة تابع تصنيف الجرائم االلكترونية Дефекты сварных соединений при дуговой сварке
Дефекты сварных соединений при дуговой сварке Вентиляция выработок при их проведении
Вентиляция выработок при их проведении Общественные организации и движения в современном мире
Общественные организации и движения в современном мире Творческий экзамен в 8 классе
Творческий экзамен в 8 классе Презентация Е.Чарушин
Презентация Е.Чарушин Открытый банк заданий по математике. Задача №16
Открытый банк заданий по математике. Задача №16 Инновационные полигоны нефтегазодобывающей отрасли НГХК
Инновационные полигоны нефтегазодобывающей отрасли НГХК Беспроводные локальные сети Wlan
Беспроводные локальные сети Wlan Необычные деревья
Необычные деревья Мастер класс Изготовление цветов из бисера
Мастер класс Изготовление цветов из бисера Эволюция жизни на Земле
Эволюция жизни на Земле Творческий проект Слово и образ Отечества
Творческий проект Слово и образ Отечества