- Генетические основы горизонтальной эволюции. Лекция 4

Содержание
- 2. I. Явление горизонтального переноса в эволюции Прочтение геномов прокариот и эукариот показало недостаточность филогенетических представлений на
- 5. Благодаря междоменным переносов генов возникли 13 крупных групп архей (в основном, порядков).
- 7. Гены могут перемещаться: А) в пределах генома одной клетки; Б). между разными клетками одного организма; В).
- 9. 3. Признаки "чужеродного" происхождения гена - высокая степень его сходства с гомологичным геном из отдаленного таксона
- 10. В горизонтальные переносы реже вовлечены гены информационных систем (транскрипции, трансляции, репликации), составляющие базовый геном. Продукты этих
- 11. 4. Механизмы переноса А). Горизонтальная передача генов реализуется через различные каналы генетической коммуникации: процессы конъюгации, трансформации,
- 13. Половой процесс отличается от более примитивных механизмов: 1) большей избирательностью (то есть смешение генов происходит в
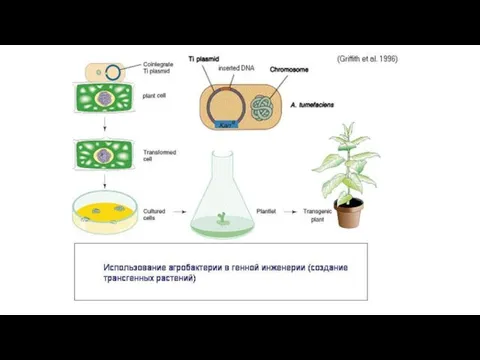
- 14. Современная генная инженерия базируется на принципах горизонтального переноса генов, т.е. на природной генной инженерии Например, бактерии,
- 17. II. Горизонтальная эволюция прокариот 1. Общие закономерности горизонтального переноса у прокариот: А). Наибольшее количество переносов характерно
- 18. Представители Proteobacteria активно обмениваются генами на уровне класса и даже отдела при условии обитания в одной
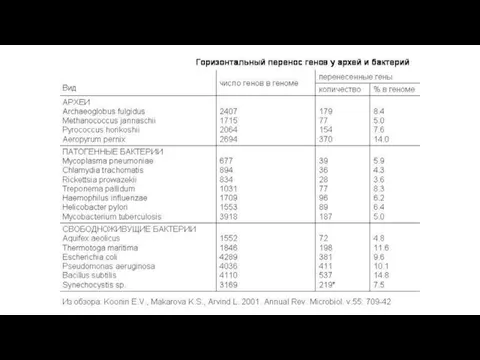
- 19. 2. Масштабы горизонтального переноса. За счет ГП разными видами бактерий в эволюции были получены примерно от
- 21. Референсное филогенетическое дерево архей. В скобках напротив каждого таксона: количество архей-специфичных семейств генов / количество семейств,
- 22. Эволюционная сеть архей. Сверху представлена референсная филогения, основанная на сцепленных последовательностях 70 универсальных генов. Шкала с
- 23. У прокариот нет жестких ограничений на размер фрагментов ДНК, получаемых из внешней среды. Возможен обмен целыми
- 25. 3. Последствия ГП у прокариот Примеры: А). Высокие адаптационные способности бактерий , которые быстро модифицируют функции
- 26. Бактерии развивают новые свойства, такие как резистентность к антибиотикам, преимущественно за счет «присвоения» генов других бактерий.
- 27. Пример 2. Эволюционные и экологические последствия ГП Существование уникального микробного сообщества с единственным видом Бактерию назвали
- 29. Прочтение генома показало, что это сульфатредуцирующая бактерия, но которая приобрела большое число генов за счет «ГП»
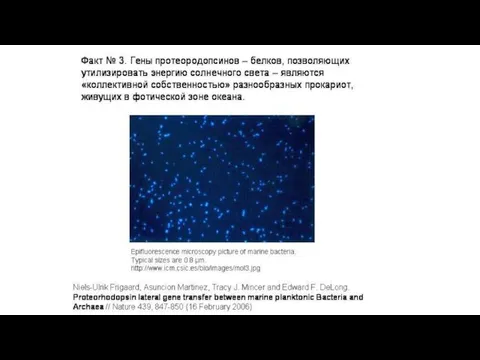
- 31. Пример 3. Кооперации за счет ГП У морских планктонных бактерий и у многих архей, живущих в
- 37. В первой колонке – процент генов, приобретенных путем горизонтального переноса в среднем на геном, если учитывать
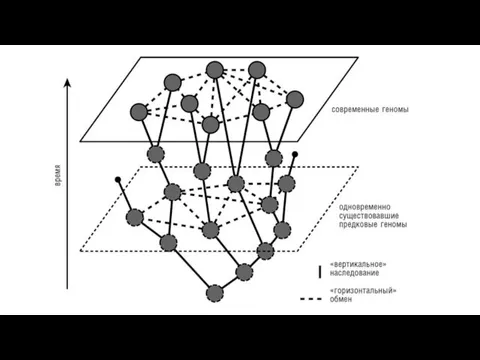
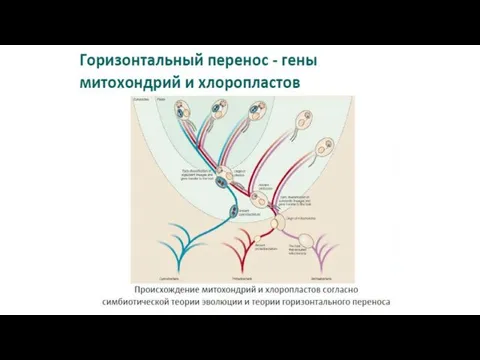
- 38. Эволюция прокариот - это не ветвящееся дерево, а сеть. Несмотря на то, что горизонтальный перенос происходит
- 39. III.
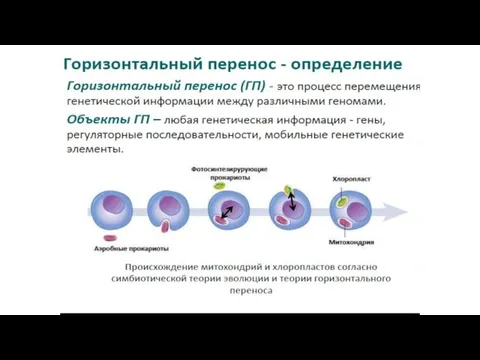
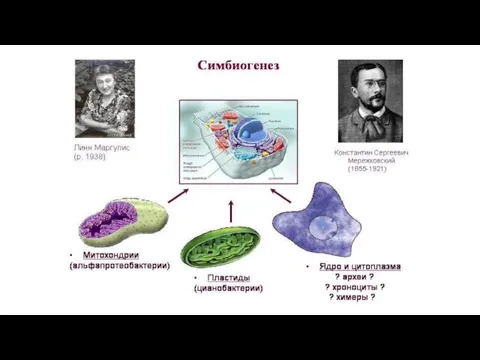
- 41. 1. Симбиогенез - один из основных путей эволюции эукариот А). Эукариотический ядерный геном является химерным с
- 46. Доказательства симбиотического происхождения органелл 1).
- 47. Доказательства симбиотического происхождения органелл 2).
- 49. Ядерный геном эукариот отличается огромным разнообразием генов, полученных в т. ч. от бактерий и архей. Чужеродная
- 50. В наши дни существует ряд организмов, содержащих внутри своих клеток другие клетки в качестве эндосимбионтов. Они,
- 51. Одноклеточная жгутиковая водоросль Cyanophora paradoxa содержит цианеллы — органоиды, напоминающие типичные хлоропласты красных водорослей, но отличающиеся
- 55. Схема эволюции эукариотических клеток. 1 — образование двойной мембраны ядра, 2 — приобретение митохондрий, 3 —
- 59. С Формирование Эукариотического ядерного генома путем симбиогенеза. У эукариот, нет механизма конъюгации для передачи генов неродственным
- 61. Эукариоты имеют: Архейную «сердцевину» (механизмы работы с генетической информацией и синтеза белка) Бактериальную «периферию» (обмен веществ
- 62. От архей эукариоты унаследовали многие компоненты информационных систем нуклеоцитоплазмы. Бактериальные эндосимбионты внесли большой вклад в формирование
- 63. По-видимому, в эволюции прото-эукариот был период ослабления изоляционных барьеров, когда происходила активная инкопрорация чужих генов. В
- 65. «Кольца жизни» (по Лейку), указывающие на формирование крупных таксонов организмов в результате эндосимбиоза
- 69. Б). Массовый горизонтальный перенос генов, связанный с симбиогенезом, происходил и после того, как сформировалась эукариотическая клетка.
- 71. Рекордсменом заимствования на сегодняшний день является одноклеточная диатомовая водоросль Phaeodactylum. В геномах диатомей, которые появились 180
- 72. Польза диатомеям от бактериальных генов Расширение биохимических возможностей. Заимствованные бактериальные гены участвуют в построении ажурных кремневых
- 73. 2. Значение для эволюционного пути. Выявлено более 170 генов, унаследованных предками диатомей от красных водорослей. Диатомеи
- 74. 2. Иные пути переноса генов, не связанные с симбиогенезом От одних одноклеточных эукариот к другим Примеры:
- 75. Б). В эволюции эукариот из группы (Apicomplexa), куда относятся токсоплазма и малярийный плазмодий, было не менее
- 76. Перенос генов у многоклеточных эукариот 1). ГРИБЫ а)
- 77. У почвенных бактерий живущие вблизи корней растений есть ген acdS, который способствует росту корневой системы. Этот
- 78. в).
- 79. Один из многочисленных внутриклеточных симбионтов тлей представляет собой двух «вложенных» друг в друга бактерий: бета-протеобактерию, внутри
- 80. 2).Растения «ГП» митохондриальных генов от одного растения к другому. Описано более 40 случаев горизонтального переноса
- 81. 2).Растения «ГП» митохондриальных генов от одного растения к другому (более 40 случаев)
- 84. Описаны и случаи обмена ядерными генами. Например, была обнаружена горизонтальная передача транспозона между двумя видами злаков
- 85. Часто в обмене участвуют паразитические или эпифитные растения, то есть нужен тесный физический контакт. Пример: Раффлезия
- 88. 3). Горизонтальный перенос у животных а) между органеллами
- 90. б).
- 91. От паразитических бактерий в хромосомы животных-хозяев. могут вставляться гены И целые геномы
- 94. Вольбахия — паразитическая бактерия, живущая в клетках многих беспозвоночных. Это «микроб-манипулятор», т.к. при помощи регуляторных белков
- 95. Вольбахия паразитирует в клетках беспозвоночных уже 100 млн. лет, ее предки тоже были внутриклеточными паразитами. Долгое
- 96. Многие гены, заимствованные мухой у бактерии, работают, или по крайней мере транскрибируются. Это значит, что инкорпорация
- 97. в).
- 98. Важнейшую роль в эволюции играют гены, которые животные заимствуют у г) вирусов, транспозонов, ретротранспозонов. Это наглядные
- 99. г)
- 100. Мобильные элементы обладают повышенной способностью к горизонтальной передаче по сравнению с другими частями генома многоклеточных.
- 101. Сравнение генома короткохвостого опоссума, прочтенного в 2007 году с геномами плацентарных (человека, мыши, крысы и собаки)
- 106. В результате анализа геномов млекопитающих: опоссум, обезьяна саймири, покусанных южноамериканским кровососущим жуком Rhodnius prolixus, был обнаружен
- 110. Примерно 42% нашего генома составляют ретротранспозоны – самоповторяющиеся генетические элементы, которые копируют сами себя и вставляют
- 111. Эволюция BovB по результатам анализа 2012 года. Эволюция BovB из статьи 2017 года.
- 119. Фрагменты ДНК вирусов и транспозонов часто «приручаются» высшими организмами и начинают выполнять полезные функции в геноме.
- 120. Примеры важных эволюционных событий, связанных с молекулярным одомашниванием: 1) Ферменты теломеразы, служащие для восстановления концевых участков
- 121. Установлен процесс превращения так называемой «мусорной» ДНК» в полезные элементы генома. многие из транспозонов и ретротранспозонов
- 122. Описано превращение прирученных генов мобильных элементов в самые настоящие транскрипционные факторы. Так, у арабидопсиса два транскрипционных
- 124. В эволюции приматов тоже имело место приобретение полезных генов «со стороны», а именно от ретровирусов. Например,
- 126. Возможные функции вирусных генов у человека и обезьян в плаценте: управление слиянием клеток в ходе формирования
- 127. Исходный ретровирус встроился в геном наших предков и стал эндогенным ретровирусом (ЭРВ) после того, как разделились
- 129. Некоторые животные могут в массовом порядке заимствовать гены не у вирусов, а у клеточных организмов: у
- 130. Бделлоидные коловратки (ок. 400 видов) представляют собой удивительное исключение. Это класс, не имеющих самцов и размножающихся
- 132. В концевых участках хромосом бделлоидной коловратки Adineta vaga было обнаружено множество генов, не встречающихся ни у
- 134. Как известно, животные стараются оберегать свои половые клетки от чужого генетического материала, в том числе вирусного.
- 135. Некоторые заимствованные бактериальные гены у коловраток сохранили структуру, характерную для прокариотических генов, а другие имеют интроны,
- 136. Новые случаи горизонтального переноса у многоклеточных разных таксономических групп обнаруживаются постоянно. Пока счет прочтенных геномов многоклеточных
- 139. Скачать презентацию

I. Явление горизонтального переноса в эволюции
Прочтение геномов
прокариот и эукариот показало
I. Явление горизонтального переноса в эволюции
Прочтение геномов
прокариот и эукариот показало


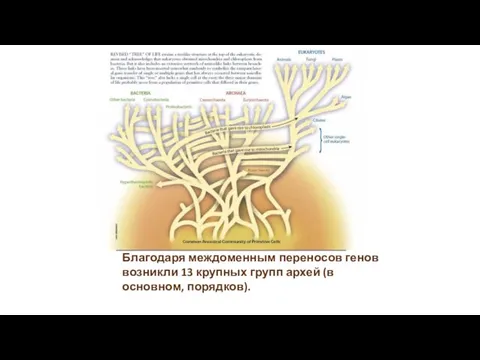
Благодаря междоменным переносов генов возникли 13 крупных групп архей (в основном,
Благодаря междоменным переносов генов возникли 13 крупных групп архей (в основном,

Гены могут перемещаться:
А) в пределах генома одной клетки;
Б). между разными
Гены могут перемещаться:
А) в пределах генома одной клетки;
Б). между разными

3. Признаки "чужеродного" происхождения гена
- высокая степень его сходства
3. Признаки "чужеродного" происхождения гена
- высокая степень его сходства

В горизонтальные переносы реже вовлечены гены информационных систем (транскрипции, трансляции, репликации),
В горизонтальные переносы реже вовлечены гены информационных систем (транскрипции, трансляции, репликации),

4. Механизмы переноса
А). Горизонтальная передача генов реализуется через различные каналы генетической
4. Механизмы переноса
А). Горизонтальная передача генов реализуется через различные каналы генетической


Половой процесс отличается от более примитивных механизмов:
1) большей избирательностью (то
1) большей избирательностью (то

Современная генная инженерия базируется на принципах горизонтального переноса генов, т.е. на
Современная генная инженерия базируется на принципах горизонтального переноса генов, т.е. на


II. Горизонтальная эволюция прокариот
1. Общие закономерности горизонтального переноса у прокариот:
А). Наибольшее
II. Горизонтальная эволюция прокариот
1. Общие закономерности горизонтального переноса у прокариот:
А). Наибольшее

Представители Proteobacteria активно обмениваются генами на уровне класса и даже отдела
Представители Proteobacteria активно обмениваются генами на уровне класса и даже отдела

2. Масштабы горизонтального переноса.
За счет ГП разными видами бактерий в
2. Масштабы горизонтального переноса.
За счет ГП разными видами бактерий в
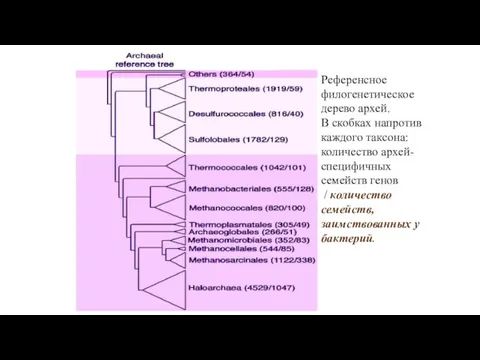
Референсное филогенетическое дерево архей.
В скобках напротив каждого таксона: количество архей-специфичных
Референсное филогенетическое дерево архей.
В скобках напротив каждого таксона: количество архей-специфичных

Эволюционная сеть архей.
Сверху представлена референсная филогения, основанная на сцепленных последовательностях
Эволюционная сеть архей.
Сверху представлена референсная филогения, основанная на сцепленных последовательностях

У прокариот нет жестких ограничений на размер фрагментов ДНК, получаемых
У прокариот нет жестких ограничений на размер фрагментов ДНК, получаемых


3. Последствия ГП у прокариот
Примеры:
А). Высокие адаптационные способности бактерий , которые
3. Последствия ГП у прокариот
Примеры:
А). Высокие адаптационные способности бактерий , которые

Бактерии развивают новые свойства, такие как резистентность к антибиотикам, преимущественно за
Бактерии развивают новые свойства, такие как резистентность к антибиотикам, преимущественно за

Пример 2. Эволюционные и экологические последствия ГП
Существование уникального микробного сообщества с
Пример 2. Эволюционные и экологические последствия ГП
Существование уникального микробного сообщества с

Прочтение генома показало, что это сульфатредуцирующая бактерия, но которая приобрела


Пример 3. Кооперации за счет ГП
У морских планктонных бактерий и у
Пример 3. Кооперации за счет ГП
У морских планктонных бактерий и у

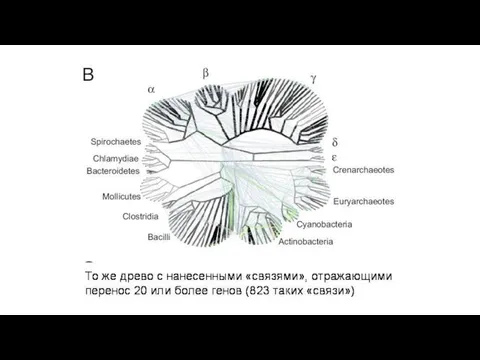
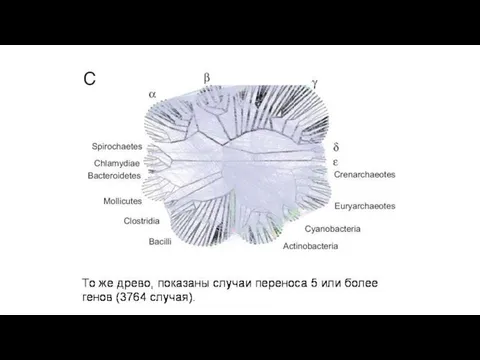
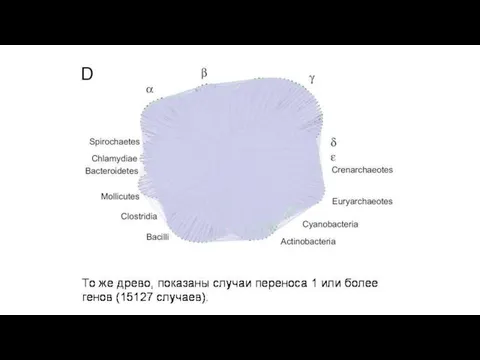

В первой колонке – процент генов, приобретенных путем горизонтального переноса в
В первой колонке – процент генов, приобретенных путем горизонтального переноса в

Эволюция прокариот -
это не ветвящееся дерево, а сеть.
Несмотря на
Эволюция прокариот -
это не ветвящееся дерево, а сеть.
Несмотря на
III.
III.


1. Симбиогенез - один из основных путей эволюции эукариот
А).
1. Симбиогенез - один из основных путей эволюции эукариот
А).
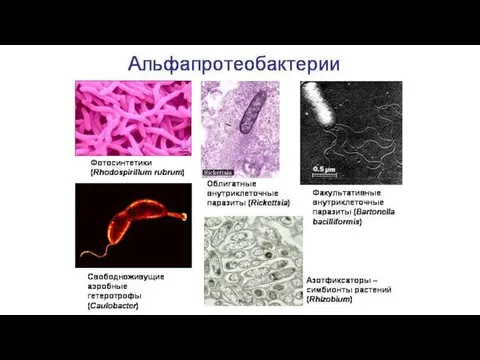
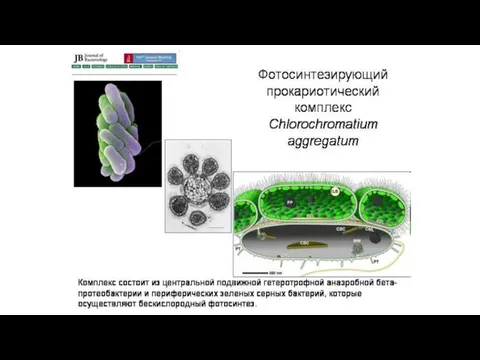
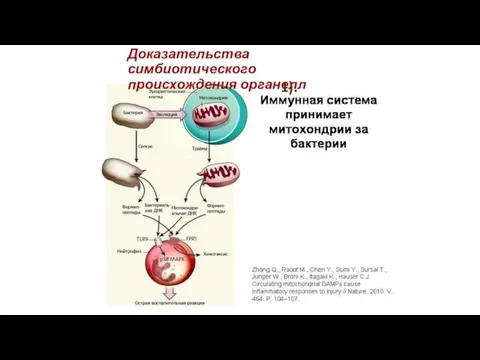
Доказательства симбиотического происхождения органелл
1).
Доказательства симбиотического происхождения органелл
1).

Доказательства симбиотического происхождения органелл
2).
Доказательства симбиотического происхождения органелл
2).

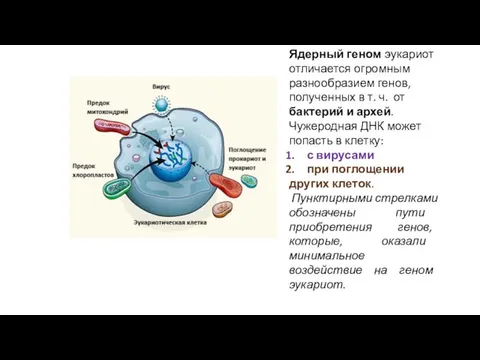
Ядерный геном эукариот отличается огромным разнообразием генов, полученных в т. ч.
Ядерный геном эукариот отличается огромным разнообразием генов, полученных в т. ч.

В наши дни существует ряд организмов, содержащих внутри своих клеток другие
В наши дни существует ряд организмов, содержащих внутри своих клеток другие

Одноклеточная жгутиковая водоросль Cyanophora paradoxa содержит цианеллы — органоиды, напоминающие типичные хлоропласты
Одноклеточная жгутиковая водоросль Cyanophora paradoxa содержит цианеллы — органоиды, напоминающие типичные хлоропласты

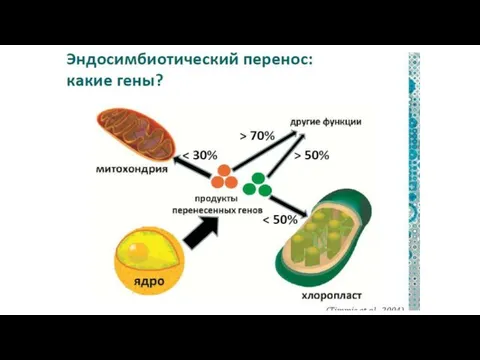
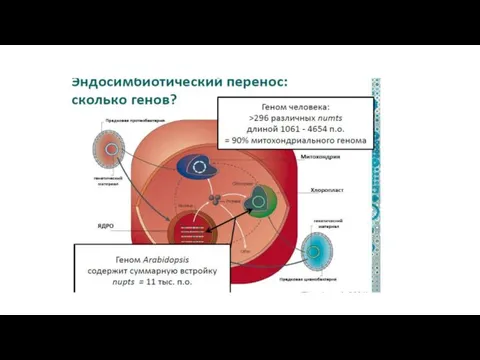

Схема эволюции эукариотических клеток.
1 — образование двойной мембраны ядра,
2 — приобретение
Схема эволюции эукариотических клеток.
1 — образование двойной мембраны ядра,
2 — приобретение




С Формирование Эукариотического ядерного генома путем симбиогенеза.
У эукариот, нет
С Формирование Эукариотического ядерного генома путем симбиогенеза.
У эукариот, нет

Эукариоты имеют:
Архейную «сердцевину» (механизмы работы с генетической информацией и
Эукариоты имеют:
Архейную «сердцевину» (механизмы работы с генетической информацией и

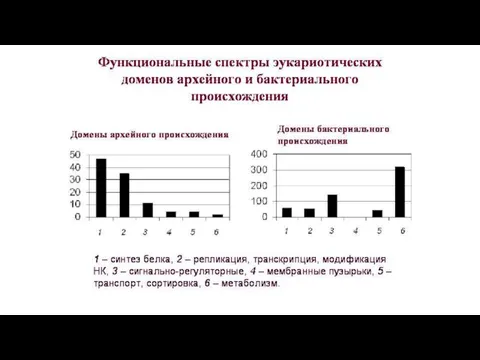
От архей эукариоты унаследовали многие компоненты информационных систем нуклеоцитоплазмы.
Бактериальные эндосимбионты
Бактериальные эндосимбионты

По-видимому, в эволюции прото-эукариот был период ослабления изоляционных барьеров, когда происходила
По-видимому, в эволюции прото-эукариот был период ослабления изоляционных барьеров, когда происходила

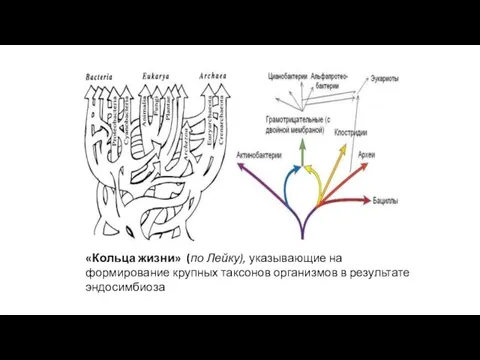
«Кольца жизни» (по Лейку), указывающие на формирование крупных таксонов организмов в
«Кольца жизни» (по Лейку), указывающие на формирование крупных таксонов организмов в
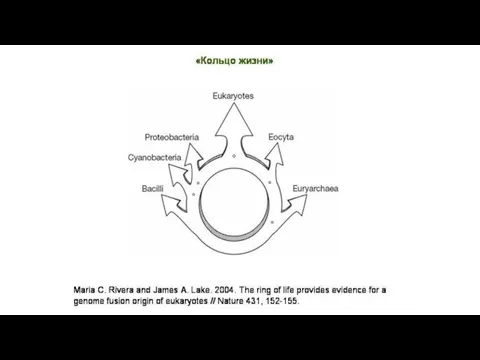
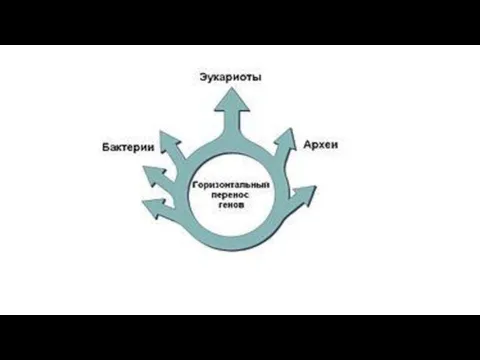
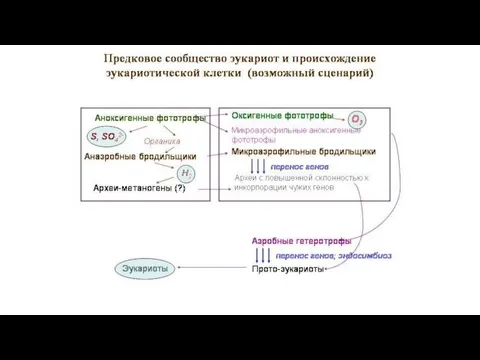

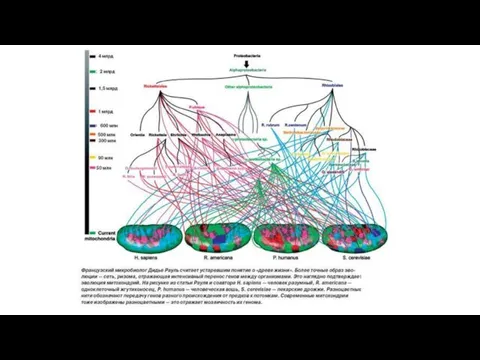
Б). Массовый горизонтальный перенос генов,
связанный с симбиогенезом,
происходил и после
связанный с симбиогенезом,
происходил и после


Рекордсменом заимствования на сегодняшний день является одноклеточная диатомовая водоросль Phaeodactylum.
В
Рекордсменом заимствования на сегодняшний день является одноклеточная диатомовая водоросль Phaeodactylum.
В

Польза диатомеям от бактериальных генов
Расширение биохимических возможностей.
Заимствованные бактериальные гены
Польза диатомеям от бактериальных генов
Расширение биохимических возможностей.
Заимствованные бактериальные гены

2. Значение для эволюционного пути.
Выявлено более 170 генов, унаследованных
2. Значение для эволюционного пути.
Выявлено более 170 генов, унаследованных

2. Иные пути переноса генов,
не связанные с симбиогенезом
От одних
2. Иные пути переноса генов,
не связанные с симбиогенезом
От одних

Б). В эволюции эукариот из группы (Apicomplexa), куда относятся токсоплазма и
Б). В эволюции эукариот из группы (Apicomplexa), куда относятся токсоплазма и

Перенос генов у многоклеточных эукариот
1). ГРИБЫ
а)
Перенос генов у многоклеточных эукариот
1). ГРИБЫ
а)
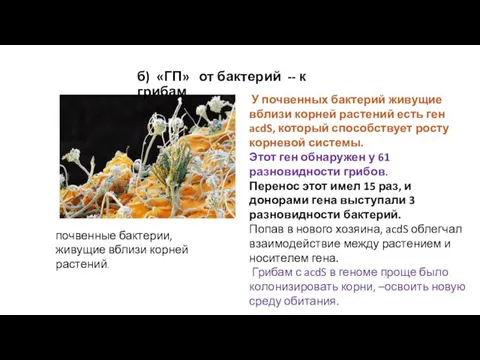
У почвенных бактерий живущие вблизи корней растений есть ген acdS,
У почвенных бактерий живущие вблизи корней растений есть ген acdS,

в).
в).

Один из многочисленных внутриклеточных симбионтов тлей представляет собой двух «вложенных» друг
Один из многочисленных внутриклеточных симбионтов тлей представляет собой двух «вложенных» друг

2).Растения
«ГП» митохондриальных генов от
одного растения к другому.
Описано
2).Растения
«ГП» митохондриальных генов от
одного растения к другому.
Описано

2).Растения «ГП» митохондриальных генов от
одного растения к другому (более 40
2).Растения «ГП» митохондриальных генов от
одного растения к другому (более 40



Описаны и случаи обмена ядерными генами.
Например, была обнаружена горизонтальная передача
Описаны и случаи обмена ядерными генами.
Например, была обнаружена горизонтальная передача

Часто в обмене участвуют паразитические или эпифитные растения, то есть нужен
Часто в обмене участвуют паразитические или эпифитные растения, то есть нужен



3). Горизонтальный перенос у животных
а) между органеллами
3). Горизонтальный перенос у животных
а) между органеллами
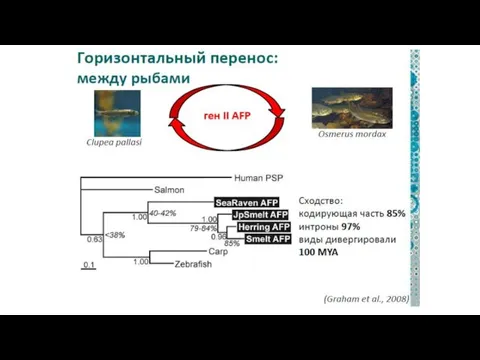
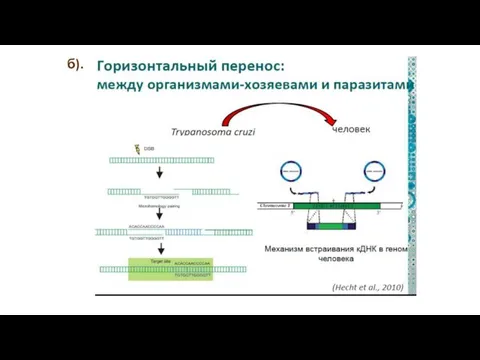
б).
б).

От паразитических бактерий
в хромосомы животных-хозяев.
могут вставляться
гены
И целые
От паразитических бактерий
в хромосомы животных-хозяев.
могут вставляться
гены
И целые
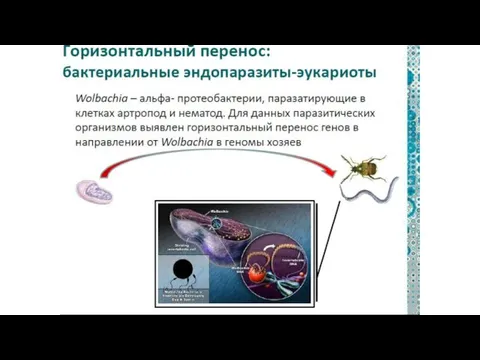
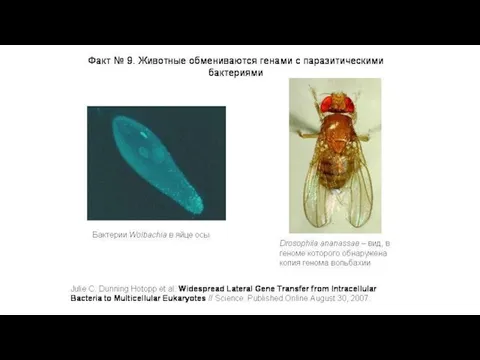

Вольбахия — паразитическая бактерия, живущая в клетках многих беспозвоночных.
Это «микроб-манипулятор»,
Это «микроб-манипулятор»,

Вольбахия паразитирует в клетках беспозвоночных уже 100 млн. лет,
ее
Вольбахия паразитирует в клетках беспозвоночных уже 100 млн. лет,
ее

Многие гены, заимствованные мухой у бактерии, работают, или по крайней мере
Многие гены, заимствованные мухой у бактерии, работают, или по крайней мере

в).
в).

Важнейшую роль в эволюции играют гены, которые животные заимствуют у
г)
г)
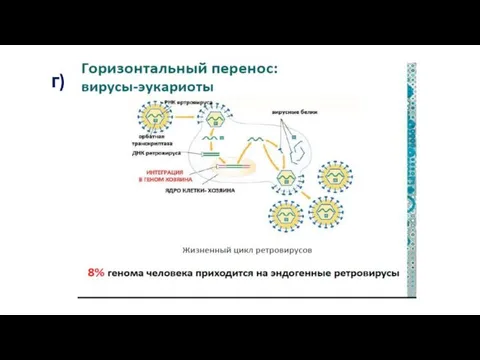
г)
г)

Мобильные элементы обладают повышенной способностью
к горизонтальной передаче
по сравнению с
Мобильные элементы обладают повышенной способностью
к горизонтальной передаче
по сравнению с

Сравнение генома короткохвостого опоссума, прочтенного в 2007 году с геномами плацентарных
Сравнение генома короткохвостого опоссума, прочтенного в 2007 году с геномами плацентарных





В результате анализа геномов млекопитающих: опоссум, обезьяна саймири,
покусанных южноамериканским кровососущим
В результате анализа геномов млекопитающих: опоссум, обезьяна саймири,
покусанных южноамериканским кровососущим




Примерно 42% нашего генома составляют ретротранспозоны – самоповторяющиеся генетические элементы,
Примерно 42% нашего генома составляют ретротранспозоны – самоповторяющиеся генетические элементы,
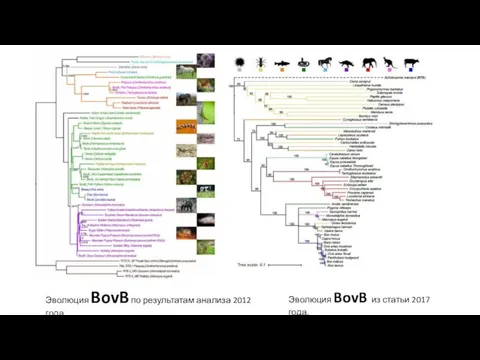
Эволюция BovB по результатам анализа 2012 года.
Эволюция BovB из статьи
Эволюция BovB по результатам анализа 2012 года.
Эволюция BovB из статьи



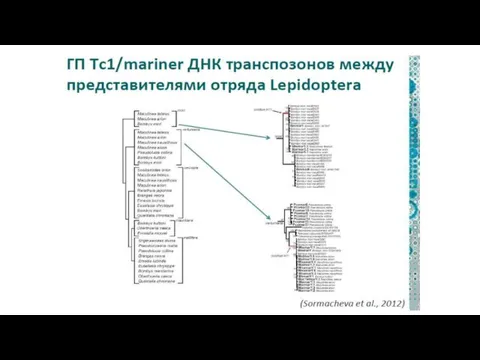




Фрагменты ДНК вирусов и транспозонов часто «приручаются» высшими организмами и начинают
Фрагменты ДНК вирусов и транспозонов часто «приручаются» высшими организмами и начинают

Примеры важных эволюционных событий, связанных с молекулярным одомашниванием:
1) Ферменты теломеразы,
Примеры важных эволюционных событий, связанных с молекулярным одомашниванием:
1) Ферменты теломеразы,

Установлен процесс превращения так называемой «мусорной» ДНК» в полезные элементы генома.

Описано превращение прирученных генов мобильных элементов в самые настоящие транскрипционные факторы.
Так,
Описано превращение прирученных генов мобильных элементов в самые настоящие транскрипционные факторы.
Так,


В эволюции приматов тоже имело место приобретение полезных генов «со стороны»,
В эволюции приматов тоже имело место приобретение полезных генов «со стороны»,
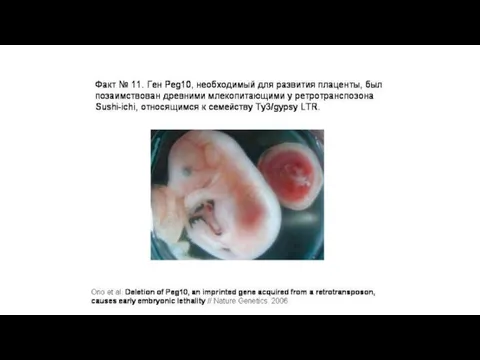

Возможные функции вирусных генов у человека и обезьян в плаценте:
управление слиянием
Возможные функции вирусных генов у человека и обезьян в плаценте:
управление слиянием

Исходный ретровирус встроился в геном наших предков и стал эндогенным ретровирусом
Исходный ретровирус встроился в геном наших предков и стал эндогенным ретровирусом
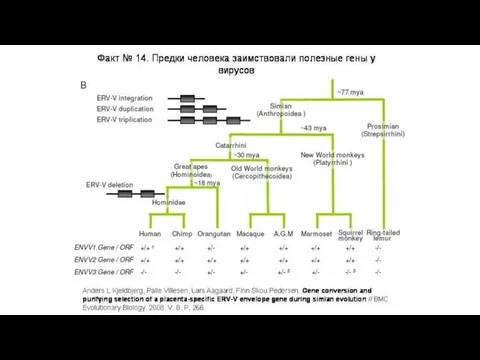

Некоторые животные могут в массовом порядке заимствовать гены не у
Некоторые животные могут в массовом порядке заимствовать гены не у

Бделлоидные коловратки (ок. 400 видов) представляют собой удивительное исключение. Это класс,
Бделлоидные коловратки (ок. 400 видов) представляют собой удивительное исключение. Это класс,


В концевых участках хромосом бделлоидной коловратки Adineta vaga было обнаружено множество
В концевых участках хромосом бделлоидной коловратки Adineta vaga было обнаружено множество
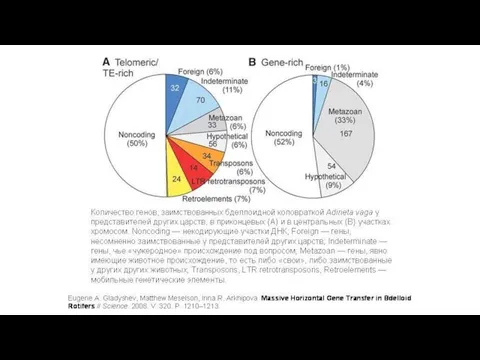

Как известно, животные стараются оберегать свои половые клетки от чужого генетического
Как известно, животные стараются оберегать свои половые клетки от чужого генетического

Некоторые заимствованные бактериальные гены у коловраток сохранили структуру, характерную для прокариотических
Некоторые заимствованные бактериальные гены у коловраток сохранили структуру, характерную для прокариотических

Новые случаи горизонтального переноса у многоклеточных разных таксономических групп обнаруживаются постоянно.
Новые случаи горизонтального переноса у многоклеточных разных таксономических групп обнаруживаются постоянно.
 Мечети России
Мечети России Клуб Молодая семья г. Великий Устюг МЦ Авангард
Клуб Молодая семья г. Великий Устюг МЦ Авангард Металлорежущие станки
Металлорежущие станки Development road map and design means
Development road map and design means Презентация к открытому занятию внеурочной деятельности в 4 классе по теме: Цветок лотоса (оригами)
Презентация к открытому занятию внеурочной деятельности в 4 классе по теме: Цветок лотоса (оригами) Шаблон Фракталы-10
Шаблон Фракталы-10 Методические рекомендации по проведению мастер- класса
Методические рекомендации по проведению мастер- класса Миграции населения
Миграции населения Как подключить EDI-сообщения с ООО АШАН и ООО АТАК
Как подключить EDI-сообщения с ООО АШАН и ООО АТАК Право (часть 2)
Право (часть 2) Уголовная и административная ответственность
Уголовная и административная ответственность Обморожение у детей
Обморожение у детей Интерактивная экскурсия Пирамиды Гизы (Египетские пирамиды)
Интерактивная экскурсия Пирамиды Гизы (Египетские пирамиды) Строитель своей души
Строитель своей души Квантовая механика и квантовая химия
Квантовая механика и квантовая химия Сооружения для приема воды из поверхностных источников
Сооружения для приема воды из поверхностных источников IPC-D-325 Table 4-1 Typical Master Drawing
IPC-D-325 Table 4-1 Typical Master Drawing Холодильное оборудование
Холодильное оборудование Направления концепции генерального плана муниципального образования город Краснодар
Направления концепции генерального плана муниципального образования город Краснодар Установочная конференция по ПП в ПСО
Установочная конференция по ПП в ПСО Модерн в архитектуре
Модерн в архитектуре Общая характеристика младшего школьного возраста
Общая характеристика младшего школьного возраста Понятия схемотехника, телекоммуникационные устройства
Понятия схемотехника, телекоммуникационные устройства Zaschita_Alexeev_SM1_81B
Zaschita_Alexeev_SM1_81B Волейбол. История
Волейбол. История Изготовление и оформление лекал. Экспорт лекал из GRAFIS (швейное дело)
Изготовление и оформление лекал. Экспорт лекал из GRAFIS (швейное дело) Молоко и молочные продукты
Молоко и молочные продукты Презентация_о_Петре_Великом,_как_о_личности
Презентация_о_Петре_Великом,_как_о_личности