Информационные технологии в биологических исследованиях. Лекция 4: Принципы построения математических моделей презентация
- Информационные технологии в биологических исследованиях. Лекция 4: Принципы построения математических моделей

Содержание
- 2. В любой науке существуют простые модели, которые поддаются аналитическому исследованию и обладают свойствами, которые позволяют описывать
- 3. Базовые модели в биологии Калибровачная зависимость Популяционные модели: - В отсутствии ограничений - С ограничениями –
- 4. Самая простая и очень нужная модель в биологии – калибровочная кривая, вернее процесс ее построения и
- 5. Самая простая и очень нужная модель в биологии – калибровочная кривая, вернее процесс ее построения и
- 6. Модели роста численности популяции Любой процесс происходит во времени. Скорость – изменение за единицу времени. Скорость
- 7. Рост показателя и скорость его изменения Фундаментальное предположение для модели роста - скорость роста пропорциональна численности
- 8. Рост колонии микроорганизмов За время Δt прирост численности равен: Δx = R - S, где R
- 9. Рост колонии микроорганизмов В простейшем случае, когда рождаемость и смертность пропорциональны численности: α скорость рождаемости, например,
- 10. Разделим переменные и проинтегрируем Решение уровнения Делим обе части равенства (уравнения) на одно и то же
- 11. Интегрирование – действие, обратное дифференцированию получаем
- 12. График зависимости численности от времени в соот-ветствии с законом экспоненциального роста (слева), а справа представлена зависимость
- 13. Варианты динамики популяции
- 14. Только в условиях неограниченных ресурсов изолированная популяция развивалась бы в соответствии с экспоненциальным законом В реальных
- 15. Примеры динамики популяций Численность поголовья овец на острове Тасмания (Davidson, 1938) Изменение численности Daphnia magna (Frail,
- 16. Ограниченный рост. Уравнение Ферхюльста Уравнение получено эмпирически, из анализа результатов наблюдений и экспериментов δx2 , второй
- 17. Аналитическое решение уравнения После интегрирования получим Произведем разделение переменных: Представим левую часть в виде суммы
- 18. Здесь С — произвольная постоянная, которая определяется начальным значением X0: Перейдем от логарифмов к переменным, помня,
- 19. Находим произвольную постоянную С t
- 20. Критические уровни численности Первый член в правой части описывает размножение двуполой популяции, скорость которого пропорциональ-на квадрату
- 21. Критические уровни численности Величины L и К различны для разных популяций и могут быть определены из
- 22. Тип поведения зависит от величины константы собствен-ной скорости роста r. Кривые зависимости значения численности в данный
- 23. Модели взаимодействия двух популяций a — константы собствен-ной скорости роста видов, c — константы внутри-видовой конкуренции,
- 24. Модель хищник-жертва При различных соотношениях параметров в системе возможно выживание только жертвы, только хищника (если у
- 26. Скачать презентацию

В любой науке существуют простые модели, которые поддаются аналитическому исследованию и
В любой науке существуют простые модели, которые поддаются аналитическому исследованию и

Базовые модели в биологии
Калибровачная зависимость
Популяционные модели:
- В отсутствии ограничений
- С
Базовые модели в биологии
Калибровачная зависимость
Популяционные модели:
- В отсутствии ограничений
- С
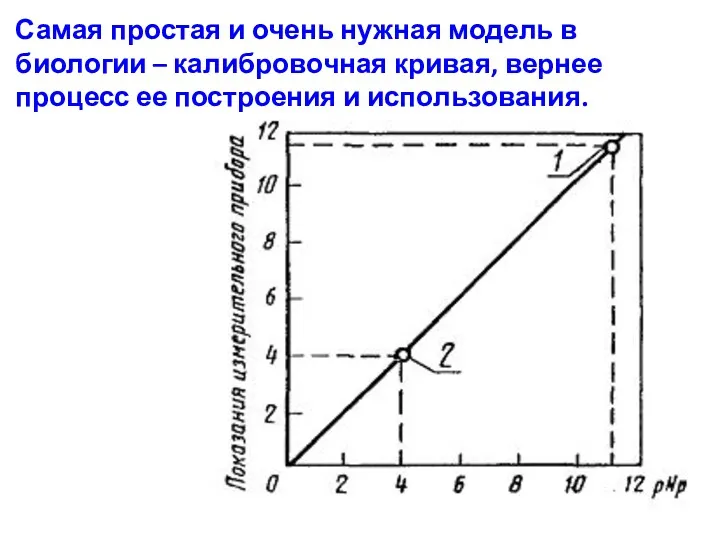
Самая простая и очень нужная модель в биологии – калибровочная кривая,
Самая простая и очень нужная модель в биологии – калибровочная кривая,

Самая простая и очень нужная модель в биологии – калибровочная кривая,
Самая простая и очень нужная модель в биологии – калибровочная кривая,

Модели роста численности популяции
Любой процесс происходит во времени.
Скорость – изменение
Модели роста численности популяции
Любой процесс происходит во времени.
Скорость – изменение
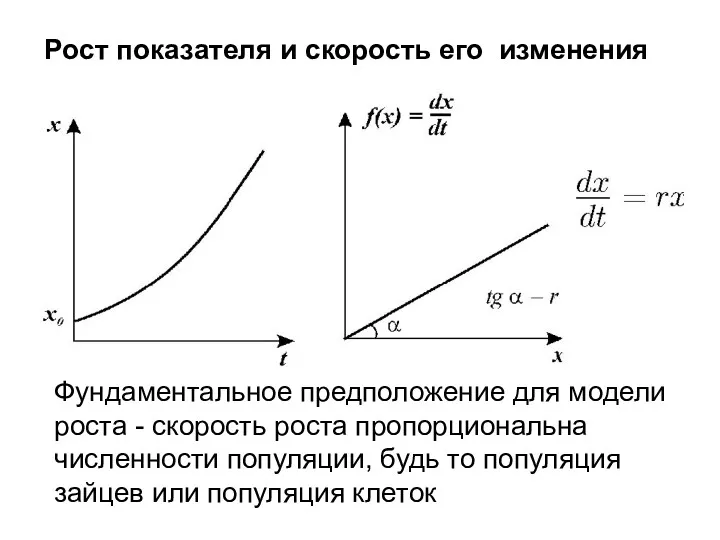
Рост показателя и скорость его изменения
Фундаментальное предположение для модели роста -
Рост показателя и скорость его изменения
Фундаментальное предположение для модели роста -

Рост колонии микроорганизмов
За время Δt прирост численности равен:
Δx = R -
Рост колонии микроорганизмов
За время Δt прирост численности равен:
Δx = R -

Рост колонии микроорганизмов
В простейшем случае, когда рождаемость и смертность пропорциональны численности:
α
Рост колонии микроорганизмов
В простейшем случае, когда рождаемость и смертность пропорциональны численности:
α

Разделим переменные и проинтегрируем
Решение уровнения
Делим обе части равенства (уравнения) на одно
Разделим переменные и проинтегрируем
Решение уровнения
Делим обе части равенства (уравнения) на одно

Интегрирование – действие, обратное дифференцированию
получаем
Интегрирование – действие, обратное дифференцированию
получаем
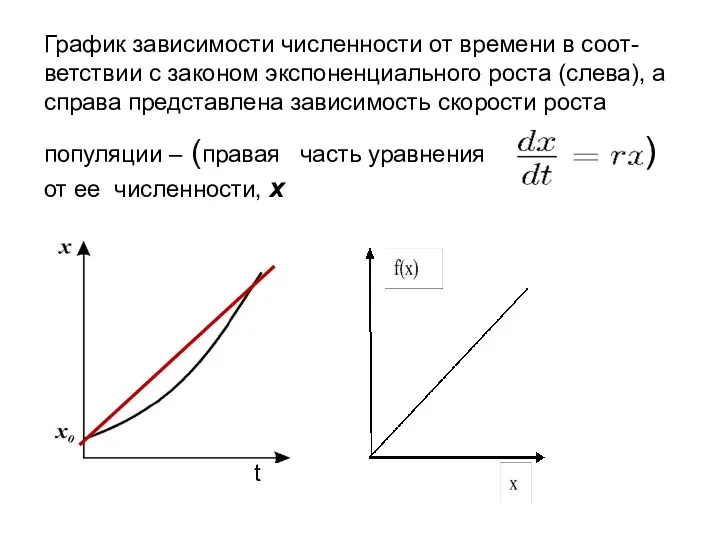
График зависимости численности от времени в соот-ветствии с законом экспоненциального роста
График зависимости численности от времени в соот-ветствии с законом экспоненциального роста
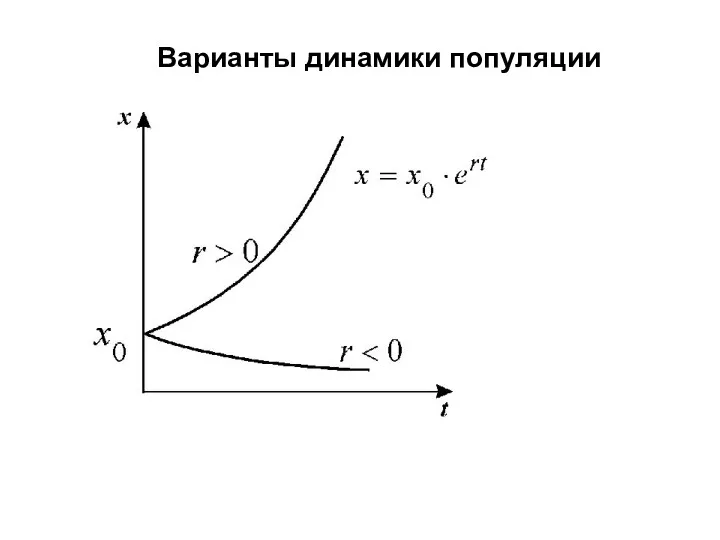
Варианты динамики популяции
Варианты динамики популяции

Только в условиях неограниченных ресурсов
изолированная популяция развивалась бы в соответствии
Только в условиях неограниченных ресурсов
изолированная популяция развивалась бы в соответствии
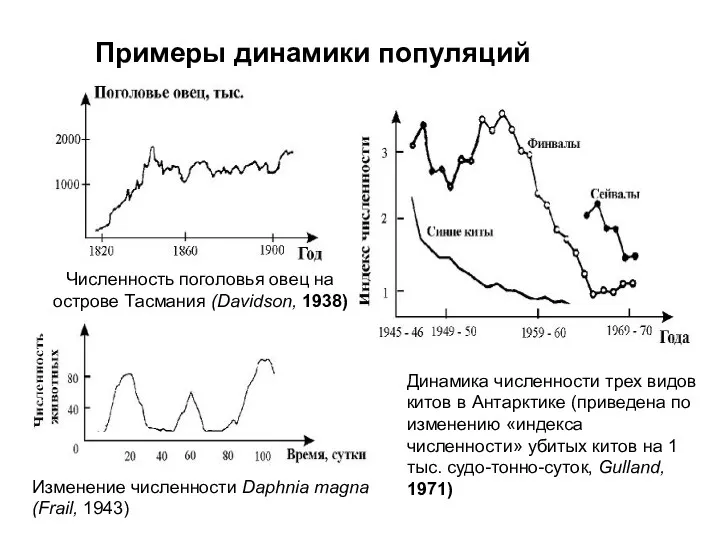
Примеры динамики популяций
Численность поголовья овец на
острове Тасмания (Davidson, 1938)
Изменение
Примеры динамики популяций
Численность поголовья овец на
острове Тасмания (Davidson, 1938)
Изменение

Ограниченный рост. Уравнение Ферхюльста
Уравнение получено эмпирически, из анализа результатов наблюдений
Ограниченный рост. Уравнение Ферхюльста
Уравнение получено эмпирически, из анализа результатов наблюдений

Аналитическое решение уравнения
После интегрирования получим
Произведем разделение переменных:
Представим левую часть в виде
Аналитическое решение уравнения
После интегрирования получим
Произведем разделение переменных:
Представим левую часть в виде

Здесь С — произвольная постоянная, которая определяется начальным значением X0:
Перейдем от
Здесь С — произвольная постоянная, которая определяется начальным значением X0:
Перейдем от
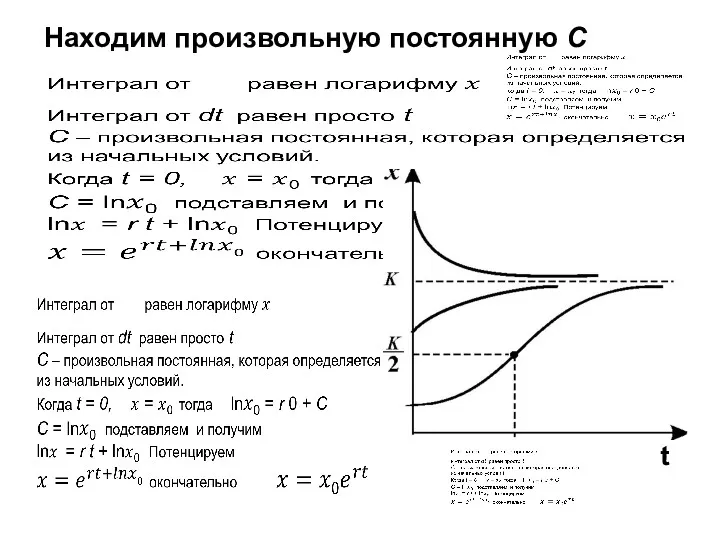
Находим произвольную постоянную С
t
Находим произвольную постоянную С
t

Критические уровни численности
Первый член в правой части описывает размножение двуполой
Критические уровни численности
Первый член в правой части описывает размножение двуполой
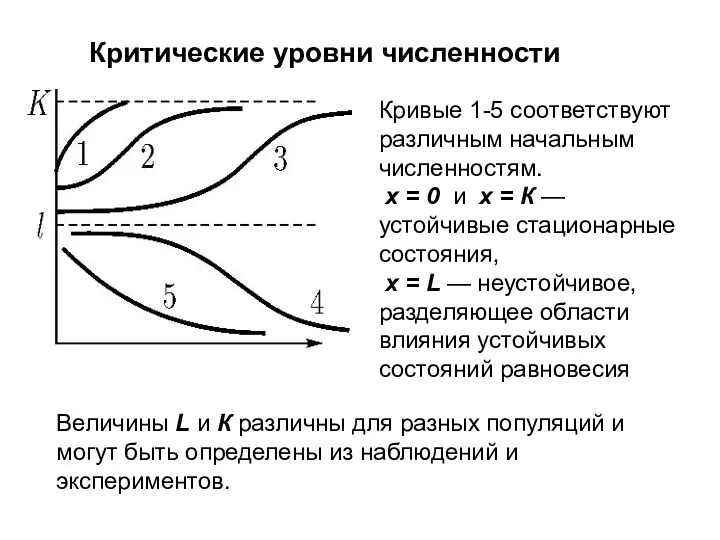
Критические уровни численности
Величины L и К различны для разных популяций
Критические уровни численности
Величины L и К различны для разных популяций

Тип поведения зависит от величины константы собствен-ной скорости роста r. Кривые
Тип поведения зависит от величины константы собствен-ной скорости роста r. Кривые

Модели взаимодействия двух популяций
a — константы собствен-ной скорости роста видов,
c
Модели взаимодействия двух популяций
a — константы собствен-ной скорости роста видов,
c

Модель хищник-жертва
При различных соотношениях параметров в системе возможно выживание только жертвы,
Модель хищник-жертва
При различных соотношениях параметров в системе возможно выживание только жертвы,
 Автоматизация Р в слогах и словах
Автоматизация Р в слогах и словах Знаменитые жители СПб
Знаменитые жители СПб A Case Bronchitis
A Case Bronchitis Идентификация и фальсификация молочных товаров
Идентификация и фальсификация молочных товаров Испытания в процессе проектирования РЭА
Испытания в процессе проектирования РЭА Шахматные фигуры. Начальная позиция.
Шахматные фигуры. Начальная позиция. Німецька класична філософія 70-ті рр XVIIІ ст. (“коперніканський переворот”)
Німецька класична філософія 70-ті рр XVIIІ ст. (“коперніканський переворот”) Онлайн курс английского языка. Ценностное предложение и УТП
Онлайн курс английского языка. Ценностное предложение и УТП Классный час: Государственные символы РОССИИ. Электронная презентация.
Классный час: Государственные символы РОССИИ. Электронная презентация. Проект: Мир сказок К. И. Чуковского. (средняя группа)
Проект: Мир сказок К. И. Чуковского. (средняя группа) Группа компаний Комитет.Орг. Hr-мероприятия. Деловые события и форумы
Группа компаний Комитет.Орг. Hr-мероприятия. Деловые события и форумы Музыкальная драматургия
Музыкальная драматургия Выселковский район. Вчера, сегодня, завтра.
Выселковский район. Вчера, сегодня, завтра. Иудаизм
Иудаизм Презентация Зимние олимпийские игры в Сочи
Презентация Зимние олимпийские игры в Сочи Знание научное и знание художественное
Знание научное и знание художественное Возбуждение и рассмотрение дела об административном правонарушении
Возбуждение и рассмотрение дела об административном правонарушении Силачи и слабаки в мире кислот и оснований.
Силачи и слабаки в мире кислот и оснований. Интеллектуально-развлекательная игра Морской бой по ПДД
Интеллектуально-развлекательная игра Морской бой по ПДД Συνδρομο υπνικης απνοιας
Συνδρομο υπνικης απνοιας ТЕАТРАЛИЗОВАННАЯ ДЕЯТЕЛЬНОСТЬ – ОДНО ИЗ ПЕРСПЕКТИВНЫХ НАПРАВЛЕНИЙ РАЗВИТИЯ СОВРЕМЕННОГО ДОШКОЛЬНОГО ОБРАЗОВАНИЯ
ТЕАТРАЛИЗОВАННАЯ ДЕЯТЕЛЬНОСТЬ – ОДНО ИЗ ПЕРСПЕКТИВНЫХ НАПРАВЛЕНИЙ РАЗВИТИЯ СОВРЕМЕННОГО ДОШКОЛЬНОГО ОБРАЗОВАНИЯ Peek-a-Boo
Peek-a-Boo День России
День России Цифро-аналоговый преобразователь: структурная схема и принцип действия
Цифро-аналоговый преобразователь: структурная схема и принцип действия Социальный состав населения России на 1897 год
Социальный состав населения России на 1897 год Германия (11 класс)
Германия (11 класс) Сестринский процесс при эпидемическом паротите, ветряной оспе
Сестринский процесс при эпидемическом паротите, ветряной оспе Экономическая политика Петра I
Экономическая политика Петра I