- Кинетика ферментативного катализа. (Лекция 5)

Содержание
- 2. Методы определения количества ферментов Наиболее часто используемые: Колориметрические - основаны на определении образующихся в ходе реакции
- 3. 2. Способы выражения активности ферментов. Используются 2 основные единицы: КАТАЛ – такое количество фермента, которое может
- 4. 3. Кинетика ферментативных реакций Ферментативная кинетика занимается исследованием закономерностей влияния химической природы реагирующих веществ (ферментов, субстратов)
- 5. Зависимость скорости от вида субстрата. Ферменты обладают избирательностью действия - специфичность действия: 1 – Абсолютная специфичность.
- 6. 2 – Относительная специфичность ( объясняется тем, что, активный центр ферментов, обладаю-щих относительной специфичностью не жесткая
- 7. 2) Влияние [S] на скорость реакции. Теоретический график зависимости скорости ферментативной реакции от концентрации субстрата при
- 8. График Лайнуивера – Берка, построенный по методу двойных обратных величин. 1 / V0 1 / [S]
- 9. 3) Зависимость скорости реакции от концентрации фермента. 4) Зависимость скорости реакции от температуры. 4Х 3Х 2Х
- 10. 4) Зависимость скорости реакции от рН среды. В норме рН цитозоля =7,2 1 2 3 4
- 11. Активность ферментов при различных рН 1 2 3 4 5 6 7 8 9 10 рН
- 12. Влияние различных веществ на активность ферментов АКТИВАТОРЫ ФЕРМЕНТОВ 1.1. Активация ферментов ионами металлов Ионы Mg+2, Mn+2,
- 13. Механизм активации ферментов металлами 1. В состав активного центра: Н2О + СО2 Н2СО3 Е + Ме
- 14. Активность амилазы в присутствии различных ионов Cl Br 1 2 3 4 5 6 7 8
- 15. Активация ферментов Ионами металлов Восстановленными соединениями S AH2 ( NADH2) SH E E S SH неактивный
- 16. Реакции ингибирования ферментативных процессов. ТИПЫ ИНГИБИРОВАНИЯ ФЕРМЕНТОВ I. Обратимое II. Необратимое Конкурентное Неконкурентное Бесконкуренетное Смешанного типа
- 17. 1. Конкурентный тип ингибирования Осуществляется веществом, близким по химическому строению к субстату. V max Vo [S]
- 18. 2. Неконкурентное торможение Ингибитор реагирует с ферментом иным образом , чем субстрат, и поэтому повышение концентрации
- 19. 3. Бесконкурентное торможение Ингибитор взаимодействует с фермент – субстратным комплексом. 4. Смешанный тип торможения Ингибитор взаимодействует
- 20. Ингибиторы взаимодействуют с ферментами различными путями, они могут: Блокировать активный центр фермента Менять четвертичную структуру фермента
- 21. Классификация ингибиторов ИНГИБИТОРЫ 1) специфические 2) неспецифические 3) Необратимого действия конкурентные неконкурентные 4) Обратимого действия 5)
- 22. Ингибирование сериновых гидролаз (АХЭ) диизопропилфторфосфатом (ДФФ) Остаток серина в активном центре ДФФ Каталитически неактивный эфир Сериновые
- 23. Необратимое ингибирование Химически модифицированный фермент ( неакт. ) Йодацетамид (йодацетат) Тиоловый фермент
- 24. Необратимое ингибирование Трансацилаза –один из ферментов, участвующих в биосинтезе клеточной стенки бактерий. - CH2OH + Трансацилаза
- 25. Необратимое ингибирование тиоловых ферментов Химически модифицированный фермент ( неакт. ) ТОКСИЧЕСКОЕ ДЕЙСТВИЕ Hg++ Pb++ , соединений
- 26. Неконкурентное ингибирование SH –групп ионами тяжелых металлов (Cu++, Hg++, Ag++, As++, Pb++) E – SH +
- 27. Неконкурентное ингибирование монотиоловых ферментов - SH - SH Активный Е + Cl As – CH =
- 28. Конкурентное ингибирование ПредшественникиТГФК Фолиевая кислота ТГФК Биоснтез ДНК, РНК Кофермент в биосинтезе пуринов и пиримидинов СУЛЬФАНИЛАМИД
- 29. Структуры основных ингибиторов Ахэ CH3 O CH3 – N – CH2 – CH2 – O –
- 30. Инактивация АХЭ
- 31. Реактивация АХЭ -I Пиридин-альдоксин-метил-йодид
- 32. Аллостерическое ингибирование Е I EI - комплекс + Аллостерический центр (НАДН2, АТФ) V max Vo [S]
- 33. Фосфатная модификация ферментов Е - ОН Е – О – РО3Н2 АТФ АДФ гликогенсинтаза Неактивный фермент
- 34. Ингибирование фермента путем ацетатной модификации PES - NH2 + Н3С-С-О || O COO- Простагландин-эндопироксид-синтаза (ее циклоге-назный
- 36. Скачать презентацию
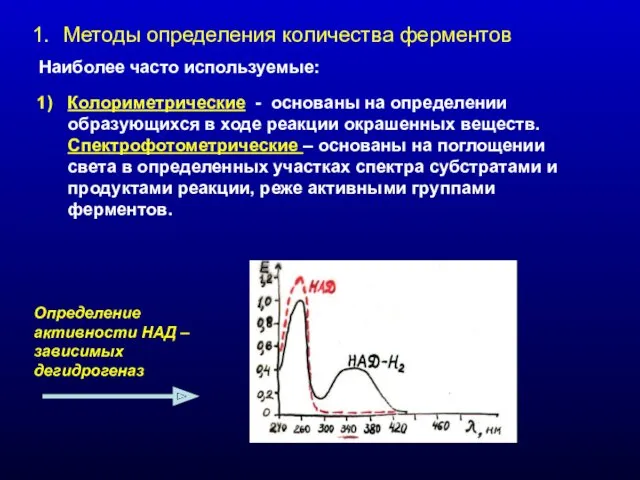
Методы определения количества ферментов
Наиболее часто используемые:
Колориметрические - основаны на определении образующихся
Методы определения количества ферментов
Наиболее часто используемые:
Колориметрические - основаны на определении образующихся

2. Способы выражения активности ферментов.
Используются 2 основные единицы:
КАТАЛ – такое количество
2. Способы выражения активности ферментов.
Используются 2 основные единицы:
КАТАЛ – такое количество

3. Кинетика ферментативных реакций
Ферментативная кинетика занимается исследованием закономерностей влияния химической природы
3. Кинетика ферментативных реакций
Ферментативная кинетика занимается исследованием закономерностей влияния химической природы

Зависимость скорости от вида субстрата.
Ферменты обладают избирательностью действия - специфичность действия:
1
Зависимость скорости от вида субстрата.
Ферменты обладают избирательностью действия - специфичность действия:
1

2 – Относительная специфичность
( объясняется тем, что, активный центр ферментов,
2 – Относительная специфичность ( объясняется тем, что, активный центр ферментов,
![2) Влияние [S] на скорость реакции. Теоретический график зависимости скорости](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/68467/slide-6.jpg)
2) Влияние [S] на скорость реакции.
Теоретический график зависимости скорости ферментативной реакции
2) Влияние [S] на скорость реакции.
Теоретический график зависимости скорости ферментативной реакции
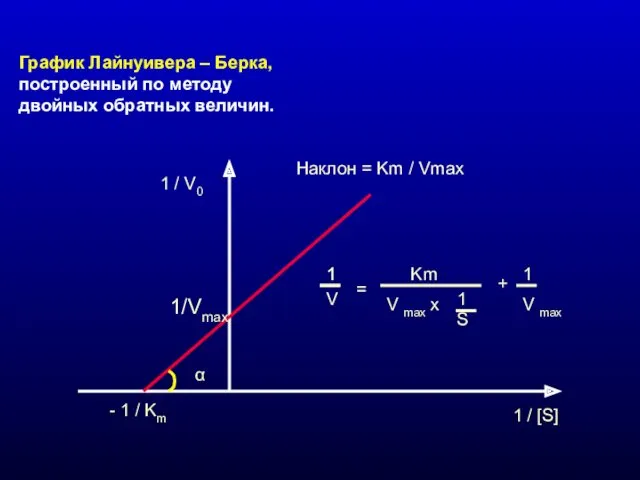
График Лайнуивера – Берка, построенный по методу двойных обратных величин.
1 /
График Лайнуивера – Берка, построенный по методу двойных обратных величин.
1 /
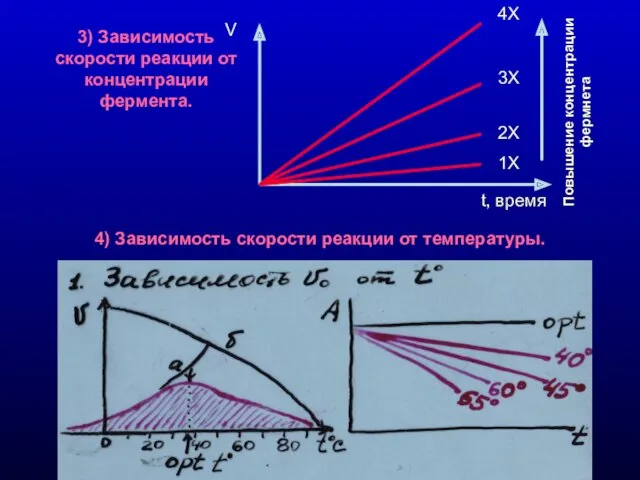
3) Зависимость скорости реакции от концентрации фермента.
4) Зависимость скорости реакции от
3) Зависимость скорости реакции от концентрации фермента.
4) Зависимость скорости реакции от
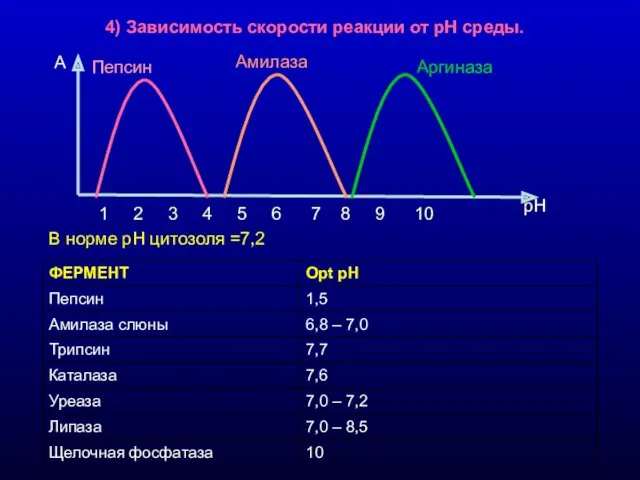
4) Зависимость скорости реакции от рН среды.
В норме рН цитозоля
4) Зависимость скорости реакции от рН среды.
В норме рН цитозоля
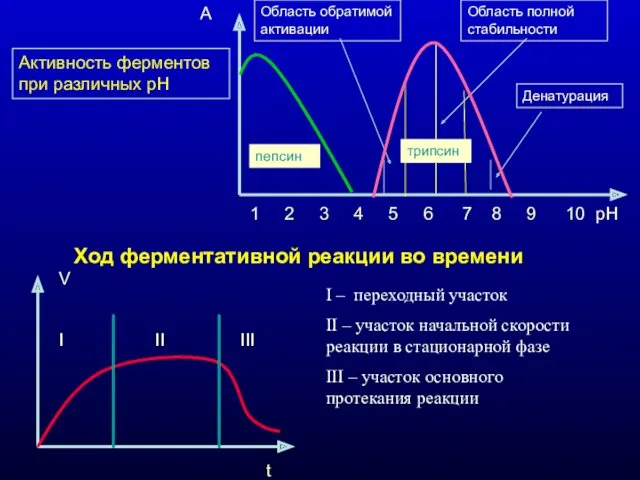
Активность ферментов при различных рН
1 2 3 4 5 6
Активность ферментов при различных рН
1 2 3 4 5 6

Влияние различных веществ на активность ферментов
АКТИВАТОРЫ ФЕРМЕНТОВ
1.1. Активация ферментов ионами металлов
Ионы
Влияние различных веществ на активность ферментов
АКТИВАТОРЫ ФЕРМЕНТОВ
1.1. Активация ферментов ионами металлов
Ионы

Механизм активации ферментов металлами
1. В состав активного центра:
Н2О + СО2 Н2СО3
Е
Механизм активации ферментов металлами
1. В состав активного центра:
Н2О + СО2 Н2СО3
Е
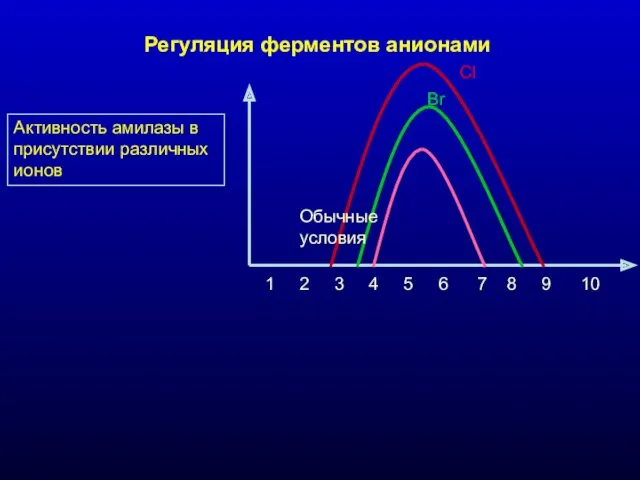
Активность амилазы в присутствии различных ионов
Cl
Br
1 2 3 4 5
Активность амилазы в присутствии различных ионов
Cl
Br
1 2 3 4 5
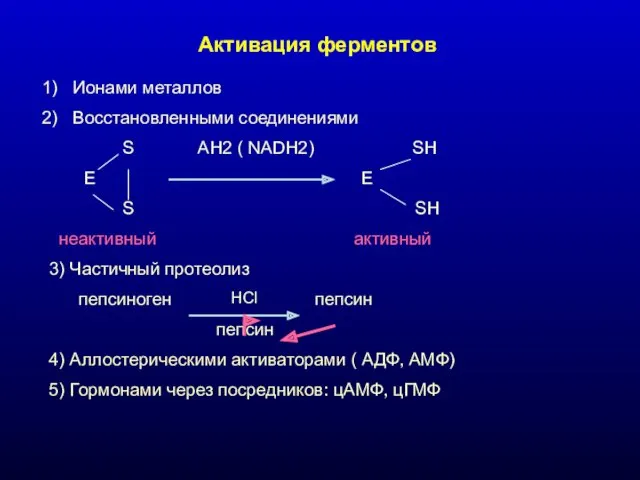
Активация ферментов
Ионами металлов
Восстановленными соединениями
S AH2 ( NADH2) SH
E E
Активация ферментов
Ионами металлов
Восстановленными соединениями
S AH2 ( NADH2) SH
E E
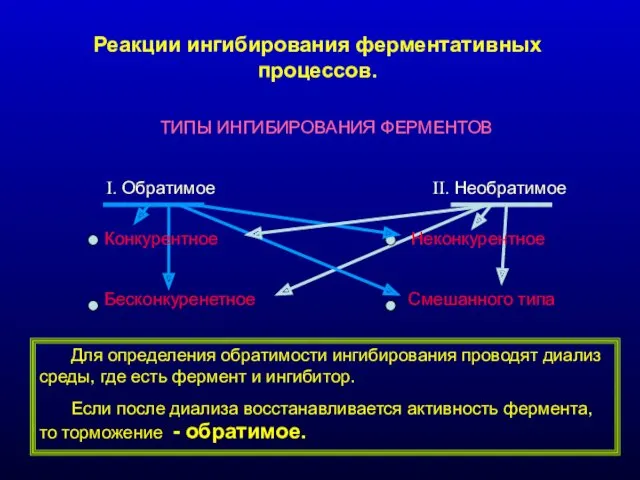
Реакции ингибирования ферментативных процессов.
ТИПЫ ИНГИБИРОВАНИЯ ФЕРМЕНТОВ
I. Обратимое II. Необратимое
Конкурентное Неконкурентное
Бесконкуренетное Смешанного
Реакции ингибирования ферментативных процессов.
ТИПЫ ИНГИБИРОВАНИЯ ФЕРМЕНТОВ
I. Обратимое II. Необратимое
Конкурентное Неконкурентное
Бесконкуренетное Смешанного
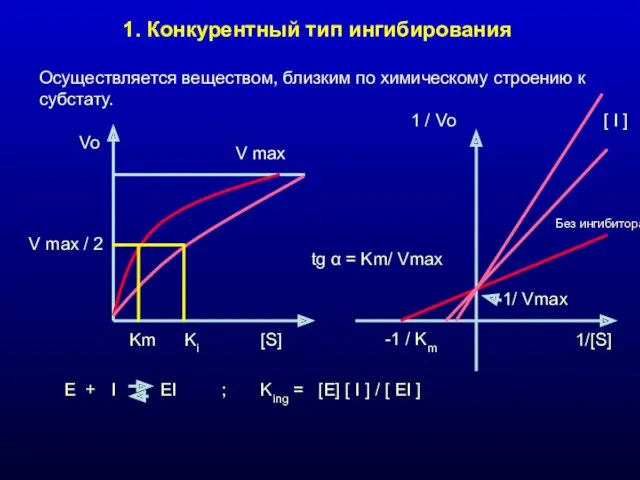
1. Конкурентный тип ингибирования
Осуществляется веществом, близким по химическому строению к субстату.
V
1. Конкурентный тип ингибирования
Осуществляется веществом, близким по химическому строению к субстату.
V
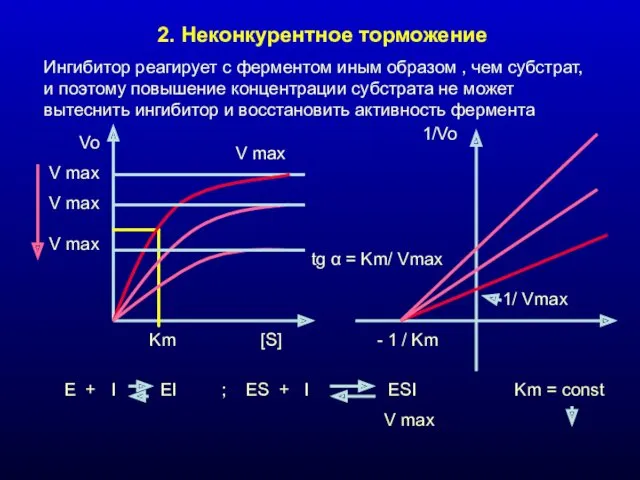
2. Неконкурентное торможение
Ингибитор реагирует с ферментом иным образом , чем субстрат,
2. Неконкурентное торможение
Ингибитор реагирует с ферментом иным образом , чем субстрат,
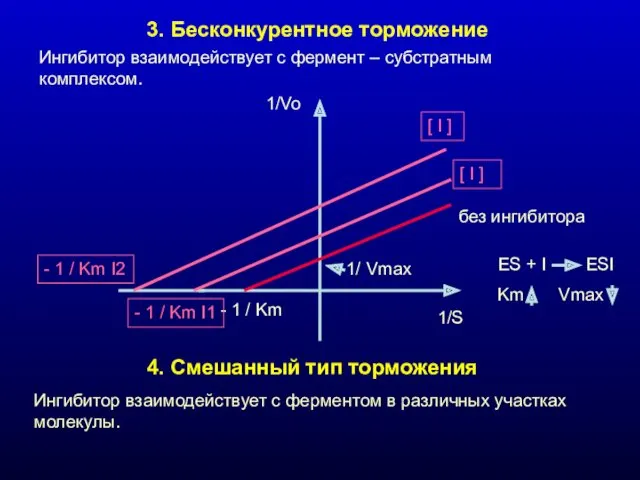
3. Бесконкурентное торможение
Ингибитор взаимодействует с фермент – субстратным комплексом.
4. Смешанный тип
3. Бесконкурентное торможение
Ингибитор взаимодействует с фермент – субстратным комплексом.
4. Смешанный тип

Ингибиторы взаимодействуют с ферментами различными путями, они могут:
Блокировать активный центр
Ингибиторы взаимодействуют с ферментами различными путями, они могут:
Блокировать активный центр
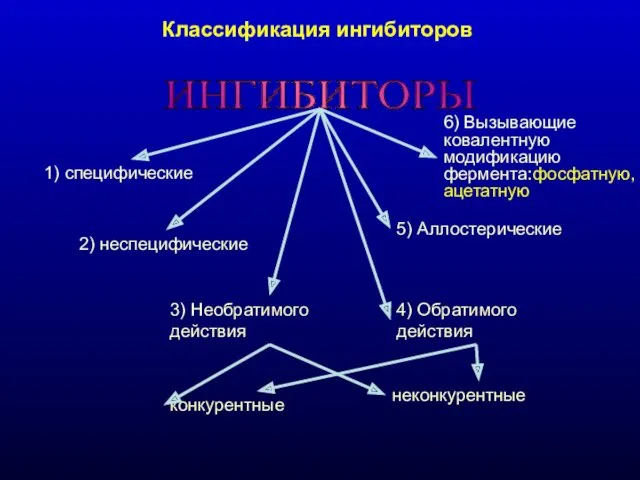
Классификация ингибиторов
ИНГИБИТОРЫ
1) специфические
2) неспецифические
3) Необратимого действия
конкурентные
неконкурентные
4) Обратимого действия
5) Аллостерические
6) Вызывающие
Классификация ингибиторов
ИНГИБИТОРЫ
1) специфические
2) неспецифические
3) Необратимого действия
конкурентные
неконкурентные
4) Обратимого действия
5) Аллостерические
6) Вызывающие
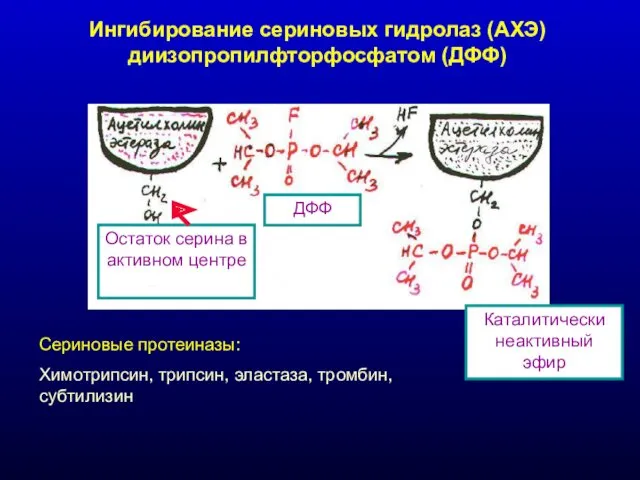
Ингибирование сериновых гидролаз (АХЭ) диизопропилфторфосфатом (ДФФ)
Остаток серина в активном центре
ДФФ
Каталитически неактивный
Ингибирование сериновых гидролаз (АХЭ) диизопропилфторфосфатом (ДФФ)
Остаток серина в активном центре
ДФФ
Каталитически неактивный
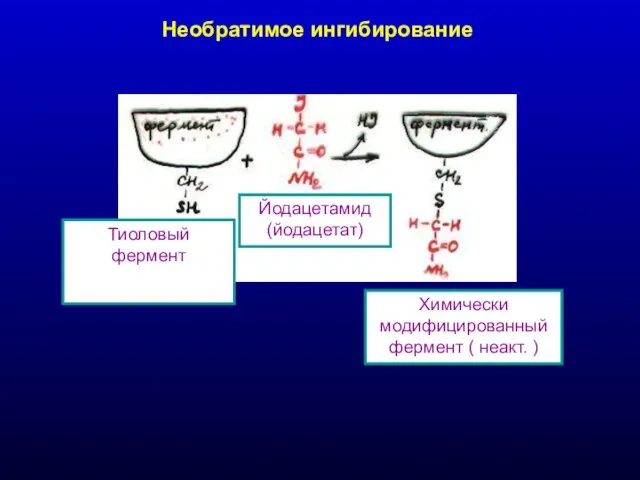
Необратимое ингибирование
Химически модифицированный фермент ( неакт. )
Йодацетамид (йодацетат)
Тиоловый фермент
Необратимое ингибирование
Химически модифицированный фермент ( неакт. )
Йодацетамид (йодацетат)
Тиоловый фермент
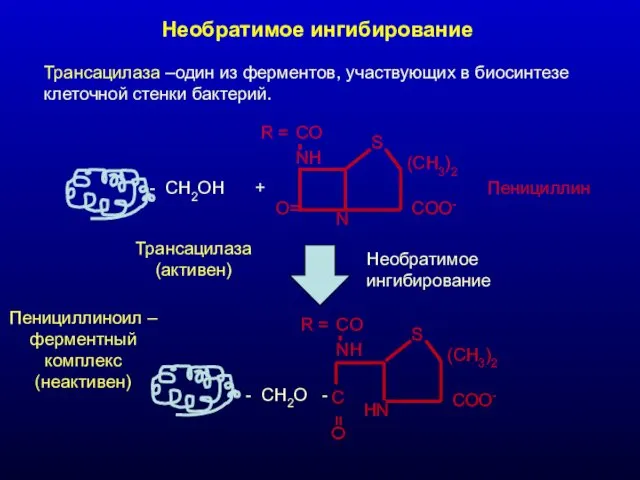
Необратимое ингибирование
Трансацилаза –один из ферментов, участвующих в биосинтезе клеточной стенки
Необратимое ингибирование
Трансацилаза –один из ферментов, участвующих в биосинтезе клеточной стенки
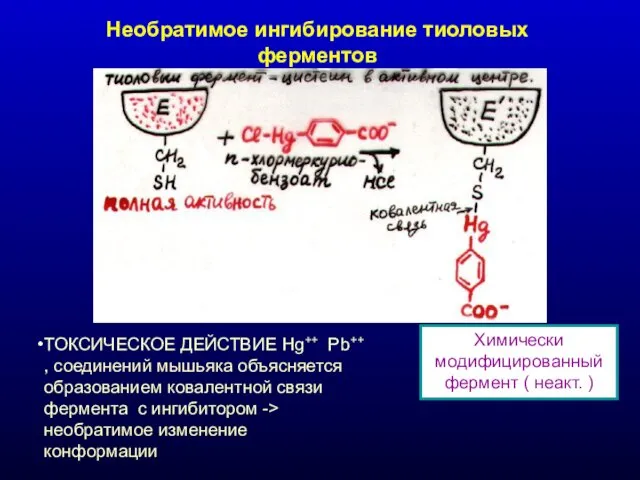
Необратимое ингибирование тиоловых ферментов
Химически модифицированный фермент ( неакт. )
ТОКСИЧЕСКОЕ ДЕЙСТВИЕ Hg++
Необратимое ингибирование тиоловых ферментов
Химически модифицированный фермент ( неакт. )
ТОКСИЧЕСКОЕ ДЕЙСТВИЕ Hg++
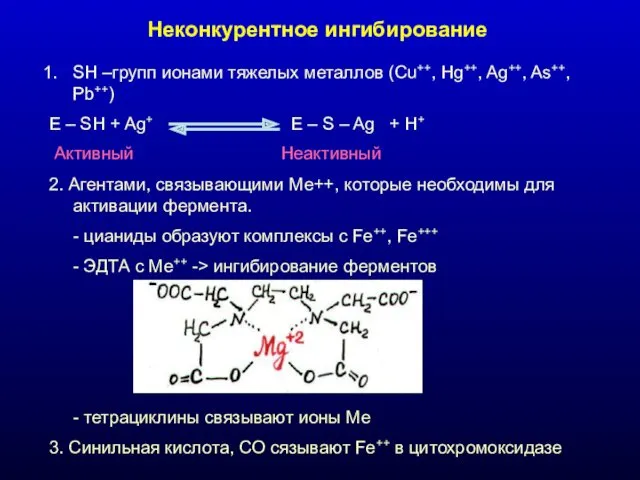
Неконкурентное ингибирование
SH –групп ионами тяжелых металлов (Cu++, Hg++, Ag++, As++, Pb++)
E
Неконкурентное ингибирование
SH –групп ионами тяжелых металлов (Cu++, Hg++, Ag++, As++, Pb++)
E
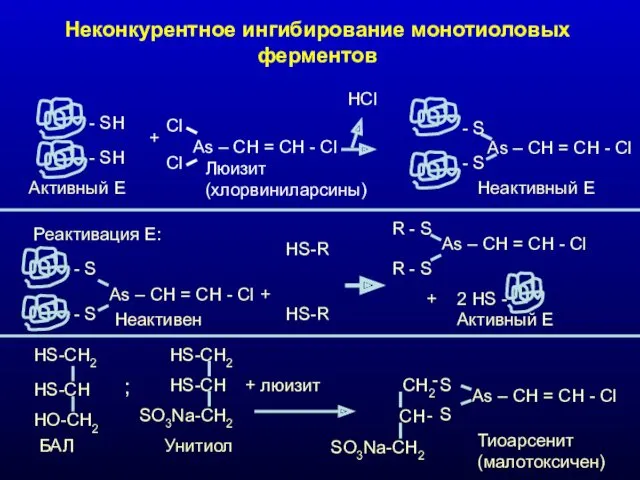
Неконкурентное ингибирование монотиоловых ферментов
- SH
- SH
Активный Е
+
Cl
As – CH
Неконкурентное ингибирование монотиоловых ферментов
- SH
- SH
Активный Е
+
Cl
As – CH
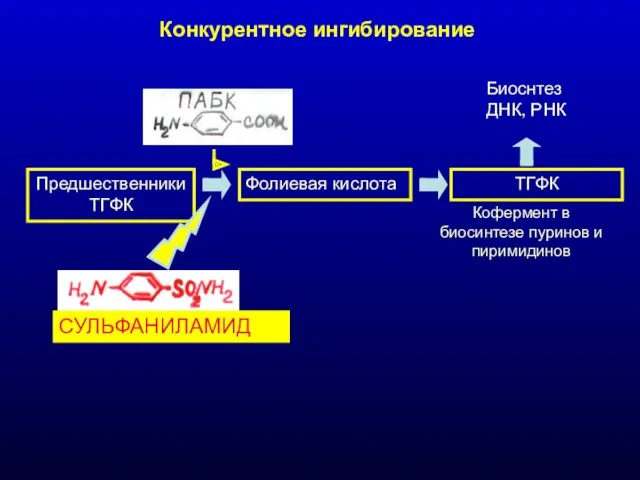
Конкурентное ингибирование
ПредшественникиТГФК
Фолиевая кислота
ТГФК
Биоснтез ДНК, РНК
Кофермент в биосинтезе пуринов и пиримидинов
СУЛЬФАНИЛАМИД
Конкурентное ингибирование
ПредшественникиТГФК
Фолиевая кислота
ТГФК
Биоснтез ДНК, РНК
Кофермент в биосинтезе пуринов и пиримидинов
СУЛЬФАНИЛАМИД
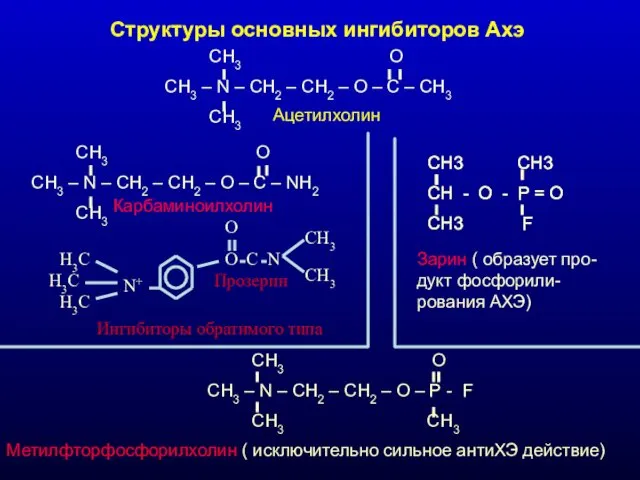
Структуры основных ингибиторов Ахэ
CH3 O
CH3 – N –
Структуры основных ингибиторов Ахэ
CH3 O
CH3 – N –
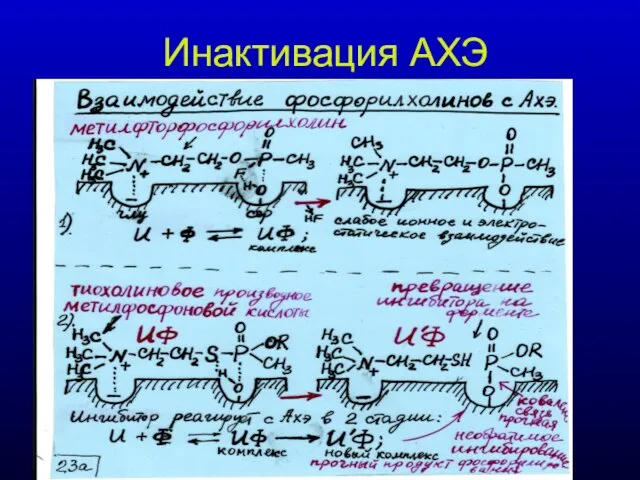
Инактивация АХЭ
Инактивация АХЭ
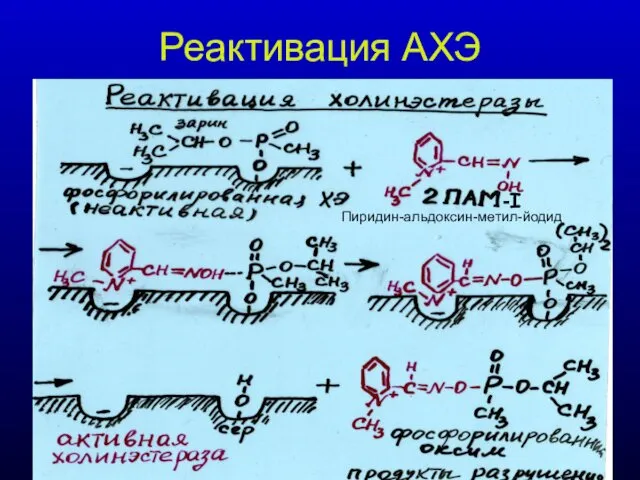
Реактивация АХЭ
-I
Пиридин-альдоксин-метил-йодид
Реактивация АХЭ
-I
Пиридин-альдоксин-метил-йодид
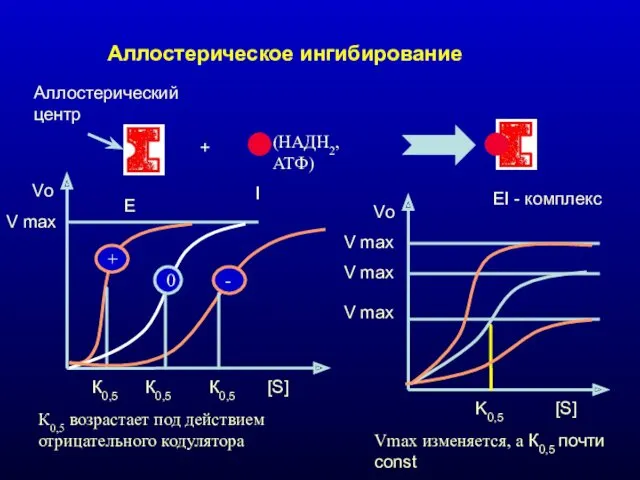
Аллостерическое ингибирование
Е
I
EI - комплекс
+
Аллостерический центр
(НАДН2, АТФ)
V max
Vo
[S]
К0,5
+
0
-
К0,5
К0,5
К0,5 возрастает под действием
Аллостерическое ингибирование
Е
I
EI - комплекс
+
Аллостерический центр
(НАДН2, АТФ)
V max
Vo
[S]
К0,5
+
0
-
К0,5
К0,5
К0,5 возрастает под действием

Фосфатная модификация ферментов
Е - ОН Е – О – РО3Н2
АТФ
АДФ
гликогенсинтаза
Неактивный
Фосфатная модификация ферментов
Е - ОН Е – О – РО3Н2
АТФ
АДФ
гликогенсинтаза
Неактивный
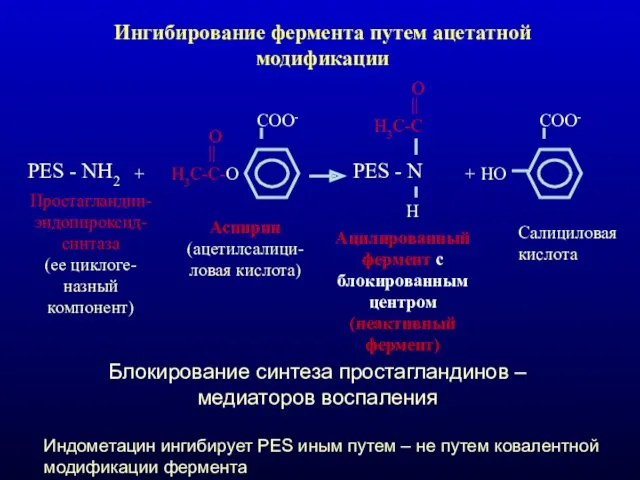
Ингибирование фермента путем ацетатной модификации
PES - NH2
+
Н3С-С-О
||
O
COO-
Простагландин-эндопироксид-синтаза (ее циклоге-назный компонент)
Аспирин (ацетилсалици-ловая
Ингибирование фермента путем ацетатной модификации
PES - NH2
+
Н3С-С-О
||
O
COO-
Простагландин-эндопироксид-синтаза (ее циклоге-назный компонент)
Аспирин (ацетилсалици-ловая
 Юбилей народного поэта к 110-летию Хасыра Сян-Белгина
Юбилей народного поэта к 110-летию Хасыра Сян-Белгина Презентация Экологическая паутина химии
Презентация Экологическая паутина химии Весёлые снеговики Зимняя физкультминутка.
Весёлые снеговики Зимняя физкультминутка.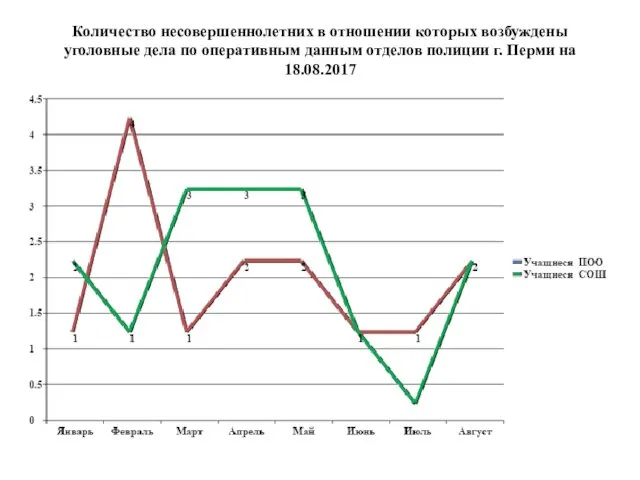 Количество несовершеннолетних, в отношении которых возбуждены уголовные дела по оперативным данным Перми на 18.08.2017
Количество несовершеннолетних, в отношении которых возбуждены уголовные дела по оперативным данным Перми на 18.08.2017 Макет рабочей программы воспитания при разработке и обновлении образовательных программ среднего профессионального образования
Макет рабочей программы воспитания при разработке и обновлении образовательных программ среднего профессионального образования Новшества культуры и быта при Петре I
Новшества культуры и быта при Петре I Новшества Windows 7
Новшества Windows 7 Таможенная стоимость, ее определение и контроль. (Лекция 5)
Таможенная стоимость, ее определение и контроль. (Лекция 5) Презентация Психологическая готовность к школе
Презентация Психологическая готовность к школе Гидрологические характеристики и гидрологическое состояние водного объекта
Гидрологические характеристики и гидрологическое состояние водного объекта Шекә авылы каһарманнары
Шекә авылы каһарманнары Праздник Троица
Праздник Троица Водный режим котла
Водный режим котла Политическая культура
Политическая культура закрепление
закрепление Магнітні матеріали. Класифікація магнітних матеріалів. Магнітом'які матеріали для постійних і низькочастотних магнітних полів
Магнітні матеріали. Класифікація магнітних матеріалів. Магнітом'які матеріали для постійних і низькочастотних магнітних полів Мероприятие в шатре. Аренда шатра, мебели, текстиля, оборудования
Мероприятие в шатре. Аренда шатра, мебели, текстиля, оборудования Основания и фундаменты исторических зданий
Основания и фундаменты исторических зданий Государственно-религиозные отношения
Государственно-религиозные отношения Результат усвоения систематизированных знаний, умений и навыков
Результат усвоения систематизированных знаний, умений и навыков Презентация Этикет
Презентация Этикет Лагеря смерти нацистской Германии
Лагеря смерти нацистской Германии Мы - Атланты. Презентация нашего детского объединения. Диск
Мы - Атланты. Презентация нашего детского объединения. Диск презентация Здоровьесбережение в воспитательном процессе
презентация Здоровьесбережение в воспитательном процессе Оценивание закономерностей окислительного изнашивания
Оценивание закономерностей окислительного изнашивания Внеурочная деятельность
Внеурочная деятельность Системы передачи измерительной информации - сельсинные
Системы передачи измерительной информации - сельсинные Перегородки
Перегородки