- МОЗГ: баланс ГАМК и глутамата

Содержание
- 2. Главными медиаторами ЦНС и головного мозга являются аминокислоты. Глутаминовая кислота (Glu) – главный возбуждающий меди-атор (около
- 3. Глутаминовая кислота и ГАМК – конкуренты и партнёры. При нарушении баланса Glu и ГАМК: синдром дефицита
- 4. Глутаминовая кислота. Пищевая аминокислота – входит в состав белков пищи и белков нашего тела; самая распростра-
- 5. R Другой («быстрый») путь синтеза, характерный для нейронов: взаимные превращения Glu и очень сходной пищевой а/к
- 6. R Другой («быстрый») путь синтеза, характерный для нейронов: взаимные превращения Glu и очень сходной пищевой а/к
- 7. Всех тканях организма (в т.ч. в мозге) очень много Glu. В связи с этим долгое время
- 8. Всех тканях организма (в т.ч. в мозге) очень много Glu. В связи с этим долгое время
- 9. Глутамат, помимо действия на рецепторы постсинаптической мембра-ны, способен влиять на вкусовые клетки-рецепторы языка («вкус белка»). Существуют
- 10. Рецепторы глутамата. В настоящее время выделяют три типа ионотропных и не менее пяти типов метаботропных рецепторов
- 11. NMDA-рецепторы (агонист N-метил-D-аспартат) АМРА-рецепторы (агонист амино-гидрокси-метил- изоксазол-пропионовая кислота) Каинатные рецепторы (агонист каиновая кислота) Рецепторы разных типов
- 12. Наиболее изучены NMDA- рецепторы. Каждый такой рец-р состоит из 4-х белковых молекул; в открытом положении он
- 13. Наиболее очевидный способ удаления Mg2+: ПД, запущенный с помощью не-NMDA-рецепторов (например, АМРА), находящихся на той же
- 14. Уникальная особенность NMDA-рецепторов состоит в том, что их канал может блокировать ион Mg2+ («магниевая пробка»). В
- 15. Более длительные изменения обеспечиваются передачей сигнала посредством ионов Са2+ на ядерную ДНК, активацией генов не-NMDA-рецепторов, синтезом
- 16. Пусть мы учим крысу прыгать на полку в ответ на звонок (иначе она получает удар электрическим
- 17. 2 дополнит. схемы, где показаны упомянутые выше пластические изменения: выбивание Mg2+-пробок, фосфорилирование АМРА-рецеп-торов, влияние на гены
- 18. Более широкое распространение в клинике продолжают сохранять агонисты ГАМК – препараты, «проверенные временем». Но сначала –
- 19. Инактивация Glu: Из синаптич. щели Glu ● переносится в глиальные клетки, где превращается в глутамин (Gln)
- 20. ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается не столько торможение ЦНС, сколько
- 21. ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается не столько торможение ЦНС, сколько
- 22. Ноотропы – производные ГАМК, улучшают выработку энергии нейронами (в сутки по 0.5-3 г). Но есть и
- 23. Рецепторы ГАМК. Выделяют два основных типа рецепторов ГАМК: (GABA – gamma-aminobutyric acid) ГАМКА – ионотропные, из
- 24. Антагонисты ГАМК: вызывают судороги. Бикукулин – мешает ГАМК присо-единяться к рецептору А-типа; токсин североамериканского травянистого
- 25. Агонисты ГАМКА-рецептора: барбитураты и бензодиазепины. Барбитураты: открыты более 100 лет назад в день Св. Варвары. продолжительное
- 26. Барбитураты: открыты более 100 лет назад в день Св. Варвары. продолжительное общее тормозящ. действие; используются для
- 27. Инактивация ГАМК. В значительной мере сходна с инактивацией Glu: обратное всасывание ГАМК в пресинаптическое окончание либо
- 28. Вальпроаты (соли вальпроевой кис-лоты) – блокаторы ГАМК трансферазы, усиливают тормозные процессы в ЦНС и, наряду с
- 29. Транквилизаторы, анксиолитики, успокаивающие средства – группа лек. препаратов (в основном агонистов ГАМК), снижающих активность центров заднего
- 30. Тестирование транквилизаторов на эксперимен-тальных животных Тестирование потен-циальных лекарств на животных – обязательный этап перед их внедрением
- 31. Сон и наркоз. Мозговые центры, регулирующие смену сна и бодрствования – одни из самых древних структур
- 32. Сон и наркоз. Мозговые центры, регулирующие смену сна и бодрствования – одни из самых древних структур
- 33. Эпилепсия. Около 0.5% населения (предрасположены ~2%). В большинстве случаев в мозге больного есть зона постоянной активации
- 34. Эпилепсия. Около 0.5% населения (предрасположены ~2%). В большинстве случаев в мозге больного есть зона постоянной активации
- 35. 5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50% взрослых; наиболее эффективны психотерапия +
- 36. 5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50% взрослых; наиболее эффективны психотерапия +
- 38. Скачать презентацию

Главными медиаторами ЦНС и головного мозга являются аминокислоты.
Глутаминовая кислота (Glu) –
Главными медиаторами ЦНС и головного мозга являются аминокислоты.
Глутаминовая кислота (Glu) –

Глутаминовая кислота и ГАМК –
конкуренты и партнёры.
При нарушении баланса Glu
Глутаминовая кислота и ГАМК –
конкуренты и партнёры.
При нарушении баланса Glu
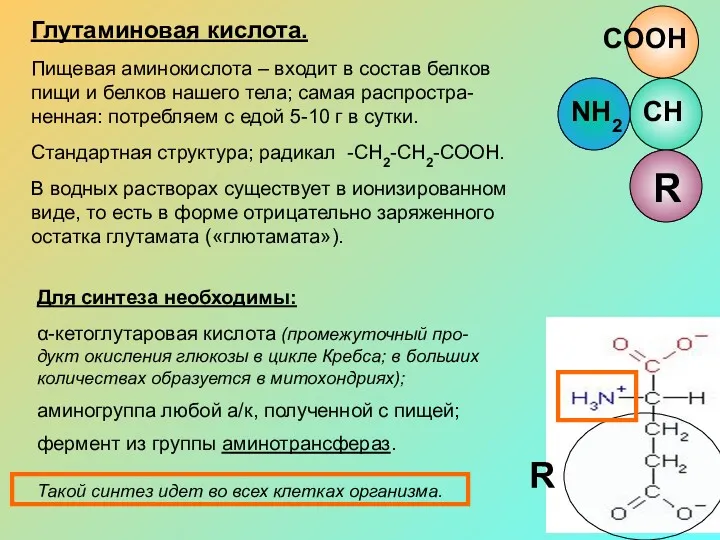
Глутаминовая кислота.
Пищевая аминокислота – входит в состав белков пищи и белков
Глутаминовая кислота.
Пищевая аминокислота – входит в состав белков пищи и белков

R
Другой («быстрый») путь синтеза, характерный для нейронов:
взаимные превращения Glu и очень
R
Другой («быстрый») путь синтеза, характерный для нейронов:
взаимные превращения Glu и очень

R
Другой («быстрый») путь синтеза, характерный для нейронов:
взаимные превращения Glu и очень
R
Другой («быстрый») путь синтеза, характерный для нейронов:
взаимные превращения Glu и очень
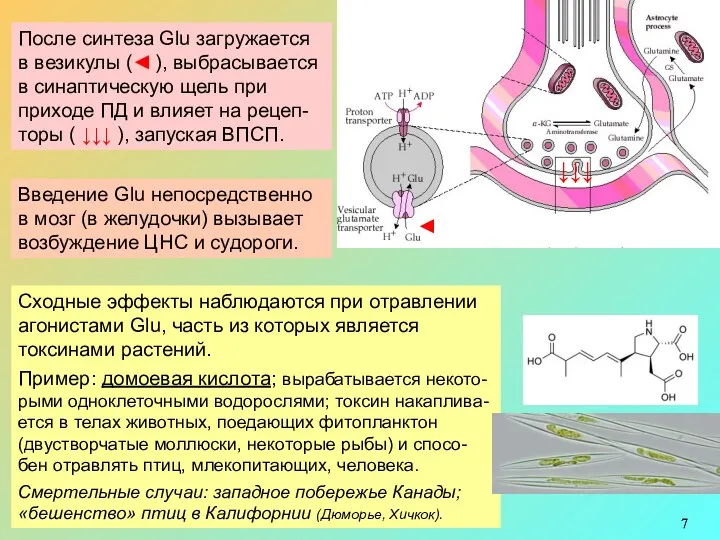
Всех тканях организма (в т.ч. в мозге) очень много Glu. В
Всех тканях организма (в т.ч. в мозге) очень много Glu. В
Всех тканях организма (в т.ч. в мозге) очень много Glu. В
Всех тканях организма (в т.ч. в мозге) очень много Glu. В
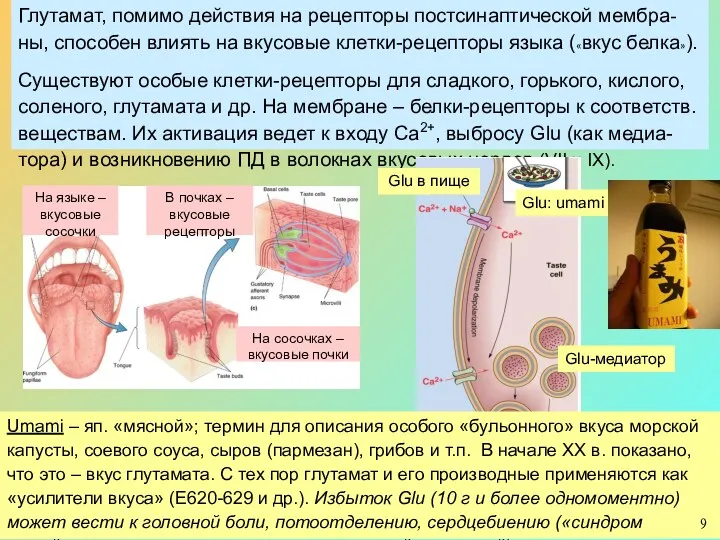
Глутамат, помимо действия на рецепторы постсинаптической мембра-ны, способен влиять на вкусовые
Глутамат, помимо действия на рецепторы постсинаптической мембра-ны, способен влиять на вкусовые

Рецепторы глутамата.
В настоящее время выделяют три типа ионотропных и не менее
Рецепторы глутамата.
В настоящее время выделяют три типа ионотропных и не менее

NMDA-рецепторы (агонист N-метил-D-аспартат)
АМРА-рецепторы (агонист амино-гидрокси-метил-
изоксазол-пропионовая кислота)
Каинатные рецепторы (агонист каиновая кислота)
Рецепторы разных
NMDA-рецепторы (агонист N-метил-D-аспартат)
АМРА-рецепторы (агонист амино-гидрокси-метил-
изоксазол-пропионовая кислота)
Каинатные рецепторы (агонист каиновая кислота)
Рецепторы разных
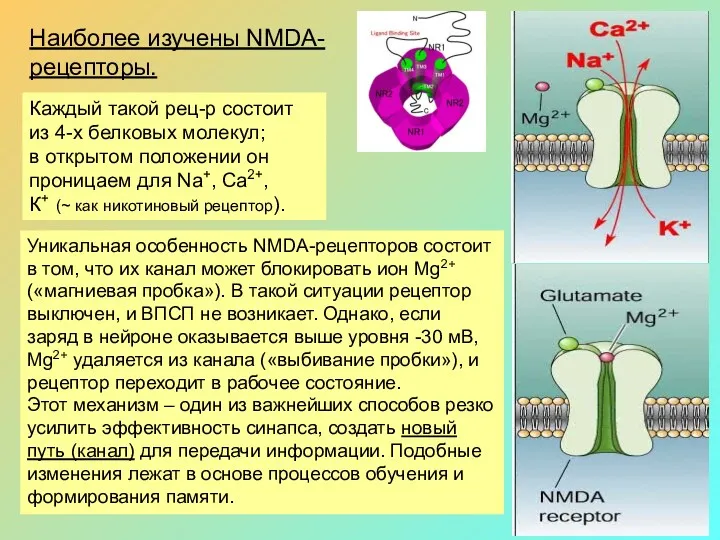
Наиболее изучены NMDA-
рецепторы.
Каждый такой рец-р состоит из 4-х белковых молекул;
в
Наиболее изучены NMDA-
рецепторы.
Каждый такой рец-р состоит из 4-х белковых молекул;
в
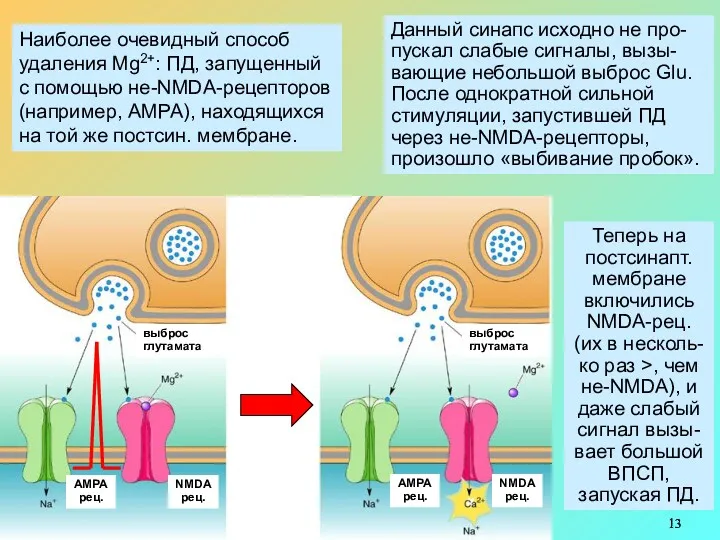
Наиболее очевидный способ
удаления Mg2+: ПД, запущенный с помощью не-NMDA-рецепторов (например, АМРА),
Наиболее очевидный способ
удаления Mg2+: ПД, запущенный с помощью не-NMDA-рецепторов (например, АМРА),

Уникальная особенность NMDA-рецепторов состоит в том, что их канал может блокировать
Уникальная особенность NMDA-рецепторов состоит в том, что их канал может блокировать
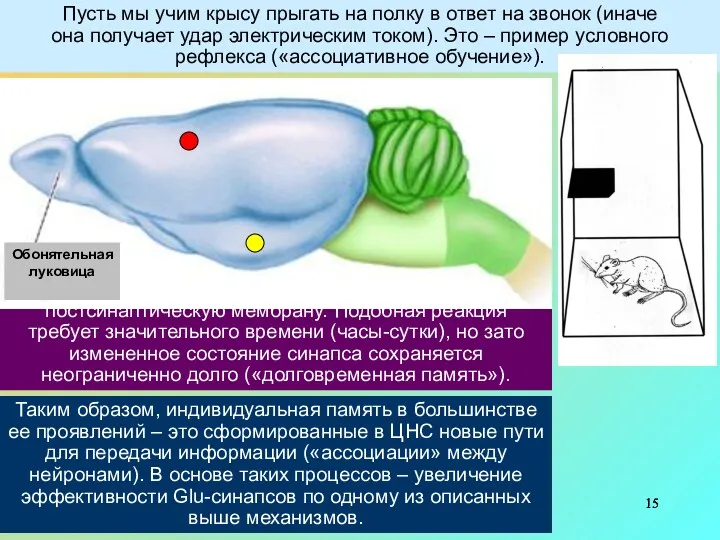
Более длительные изменения обеспечиваются передачей сигнала посредством ионов Са2+ на ядерную
Более длительные изменения обеспечиваются передачей сигнала посредством ионов Са2+ на ядерную
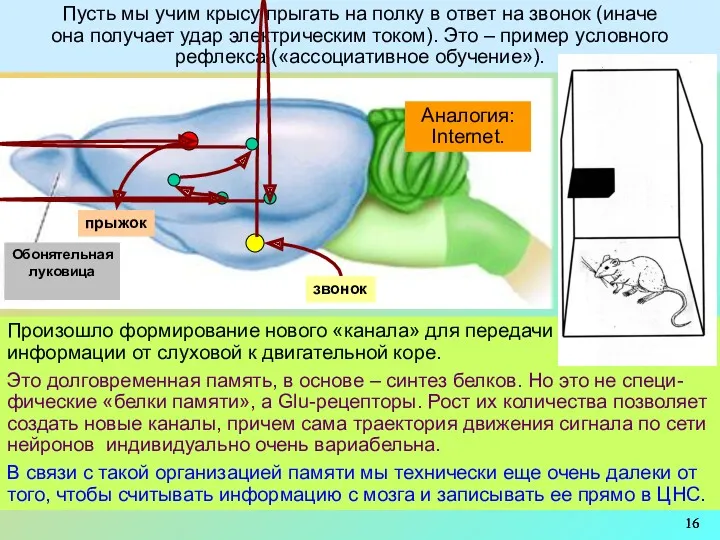
Пусть мы учим крысу прыгать на полку в ответ на звонок
Пусть мы учим крысу прыгать на полку в ответ на звонок
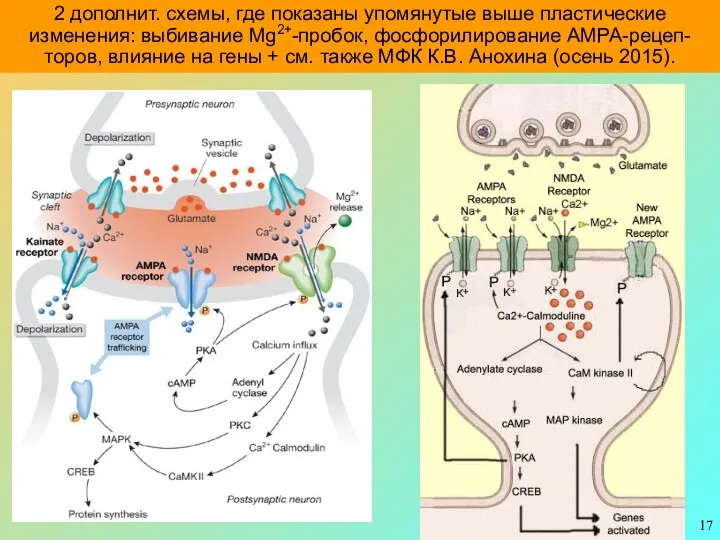
2 дополнит. схемы, где показаны упомянутые выше пластические изменения: выбивание Mg2+-пробок,
2 дополнит. схемы, где показаны упомянутые выше пластические изменения: выбивание Mg2+-пробок,

Более широкое распространение в клинике продолжают сохранять агонисты ГАМК – препараты,
Более широкое распространение в клинике продолжают сохранять агонисты ГАМК – препараты,
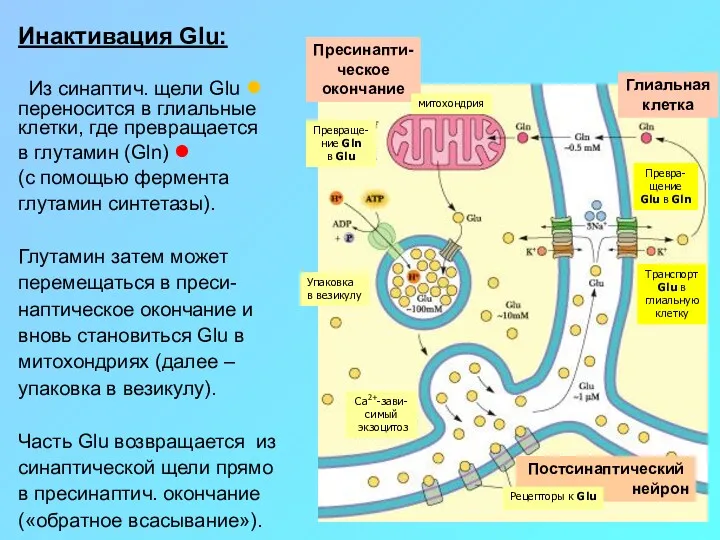
Инактивация Glu:
Из синаптич. щели Glu ● переносится в глиальные клетки, где
Инактивация Glu:
Из синаптич. щели Glu ● переносится в глиальные клетки, где
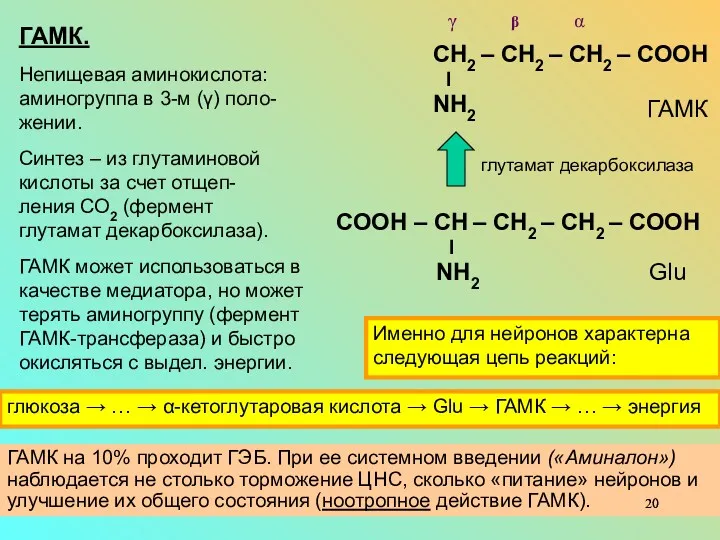
ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается
ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается
ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается
ГАМК на 10% проходит ГЭБ. При ее системном введении («Аминалон») наблюдается
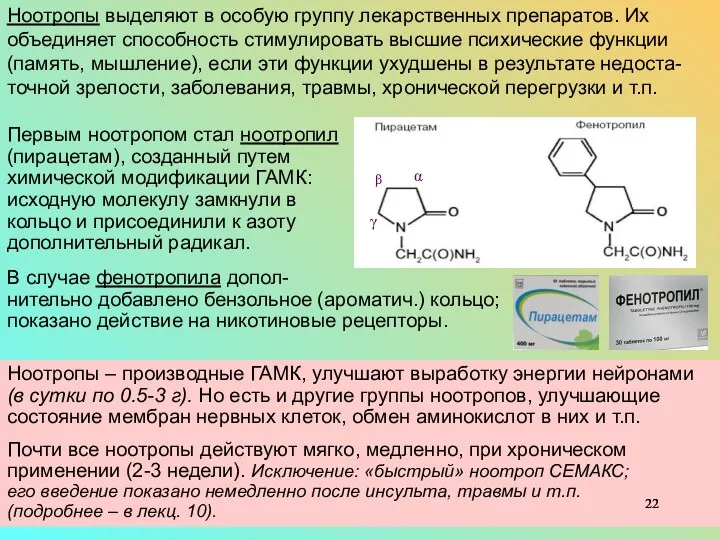
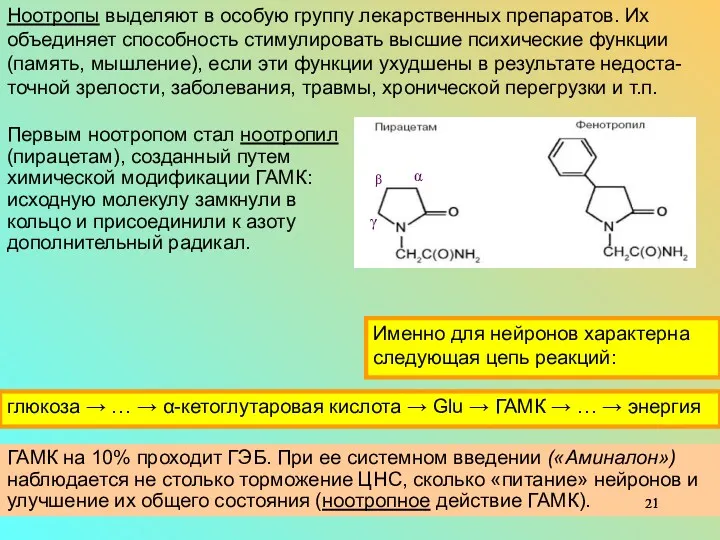
Ноотропы – производные ГАМК, улучшают выработку энергии нейронами (в сутки по
Ноотропы – производные ГАМК, улучшают выработку энергии нейронами (в сутки по
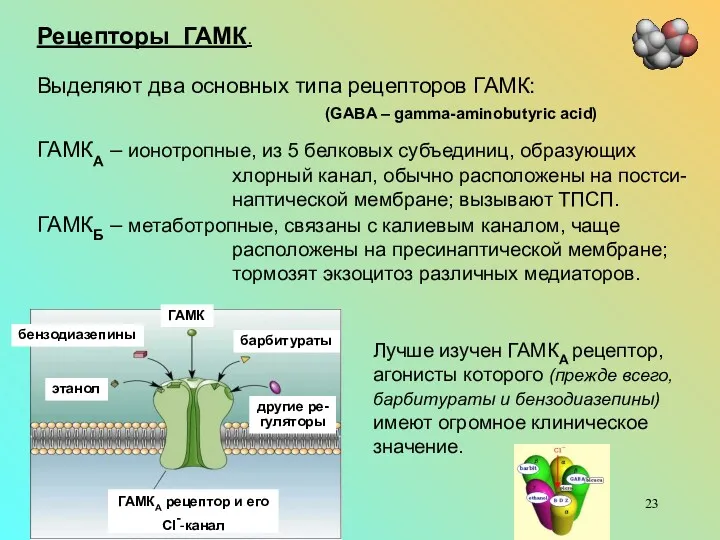
Рецепторы ГАМК.
Выделяют два основных типа рецепторов ГАМК:
(GABA – gamma-aminobutyric acid)
Рецепторы ГАМК.
Выделяют два основных типа рецепторов ГАМК:
(GABA – gamma-aminobutyric acid)

Антагонисты ГАМК: вызывают судороги.
Бикукулин – мешает ГАМК присо-единяться к рецептору А-типа;
Антагонисты ГАМК: вызывают судороги.
Бикукулин – мешает ГАМК присо-единяться к рецептору А-типа;

Агонисты ГАМКА-рецептора: барбитураты и бензодиазепины.
Барбитураты: открыты более 100 лет назад в
Агонисты ГАМКА-рецептора: барбитураты и бензодиазепины.
Барбитураты: открыты более 100 лет назад в

Барбитураты: открыты более 100 лет назад в день Св. Варвары.
продолжительное
Барбитураты: открыты более 100 лет назад в день Св. Варвары.
продолжительное
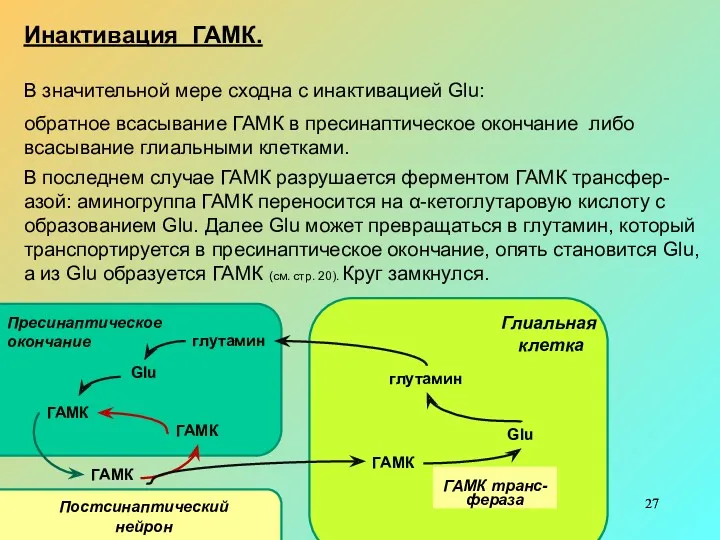
Инактивация ГАМК.
В значительной мере сходна с инактивацией Glu:
обратное всасывание ГАМК
Инактивация ГАМК.
В значительной мере сходна с инактивацией Glu:
обратное всасывание ГАМК
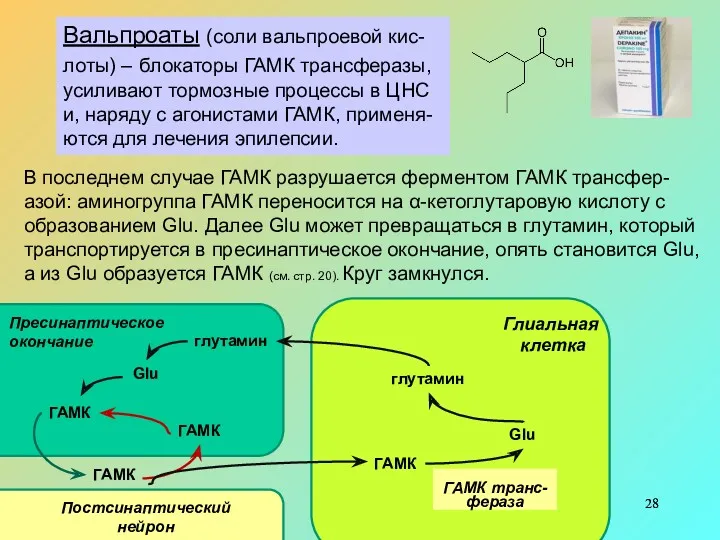
Вальпроаты (соли вальпроевой кис-лоты) – блокаторы ГАМК трансферазы, усиливают тормозные процессы
Вальпроаты (соли вальпроевой кис-лоты) – блокаторы ГАМК трансферазы, усиливают тормозные процессы

Транквилизаторы, анксиолитики, успокаивающие средства – группа лек. препаратов (в основном агонистов
Транквилизаторы, анксиолитики, успокаивающие средства – группа лек. препаратов (в основном агонистов

Тестирование транквилизаторов на эксперимен-тальных животных
Тестирование потен-циальных лекарств на животных – обязательный
Тестирование транквилизаторов на эксперимен-тальных животных
Тестирование потен-циальных лекарств на животных – обязательный
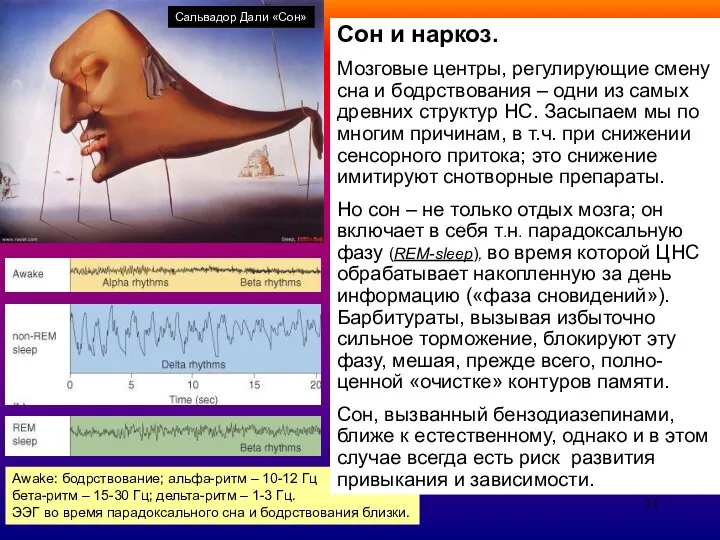
Сон и наркоз.
Мозговые центры, регулирующие смену сна и бодрствования – одни
Сон и наркоз.
Мозговые центры, регулирующие смену сна и бодрствования – одни

Сон и наркоз.
Мозговые центры, регулирующие смену сна и бодрствования – одни
Сон и наркоз.
Мозговые центры, регулирующие смену сна и бодрствования – одни
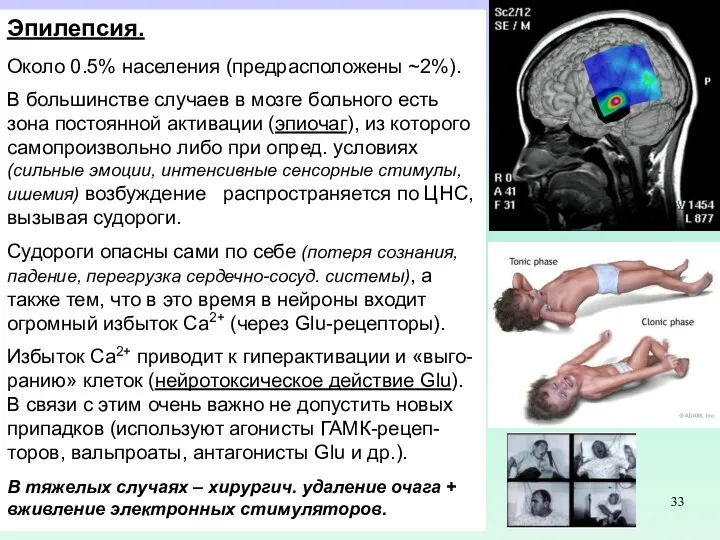
Эпилепсия.
Около 0.5% населения (предрасположены ~2%).
В большинстве случаев в мозге больного есть
Эпилепсия.
Около 0.5% населения (предрасположены ~2%).
В большинстве случаев в мозге больного есть

Эпилепсия.
Около 0.5% населения (предрасположены ~2%).
В большинстве случаев в мозге больного есть
Эпилепсия.
Около 0.5% населения (предрасположены ~2%).
В большинстве случаев в мозге больного есть

5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50%
5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50%

5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50%
5-15 % детей в возрасте 6-8 лет; симптомы сохраняются у 50%
 Повторялочка
Повторялочка Тошкент туманида дехқон хўжаликлари ва томорқа ер эгаларига бириктирилган экин ерларидан самарали фойдаланишни ташкил этиш
Тошкент туманида дехқон хўжаликлари ва томорқа ер эгаларига бириктирилган экин ерларидан самарали фойдаланишни ташкил этиш Презентация Наша Масленица
Презентация Наша Масленица Машины для приготовления бетонных и растворных смесей
Машины для приготовления бетонных и растворных смесей презентация программы творческого объединения Активисты школьного музея
презентация программы творческого объединения Активисты школьного музея Умный Дом. Базовый пакет для 1-2-х комнатных квартир
Умный Дом. Базовый пакет для 1-2-х комнатных квартир Реки окрестностей села Кузнецкого Диск
Реки окрестностей села Кузнецкого Диск Сборочные работы при различных видах соединений
Сборочные работы при различных видах соединений Трудности при овладении чтением учащимися и пути их преодоления.
Трудности при овладении чтением учащимися и пути их преодоления. Снегурочка. Кроссворд
Снегурочка. Кроссворд Феномен тоталитаризма: Италия и Германия. Часть 2
Феномен тоталитаризма: Италия и Германия. Часть 2 Шаблон Фракталы-5
Шаблон Фракталы-5 Занятия физкультурой на открытом воздухе, организация занятий
Занятия физкультурой на открытом воздухе, организация занятий Жостовские подносы
Жостовские подносы Презентация к родительскому собранию 2 класс Проектная деятельность
Презентация к родительскому собранию 2 класс Проектная деятельность Коммунизм. Марксизм, как наука. Старая новая политика, как временный отход
Коммунизм. Марксизм, как наука. Старая новая политика, как временный отход Организация проектирования и строительства горных предприятий. Лекция 2. Подготовительный период
Организация проектирования и строительства горных предприятий. Лекция 2. Подготовительный период Страховая пенсия по случаю потери кормильца
Страховая пенсия по случаю потери кормильца Благоустройсто сквера Нижнекисляйского городского поселения Нижний Кисляй Бутурлиновского муниципального района
Благоустройсто сквера Нижнекисляйского городского поселения Нижний Кисляй Бутурлиновского муниципального района Ғимараттарды жылумен қамдау жүйелері
Ғимараттарды жылумен қамдау жүйелері Moxito - узко-специализированный формат салона красоты по франшизе
Moxito - узко-специализированный формат салона красоты по франшизе Схемотехническое проектирование электронных приборов
Схемотехническое проектирование электронных приборов Морское право в правовой системе России
Морское право в правовой системе России Архитектура параллельных вычислительных систем. Часть 1. История и проблематика. Основы параллельного программирования
Архитектура параллельных вычислительных систем. Часть 1. История и проблематика. Основы параллельного программирования Виникнення та розвиток держави і права США
Виникнення та розвиток держави і права США Клинико-анатомические особенности строения зубочелюстной системы
Клинико-анатомические особенности строения зубочелюстной системы Какие вклады являются застрахованными
Какие вклады являются застрахованными развитие воображения у детей дошкольного возраста
развитие воображения у детей дошкольного возраста