- NK-клетки - большие гранулярные лимфоциты

Содержание
- 2. NK-клетки большие гранулярные лимфоциты, обладающие цитотоксичностью противопухолевых клеток и клеток, зараженных вирусами. В настоящее время NK-клетки
- 3. Характеристика Задача- выявлять и уничтожать собственные клетки организма, в которых что-то нарушилось. Составляют 5% лимфоцитов периферической
- 5. Маркёры NK-клетки не имеют основных маркёров Т- или B-лимфоцитов (поэтому их также называют нулевые лимфоциты), но
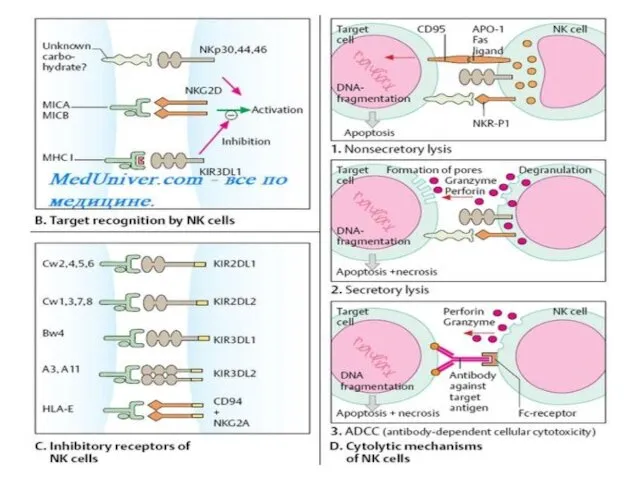
- 6. Рецепторы NK-клетки уничтожают клетку-мишень после установления с ней прямого контакта при помощи специальных белков — перфоринов.
- 8. Роль цитокинов Активность NK-клеток регулируют цитокины (у-ИФН и ИЛ-2 усиливают их цитолитическую активность). Наряду с макрофагами,
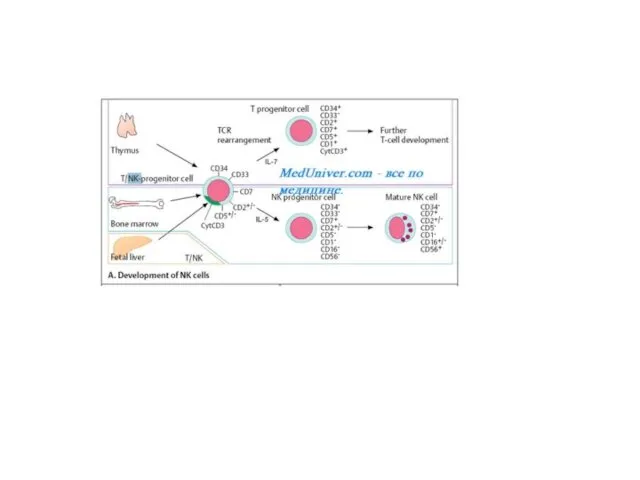
- 9. Развитие
- 10. Современные методы выделения лимфоцитов и других клеток
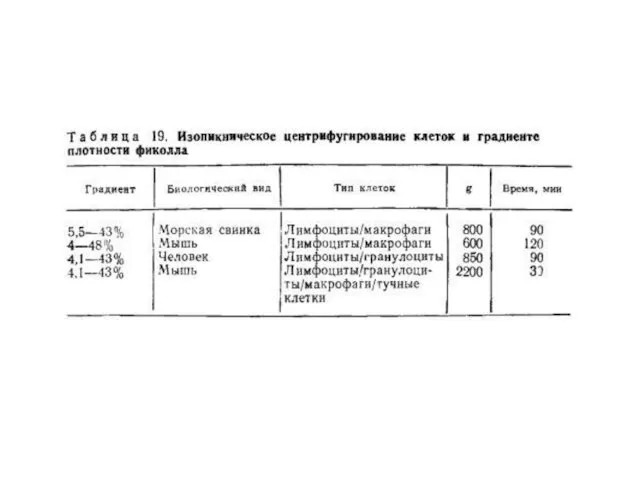
- 11. Для выделения мононуклеаров крови наиболее широкое распространение получил метод дифференциального центрифугирования в градиенте плотности фиколл-верографин (с
- 12. Для выделения моноцитов из суммарной фракции мононуклеаров самый простой и доступный метод основан на избирательной способности
- 13. Разделение клеток в градиенте плотности Материалы и оборудование. Для работы необходимы: центрифуга с охлаждением, бакет-ротор, градиентный
- 14. Центрифугирование в градиенте плотности Ввиду разнообразия применяемых методов фракционирования клеток в градиенте плотности невозможно в рамках
- 16. ступенчатый градиент плотности Одновременное разделение эозинофилов, нейтрофилов и моноцитов человека в градиенте фиколл/гипак: (A) 15,0 мл
- 17. Составляют градиент осторожно, наслаивая друг на друга по 2,0 мл растворов уменьшающейся плотности. Сверху наслаивают 2,0
- 18. Разделение гранулоцитов и фракции лимфоциты/моноциты человека в градиенте фиколл/триомбраст: (А) 10 объемов 34% триомбраста плотность 1,075
- 19. Изокинетическое разделение Изокинетическое разделение используют при необходимости разделять клетки одинаковой плотности, но различной величины. Если используется
- 20. Все многообразие методов разделения клеток можно видеть на примере разделения моноцитов и лимфоцитов человека в градиенте
- 21. Седиментация в градиенте плотности Фракционирование клеток различной величины чаще всего проводят в седиментационной камере по принципу
- 22. Идентификация T-лимфоцитов 1. Выделение чистых лимфоцитов в градиенте определенной плотности (различной для разных видов животных) методом
- 23. 4. Постановка реакции. К 0,25 мл рабочей взвеси лимфоцитов добавляют 0,25 мл 0,5%-ной взвеси отмытых эритроцитов.
- 24. 5. Готовят мазки па предметных стеклах методом толстой капли; фиксируют метиловым спиртом 3—5 мин и окрашивают
- 25. 6. Учет реакции. Под иммерсионной системой микроскопа подсчитывают 100 лимфоцитов (увеличение 90х7), неприсоединившие и присоединившие 3
- 27. Идентификация В-лимфоцитов 1. Эритроциты барана трижды отмывают раствором Хенкса и готовят 5%-ную взвесь в этом растворе.
- 28. 5. Отмытые эритроциты инкубируют с равным объемом комплемента при 37 °C в течение 30 мин и
- 30. Оценка функциональной активности NK-клеток с использованием проточной цитометрии
- 31. В качестве метки для КМ применяют флуоресцентный краситель 5-, 6-карбоксифлуоресцеин диацетатсукцинилмидиловый эфир (КФДЭ), который образует прочную
- 32. Первоначально нефлуоресцирующий КФДЭ проникает через клеточную мембрану. Карбоксифлуоресцеины связываются с внутриклеточными молекулами, формируя конъюгаты, обеспечивая стойкое
- 34. Скачать презентацию

NK-клетки
большие гранулярные лимфоциты, обладающие цитотоксичностью противопухолевых клеток и клеток, зараженных вирусами. В настоящее время
NK-клетки
большие гранулярные лимфоциты, обладающие цитотоксичностью противопухолевых клеток и клеток, зараженных вирусами. В настоящее время

Характеристика
Задача- выявлять и уничтожать собственные клетки организма, в которых что-то нарушилось.
Составляют
Характеристика
Задача- выявлять и уничтожать собственные клетки организма, в которых что-то нарушилось.
Составляют

Маркёры
NK-клетки не имеют основных маркёров Т- или B-лимфоцитов (поэтому их также
Маркёры
NK-клетки не имеют основных маркёров Т- или B-лимфоцитов (поэтому их также

Рецепторы
NK-клетки уничтожают клетку-мишень после установления с ней прямого контакта при помощи
Рецепторы
NK-клетки уничтожают клетку-мишень после установления с ней прямого контакта при помощи

Роль цитокинов
Активность NK-клеток регулируют цитокины (у-ИФН и ИЛ-2 усиливают их цитолитическую
Роль цитокинов
Активность NK-клеток регулируют цитокины (у-ИФН и ИЛ-2 усиливают их цитолитическую
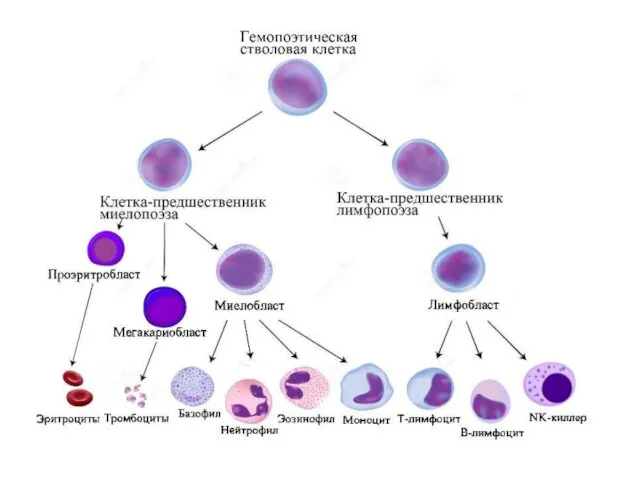
Развитие
Развитие

Современные методы выделения лимфоцитов и других клеток
Современные методы выделения лимфоцитов и других клеток

Для выделения мононуклеаров крови наиболее широкое распространение получил метод дифференциального центрифугирования
Для выделения мононуклеаров крови наиболее широкое распространение получил метод дифференциального центрифугирования

Для выделения моноцитов из суммарной фракции мононуклеаров самый простой и доступный
Для выделения моноцитов из суммарной фракции мононуклеаров самый простой и доступный

Разделение клеток в градиенте плотности
Материалы и оборудование. Для работы необходимы: центрифуга
Разделение клеток в градиенте плотности
Материалы и оборудование. Для работы необходимы: центрифуга

Центрифугирование в градиенте плотности
Ввиду разнообразия применяемых методов фракционирования клеток в градиенте
Центрифугирование в градиенте плотности
Ввиду разнообразия применяемых методов фракционирования клеток в градиенте

ступенчатый градиент плотности
Одновременное разделение эозинофилов, нейтрофилов и моноцитов человека в градиенте
ступенчатый градиент плотности
Одновременное разделение эозинофилов, нейтрофилов и моноцитов человека в градиенте

Составляют градиент осторожно, наслаивая друг на друга по 2,0 мл растворов
Составляют градиент осторожно, наслаивая друг на друга по 2,0 мл растворов

Разделение гранулоцитов и фракции лимфоциты/моноциты человека в градиенте фиколл/триомбраст:
(А) 10 объемов
Разделение гранулоцитов и фракции лимфоциты/моноциты человека в градиенте фиколл/триомбраст:
(А) 10 объемов

Изокинетическое разделение
Изокинетическое разделение используют при необходимости разделять клетки одинаковой плотности, но
Изокинетическое разделение
Изокинетическое разделение используют при необходимости разделять клетки одинаковой плотности, но

Все многообразие методов разделения клеток можно видеть на примере разделения моноцитов
Все многообразие методов разделения клеток можно видеть на примере разделения моноцитов

Седиментация в градиенте плотности
Фракционирование клеток различной величины чаще всего проводят в
Седиментация в градиенте плотности
Фракционирование клеток различной величины чаще всего проводят в

Идентификация T-лимфоцитов
1. Выделение чистых лимфоцитов в градиенте определенной плотности (различной для
Идентификация T-лимфоцитов
1. Выделение чистых лимфоцитов в градиенте определенной плотности (различной для

4. Постановка реакции. К 0,25 мл рабочей взвеси лимфоцитов добавляют 0,25
4. Постановка реакции. К 0,25 мл рабочей взвеси лимфоцитов добавляют 0,25

5. Готовят мазки па предметных стеклах методом толстой капли; фиксируют метиловым
5. Готовят мазки па предметных стеклах методом толстой капли; фиксируют метиловым

6. Учет реакции. Под иммерсионной системой микроскопа подсчитывают 100 лимфоцитов (увеличение
6. Учет реакции. Под иммерсионной системой микроскопа подсчитывают 100 лимфоцитов (увеличение
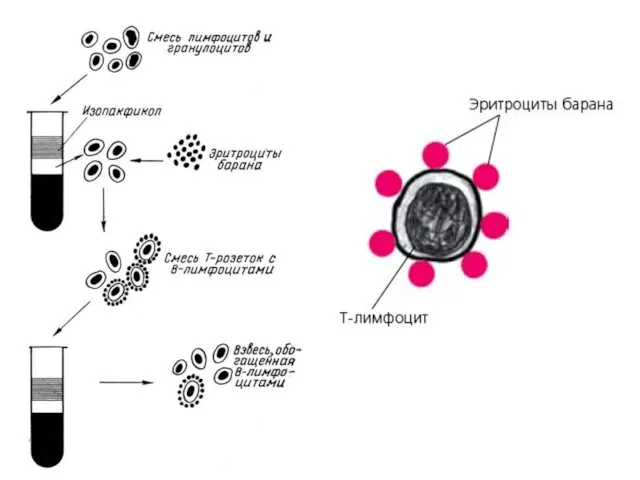

Идентификация В-лимфоцитов
1. Эритроциты барана трижды отмывают раствором Хенкса и готовят 5%-ную
Идентификация В-лимфоцитов
1. Эритроциты барана трижды отмывают раствором Хенкса и готовят 5%-ную

5. Отмытые эритроциты инкубируют с равным объемом комплемента при 37 °C
5. Отмытые эритроциты инкубируют с равным объемом комплемента при 37 °C
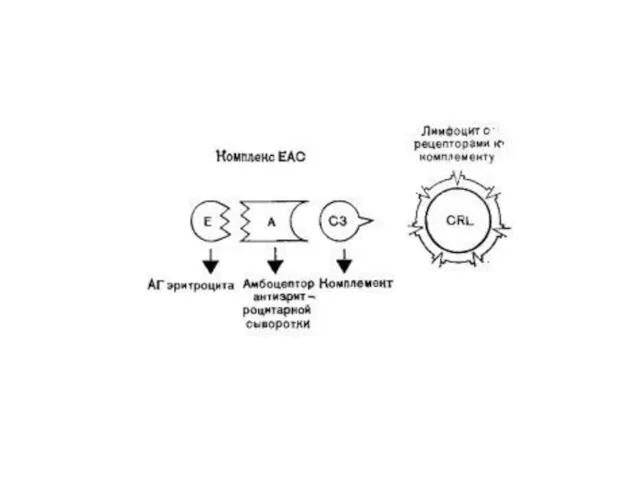

Оценка функциональной активности NK-клеток с использованием проточной цитометрии
Оценка функциональной активности NK-клеток с использованием проточной цитометрии

В качестве метки для КМ применяют флуоресцентный краситель 5-, 6-карбоксифлуоресцеин диацетатсукцинилмидиловый
В качестве метки для КМ применяют флуоресцентный краситель 5-, 6-карбоксифлуоресцеин диацетатсукцинилмидиловый

Первоначально нефлуоресцирующий КФДЭ проникает через клеточную мембрану. Карбоксифлуоресцеины связываются с внутриклеточными
Первоначально нефлуоресцирующий КФДЭ проникает через клеточную мембрану. Карбоксифлуоресцеины связываются с внутриклеточными
 Устройство трансформатор
Устройство трансформатор Wall Calendar 2017: NASA
Wall Calendar 2017: NASA Здоровье сберегающие технологии в ДОУ
Здоровье сберегающие технологии в ДОУ Игра- викторина Лучший пешеход
Игра- викторина Лучший пешеход Вазоренальная гипертензия
Вазоренальная гипертензия Возникновение христианства
Возникновение христианства Unusual weddings
Unusual weddings Электрооборудование пассажирских вагонов в поездах
Электрооборудование пассажирских вагонов в поездах Наша группа не простая, наша группа- речевая
Наша группа не простая, наша группа- речевая Машины постоянного тока
Машины постоянного тока Паспорт автоматизированной блочномодульной котельной АБМКУ-П-2,15
Паспорт автоматизированной блочномодульной котельной АБМКУ-П-2,15 Советы родителям первоклассников
Советы родителям первоклассников Липиды. (Лекция 10)
Липиды. (Лекция 10) Проект Агенство Визит
Проект Агенство Визит Презентация Гимн,герб, флаг
Презентация Гимн,герб, флаг Короткие замыкания
Короткие замыкания Презентация к итоговому занятию кружка В гостях у сказки
Презентация к итоговому занятию кружка В гостях у сказки Ручной и электрифицированный столярный инструмент. Столярная обработка древесины
Ручной и электрифицированный столярный инструмент. Столярная обработка древесины Маркетинг-план
Маркетинг-план Расточные резцы
Расточные резцы ФГОС как система обязательных требований.
ФГОС как система обязательных требований. Стандарт организации
Стандарт организации Геометрический смысл производной
Геометрический смысл производной Луна-естественный спутник Земли
Луна-естественный спутник Земли Интерактивный портал службы занятости населения Вологодской области. Инструкция для граждан на признание безработными
Интерактивный портал службы занятости населения Вологодской области. Инструкция для граждан на признание безработными Путешествие в страну сказок К.И. Чуковского. Сказка Айболит
Путешествие в страну сказок К.И. Чуковского. Сказка Айболит Модели и методы планирования и прогнозирования. Тема 3
Модели и методы планирования и прогнозирования. Тема 3 Презентация по мини-музею Домашние животные
Презентация по мини-музею Домашние животные