- Нуклеиновые кислоты. Лекция №2. 11 класс

Содержание
- 2. Фридрих Мишер Открыл дезоксирибонуклеиновую кислоту в 1869 году молодой швейцарский врач Фридрих Мишер, работавший тогда в
- 3. Нуклеиновые кислоты Нуклеиновая кислота (от лат. nucleus — ядро) — высокомолекулярное органическое соединение, биополимер (полинуклеотид), образованный
- 4. Нуклеотиды и нуклеозиды Нуклеотиды (нуклеозидфосфаты) — фосфорные эфиры нуклеозидов. Нуклеозиды — это гликозиламины, содержащие азотистое основание
- 5. Пурин и пуриновые основания Пурин Гуанин Аденин
- 6. Пиримидин и пиримидиновые основания Пиримидин Тимин Цитозин Урацил
- 7. Строение мономера нуклеиновой кислоты
- 8. История изучения структуры ДНК В 1921 году Левин выдвинул гипотезу «тетрануклеотидной структуры ДНК», оказавшуюся впоследствии ошибочной.
- 9. Правила Чаргаффа 1) Количество аденина равно количеству тимина, а гуанина — цитозину: А=Т, Г=Ц. 2) Количество
- 10. Первая рентгенограмма молекулы ДНК Уилкинс Морис Рентгенограмма волокон ДНК кишечной палочки, полученная Уилкинсом В 1950г этот
- 11. Революция имени Уотсона и Крика Структура двойной спирали ДНК была предложена Френсисом Криком и Джеймсом Уотсоном
- 12. Строение ДНК ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Связи между
- 13. Каждая цепь ДНК имеет определенную ориентацию. Один конец несет гидроксильную группу (-ОН), присоединенную к 3'-углероду в
- 14. Репликация ДНК
- 15. Проблема концевой недорепликации ДНК ДНК-полимераза не может начинать цепочку ДНК, для этого существует фермент праймаза, который
- 16. Теория А.М. Оловнкова Больше 60 раз (число Хейфлика) клетки человека поделиться не могут, поскольку концевая недорепликация
- 17. Биологически активные формы ДНК Эта двухцепочечная молекула закручена по винтовой линии. В целом структура молекулы ДНК
- 18. В-семейство ДНК B-форма ДНК — одно из трех основных конформационных состояний двухцепочечной ДНК, в которой 2
- 19. В-ДНК и родственные ей C-ДНК , D-ДНК и T-ДНК принадлежат В-семейству двойных спиралей. С-ДНК имеет структуру,
- 20. А-семейство ДНК Одним из важнейших вкладов Франклин в модель Уотсона-Крика стала её лекция, проведённая в ноябре
- 21. A-ДНК— одна из форм молекулы ДНК. Представляет собой правозакрученную двусхпиральную молекулу ДНК. Эта форма имеет 11
- 22. Z-семейство ДНК Z-ДНК значительно отличается от правозакрученных форм. Z-ДНК — левозакрученная и имеет первичную структуру, повторяющуюся
- 24. Список использованной литературы 1) «Общая биология», Л.В. Высоцкая, С.М. Глаголев, Г.М. Дымшиц и др. 2) «Клетки»,
- 26. Скачать презентацию

Фридрих Мишер
Открыл дезоксирибонуклеиновую кислоту в 1869 году молодой швейцарский врач Фридрих
Фридрих Мишер
Открыл дезоксирибонуклеиновую кислоту в 1869 году молодой швейцарский врач Фридрих

Нуклеиновые кислоты
Нуклеиновая кислота (от лат. nucleus — ядро) — высокомолекулярное органическое
Нуклеиновые кислоты
Нуклеиновая кислота (от лат. nucleus — ядро) — высокомолекулярное органическое

Нуклеотиды и нуклеозиды
Нуклеотиды (нуклеозидфосфаты) — фосфорные эфиры нуклеозидов.
Нуклеозиды — это гликозиламины,
Нуклеотиды и нуклеозиды
Нуклеотиды (нуклеозидфосфаты) — фосфорные эфиры нуклеозидов.
Нуклеозиды — это гликозиламины,
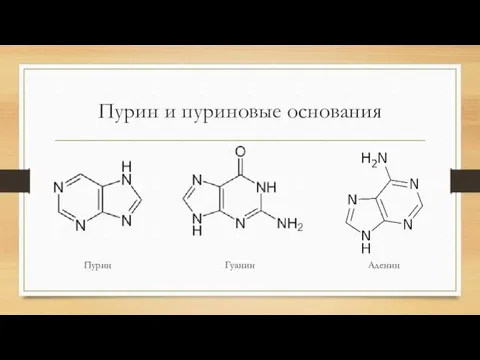
Пурин и пуриновые основания
Пурин
Гуанин
Аденин
Пурин и пуриновые основания
Пурин
Гуанин
Аденин
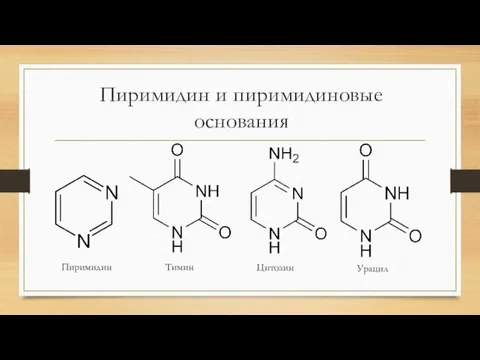
Пиримидин и пиримидиновые
основания
Пиримидин
Тимин
Цитозин
Урацил
Пиримидин и пиримидиновые
основания
Пиримидин
Тимин
Цитозин
Урацил
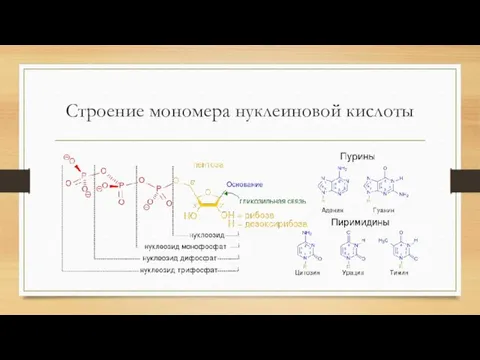
Строение мономера нуклеиновой кислоты
Строение мономера нуклеиновой кислоты

История изучения структуры ДНК
В 1921 году Левин выдвинул гипотезу «тетрануклеотидной структуры
История изучения структуры ДНК
В 1921 году Левин выдвинул гипотезу «тетрануклеотидной структуры

Правила Чаргаффа
1) Количество аденина равно количеству тимина, а гуанина — цитозину:
Правила Чаргаффа
1) Количество аденина равно количеству тимина, а гуанина — цитозину:

Первая рентгенограмма молекулы ДНК
Уилкинс Морис
Рентгенограмма волокон ДНК кишечной палочки, полученная Уилкинсом
В
Первая рентгенограмма молекулы ДНК
Уилкинс Морис
Рентгенограмма волокон ДНК кишечной палочки, полученная Уилкинсом
В

Революция имени Уотсона и Крика
Структура двойной спирали ДНК была предложена Френсисом
Революция имени Уотсона и Крика
Структура двойной спирали ДНК была предложена Френсисом
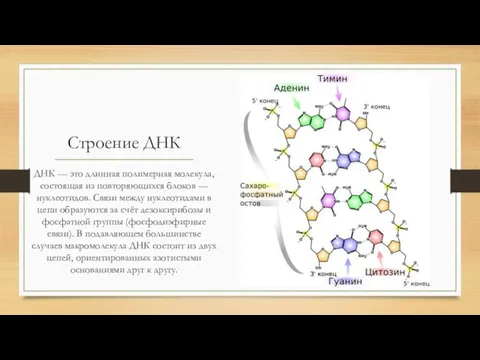
Строение ДНК
ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков
Строение ДНК
ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков

Каждая цепь ДНК имеет определенную ориентацию. Один конец несет гидроксильную группу
Каждая цепь ДНК имеет определенную ориентацию. Один конец несет гидроксильную группу

Репликация ДНК
Репликация ДНК

Проблема концевой недорепликации ДНК
ДНК-полимераза не может начинать цепочку ДНК, для
Проблема концевой недорепликации ДНК
ДНК-полимераза не может начинать цепочку ДНК, для

Теория А.М. Оловнкова
Больше 60 раз (число Хейфлика) клетки человека поделиться
Теория А.М. Оловнкова
Больше 60 раз (число Хейфлика) клетки человека поделиться
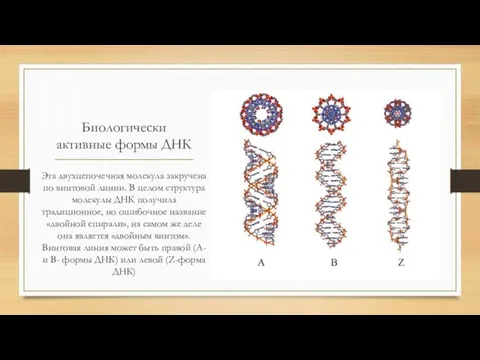
Биологически активные формы ДНК
Эта двухцепочечная молекула закручена по винтовой линии. В
Биологически активные формы ДНК
Эта двухцепочечная молекула закручена по винтовой линии. В

В-семейство ДНК
B-форма ДНК — одно из трех основных конформационных состояний двухцепочечной
В-семейство ДНК
B-форма ДНК — одно из трех основных конформационных состояний двухцепочечной

В-ДНК и родственные ей C-ДНК , D-ДНК и T-ДНК принадлежат В-семейству
В-ДНК и родственные ей C-ДНК , D-ДНК и T-ДНК принадлежат В-семейству

А-семейство ДНК
Одним из важнейших вкладов Франклин в модель Уотсона-Крика стала её
А-семейство ДНК
Одним из важнейших вкладов Франклин в модель Уотсона-Крика стала её

A-ДНК— одна из форм молекулы ДНК. Представляет собой правозакрученную двусхпиральную молекулу
A-ДНК— одна из форм молекулы ДНК. Представляет собой правозакрученную двусхпиральную молекулу

Z-семейство ДНК
Z-ДНК значительно отличается от правозакрученных форм. Z-ДНК — левозакрученная и
Z-семейство ДНК
Z-ДНК значительно отличается от правозакрученных форм. Z-ДНК — левозакрученная и

Список использованной литературы
1) «Общая биология», Л.В. Высоцкая, С.М. Глаголев, Г.М. Дымшиц
Список использованной литературы
1) «Общая биология», Л.В. Высоцкая, С.М. Глаголев, Г.М. Дымшиц
 Проблемы мотивации современных детей
Проблемы мотивации современных детей Биологическое оружие
Биологическое оружие Обработка записанного вокала
Обработка записанного вокала Организация развивающей предметно-пространственной среды ДО в связи с введением ФГОС
Организация развивающей предметно-пространственной среды ДО в связи с введением ФГОС Природный и попутный нефтяной газы
Природный и попутный нефтяной газы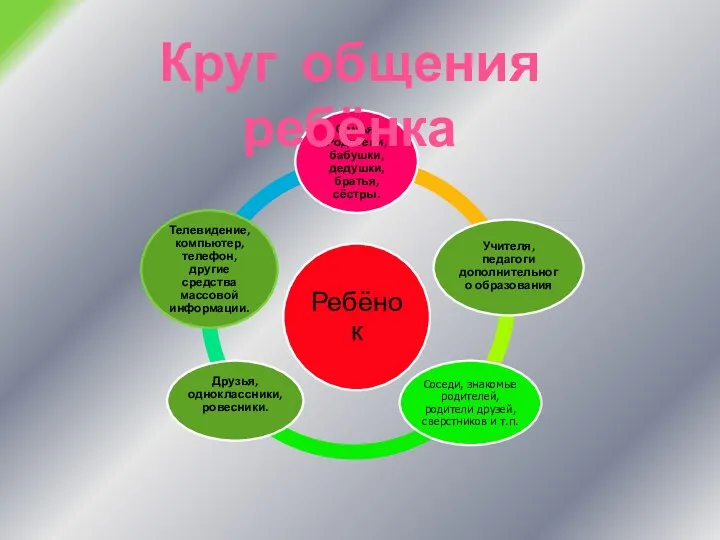 Общение в жизни ребенка
Общение в жизни ребенка Миофункциональная коррекция в работе логопеда (здоровьесберегающие технологии)
Миофункциональная коррекция в работе логопеда (здоровьесберегающие технологии) Псориаз. Красный плоский лишай
Псориаз. Красный плоский лишай Итоги работы путевой машинной станции № 55
Итоги работы путевой машинной станции № 55 Отражение и преломление света. Оптическое изображение
Отражение и преломление света. Оптическое изображение Програмне забезпечення мікропроцесорних систем
Програмне забезпечення мікропроцесорних систем Создание и размещение таможенных органов
Создание и размещение таможенных органов Составление числовых выражений
Составление числовых выражений Подготовка данных к анализу. (Лекция 5)
Подготовка данных к анализу. (Лекция 5) Православные храмы города Саратова
Православные храмы города Саратова Алгоритм диагностики и оказания скорой помощи при кровотечениях
Алгоритм диагностики и оказания скорой помощи при кровотечениях Легенды о короле Артуре: Мифы и Реальность
Легенды о короле Артуре: Мифы и Реальность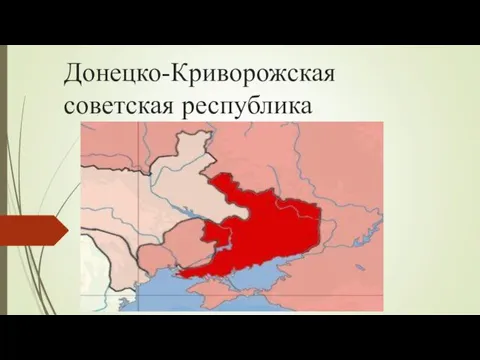 Донецко-Криворожская советская республика
Донецко-Криворожская советская республика Теорема косинусов
Теорема косинусов Комплекс устройств и схема электроснабжения железных дорог
Комплекс устройств и схема электроснабжения железных дорог Религия иудаизм
Религия иудаизм Специальное сезонное предложение. Корпоративное сервисное обслуживание. Группа компаний Авторусь
Специальное сезонное предложение. Корпоративное сервисное обслуживание. Группа компаний Авторусь Проектирование и эксплуатация объектов размещения отходов
Проектирование и эксплуатация объектов размещения отходов План квартиры с привязкой перегородок
План квартиры с привязкой перегородок Предмет, задачи и объекты дактилоскопической экспертизы
Предмет, задачи и объекты дактилоскопической экспертизы Защита древесины
Защита древесины ОГАС 2.0. Программно-техническое решение системы (часть 2)
ОГАС 2.0. Программно-техническое решение системы (часть 2) Клинико-фармакологическая характеристика лекарственных средств, применяемых при бронхиальной астме
Клинико-фармакологическая характеристика лекарственных средств, применяемых при бронхиальной астме