- Основная роль ЦТК

Содержание
- 2. лимонная кислота ЩУК Цикл Кребса НАД НАД НАД НАД Н НАД Н НАД Н ФАД Н2
- 3. Основная роль ЦТК Основная роль ЦТК заключается в 1) генерации атомов водорода для работы дыхательной цепи,
- 4. Регуляция цикла трикарбоновых кислот Главным и основным регулятором ЦТК является оксалоацетат, а точнее его доступность. Наличие
- 5. Регуляторные ферменты ЦТК Некоторые ферменты ЦТК являются чувствительными к аллостерической регуляции метаболитами:
- 6. Окислительное фосфорилирование Молекулы НАДН и ФАДН2, образуемые в реакциях окисления углеводов, жирных кислот, спиртов и аминокислот,
- 7. Механизм окислительного фосфорилирования По современным представлениям внутренняя митохондриальная мембрана содержит ряд мультиферментных комплексов, включающих множество ферментов.
- 8. Принцип работы дыхательной цепи В целом работа дыхательной цепи заключается в следующем: 1) Образующиеся в реакциях
- 9. АТФ-синтаза
- 10. Общие принцип окислительного фосфорилирования
- 11. Дыхательная цепь включает множество белков-переносчиков Всего цепь переноса электронов включает в себя около 40 разнообразных белков,
- 12. Блок-схема дыхательной цепи
- 13. Первый комплекс. 1 комплекс - НАДН-КоQ-оксидоредуктаза Этот комплекс также имеет рабочее название НАДН-дегидрогеназа, содержит ФМН, 22
- 14. Второй комплекс 2 комплекс - ФАД-зависимые дегидрогеназы Данный комплекс как таковой не существует, его выделение условно.
- 15. Третий комплекс 3 комплекс - КоQ-цитохром с-оксидоредуктаза Данный комплекс включает цитохромы b и c1. Кроме цитохромов
- 16. Четвёртый комплекс 4 комплекс. - Цитохром с-кислород-оксидоредуктаза В этом комплексе находятся цитохромы а и а3, он
- 17. Пятый комплекс 5 комплекс – это фермент АТФ-синтаза, состоящий из множества белковых цепей, подразделенных на две
- 18. Строение дыхательной цепи и механизм окислительного фосфорилирования
- 19. Механизм окислительного фосфорилирования На основании строения и функций компонентов дыхательной цепи предложен механизм окислительного фосфорилирования: 1)
- 20. 3) На внутренней мембране митохондрий атомы водорода (от НАДН и ФАДН2) передают свои электроны в дыхательную
- 21. Участки сопряжения 5) Перенос ионов водорода через мембрану (выкачивание) происходит не случайно, а в строго определенных
- 22. Наработка АТФ 6. Как завершение всех предыдущих событий и необходимый их результат происходит наработка АТФ: ионы
- 23. Гипоэнергетические состояния Причиной гипоэнергетических состояний может быть следующее: гиповитаминозы экзогенные и/или эндогенные – снижается скорость и
- 24. Разобщители окисления и фосфорилирования К разобщителям в первую очередь относят "протонофоры" – вещества переносящие ионы водорода.
- 25. Ингибиторы ферментов дыхательной цепи Ряд веществ может ингибировать ферменты дыхательной цепи и блокировать движение электронов от
- 27. Скачать презентацию
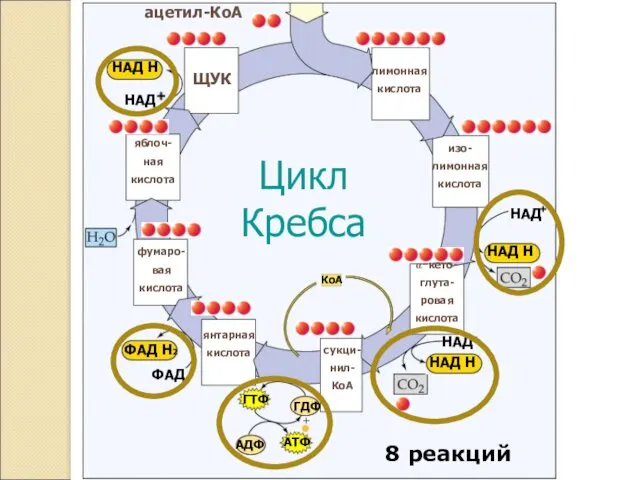
лимонная
кислота
ЩУК
Цикл
Кребса
НАД
НАД
НАД
НАД Н
НАД Н
НАД Н
ФАД Н2
ФАД
ГДФ
ГТФ
АДФ
АТФ
изо-
лимонная
кислота
α−кето-
глута-
ровая
кислота
янтарная
кислота
фумаро-
вая
кислота
яблоч-
ная
кислота
КоА
сукци-
нил-
КоА
ацетил-КоА
8 реакций
лимонная
кислота
ЩУК
Цикл
Кребса
НАД
НАД
НАД
НАД Н
НАД Н
НАД Н
ФАД Н2
ФАД
ГДФ
ГТФ
АДФ
АТФ
изо-
лимонная
кислота
α−кето-
глута-
ровая
кислота
янтарная
кислота
фумаро-
вая
кислота
яблоч-
ная
кислота
КоА
сукци-
нил-
КоА
ацетил-КоА
8 реакций

Основная роль ЦТК
Основная роль ЦТК заключается в
1) генерации атомов водорода для
Основная роль ЦТК
Основная роль ЦТК заключается в
1) генерации атомов водорода для

Регуляция цикла трикарбоновых кислот
Главным и основным регулятором ЦТК является оксалоацетат, а
Регуляция цикла трикарбоновых кислот
Главным и основным регулятором ЦТК является оксалоацетат, а
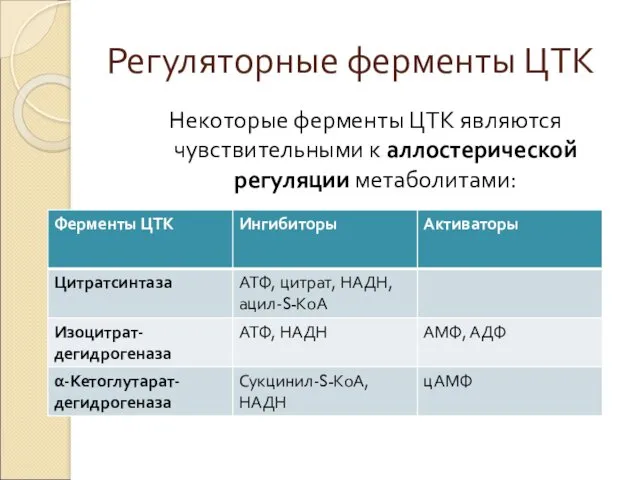
Регуляторные ферменты ЦТК
Некоторые ферменты ЦТК являются чувствительными к аллостерической регуляции метаболитами:
Регуляторные ферменты ЦТК
Некоторые ферменты ЦТК являются чувствительными к аллостерической регуляции метаболитами:

Окислительное фосфорилирование
Молекулы НАДН и ФАДН2, образуемые в реакциях окисления углеводов, жирных
Окислительное фосфорилирование
Молекулы НАДН и ФАДН2, образуемые в реакциях окисления углеводов, жирных

Механизм окислительного фосфорилирования
По современным представлениям внутренняя митохондриальная мембрана содержит ряд мультиферментных
Механизм окислительного фосфорилирования
По современным представлениям внутренняя митохондриальная мембрана содержит ряд мультиферментных

Принцип работы дыхательной цепи
В целом работа дыхательной цепи заключается в следующем:
1)
Принцип работы дыхательной цепи
В целом работа дыхательной цепи заключается в следующем:
1)
АТФ-синтаза
АТФ-синтаза
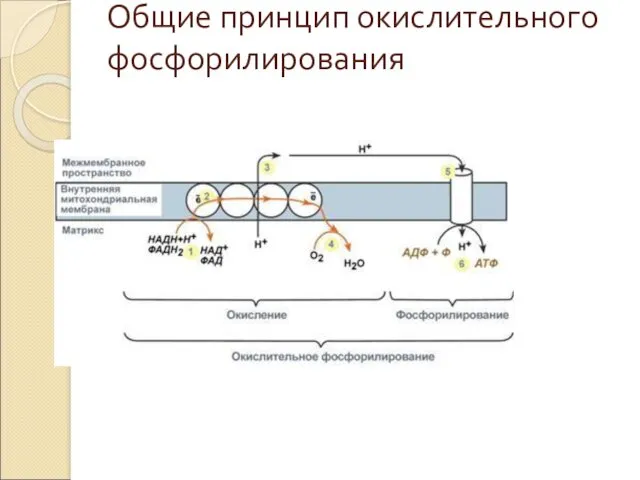
Общие принцип окислительного фосфорилирования
Общие принцип окислительного фосфорилирования

Дыхательная цепь включает множество белков-переносчиков
Всего цепь переноса электронов включает в
Дыхательная цепь включает множество белков-переносчиков
Всего цепь переноса электронов включает в
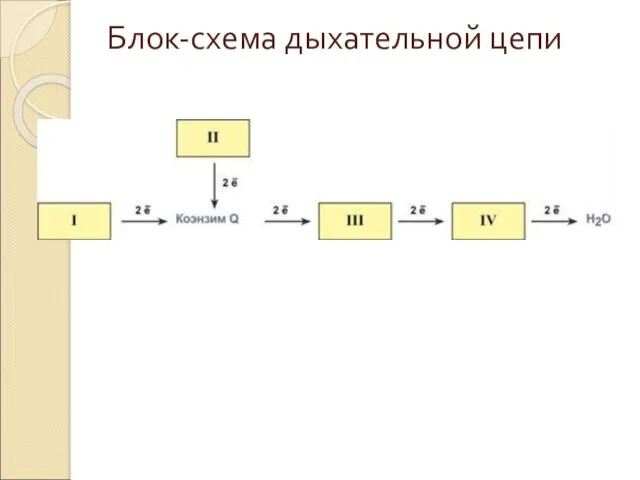
Блок-схема дыхательной цепи
Блок-схема дыхательной цепи

Первый комплекс.
1 комплекс - НАДН-КоQ-оксидоредуктаза
Этот комплекс также имеет рабочее название НАДН-дегидрогеназа, содержит ФМН,
Первый комплекс.
1 комплекс - НАДН-КоQ-оксидоредуктаза
Этот комплекс также имеет рабочее название НАДН-дегидрогеназа, содержит ФМН,

Второй комплекс
2 комплекс - ФАД-зависимые дегидрогеназы
Данный комплекс как таковой не существует,
Второй комплекс
2 комплекс - ФАД-зависимые дегидрогеназы
Данный комплекс как таковой не существует,

Третий комплекс
3 комплекс - КоQ-цитохром с-оксидоредуктаза
Данный комплекс включает цитохромы b и
Третий комплекс
3 комплекс - КоQ-цитохром с-оксидоредуктаза
Данный комплекс включает цитохромы b и

Четвёртый комплекс
4 комплекс. - Цитохром с-кислород-оксидоредуктаза
В этом комплексе находятся цитохромы а
Четвёртый комплекс
4 комплекс. - Цитохром с-кислород-оксидоредуктаза
В этом комплексе находятся цитохромы а

Пятый комплекс
5 комплекс – это фермент АТФ-синтаза, состоящий из множества белковых
Пятый комплекс
5 комплекс – это фермент АТФ-синтаза, состоящий из множества белковых
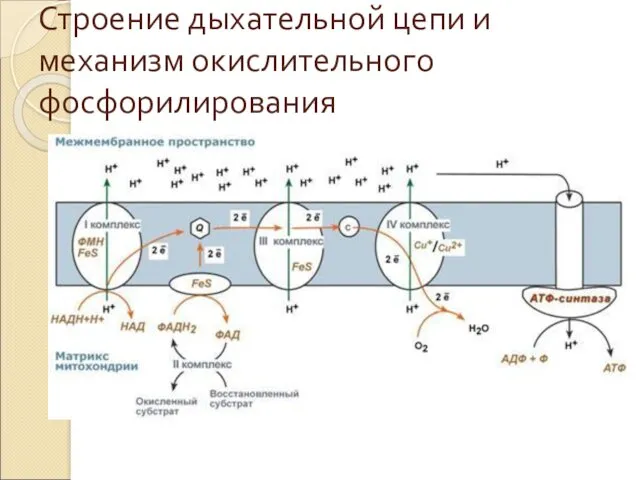
Строение дыхательной цепи и механизм окислительного фосфорилирования
Строение дыхательной цепи и механизм окислительного фосфорилирования

Механизм окислительного фосфорилирования
На основании строения и функций компонентов дыхательной цепи предложен
Механизм окислительного фосфорилирования
На основании строения и функций компонентов дыхательной цепи предложен

3) На внутренней мембране митохондрий атомы водорода (от НАДН и ФАДН2)
3) На внутренней мембране митохондрий атомы водорода (от НАДН и ФАДН2)

Участки сопряжения
5) Перенос ионов водорода через мембрану (выкачивание) происходит не случайно,
Участки сопряжения
5) Перенос ионов водорода через мембрану (выкачивание) происходит не случайно,

Наработка АТФ
6. Как завершение всех предыдущих событий и необходимый их результат
Наработка АТФ
6. Как завершение всех предыдущих событий и необходимый их результат

Гипоэнергетические состояния
Причиной гипоэнергетических состояний может быть следующее:
гиповитаминозы экзогенные и/или эндогенные – снижается
Гипоэнергетические состояния
Причиной гипоэнергетических состояний может быть следующее:
гиповитаминозы экзогенные и/или эндогенные – снижается

Разобщители окисления и фосфорилирования
К разобщителям в первую очередь относят "протонофоры" –
Разобщители окисления и фосфорилирования
К разобщителям в первую очередь относят "протонофоры" –

Ингибиторы ферментов дыхательной цепи
Ряд веществ может ингибировать ферменты дыхательной цепи и
Ингибиторы ферментов дыхательной цепи
Ряд веществ может ингибировать ферменты дыхательной цепи и
 Тур по Горному Алтаю Большая прогулка
Тур по Горному Алтаю Большая прогулка Основы моделирования вычислительных систем
Основы моделирования вычислительных систем Who is the pride of your country
Who is the pride of your country Алмаз - Жемчужина кен орны
Алмаз - Жемчужина кен орны Информационные технологии в профессии. Сфера IT
Информационные технологии в профессии. Сфера IT Теплообменное оборудование тепловых пунктов. Кожухотрубные теплообменники
Теплообменное оборудование тепловых пунктов. Кожухотрубные теплообменники Методический семинар Методика проведения нестандартного урока
Методический семинар Методика проведения нестандартного урока Электронное портфолио - папка (файлы) с документами, фото, диагностическими,воспитательными и методическими материалами.
Электронное портфолио - папка (файлы) с документами, фото, диагностическими,воспитательными и методическими материалами. Непозиционные системы счисления
Непозиционные системы счисления Малі тіла Сонячної системи
Малі тіла Сонячної системи Проект эффективной разработки месторождения Снежное
Проект эффективной разработки месторождения Снежное Автоматизация хранения сельскохозяйственной продукции
Автоматизация хранения сельскохозяйственной продукции Отчет по нравственно - патриотическому воспитанию за учебный год в группе старшего дошкольного возраста с 5 до 6 лет
Отчет по нравственно - патриотическому воспитанию за учебный год в группе старшего дошкольного возраста с 5 до 6 лет Innovation electric car: Tesla
Innovation electric car: Tesla Объем и скорость передачи цифрового потока ТВ-информации. Резервы сокращения цифрового потока ТВ-систем. (Лекция 5)
Объем и скорость передачи цифрового потока ТВ-информации. Резервы сокращения цифрового потока ТВ-систем. (Лекция 5) Презентация к беседе Воздух, которым мы дышим
Презентация к беседе Воздух, которым мы дышим Землетрясения. Презентация к урокам географии и ОБЖ.
Землетрясения. Презентация к урокам географии и ОБЖ. Буровые технологические жидкости. Скважины. (Лекция 1)
Буровые технологические жидкости. Скважины. (Лекция 1) Интеллектуальная игра Самый умный
Интеллектуальная игра Самый умный Современные постановки пьесы Чайка А.П. Чехова на сцене русских театров
Современные постановки пьесы Чайка А.П. Чехова на сцене русских театров Эбру специализация
Эбру специализация Рынок услуг интернет-маркетинга
Рынок услуг интернет-маркетинга Модернизация поездной радиосвязи с применением цифровой радиосвязи стандарта DMR
Модернизация поездной радиосвязи с применением цифровой радиосвязи стандарта DMR Нагревание воздуха и его температура
Нагревание воздуха и его температура Проблемы российского образования в современных условиях
Проблемы российского образования в современных условиях Движение воды в океане
Движение воды в океане Финансовая система РФ, ее основные звенья, их взаимосвязь
Финансовая система РФ, ее основные звенья, их взаимосвязь Загадки о Великой Отечественной войне
Загадки о Великой Отечественной войне