- Пероксисомы/Микротельца

Содержание
- 2. Пероксисомы или микротельца – органеллы, окруженные однослойной мембраной. Пероксисомы присутствуют у большинства эукариотических организмов, имеют, как
- 3. Пероксисомы содержат ферменты, ответственные за множество биохимических путей, в частности, β-окисление жирных кислот и метаболизм перекиси
- 4. Кроме окислительных ферментов, пероксисома обязательно содержит каталазу. Каталаза разлагает образовавшуюся перекись водорода, поскольку она достаточно токсична.
- 5. Последние данные продемонстрировали, что пероксисомы почкующихся дрожжей, мицелиального гриба Рodospora anserina и цветкового растения Arаbidopsis thaliána
- 6. Функции пероксисом у эукариот Пероксисомы участвуют в регуляции разнообразных процессов развития, таких как узнавание женских/мужских гаметофитов
- 7. Общие функции Общие функции включают пероксисомальный путь β-окисления жирных кислот. Этот путь вместе с митохондриальным путем
- 8. Специфические функции Последняя стадия биосинтеза пенициллина у грибов (проводимая ацилтрансферазой, IAT) происходит в пероксисомах. Метилотрофность дрожжей.
- 9. Распространение Пероксисомы присутствуют у большинства групп грибов. Исключение составляет группа анаэробных грибов, такие как Microsporidia, у
- 10. Пероксисомы формируются при росте клеток дрожжей (Candida, Hansenula, Pichia) на метаноле (метилотрофные дрожжи) и содержат три
- 11. Пероксисомы Pichia pastoris Pichia pastoris Рост на метаноле дикий штамм
- 12. Пероксисомы Hansenula polymorpha
- 13. Биогенез Новые пероксисомы всегда возникают из предсуществующих и формируются путем роста и деления органеллы, т. е.
- 14. Молекулярные основы биогенеза пероксисом достаточно хорошо изучены. Формирование пероксисом у разных эукариотических организмов характеризуется общей основой
- 15. В настоящее время общепринято, что практически все пероксисомальные матриксные белки (ПМБ) синтезируются на свободных полисомах в
- 16. Для мембранных белков ситуация более сложная; в зависимости от белка и изучаемого организма, биосинтез этого класса
- 17. В биогенезе и включении в мембрану пероксисомальных мембранных белков аскомицета Podospora anserina участвуют три пероксисомальных матриксных
- 18. В импорте пероксисомальных матриксных белков используется большой набор пероксинов: PEX1, PEX2, PEX4-PEX10, PEX12-PEX15, PEX17-PEX18, PEX20-PEX22, Pex26,
- 19. Биогенез пероксисом
- 20. Пероксиновые гены имеют эукариотическое происхождение. Имеется множество высоко консервативных пероксинов, которые, по-видимому, представляют собой основу биогенеза
- 21. белок наследования пероксисом 2 = Inheritance of peroxisomes protein 2 (Inp2), рецептор Myo2 белок наследования пероксисом
- 22. Пексофагия Для регуляции функций пероксисом и ограничения процесса клеточного старения, лишние и нефункциональные пероксисомы должны быть
- 23. Макро- и микропексофагия (аналогичные макро- и микроавтофагии) два морфологически четких типа пероксисомспецифичных вакуолярных/лизосомальных путей деградации, которые
- 24. Пексофагия у Pichia pastoris Мембрана вакуоли пексофагосома макропексофагия микропексофагия
- 25. Макропексофагия у Hansenula polymorpha
- 26. Индуцированная глюкозой микропексофагия у Pichia pastoris
- 27. Транспорт и наследование пероксисом у дрожжей
- 28. Пероксисомы занимают строго фиксированное положение в процессе роста почки, что связано с существованием заякоривающего белка/ов. У
- 29. Пока большинство пероксисом остаются неподвижными у кортекса материнской клетки во время роста почки, некоторые пероксисомы отрываются
- 30. Деление и перенос пероксисом в дочернюю клетку
- 31. Тельца Воронина Тельца Воронина названы в честь русского миколога Михаил Степановича Воронина (Buller, 1933), который впервые
- 32. Функция телец Воронина – встраивание в поровый канал септы для изоляции неповрежденной клетки мицелия Недавно было
- 33. Филогенетическое древо грибов дополненное некоторыми характеристиками (рабочая модель), построенное на 18S rRNA (мод. по Berbee and
- 34. Тельца Воронина были впервые изолированы из N. crassa при использовании центрифугирования в градиенте плотности и показано
- 35. Тельца Воронина в апексе гифы Aspergillus nidulans. Окрашивание с использованием антител к матриксному белку микротелец Neurospora
- 36. Микротельца Воронина в районе септы у гифы Aspergillus nidulans (размер от 100 нм до 1 мкм).
- 37. Тельца Воронина у N. crassa Масштабный отрезок 500 нм.
- 38. Структура кристаллической решетки HEX-1 Очищенный рекомбинантный HEX-1 N. crassa спонтанно самособирается в гексагональные кристаллы
- 39. Биогенез телец Воронина Тельца Воронина обнаружены во всех вегетативных гифах и формируются de novo без деления
- 40. Распределение телец Воронина в пяти образцах мицелия при прорастании конидий Aspergillus nidulans до образования первой септы
- 41. Дифференциация телец Воронина из пероксисом
- 43. Скачать презентацию

Пероксисомы или микротельца – органеллы, окруженные однослойной мембраной. Пероксисомы присутствуют у
Пероксисомы или микротельца – органеллы, окруженные однослойной мембраной. Пероксисомы присутствуют у

Пероксисомы содержат ферменты, ответственные за множество биохимических путей, в частности, β-окисление
Пероксисомы содержат ферменты, ответственные за множество биохимических путей, в частности, β-окисление

Кроме окислительных ферментов, пероксисома обязательно содержит каталазу. Каталаза разлагает образовавшуюся перекись
Кроме окислительных ферментов, пероксисома обязательно содержит каталазу. Каталаза разлагает образовавшуюся перекись

Последние данные продемонстрировали, что пероксисомы почкующихся дрожжей, мицелиального гриба Рodospora anserina
Последние данные продемонстрировали, что пероксисомы почкующихся дрожжей, мицелиального гриба Рodospora anserina

Функции пероксисом у эукариот
Пероксисомы участвуют в регуляции разнообразных процессов развития, таких
Функции пероксисом у эукариот
Пероксисомы участвуют в регуляции разнообразных процессов развития, таких

Общие функции
Общие функции включают пероксисомальный путь β-окисления жирных кислот. Этот путь
Общие функции
Общие функции включают пероксисомальный путь β-окисления жирных кислот. Этот путь

Специфические функции
Последняя стадия биосинтеза пенициллина у грибов (проводимая ацилтрансферазой, IAT)
Специфические функции
Последняя стадия биосинтеза пенициллина у грибов (проводимая ацилтрансферазой, IAT)

Распространение
Пероксисомы присутствуют у большинства групп грибов. Исключение составляет группа анаэробных грибов,
Распространение
Пероксисомы присутствуют у большинства групп грибов. Исключение составляет группа анаэробных грибов,

Пероксисомы формируются при росте клеток дрожжей (Candida, Hansenula, Pichia) на метаноле
Пероксисомы формируются при росте клеток дрожжей (Candida, Hansenula, Pichia) на метаноле
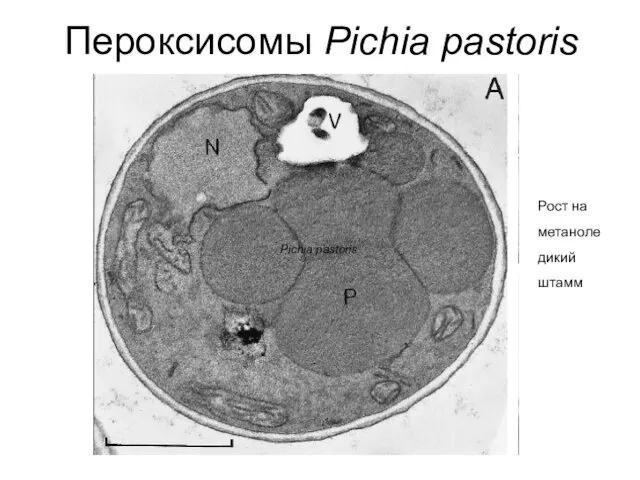
Пероксисомы Pichia pastoris
Pichia pastoris
Рост на метаноле дикий штамм
Пероксисомы Pichia pastoris
Pichia pastoris
Рост на метаноле дикий штамм
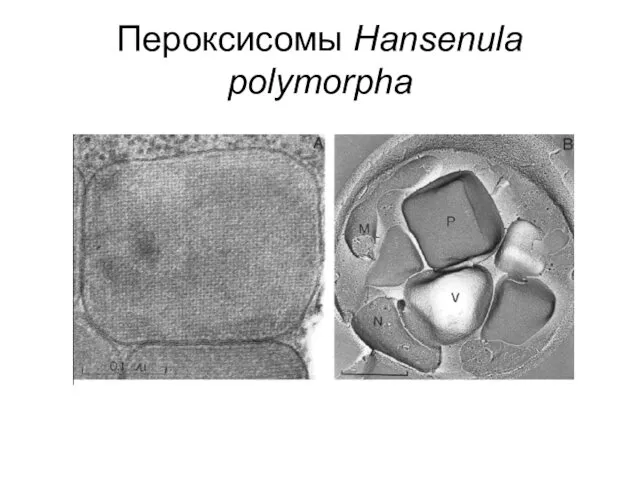
Пероксисомы Hansenula polymorpha
Пероксисомы Hansenula polymorpha

Биогенез
Новые пероксисомы всегда возникают из предсуществующих и формируются путем роста и
Биогенез
Новые пероксисомы всегда возникают из предсуществующих и формируются путем роста и

Молекулярные основы биогенеза пероксисом достаточно хорошо изучены. Формирование пероксисом у разных
Молекулярные основы биогенеза пероксисом достаточно хорошо изучены. Формирование пероксисом у разных

В настоящее время общепринято, что практически все пероксисомальные матриксные белки (ПМБ)
В настоящее время общепринято, что практически все пероксисомальные матриксные белки (ПМБ)

Для мембранных белков ситуация более сложная; в зависимости от белка и
Для мембранных белков ситуация более сложная; в зависимости от белка и

В биогенезе и включении в мембрану пероксисомальных мембранных белков аскомицета Podospora
В биогенезе и включении в мембрану пероксисомальных мембранных белков аскомицета Podospora

В импорте пероксисомальных матриксных белков используется большой набор пероксинов: PEX1, PEX2,
В импорте пероксисомальных матриксных белков используется большой набор пероксинов: PEX1, PEX2,
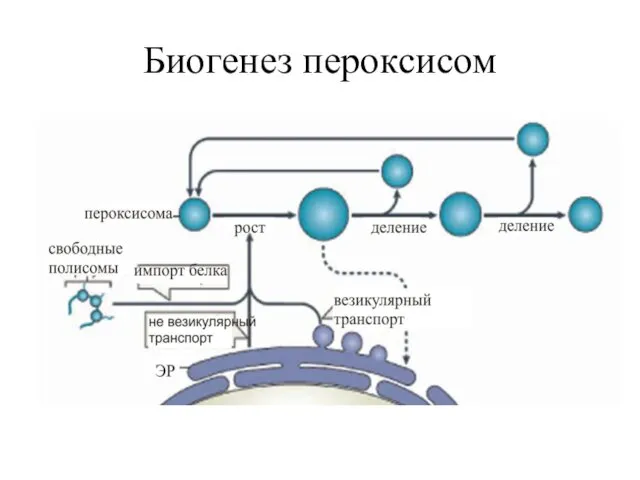
Биогенез пероксисом
Биогенез пероксисом

Пероксиновые гены имеют эукариотическое происхождение. Имеется множество высоко консервативных пероксинов, которые,
Пероксиновые гены имеют эукариотическое происхождение. Имеется множество высоко консервативных пероксинов, которые,
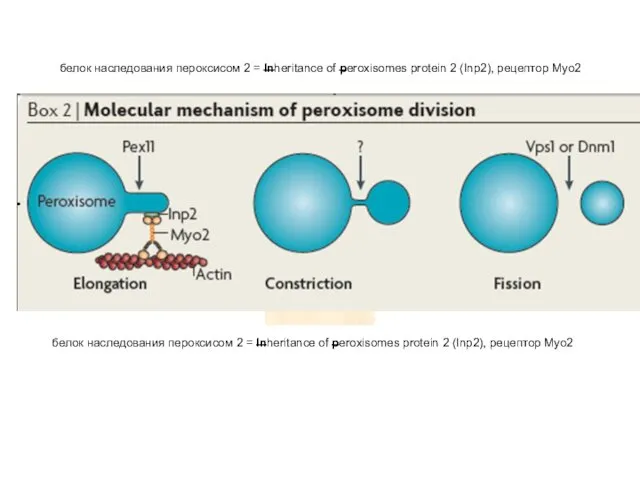
белок наследования пероксисом 2 = Inheritance of peroxisomes protein 2 (Inp2),
белок наследования пероксисом 2 = Inheritance of peroxisomes protein 2 (Inp2),

Пексофагия
Для регуляции функций пероксисом и ограничения процесса клеточного старения, лишние и
Пексофагия
Для регуляции функций пероксисом и ограничения процесса клеточного старения, лишние и

Макро- и микропексофагия (аналогичные макро- и микроавтофагии) два морфологически четких типа
Макро- и микропексофагия (аналогичные макро- и микроавтофагии) два морфологически четких типа
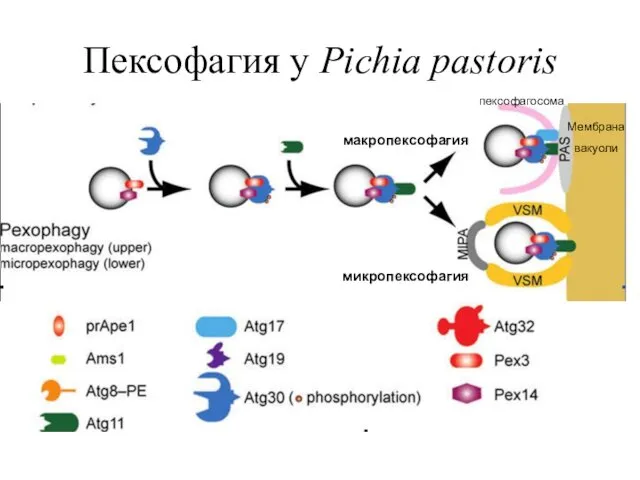
Пексофагия у Pichia pastoris
Мембрана вакуоли
пексофагосома
макропексофагия
микропексофагия
Пексофагия у Pichia pastoris
Мембрана вакуоли
пексофагосома
макропексофагия
микропексофагия
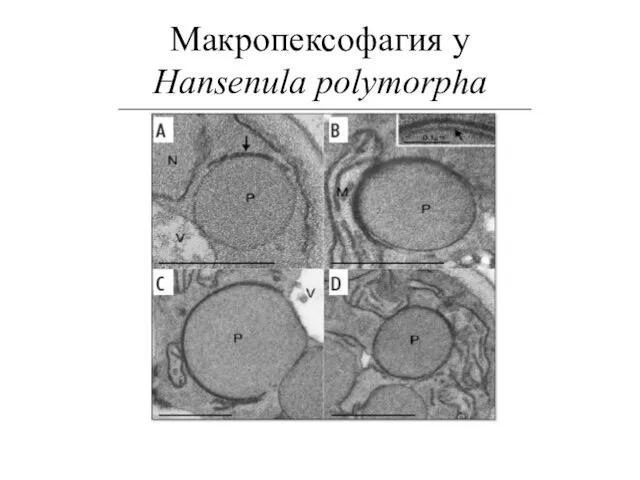
Макропексофагия у
Hansenula polymorpha
Макропексофагия у
Hansenula polymorpha
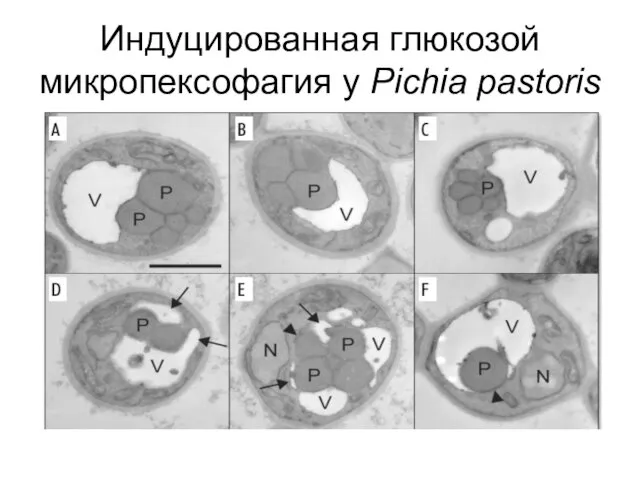
Индуцированная глюкозой микропексофагия у Pichia pastoris
Индуцированная глюкозой микропексофагия у Pichia pastoris
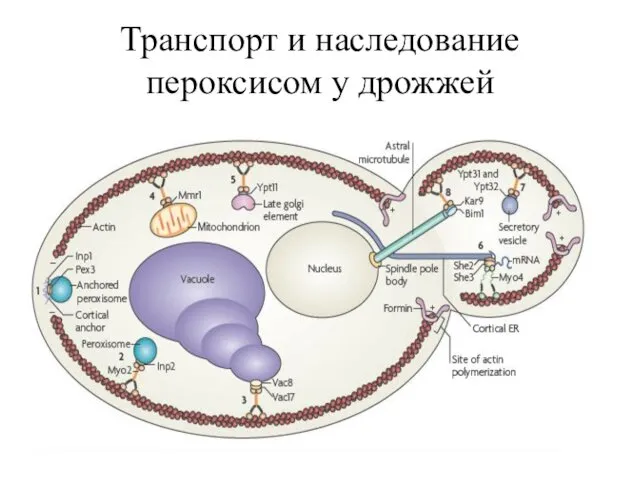
Транспорт и наследование пероксисом у дрожжей
Транспорт и наследование пероксисом у дрожжей

Пероксисомы занимают строго фиксированное положение в процессе роста почки, что связано
Пероксисомы занимают строго фиксированное положение в процессе роста почки, что связано

Пока большинство пероксисом остаются неподвижными у кортекса материнской клетки во время
Пока большинство пероксисом остаются неподвижными у кортекса материнской клетки во время
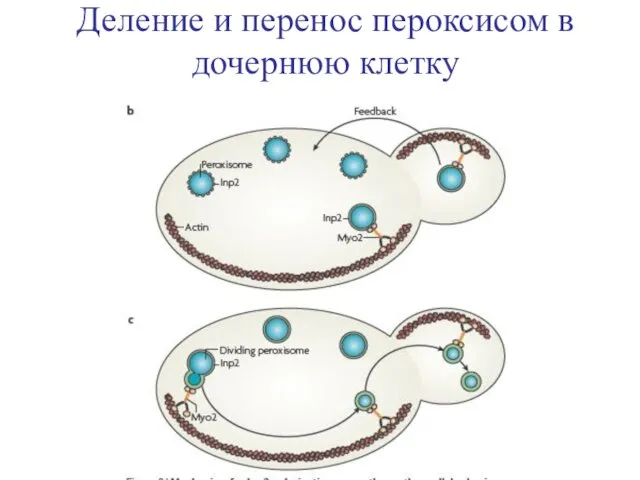
Деление и перенос пероксисом в дочернюю клетку
Деление и перенос пероксисом в дочернюю клетку

Тельца Воронина
Тельца Воронина названы в честь русского миколога Михаил Степановича Воронина
Тельца Воронина
Тельца Воронина названы в честь русского миколога Михаил Степановича Воронина
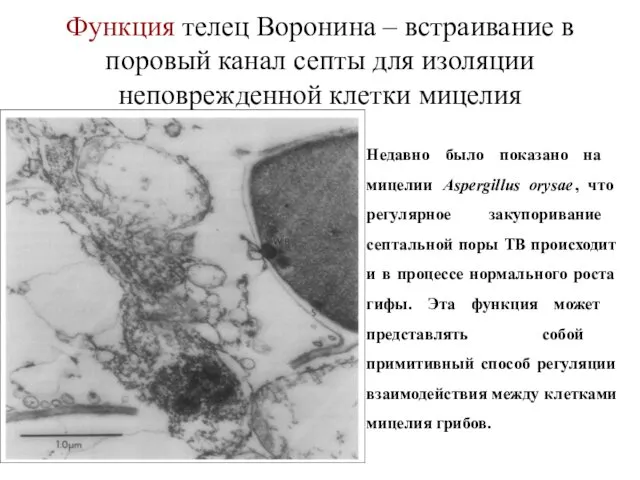
Функция телец Воронина – встраивание в поровый канал септы для изоляции
Функция телец Воронина – встраивание в поровый канал септы для изоляции
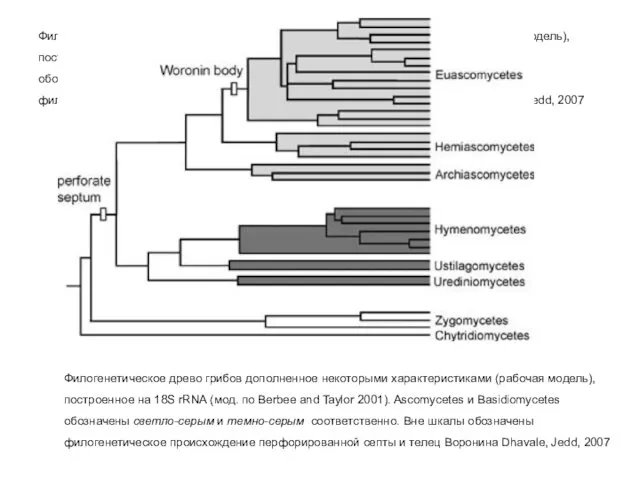
Филогенетическое древо грибов дополненное некоторыми характеристиками (рабочая модель), построенное на 18S
Филогенетическое древо грибов дополненное некоторыми характеристиками (рабочая модель), построенное на 18S

Тельца Воронина были впервые изолированы из N. crassa при использовании центрифугирования
Тельца Воронина были впервые изолированы из N. crassa при использовании центрифугирования
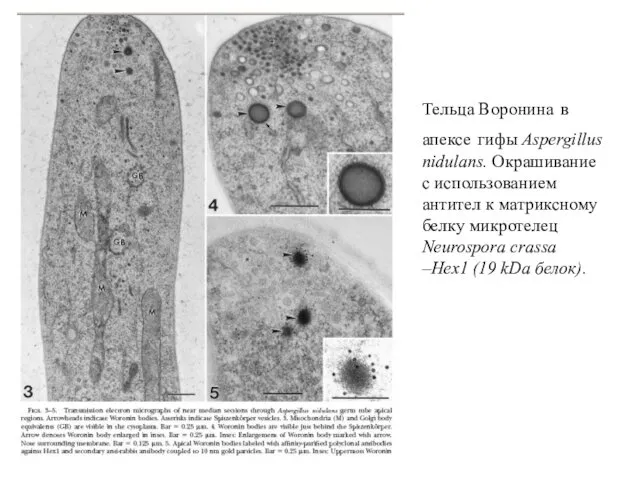
Тельца Воронина в апексе гифы Aspergillus nidulans. Окрашивание с использованием антител
Тельца Воронина в апексе гифы Aspergillus nidulans. Окрашивание с использованием антител
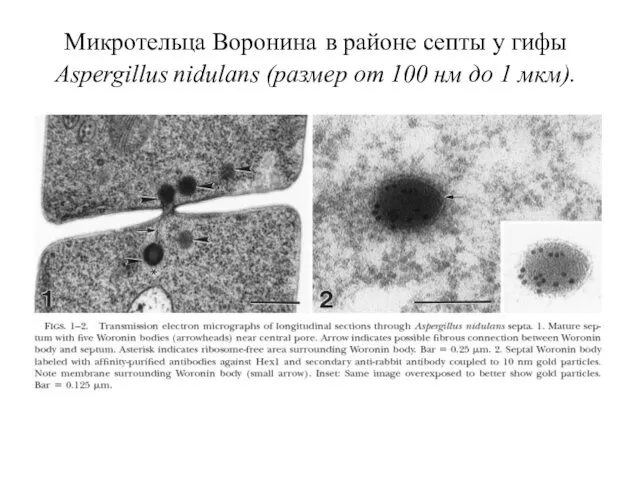
Микротельца Воронина в районе септы у гифы Aspergillus nidulans (размер от
Микротельца Воронина в районе септы у гифы Aspergillus nidulans (размер от
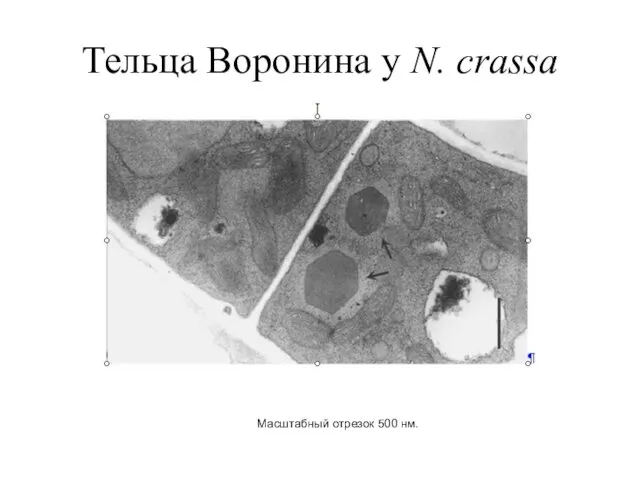
Тельца Воронина у N. crassa
Масштабный отрезок 500 нм.
Тельца Воронина у N. crassa
Масштабный отрезок 500 нм.

Структура кристаллической решетки HEX-1
Очищенный рекомбинантный HEX-1 N. crassa спонтанно самособирается
Структура кристаллической решетки HEX-1
Очищенный рекомбинантный HEX-1 N. crassa спонтанно самособирается

Биогенез телец Воронина
Тельца Воронина обнаружены во всех вегетативных гифах и
Биогенез телец Воронина
Тельца Воронина обнаружены во всех вегетативных гифах и
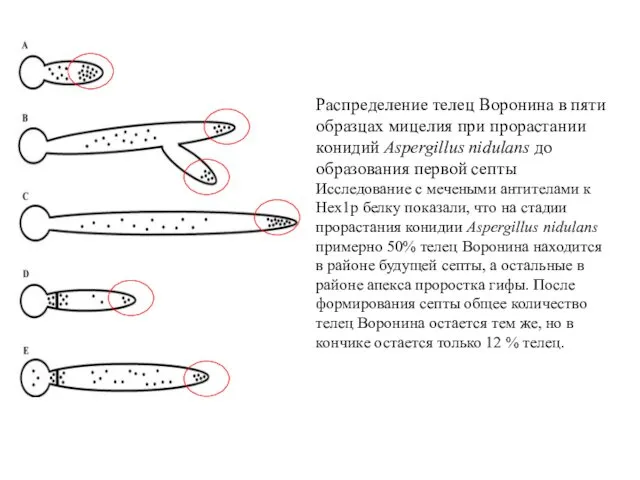
Распределение телец Воронина в пяти образцах мицелия при прорастании конидий Aspergillus
Распределение телец Воронина в пяти образцах мицелия при прорастании конидий Aspergillus
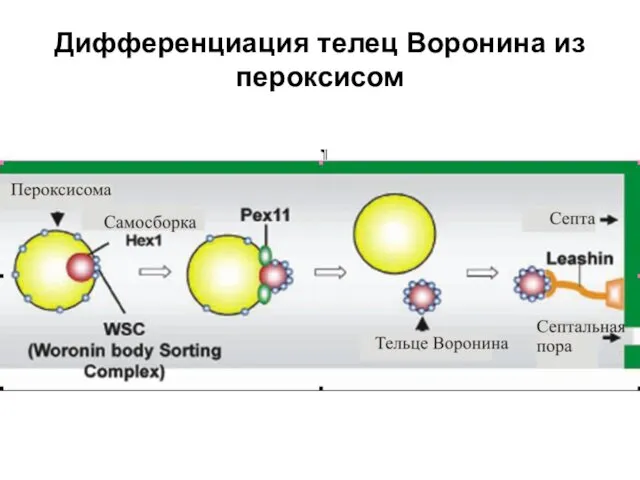
Дифференциация телец Воронина из пероксисом
Дифференциация телец Воронина из пероксисом
 Знаменитые люди Великобритании
Знаменитые люди Великобритании Возникновение ислама. Арабский халифат и его распад
Возникновение ислама. Арабский халифат и его распад Падение редуцированных и последствия этого процесса в истории языка
Падение редуцированных и последствия этого процесса в истории языка Презентация к исследовательской работе Жевательная резинка - друг или враг
Презентация к исследовательской работе Жевательная резинка - друг или враг Роль школьной библиотеки в формированииустойчивого познавательного интереса школьников.
Роль школьной библиотеки в формированииустойчивого познавательного интереса школьников. Тамақ және жеңіл өнеркәсіптерінің салалары
Тамақ және жеңіл өнеркәсіптерінің салалары Краткосрочный проект для детей пятого года жизни
Краткосрочный проект для детей пятого года жизни Хто хоче стати міліонером на тему: Перша світова війна
Хто хоче стати міліонером на тему: Перша світова війна Баскетбол
Баскетбол Открытка к 9 Мая. Мастер и мастерица
Открытка к 9 Мая. Мастер и мастерица Сложные эфиры
Сложные эфиры Алкадиены.
Алкадиены. Болезнь Альцгеймера
Болезнь Альцгеймера Развивающая предметно-пространственная среда по физическому развитию детей дошкольного возраста
Развивающая предметно-пространственная среда по физическому развитию детей дошкольного возраста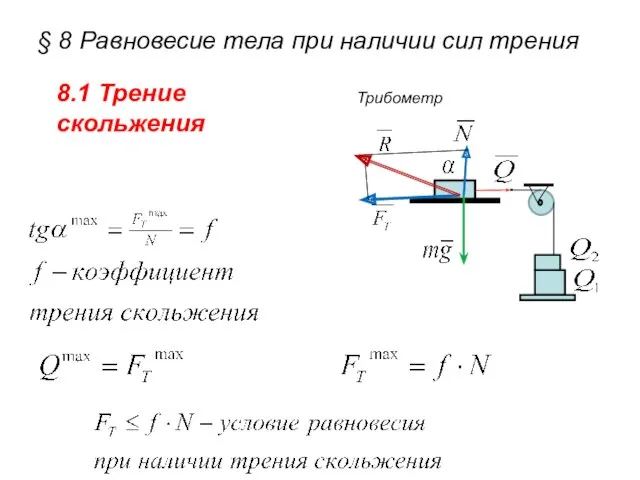 §8. Равновесие тела при наличии сил трения
§8. Равновесие тела при наличии сил трения Энергия ветра
Энергия ветра Бог нас любит
Бог нас любит Свободные экономические зоны и их роль в мировой экономике
Свободные экономические зоны и их роль в мировой экономике Презентация по обучению детей грамоте
Презентация по обучению детей грамоте как определить падеж сущ-х
как определить падеж сущ-х Психология развития: концепция Э. Эриксона психостадиального развития личности
Психология развития: концепция Э. Эриксона психостадиального развития личности Алтайская ГЭС. Фундамент устойчивого развития Республики Алтай
Алтайская ГЭС. Фундамент устойчивого развития Республики Алтай Презентация к уроку по теме Страны Европы
Презентация к уроку по теме Страны Европы Особо охраняемые природные территории. Тамбовская область
Особо охраняемые природные территории. Тамбовская область Раздел Портфолио 2
Раздел Портфолио 2 Религия, её виды, функции, структура
Религия, её виды, функции, структура Работа по проекту Классическая музыка в детском саду С ДМШ имени Йозефа Гайдна.
Работа по проекту Классическая музыка в детском саду С ДМШ имени Йозефа Гайдна. Проект в средней группе на тему: День рожденье куклы Даши
Проект в средней группе на тему: День рожденье куклы Даши