- Пространственная структура экосистем. Вертикальная структура экосистем

Содержание
- 3. Разновысотность растений и «разноглубинность» - важные оси дифференциации экологических ниш в биогеоценозе. В сомкнутом фитоценозе на
- 5. Два основных варианта разновысотности: Ярусность, когда на глаз видно расчленение биоценоза по вертикали на четко отграниченные
- 6. Пример экосистемы с четкой ярусной дифференциацией
- 7. Пример экосистемы с нечетко выраженной ярусностью
- 9. Горизонтальная структура экосистемы. Мозаичность – это явление неоднородности горизонтального сложения биоценоза. Горизонтально неоднородные структуры внутри биоценоза
- 12. Соотношение дискретности и непрерывности в экосистемах – один из интереснейших и важнейших вопросов современной экологии. Косвенным
- 13. Представления о дискретности экосистем (организмистские аналогии) связаны с работами американского эколога Ф. Клементса начала ХХ в.
- 14. Новые представления о непрерывности растительного покрова – теория континуума Леонтий Григорьевич Раменский (1884-1953) Генри Глизон Henry
- 15. Идеи континуума "пробили себе дорогу" лишь в 50-60-х годах ХХ в., когда были выполнены оригинальные исследования
- 16. Зависимость степени непрерывности растительного покрова от числа доминантов и видового разнообразия непрерывность (тропический лес) дискретность (бореальный
- 17. Коцепция единства дискретности и непрерывности в экологии Экосистемы представляют собой явление природы единое в проявлении дискретности
- 18. МЕТОДЫ (ЗАКОНЫ) ОРДИНАЦИИ Методы анализа и описания закономерностей распределения видов или сообществ вдоль некоторых осей (гипотеза
- 19. По методам различают ординации: прямую (ординация ведется по реальным факторам среды – экологическим, пространственным, временным), непрямую
- 20. Прямой градиентный анализ – один из наиболее эффективных методов ординации, который выполняется при возможности прямого измерения
- 21. Факторный анализ – раздел статистического многомерного анализа, объединяющий методы оценки размерности множества наблюдаемых переменных путем исследования
- 22. Пример результатов непрямой ординации лесных экосистем
- 23. Биоразнообразие экосистем Видовое богатство – общее количество видов живых существ в сообществе. Чем больше видов –
- 24. Научный (академический) интерес к проблеме биоразнообразия связан с возможностью познания механизмов формирования структуры сообществ и экосистем
- 25. ГИПОТЕЗЫ АЛЬФА-, БЕТА- И ГАММА-РАЗНООБРАЗИЯ Роберт Уиттекер (R. Whittaker) в 1960 г. предложил различать следующие типы
- 26. бета-разнообразие (разнообразие между сообществами, показатель степени дифференцированности распределения видов или скорости изменения видового состава, видовой структуры
- 27. БИОЦЕНОТИЧЕСКИЕ ПРИНЦИПЫ Тинемана Сформулированные немец- ким гидробиологом Августом Тинеманом в 1939 г. законы экологического разнообразия, согласно
- 28. чем больше отклоняются от нормы (оптимума) условия существования в пределах биотопа, тем беднее видами становится биоценоз
- 29. В качестве примеров можно назвать: процесс «цветения» водохранилищ равнинного типа (массовое развитие сине-зеленых водорослей в условиях
- 30. Индексы биоразнообразия индекс (показатель) Симпсона (Еdward Hugh Simpson; публикация 1951 г.; или Симпсона – Джини) S
- 31. МОДЕЛИ РАСПРЕДЕЛЕНИЯ ЗНАЧИМОСТИ ВИДОВ Формализация кривых относительных оценок значимости видов в сообществе: Роберт Мак-Артур Robert H.
- 32. ПОСТУЛАТЫ ВИДОВОГО ОБЕДНЕНИЯ Основные закономерности, которые автоматиче-ски осуществляются в ходе нарушения эколо-гического разнообразия в сообществе и
- 33. вновь внедрившийся вид приводит к перераспределению пространства экологических ниш сообщества, сужает возможности менее конкурентоспособных видов и
- 34. с антропоцентристской точки зрения замена видов или трофических цепей (сетей) может быть в хозяйственном плане как
- 35. ПРАВИЛО КРОГЕРУСА О ДОМИНИРОВАНИИ ВИДОВ В ЭКСТРЕМАЛЬНЫХ УСЛОВИЯХ В биотопах с экстремальными условиями, как правило, доминируют
- 36. ПРАВИЛО ДЕ КАНДОЛЯ – УОЛЛЕСА (ГЕОГРАФИЧЕСКАЯ ОБУСЛОВЛЕННОСТЬ ИЗМЕНЕНИЯ РАЗНООБРАЗИЯ) По мере продвижения с севера на юг,
- 37. ПРАВИЛО МИНИМУМА ВИДОВ Ремане Парадокс солоноватых вод, согласно которому минимум разнообразия морских и пресноводных видов животных
- 38. ПРАВИЛО ДАРЛИНГТОНА (СВЯЗИ РАЗМЕРОВ ОСТРОВА С ЧИСЛОМ ВИДОВ) Уменьшение площади острова в десять раз, как правило,
- 39. Энергетическая структура экосистемы
- 40. С точки зрения трофических отношений экосистема состоит из двух групп организмов: автотрофных (самостоятельно «питающихся», осуществляющих, в
- 41. В составе экосистемы выделяют следующие компоненты: неорганические вещества (С, N, Р, CO2, H2O и т.д.); органические
- 42. макроконсументы или фаготрофы (гетеротрофные организмы, главным образом животные, которые поедают другие организмы или частицы органического вещества);
- 43. Р. Вигерт и Д. Оуэнс разделяют гетеротрофов на две группы (учитывается разрыв во времени между потреблением
- 44. ПИЩЕВЫЕ ЦЕПИ И ЭКОЛОГИЧЕСКИЕ ПИРАМИДЫ Внутри экосистемы органические вещества создаются автотрофными организмами (например, растениями). Растения поедают
- 45. Организмы первого трофического уровня называются первичными продуцентами. На суше большую часть продуцентов составляют растения лесов и
- 46. Организмы второго трофического уровня называются первичными консументами, третьего трофического уровня – вторичными консументами и т. д.
- 47. Существует ещё одна группа организмов, называемых редуцентами. Это сапрофиты (обычно, бактерии и грибы), питающие-ся органическими остатками
- 48. В схемах пищевых цепей каждый организм представлен питающим-ся организмами какого-то определённого типа. Действительность намного сложнее, и
- 49. ЗАКОН ПИРАМИДЫ ЧИСЕЛ Элтона Число индивидуумов в последовательности трофических уровней убывает и формирует пирамиду чисел. Закон
- 50. Упрощённый вариант экологической пирамиды В трофических цепях, где энергия передается в основном через связи системы «хищник
- 51. ЗАКОН ПИРАМИДЫ БИОМАСС Пирамиды биомасс представляют более фундаментальный интерес, так как они дают «...картину общего влияния
- 52. ЗАКОН ПИРАМИДЫ ПРОДУКТИВНОСТИ Пирамида продуктивности - более стабильная пирамида, чем пирамида чисел или пирамида биомасс, которая
- 53. Рис. Три типа экологических пирамид для пищевой цепи «люцерна – теленок – мальчик» (Одум, 1975, с.
- 54. ГИПОТЕЗА КОНСОРТИВНЫХ СВЯЗЕЙ Беклемишева – Раменского Консорция - основная ячейка трансформации энергии в экосистеме, являющаяся ее
- 56. Скачать презентацию


Разновысотность растений и «разноглубинность» - важные оси дифференциации экологических ниш в
Разновысотность растений и «разноглубинность» - важные оси дифференциации экологических ниш в


Два основных варианта разновысотности:
Ярусность, когда на глаз видно расчленение биоценоза по
Два основных варианта разновысотности:
Ярусность, когда на глаз видно расчленение биоценоза по

Пример экосистемы с четкой ярусной дифференциацией
Пример экосистемы с четкой ярусной дифференциацией

Пример экосистемы с нечетко выраженной ярусностью
Пример экосистемы с нечетко выраженной ярусностью
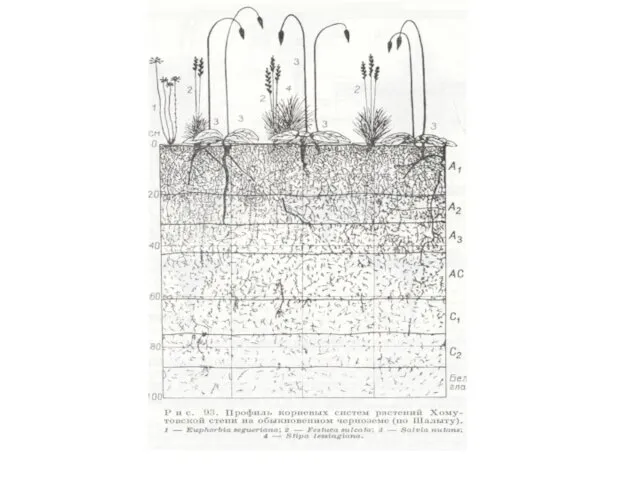

Горизонтальная структура экосистемы.
Мозаичность – это явление неоднородности горизонтального сложения биоценоза. Горизонтально
Горизонтальная структура экосистемы.
Мозаичность – это явление неоднородности горизонтального сложения биоценоза. Горизонтально



Соотношение дискретности и непрерывности в экосистемах – один из интереснейших и
Соотношение дискретности и непрерывности в экосистемах – один из интереснейших и

Представления о дискретности экосистем (организмистские аналогии) связаны с работами американского эколога
Представления о дискретности экосистем (организмистские аналогии) связаны с работами американского эколога

Новые представления о непрерывности растительного покрова – теория континуума
Леонтий Григорьевич
Раменский (1884-1953)
Генри
Новые представления о непрерывности растительного покрова – теория континуума
Леонтий Григорьевич
Раменский (1884-1953)
Генри

Идеи континуума "пробили себе дорогу" лишь в 50-60-х годах ХХ в.,
Идеи континуума "пробили себе дорогу" лишь в 50-60-х годах ХХ в.,

Зависимость степени непрерывности растительного
покрова от числа доминантов и видового разнообразия
непрерывность
(тропический лес)
дискретность
(бореальный
покрова от числа доминантов и видового разнообразия
непрерывность
(тропический лес)
дискретность
(бореальный

Коцепция единства дискретности и непрерывности в экологии
Экосистемы представляют собой явление природы
Коцепция единства дискретности и непрерывности в экологии
Экосистемы представляют собой явление природы

МЕТОДЫ (ЗАКОНЫ) ОРДИНАЦИИ
Методы анализа и описания закономерностей распределения видов или
МЕТОДЫ (ЗАКОНЫ) ОРДИНАЦИИ
Методы анализа и описания закономерностей распределения видов или

По методам различают ординации:
прямую (ординация ведется по реальным факторам среды
По методам различают ординации:
прямую (ординация ведется по реальным факторам среды

Прямой градиентный анализ – один из наиболее эффективных методов ординации, который
Прямой градиентный анализ – один из наиболее эффективных методов ординации, который

Факторный анализ – раздел статистического многомерного анализа, объединяющий методы оценки размерности
Факторный анализ – раздел статистического многомерного анализа, объединяющий методы оценки размерности
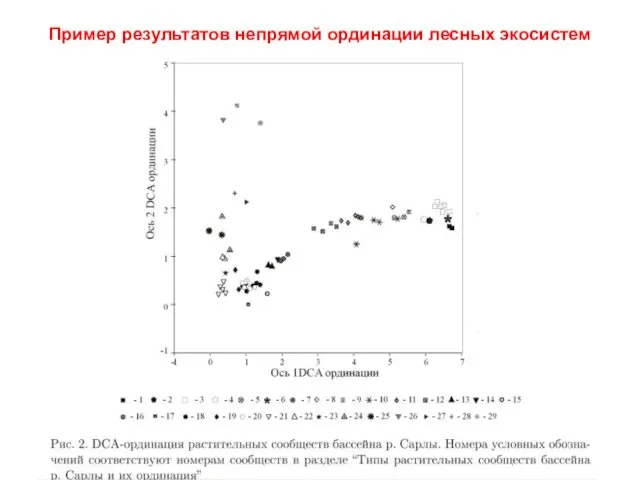
Пример результатов непрямой ординации лесных экосистем
Пример результатов непрямой ординации лесных экосистем

Биоразнообразие экосистем
Видовое богатство – общее количество видов живых существ в сообществе.
Чем
Биоразнообразие экосистем
Видовое богатство – общее количество видов живых существ в сообществе.
Чем

Научный (академический) интерес к проблеме биоразнообразия связан с возможностью познания механизмов
Научный (академический) интерес к проблеме биоразнообразия связан с возможностью познания механизмов

ГИПОТЕЗЫ АЛЬФА-, БЕТА- И ГАММА-РАЗНООБРАЗИЯ
Роберт Уиттекер (R. Whittaker) в 1960
ГИПОТЕЗЫ АЛЬФА-, БЕТА- И ГАММА-РАЗНООБРАЗИЯ
Роберт Уиттекер (R. Whittaker) в 1960

бета-разнообразие (разнообразие между сообществами, показатель степени дифференцированности распределения видов или скорости
бета-разнообразие (разнообразие между сообществами, показатель степени дифференцированности распределения видов или скорости

БИОЦЕНОТИЧЕСКИЕ ПРИНЦИПЫ Тинемана
Сформулированные немец- ким гидробиологом Августом Тинеманом в 1939
БИОЦЕНОТИЧЕСКИЕ ПРИНЦИПЫ Тинемана
Сформулированные немец- ким гидробиологом Августом Тинеманом в 1939

чем больше отклоняются от нормы (оптимума) условия существования в пределах биотопа,
чем больше отклоняются от нормы (оптимума) условия существования в пределах биотопа,

В качестве примеров можно назвать:
процесс «цветения» водохранилищ равнинного типа (массовое развитие
В качестве примеров можно назвать:
процесс «цветения» водохранилищ равнинного типа (массовое развитие

Индексы биоразнообразия
индекс (показатель) Симпсона
(Еdward Hugh Simpson; публикация 1951 г.;
Индексы биоразнообразия
индекс (показатель) Симпсона
(Еdward Hugh Simpson; публикация 1951 г.;

МОДЕЛИ РАСПРЕДЕЛЕНИЯ ЗНАЧИМОСТИ ВИДОВ
Формализация кривых относительных оценок значимости видов в сообществе:
Роберт
МОДЕЛИ РАСПРЕДЕЛЕНИЯ ЗНАЧИМОСТИ ВИДОВ
Формализация кривых относительных оценок значимости видов в сообществе:
Роберт

ПОСТУЛАТЫ ВИДОВОГО ОБЕДНЕНИЯ
Основные закономерности, которые автоматиче-ски осуществляются в ходе нарушения
ПОСТУЛАТЫ ВИДОВОГО ОБЕДНЕНИЯ
Основные закономерности, которые автоматиче-ски осуществляются в ходе нарушения

вновь внедрившийся вид приводит к перераспределению пространства экологических ниш сообщества, сужает
вновь внедрившийся вид приводит к перераспределению пространства экологических ниш сообщества, сужает

с антропоцентристской точки зрения замена видов или трофических цепей (сетей) может
с антропоцентристской точки зрения замена видов или трофических цепей (сетей) может

ПРАВИЛО КРОГЕРУСА О ДОМИНИРОВАНИИ ВИДОВ В ЭКСТРЕМАЛЬНЫХ УСЛОВИЯХ
В биотопах с
ПРАВИЛО КРОГЕРУСА О ДОМИНИРОВАНИИ ВИДОВ В ЭКСТРЕМАЛЬНЫХ УСЛОВИЯХ
В биотопах с

ПРАВИЛО ДЕ КАНДОЛЯ – УОЛЛЕСА (ГЕОГРАФИЧЕСКАЯ ОБУСЛОВЛЕННОСТЬ ИЗМЕНЕНИЯ РАЗНООБРАЗИЯ)
По мере
ПРАВИЛО ДЕ КАНДОЛЯ – УОЛЛЕСА (ГЕОГРАФИЧЕСКАЯ ОБУСЛОВЛЕННОСТЬ ИЗМЕНЕНИЯ РАЗНООБРАЗИЯ)
По мере

ПРАВИЛО МИНИМУМА ВИДОВ Ремане
Парадокс солоноватых вод, согласно которому минимум разнообразия
ПРАВИЛО МИНИМУМА ВИДОВ Ремане
Парадокс солоноватых вод, согласно которому минимум разнообразия
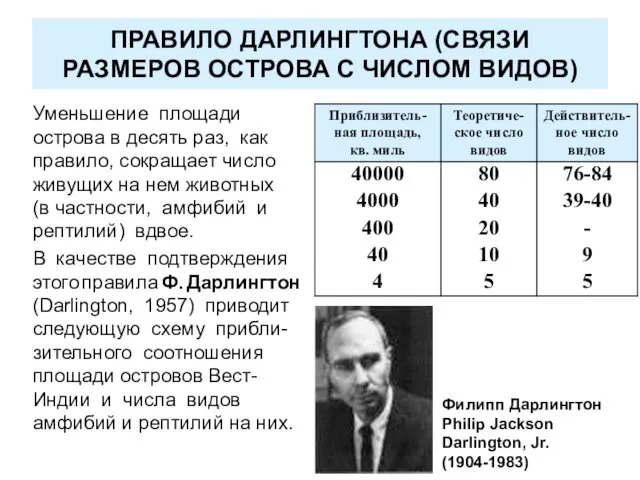
ПРАВИЛО ДАРЛИНГТОНА (СВЯЗИ РАЗМЕРОВ ОСТРОВА С ЧИСЛОМ ВИДОВ)
Уменьшение площади острова
ПРАВИЛО ДАРЛИНГТОНА (СВЯЗИ РАЗМЕРОВ ОСТРОВА С ЧИСЛОМ ВИДОВ)
Уменьшение площади острова

Энергетическая структура экосистемы
Энергетическая структура экосистемы

С точки зрения трофических отношений экосистема состоит из двух групп организмов:
С точки зрения трофических отношений экосистема состоит из двух групп организмов:

В составе экосистемы выделяют следующие компоненты:
неорганические вещества (С, N, Р, CO2,
В составе экосистемы выделяют следующие компоненты:
неорганические вещества (С, N, Р, CO2,

макроконсументы или фаготрофы (гетеротрофные организмы, главным образом животные, которые поедают другие
макроконсументы или фаготрофы (гетеротрофные организмы, главным образом животные, которые поедают другие

Р. Вигерт и Д. Оуэнс разделяют гетеротрофов на две группы (учитывается
Р. Вигерт и Д. Оуэнс разделяют гетеротрофов на две группы (учитывается
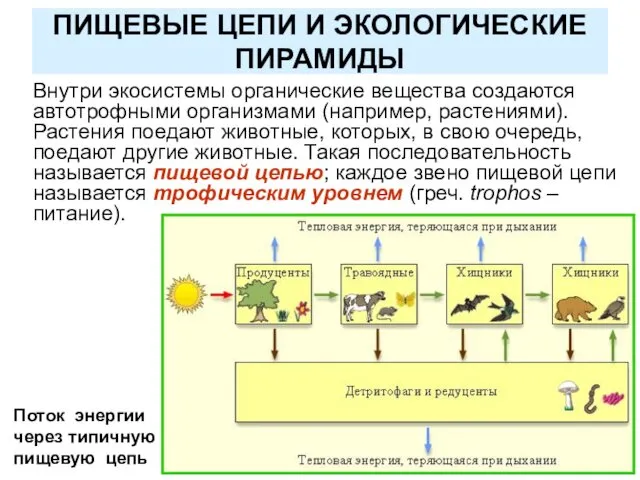
ПИЩЕВЫЕ ЦЕПИ И ЭКОЛОГИЧЕСКИЕ ПИРАМИДЫ
Внутри экосистемы органические вещества создаются автотрофными
ПИЩЕВЫЕ ЦЕПИ И ЭКОЛОГИЧЕСКИЕ ПИРАМИДЫ
Внутри экосистемы органические вещества создаются автотрофными

Организмы первого трофического уровня называются первичными продуцентами.
На суше большую часть
Организмы первого трофического уровня называются первичными продуцентами.
На суше большую часть

Организмы второго трофического уровня называются первичными консументами, третьего трофического уровня –
Организмы второго трофического уровня называются первичными консументами, третьего трофического уровня –

Существует ещё одна группа организмов, называемых редуцентами.
Это сапрофиты (обычно, бактерии
Существует ещё одна группа организмов, называемых редуцентами.
Это сапрофиты (обычно, бактерии
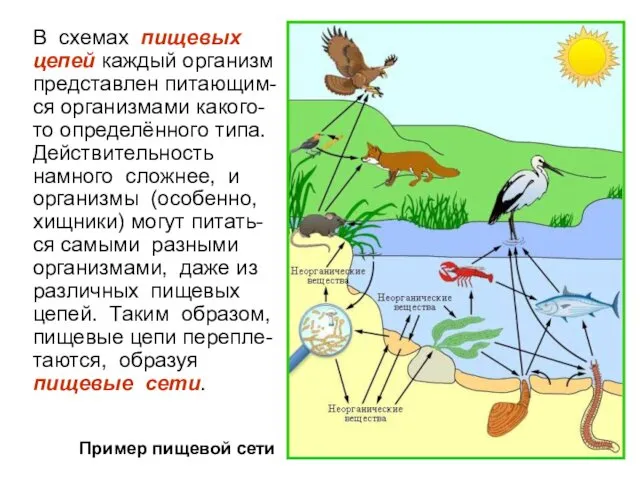
В схемах пищевых цепей каждый организм представлен питающим-ся организмами какого-то определённого
В схемах пищевых цепей каждый организм представлен питающим-ся организмами какого-то определённого

ЗАКОН ПИРАМИДЫ ЧИСЕЛ Элтона
Число индивидуумов в последовательности трофических уровней убывает и
ЗАКОН ПИРАМИДЫ ЧИСЕЛ Элтона
Число индивидуумов в последовательности трофических уровней убывает и
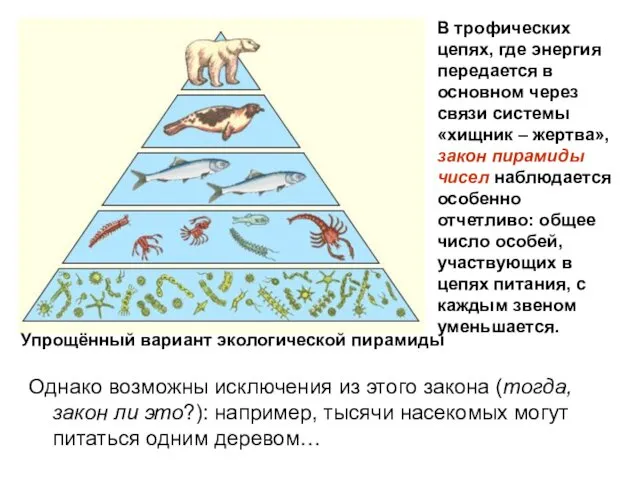
Упрощённый вариант экологической пирамиды
В трофических цепях, где энергия передается в основном
Упрощённый вариант экологической пирамиды
В трофических цепях, где энергия передается в основном
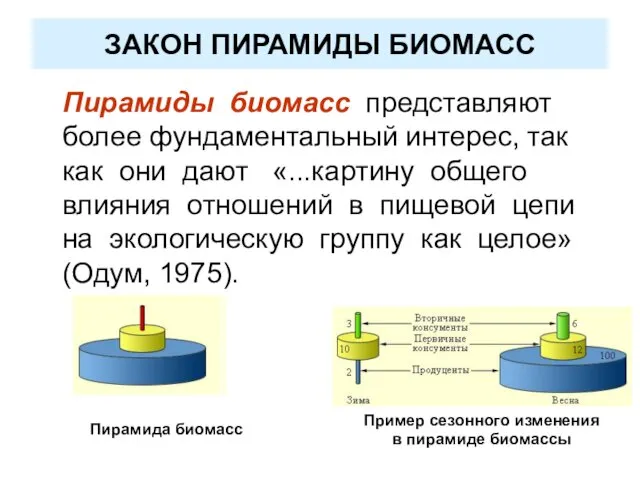
ЗАКОН ПИРАМИДЫ БИОМАСС
Пирамиды биомасс представляют более фундаментальный интерес, так как они
ЗАКОН ПИРАМИДЫ БИОМАСС
Пирамиды биомасс представляют более фундаментальный интерес, так как они

ЗАКОН ПИРАМИДЫ ПРОДУКТИВНОСТИ
Пирамида продуктивности - более стабильная пирамида, чем пирамида чисел
ЗАКОН ПИРАМИДЫ ПРОДУКТИВНОСТИ
Пирамида продуктивности - более стабильная пирамида, чем пирамида чисел
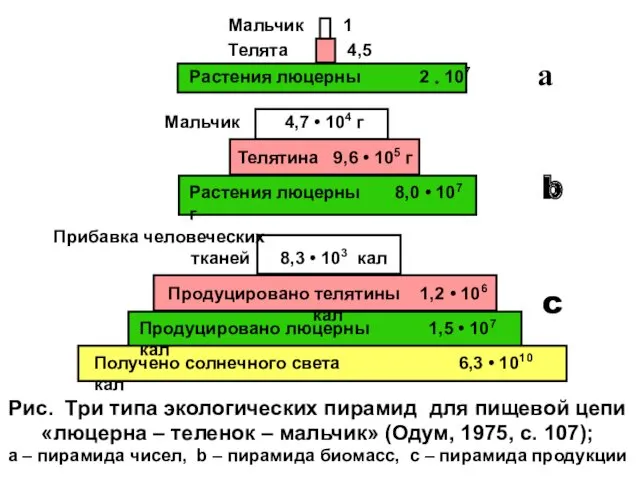
Рис. Три типа экологических пирамид для пищевой цепи
«люцерна – теленок –
Рис. Три типа экологических пирамид для пищевой цепи
«люцерна – теленок –

ГИПОТЕЗА КОНСОРТИВНЫХ СВЯЗЕЙ Беклемишева – Раменского
Консорция - основная ячейка
ГИПОТЕЗА КОНСОРТИВНЫХ СВЯЗЕЙ Беклемишева – Раменского
Консорция - основная ячейка
 Презентация Мой город
Презентация Мой город Адмiнiстративне i кримiнальне право. Головнi аспекти
Адмiнiстративне i кримiнальне право. Головнi аспекти Напряженно-деформированное состояние оболочечных конструкций, выполненных из материалов с усложненными механическими свойствами
Напряженно-деформированное состояние оболочечных конструкций, выполненных из материалов с усложненными механическими свойствами Познавательный проект на тему: Удомельский район в годы Великой Отечественной войны
Познавательный проект на тему: Удомельский район в годы Великой Отечественной войны Матрицы. Определитель матрицы
Матрицы. Определитель матрицы Дизартрия. Классификация клинических форм дизартрии
Дизартрия. Классификация клинических форм дизартрии radio_siti
radio_siti Тренажёр для подготовки к аттестационному тестированию по педагогике
Тренажёр для подготовки к аттестационному тестированию по педагогике Сюжетно-ролевая игра – средство формирования у дошкольников элементарных знаний об окружающем мире презентация
Сюжетно-ролевая игра – средство формирования у дошкольников элементарных знаний об окружающем мире презентация Резьбообрабатывающие станки
Резьбообрабатывающие станки Литературный поиск. Базы данных
Литературный поиск. Базы данных Наречие как часть речи. Правописание наречий
Наречие как часть речи. Правописание наречий Секты современной России
Секты современной России Проектирование, строительство и эксплуатация автомобильных дорог. Исследование характеристик колейности по Башкортостану
Проектирование, строительство и эксплуатация автомобильных дорог. Исследование характеристик колейности по Башкортостану Алексей Николаевич Плещеев В бурю
Алексей Николаевич Плещеев В бурю Буклет для родителей Информационная безопасность детей в Интернете
Буклет для родителей Информационная безопасность детей в Интернете Карцов Владимир Геннадьевич и Гиттис Изабелла Васильевна
Карцов Владимир Геннадьевич и Гиттис Изабелла Васильевна Собрание для родителей будущих первоклассников
Собрание для родителей будущих первоклассников Контур многократной принудительной циркуляции
Контур многократной принудительной циркуляции Анимация. Мультипликация. Виды анимации
Анимация. Мультипликация. Виды анимации Мягкие лекарственные формы
Мягкие лекарственные формы Неравновесные явления в растворах электролитов. Электрическая проводимость растворов электролитов
Неравновесные явления в растворах электролитов. Электрическая проводимость растворов электролитов урок Серная кислота
урок Серная кислота Творческий проект Логарифмическая линейка
Творческий проект Логарифмическая линейка Разработка ресурсосберегающего способа производства топливно-плавильных материалов на основе техногенного сырья (Н3)
Разработка ресурсосберегающего способа производства топливно-плавильных материалов на основе техногенного сырья (Н3) Представление целых чисел на координатной оси
Представление целых чисел на координатной оси Основы ландшафтной колористики
Основы ландшафтной колористики Песня по картинке
Песня по картинке