- Сцепленное наследование и рекомбинация

Содержание
- 2. Хромосомы Гены Последовательности нуклеотидов в ДНК Располагаются на хромосоме линейно Сцепленные гены Гены расположенные на одной
- 3. Генетическое сцепление и его последствия Генетическое сцепление - это явление, когда определенные гены, передаются по наследству
- 4. Генетическое сцепление. (а) Сцепленные гены AB/ab передаются одним блоком от одного поколения к другому. (б) Генетическое
- 5. Рекомбинация
- 6. Генетическая рекомбинация Гомологичные хромосомы обмениваются участками, содержащими альтернативные аллели генов Обмен участками (кроссинговер) происходит в пахитене
- 7. Сцепленное наследование Эксперимент. Вопрос? Наследуются ли признаки «окраска цветка» и «форма пыльцевого зерна» независимо?
- 8. Сцепленное наследование Вывод: Расщепление фенотипов в потомстве F2 не соответствует ожидаемому при независимом наследовании признаков соотношению
- 9. Сцепленные гены наследуются совместно, а кроссинговер приводит к их рекомбинации Гены одной хромосомы Сцепленные гены Группа
- 10. Особенности сцепленного наследования Полное сцепление приводит к формированию нерекомбинантных гамет и нерекомбинантного потомства. Кроссинговер между сцепленными
- 11. Доказательство сцепления генов Плодовая мушка Drosophila melanogaster
- 12. Особенности сцепленного наследования Аллели указывают на гомологичных хромосомах X A B A B a b a
- 13. Особенности сцепленного наследования Принятые обозначения : X A B A B a b a b A
- 14. Особенности сцепленного наследования Необходимо запомнить, что Аллели генов располагаются на одном участке гомологичных хромосом A a
- 15. Особенности сцепленного наследования Необходимо запомнить, что Порядок расположения генов на гомологичных хромосомах одинаков A B b
- 16. К счастью для селекционеров гены в хромосомах сцеплены не полностью. Если бы это было так, то
- 17. Лекция 4 Кроссинговер Кроссинговер – процесс обмена участками несестринских хроматид гомологичных хромосом в профазе I мейоза
- 18. Схема кроссинговера Нет кроссинговера Все хромосомы в гаметах нерекомбинантные Кроссинговер в профазе I Половина гамет содержит
- 19. Цитологическое Доказательство кроссинговера Г. Крейтон и Б. Мак-Клинток создали линию кукурузы, в которой хромосомы IX пары
- 20. Лекция 4 Частота рекомбинации Т. Морган и А. Стертевант первыми высказали гипотезу о линейном расположении генов:
- 21. Расчет частоты рекомбинации Частоту рекомбинации (rf) определяют в потомствах от анализирующего скрещивания, как отношения числа кроссоверных
- 22. Расчет частоты рекомбинации Пример:
- 23. Частота кроссинговера и линейное расположение генов в хромосоме В одном из опытов Морган и его сотрудники
- 24. Данные о наследовании, сцепленном с полом, о нерасхождении хромосом, о сцепленном наследовании и кроссинговере, наряду с
- 25. Частота рекомбинации Частота рекомбинации эквивалентна расстоянию между генами Чем дальше расположены ген от гена, тем больше
- 26. Частота рекомбинации Сильно удаленные друг от друга гены на хромосоме Проявляют практически независимое наследование Сцепление между
- 27. Хромосомные (генетические) карты Частота рекомбинации используется для определения относительного положения генов на хромосоме Сцепление генов a,
- 28. Картирование хромосом Селекционеры растений создают и используют «биологические карты» в качестве проводника в своей работе. Существует
- 29. Генетической картой хромосом называют схему относительного расположения генов на хромосоме, выраженную в единицах кроссинговера. На генной
- 30. Если в промежутке между двумя локусами кроссинговер происходит довольно часто, то гены находятся на сравнительно большом
- 31. Генетическая карта: Drosophila Хромосома 2
- 32. Цитоплазматическая наследственность
- 33. Схема организации генетического материала клетки
- 34. Цитоплазматическое наследование Митохондрии: 37 генов Множественные органеллы Каждая содержит множество копий ДНК (2-10) Хлоропласты Органеллы передаются
- 35. Цитоплазматическое наследование Особенности расщепления в прямом и обратном скрещивании различны (реципрокный эффект); органеллы распределяются при цитокинезе
- 36. Нехромосомная наследственность Явление нехромосомной наследственности было открыто в 1909 г. К.Корренсом и Э.Бауром при анализе наследования
- 37. Наследование пестролистности у ночной красавицы
- 38. Случайное распределение митохондрий при делении клетки
- 39. Мужская стерильность Мужская стерильность - это явление, при котором у растения пыльники или пыльца нефункциональны. Может
- 40. Мужская стерильность 1. Истинная мужская стерильность. Связана с однополыми цветками у которых отсутствуют тычинки (при двудомности
- 41. Генетическая (ядерная, генная) мужская стерильность Генетическая (ядерная, генная) мужская стерильность широко распространена у растений. Ген стерильность
- 42. Селекционер в случае ядерной стерильности не может производить и поддерживать популяцию, имеющую 100% растений с мужской
- 43. Цитоплазматическая мужская стерильность Цитоплазматическая мужская стерильность (ЦМС) контролируется генами цитоплазмы (митохондриальным геном). ЦМС передается только через
- 44. Морфологическое проявление цитоплазматической мужской стерильности у капусты Схема получения трехлинейных гибридов на основе ЦМС Цветок фертильный
- 45. Схема получения трехлинейных гибридов на основе ЦМС
- 46. Цитоплазматическая мужская стерильность Обнаружена у многих высших растений (более 300). Среди сельскохозяйственных культур у: кукурузы, лука,
- 47. Ядерно-цитоплазматическая мужская стерильность У кукурузы обнаружены 8 ядерных генов (Rf1-Rf8), взаимодействующих со стерильной цитоплазмой Взаимодействие генов
- 48. Ядерно-цитоплазматическая мужская стерильность (ЯЦМС) Используется в производстве гибридных семян лука и моркови Этот тип стерильности определяется
- 49. Схема получения гибридных семян на основе ЯЦМС S msms S msms S msms N msms N
- 50. Использование мужской стерильности в селекции Мужская стерильность в основном используется в качестве инструмента в селекции растений
- 51. Cytoplasmic Inheritance WT and mutant genes in mitochondria Random segregation Progeny cells contain different numbers of
- 52. Four O’clock Flowers Variegated leaves Progeny phenotype not determined providing pollen Progeny phenotype determined by branch
- 53. Sex linkage explained Thomas Hunt Morgan in The Fly Room! (Columbia University 1910) Fruit Flies (Drosophila
- 54. Tortioseshell Cats are Female © 2007 Paul Billiet ODWS
- 55. Daltonism = Red-Green Colourblindness Normal vision Colour blind simulation http://www.onset.unsw.edu.au/issue1/colourblindness/colourblindness_print.htm © 2007 Paul Billiet ODWS
- 56. Комплементарное взаимодействие генов Наследование формы гребня у кур: доминантные аллели каждого из генов имеют собственное фенотипическое
- 57. Комплементарное взаимодействие генов Наследование окраски цветка у гороха: доминантные аллели каждого из генов А (А-bb) и
- 58. Комплементарное взаимодействие генов Наследование окраски чешуй у лука: доминантные аллели каждого из генов С (C-rr) и
- 59. Эпистатические взаимодействия. Рецессивный эпистаз. Наследование окраски чешуй у лука: доминантные аллели каждого из генов С (C-rr)
- 60. Эпистатическое взаимодействие генов
- 61. Эпистатические взаимодействия. Рецессивный эпистаз.
- 62. Эпистатические взаимодействия. Супрессия. Наследование окраски оперения у кур: Доминантный аллель I – гена-супрессора не имеет собственного
- 63. Пенетрантность и экспрессивность Пенетрантность – доля особей, проявляющих данный признак, среди всех особей одного генотипа по
- 64. Пенетрантность и экспрессивность Экспрессивность – степень выраженности признака.
- 65. Хромосомное определение пола У большинства организмов соматические клетки женских особей имеют две гомологичные половые хромосомы (обычно
- 66. Environmental Effects on Plant Phenotypes
- 68. Скачать презентацию

Хромосомы
Гены
Последовательности нуклеотидов в ДНК
Располагаются на хромосоме линейно
Сцепленные гены
Гены расположенные на одной
Хромосомы
Гены
Последовательности нуклеотидов в ДНК
Располагаются на хромосоме линейно
Сцепленные гены
Гены расположенные на одной

Генетическое сцепление и его последствия
Генетическое сцепление - это явление, когда определенные
Генетическое сцепление и его последствия
Генетическое сцепление - это явление, когда определенные
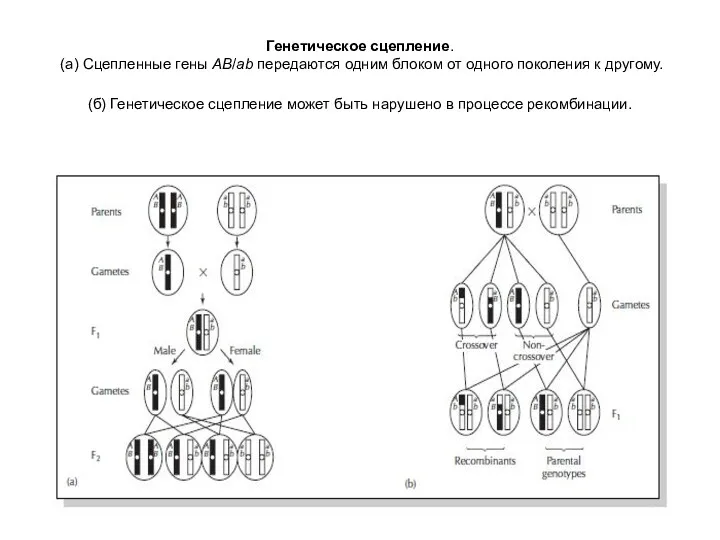
Генетическое сцепление.
(а) Сцепленные гены AB/ab передаются одним блоком от одного
Генетическое сцепление. (а) Сцепленные гены AB/ab передаются одним блоком от одного
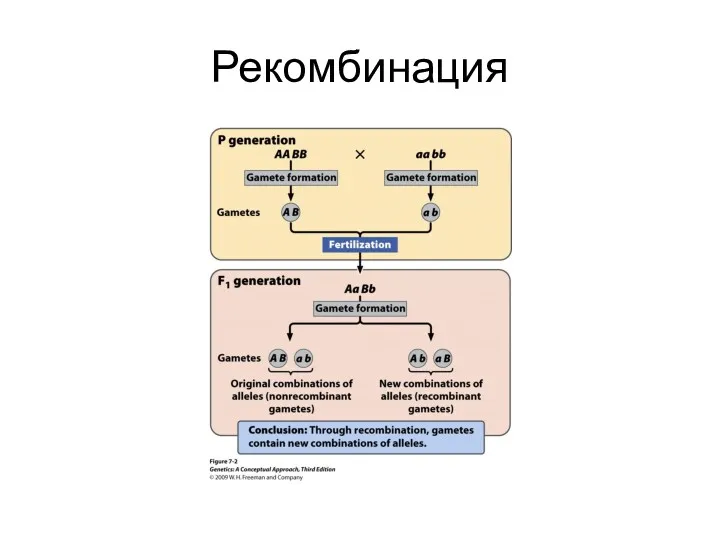
Рекомбинация
Рекомбинация

Генетическая рекомбинация
Гомологичные хромосомы обмениваются участками, содержащими альтернативные аллели генов
Обмен участками
Генетическая рекомбинация
Гомологичные хромосомы обмениваются участками, содержащими альтернативные аллели генов
Обмен участками
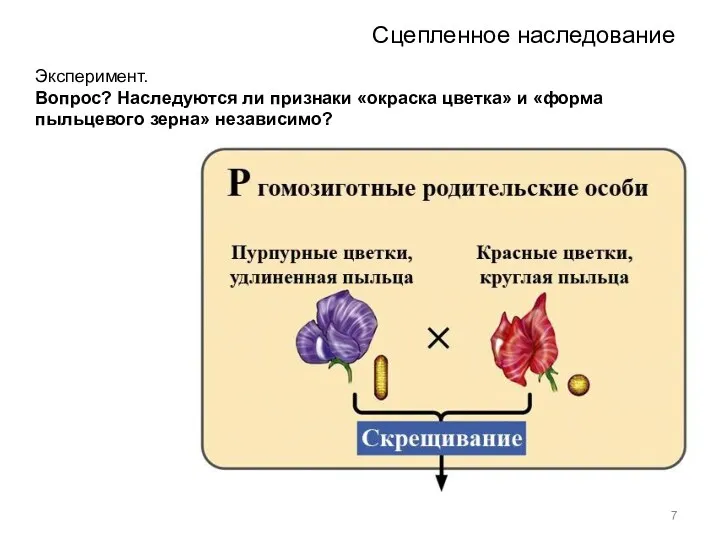
Сцепленное наследование
Эксперимент.
Вопрос? Наследуются ли признаки «окраска цветка» и «форма пыльцевого зерна»
Сцепленное наследование
Эксперимент.
Вопрос? Наследуются ли признаки «окраска цветка» и «форма пыльцевого зерна»
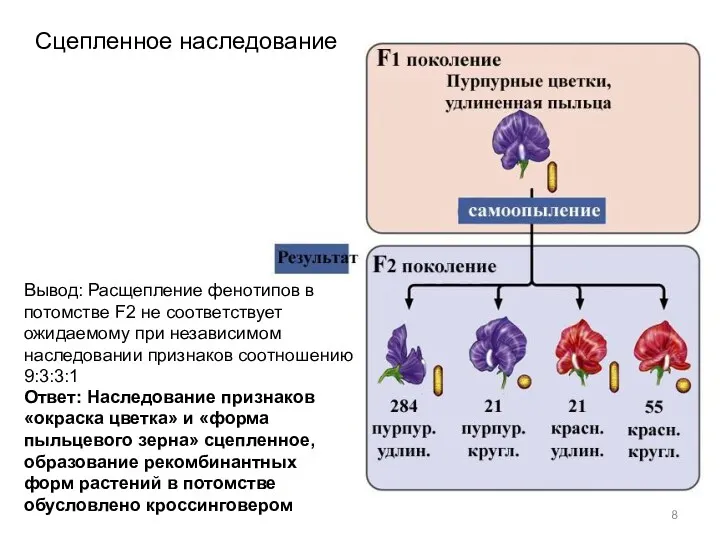
Сцепленное наследование
Вывод: Расщепление фенотипов в потомстве F2 не соответствует ожидаемому при
Сцепленное наследование
Вывод: Расщепление фенотипов в потомстве F2 не соответствует ожидаемому при

Сцепленные гены наследуются совместно, а кроссинговер приводит к их рекомбинации
Гены
Сцепленные гены наследуются совместно, а кроссинговер приводит к их рекомбинации
Гены
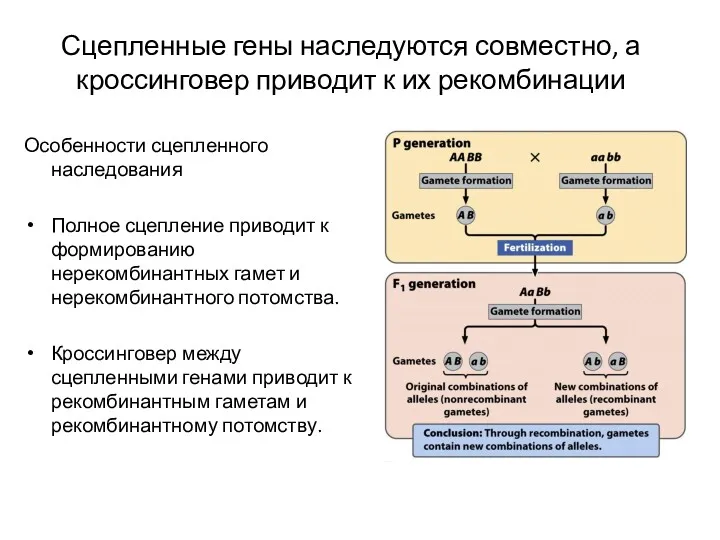
Особенности сцепленного наследования
Полное сцепление приводит к формированию нерекомбинантных гамет и нерекомбинантного
Особенности сцепленного наследования
Полное сцепление приводит к формированию нерекомбинантных гамет и нерекомбинантного

Доказательство сцепления генов
Плодовая мушка Drosophila melanogaster
Доказательство сцепления генов
Плодовая мушка Drosophila melanogaster
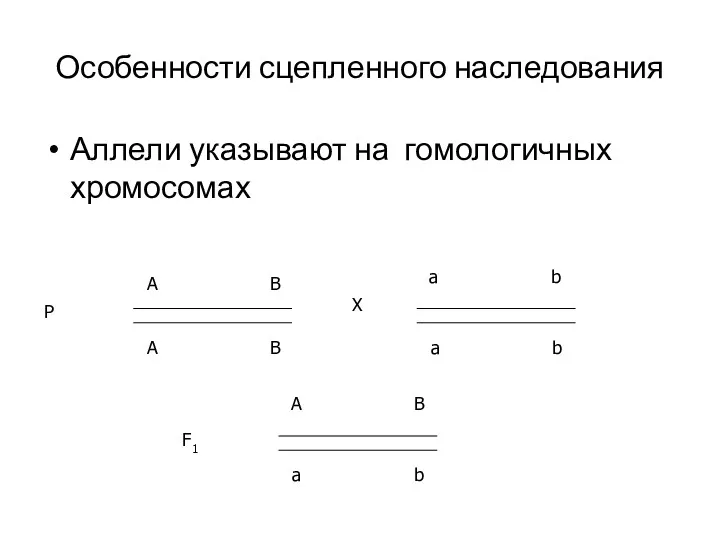
Особенности сцепленного наследования
Аллели указывают на гомологичных хромосомах
X
A B
A B
a b
a b
A
Особенности сцепленного наследования
Аллели указывают на гомологичных хромосомах
X
A B
A B
a b
a b
A
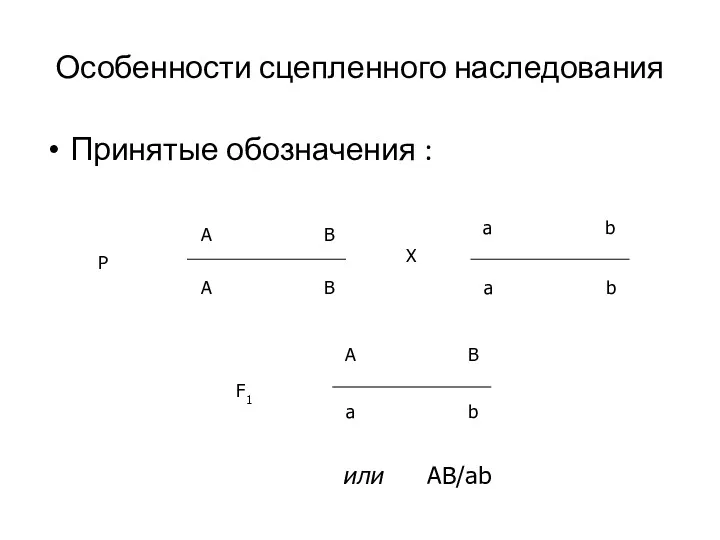
Особенности сцепленного наследования
Принятые обозначения :
X
A B
A B
a b
a b
A B
a b
P
F1
или
Особенности сцепленного наследования
Принятые обозначения :
X
A B
A B
a b
a b
A B
a b
P
F1
или

Особенности сцепленного наследования
Необходимо запомнить, что
Аллели генов располагаются на одном участке гомологичных
Особенности сцепленного наследования
Необходимо запомнить, что
Аллели генов располагаются на одном участке гомологичных

Особенности сцепленного наследования
Необходимо запомнить, что
Порядок расположения генов на гомологичных хромосомах одинаков
A
Особенности сцепленного наследования
Необходимо запомнить, что
Порядок расположения генов на гомологичных хромосомах одинаков
A

К счастью для селекционеров гены в хромосомах сцеплены не полностью. Если
К счастью для селекционеров гены в хромосомах сцеплены не полностью. Если

Лекция 4
Кроссинговер
Кроссинговер – процесс обмена участками несестринских хроматид гомологичных хромосом в
Лекция 4
Кроссинговер
Кроссинговер – процесс обмена участками несестринских хроматид гомологичных хромосом в
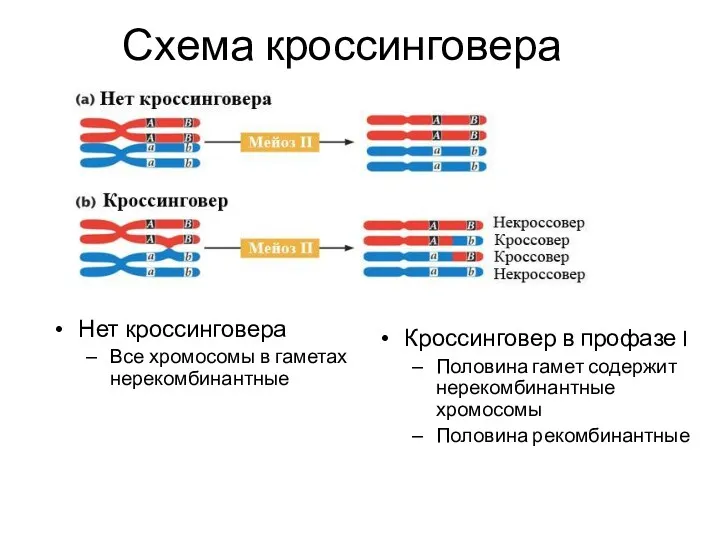
Схема кроссинговера
Нет кроссинговера
Все хромосомы в гаметах нерекомбинантные
Кроссинговер в профазе I
Половина гамет
Схема кроссинговера
Нет кроссинговера
Все хромосомы в гаметах нерекомбинантные
Кроссинговер в профазе I
Половина гамет

Цитологическое Доказательство кроссинговера
Г. Крейтон и Б. Мак-Клинток создали линию кукурузы, в
Цитологическое Доказательство кроссинговера
Г. Крейтон и Б. Мак-Клинток создали линию кукурузы, в

Лекция 4
Частота рекомбинации
Т. Морган и А. Стертевант первыми высказали гипотезу о
Лекция 4
Частота рекомбинации
Т. Морган и А. Стертевант первыми высказали гипотезу о

Расчет частоты рекомбинации
Частоту рекомбинации (rf) определяют в потомствах от анализирующего скрещивания,
Расчет частоты рекомбинации
Частоту рекомбинации (rf) определяют в потомствах от анализирующего скрещивания,
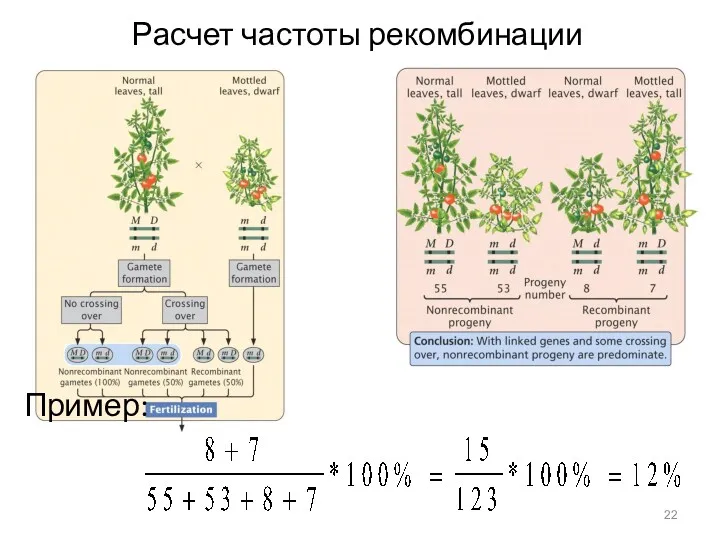
Расчет частоты рекомбинации
Пример:
Расчет частоты рекомбинации
Пример:

Частота кроссинговера и линейное расположение генов в хромосоме
В одном из опытов
Частота кроссинговера и линейное расположение генов в хромосоме
В одном из опытов

Данные о наследовании, сцепленном с полом, о нерасхождении хромосом, о сцепленном
Данные о наследовании, сцепленном с полом, о нерасхождении хромосом, о сцепленном

Частота рекомбинации
Частота рекомбинации эквивалентна расстоянию между генами
Чем дальше расположены ген от
Частота рекомбинации
Частота рекомбинации эквивалентна расстоянию между генами
Чем дальше расположены ген от

Частота рекомбинации
Сильно удаленные друг от друга гены на хромосоме
Проявляют практически независимое
Частота рекомбинации
Сильно удаленные друг от друга гены на хромосоме
Проявляют практически независимое
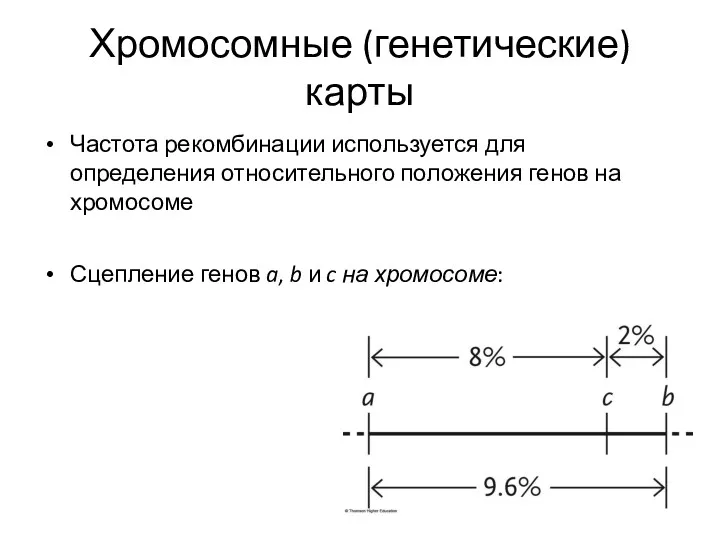
Хромосомные (генетические) карты
Частота рекомбинации используется для определения относительного положения генов на
Хромосомные (генетические) карты
Частота рекомбинации используется для определения относительного положения генов на

Картирование хромосом
Селекционеры растений создают и используют «биологические карты» в качестве проводника
Картирование хромосом
Селекционеры растений создают и используют «биологические карты» в качестве проводника

Генетической картой хромосом называют схему относительного расположения генов на хромосоме, выраженную
Генетической картой хромосом называют схему относительного расположения генов на хромосоме, выраженную

Если в промежутке между двумя локусами кроссинговер происходит довольно часто, то
Если в промежутке между двумя локусами кроссинговер происходит довольно часто, то
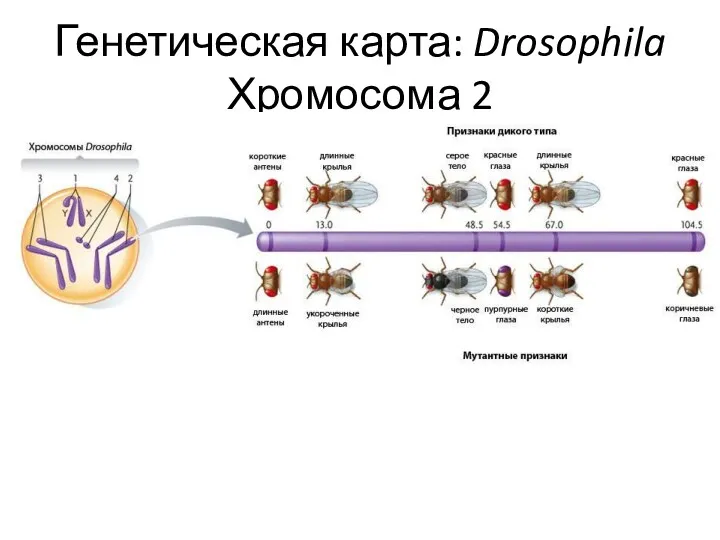
Генетическая карта: Drosophila Хромосома 2
Генетическая карта: Drosophila Хромосома 2

Цитоплазматическая наследственность
Цитоплазматическая наследственность
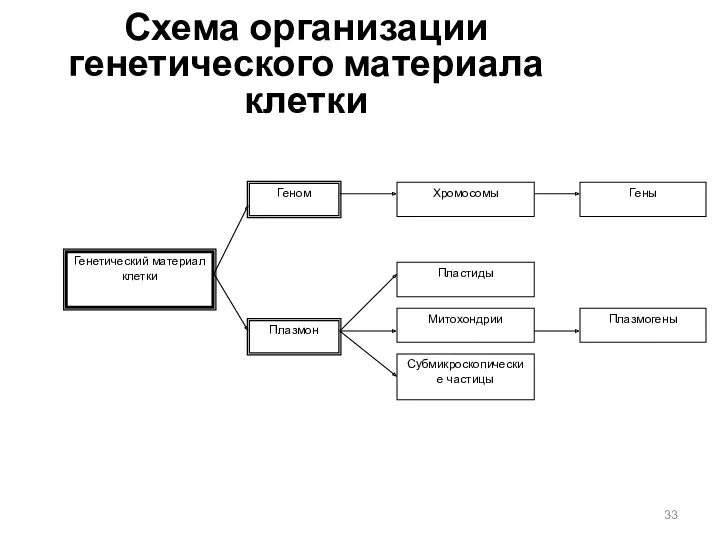
Схема организации генетического материала клетки
Схема организации генетического материала клетки

Цитоплазматическое наследование
Митохондрии: 37 генов
Множественные органеллы
Каждая содержит множество копий ДНК (2-10)
Хлоропласты
Органеллы передаются
Цитоплазматическое наследование
Митохондрии: 37 генов
Множественные органеллы
Каждая содержит множество копий ДНК (2-10)
Хлоропласты
Органеллы передаются

Цитоплазматическое наследование
Особенности
расщепления в прямом и обратном скрещивании различны (реципрокный эффект);
органеллы распределяются
Цитоплазматическое наследование
Особенности
расщепления в прямом и обратном скрещивании различны (реципрокный эффект);
органеллы распределяются

Нехромосомная наследственность
Явление нехромосомной наследственности было открыто в 1909 г. К.Корренсом и
Нехромосомная наследственность
Явление нехромосомной наследственности было открыто в 1909 г. К.Корренсом и

Наследование пестролистности у ночной красавицы
Наследование пестролистности у ночной красавицы
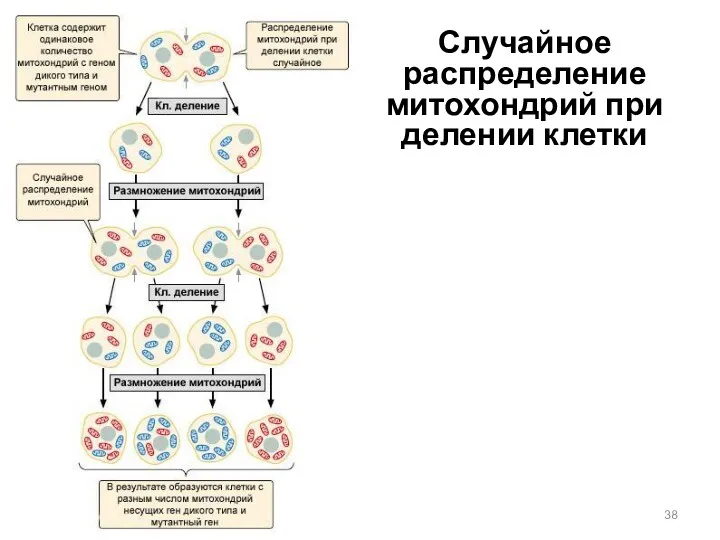
Случайное распределение митохондрий при делении клетки
Случайное распределение митохондрий при делении клетки

Мужская стерильность
Мужская стерильность - это явление, при котором у растения пыльники
Мужская стерильность
Мужская стерильность - это явление, при котором у растения пыльники

Мужская стерильность
1. Истинная мужская стерильность. Связана с однополыми цветками у которых
Мужская стерильность
1. Истинная мужская стерильность. Связана с однополыми цветками у которых

Генетическая (ядерная, генная) мужская стерильность
Генетическая (ядерная, генная) мужская стерильность широко распространена
Генетическая (ядерная, генная) мужская стерильность
Генетическая (ядерная, генная) мужская стерильность широко распространена
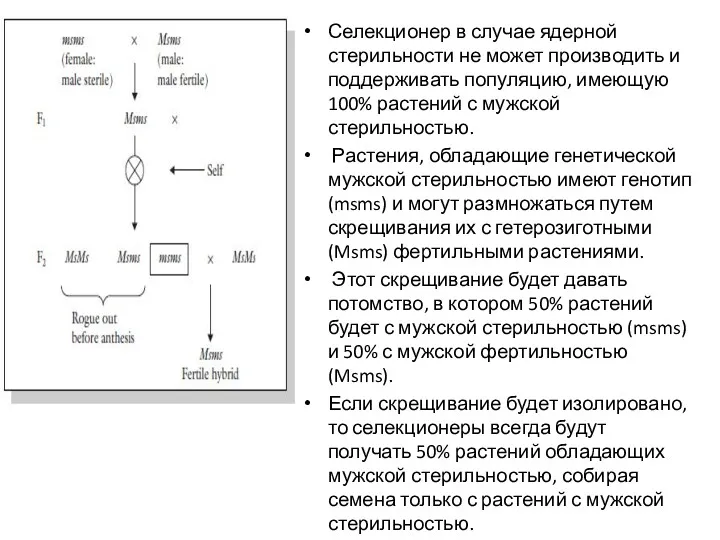
Селекционер в случае ядерной стерильности не может производить и поддерживать популяцию,
Селекционер в случае ядерной стерильности не может производить и поддерживать популяцию,

Цитоплазматическая мужская стерильность
Цитоплазматическая мужская стерильность (ЦМС) контролируется генами цитоплазмы (митохондриальным геном).
Цитоплазматическая мужская стерильность
Цитоплазматическая мужская стерильность (ЦМС) контролируется генами цитоплазмы (митохондриальным геном).
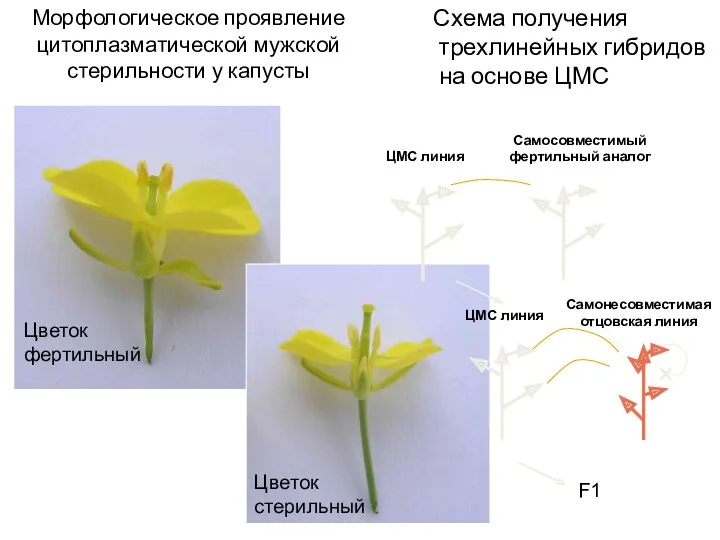
Морфологическое проявление цитоплазматической мужской стерильности у капусты
Схема получения трехлинейных гибридов
Морфологическое проявление цитоплазматической мужской стерильности у капусты
Схема получения трехлинейных гибридов
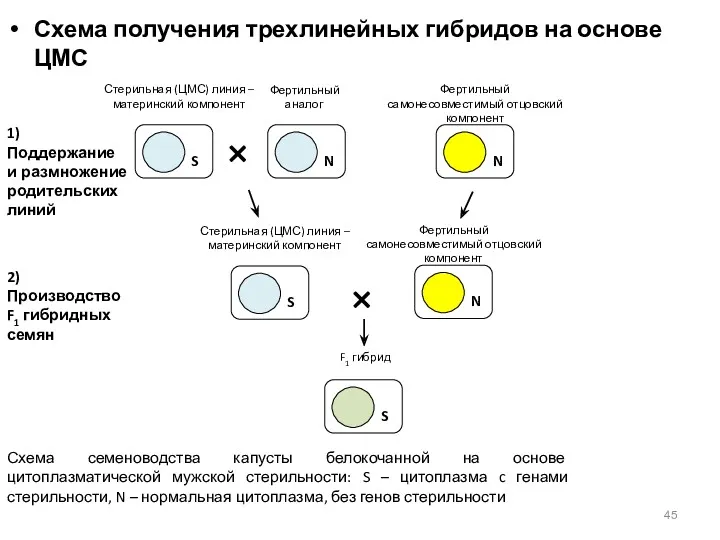
Схема получения трехлинейных гибридов на основе ЦМС
Схема получения трехлинейных гибридов на основе ЦМС

Цитоплазматическая мужская стерильность
Обнаружена у многих высших растений (более 300). Среди сельскохозяйственных
Цитоплазматическая мужская стерильность
Обнаружена у многих высших растений (более 300). Среди сельскохозяйственных

Ядерно-цитоплазматическая мужская стерильность
У кукурузы обнаружены 8 ядерных генов (Rf1-Rf8), взаимодействующих со
Ядерно-цитоплазматическая мужская стерильность
У кукурузы обнаружены 8 ядерных генов (Rf1-Rf8), взаимодействующих со

Ядерно-цитоплазматическая мужская стерильность (ЯЦМС)
Используется в производстве гибридных семян лука и моркови
Этот
Ядерно-цитоплазматическая мужская стерильность (ЯЦМС)
Используется в производстве гибридных семян лука и моркови
Этот
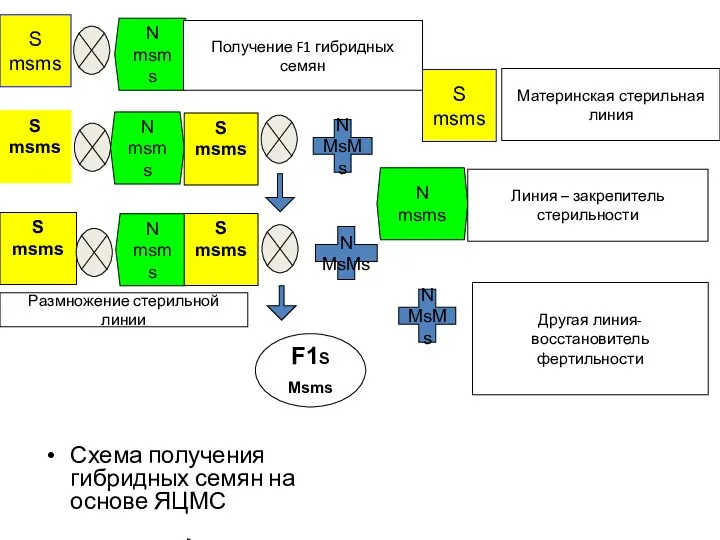
Схема получения гибридных семян на основе ЯЦМС
S msms
S msms
S msms
N msms
N
Схема получения гибридных семян на основе ЯЦМС
S msms
S msms
S msms
N msms
N

Использование мужской стерильности в селекции
Мужская стерильность в основном используется в качестве
Использование мужской стерильности в селекции
Мужская стерильность в основном используется в качестве
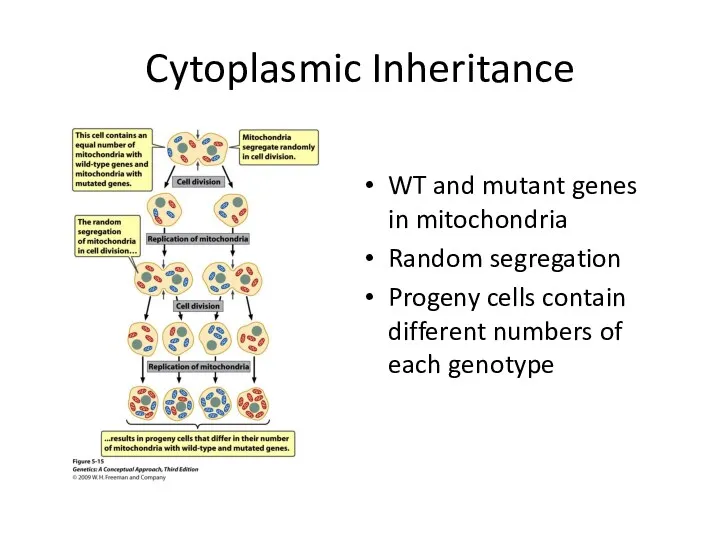
Cytoplasmic Inheritance
WT and mutant genes in mitochondria
Random segregation
Progeny cells contain different
Cytoplasmic Inheritance
WT and mutant genes in mitochondria
Random segregation
Progeny cells contain different
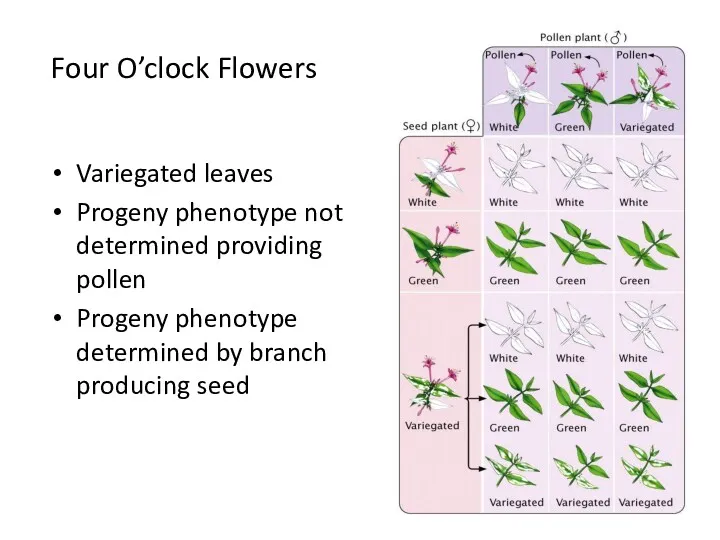
Four O’clock Flowers
Variegated leaves
Progeny phenotype not determined providing pollen
Progeny phenotype determined
Four O’clock Flowers
Variegated leaves
Progeny phenotype not determined providing pollen
Progeny phenotype determined

Sex linkage explained
Thomas Hunt Morgan in The Fly Room!
(Columbia University
Sex linkage explained
Thomas Hunt Morgan in The Fly Room! (Columbia University

Tortioseshell Cats are Female
© 2007 Paul Billiet ODWS
Tortioseshell Cats are Female
© 2007 Paul Billiet ODWS

Daltonism = Red-Green Colourblindness
Normal vision
Colour blind simulation
http://www.onset.unsw.edu.au/issue1/colourblindness/colourblindness_print.htm
© 2007 Paul Billiet
Daltonism = Red-Green Colourblindness
Normal vision
Colour blind simulation
http://www.onset.unsw.edu.au/issue1/colourblindness/colourblindness_print.htm
© 2007 Paul Billiet
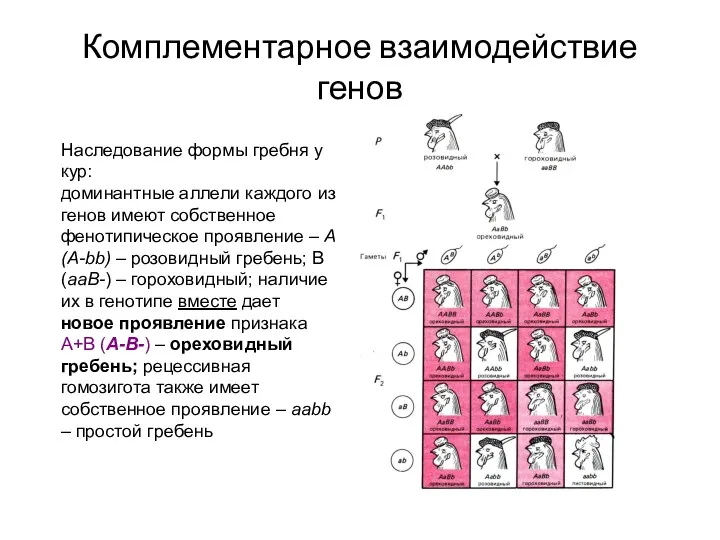
Комплементарное взаимодействие генов
Наследование формы гребня у кур:
доминантные аллели каждого из генов
Комплементарное взаимодействие генов
Наследование формы гребня у кур:
доминантные аллели каждого из генов
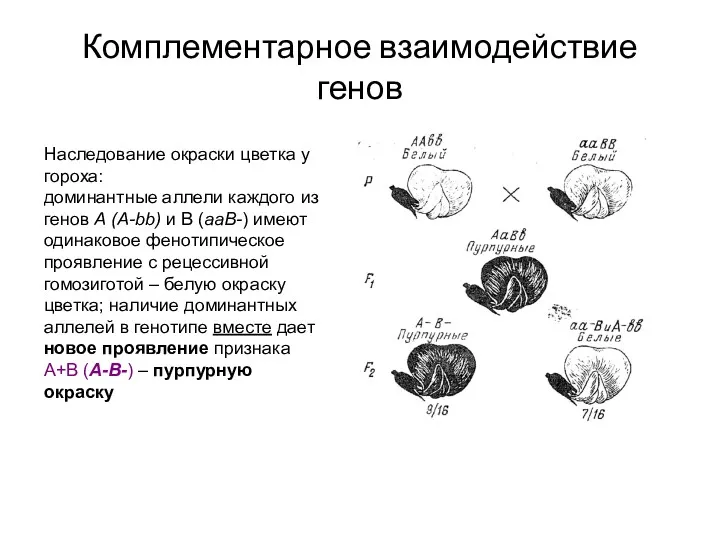
Комплементарное взаимодействие генов
Наследование окраски цветка у гороха:
доминантные аллели каждого из генов
Комплементарное взаимодействие генов
Наследование окраски цветка у гороха:
доминантные аллели каждого из генов
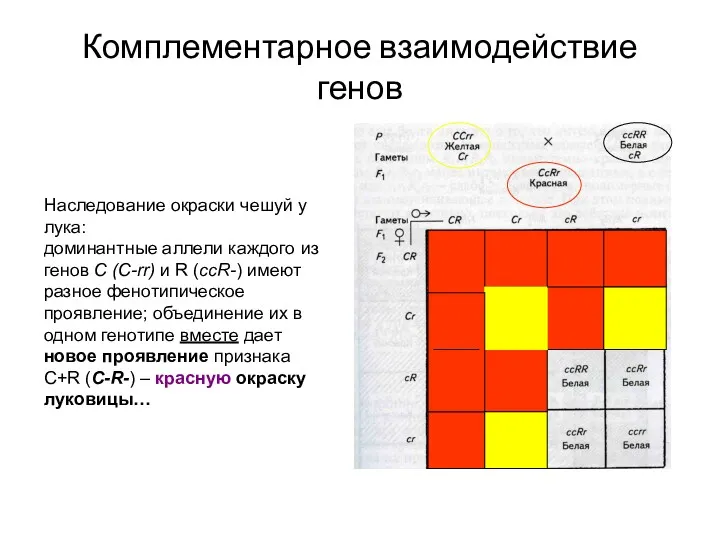
Комплементарное взаимодействие генов
Наследование окраски чешуй у лука:
доминантные аллели каждого из генов
Комплементарное взаимодействие генов
Наследование окраски чешуй у лука:
доминантные аллели каждого из генов

Эпистатические взаимодействия. Рецессивный эпистаз.
Наследование окраски чешуй у лука:
доминантные аллели каждого из
Эпистатические взаимодействия. Рецессивный эпистаз.
Наследование окраски чешуй у лука:
доминантные аллели каждого из

Эпистатическое взаимодействие генов
Эпистатическое взаимодействие генов
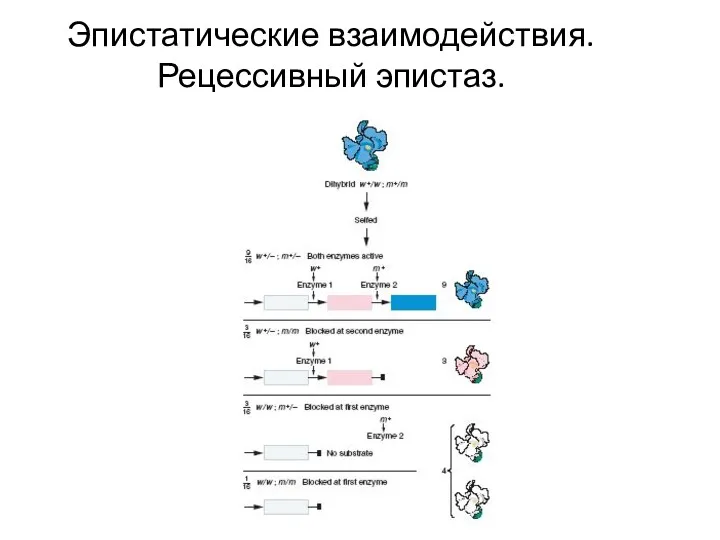
Эпистатические взаимодействия. Рецессивный эпистаз.
Эпистатические взаимодействия. Рецессивный эпистаз.
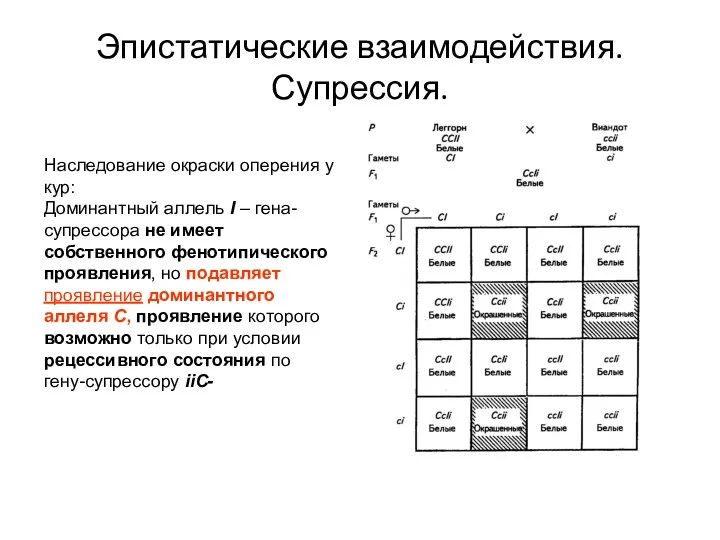
Эпистатические взаимодействия. Супрессия.
Наследование окраски оперения у кур:
Доминантный аллель I – гена-супрессора
Эпистатические взаимодействия. Супрессия.
Наследование окраски оперения у кур:
Доминантный аллель I – гена-супрессора
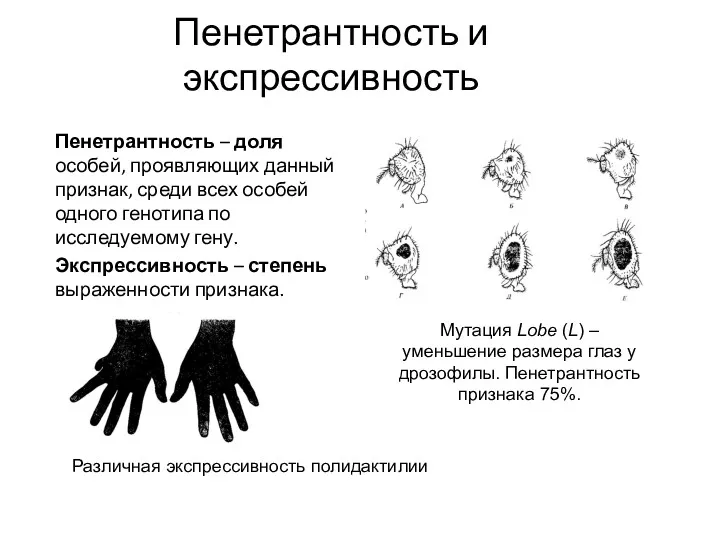
Пенетрантность и экспрессивность
Пенетрантность – доля особей, проявляющих данный признак, среди всех
Пенетрантность и экспрессивность
Пенетрантность – доля особей, проявляющих данный признак, среди всех

Пенетрантность и экспрессивность
Экспрессивность – степень выраженности признака.
Пенетрантность и экспрессивность
Экспрессивность – степень выраженности признака.

Хромосомное определение пола
У большинства организмов соматические клетки женских особей имеют две
Хромосомное определение пола
У большинства организмов соматические клетки женских особей имеют две

Environmental Effects
on Plant Phenotypes
Environmental Effects
on Plant Phenotypes
 Матрешки. Виды матрешек
Матрешки. Виды матрешек Simatic S7
Simatic S7 Гуманитариум. Вопросы трудоустройства выпускников гуманитарных специальностей
Гуманитариум. Вопросы трудоустройства выпускников гуманитарных специальностей 20230626_summa_arifmeticheskoy_progressii
20230626_summa_arifmeticheskoy_progressii Введение в робототехнику
Введение в робототехнику Стандартные образцы в цветной металлургии
Стандартные образцы в цветной металлургии Презентация День Героев Отечества
Презентация День Героев Отечества организация проектной и исследовательской деятельности школьников в разных возрастных группах в условиях реализации современных образовательных стандартов.
организация проектной и исследовательской деятельности школьников в разных возрастных группах в условиях реализации современных образовательных стандартов. Гельминты в почве. Аскаридоз
Гельминты в почве. Аскаридоз Древняя Русь IX-XII вв.: генезис и специфика феодальных отношений
Древняя Русь IX-XII вв.: генезис и специфика феодальных отношений Калькулятор. Принцип работі
Калькулятор. Принцип работі Автоматическая стабилизация и управление углом тангажа
Автоматическая стабилизация и управление углом тангажа Множество и его элементы
Множество и его элементы Рисуем Прелюдию до мажор I том ХТК. 5 класс
Рисуем Прелюдию до мажор I том ХТК. 5 класс Вспомогательные здания и сооружения
Вспомогательные здания и сооружения Основы работы в Naumen Service Desk Почты России
Основы работы в Naumen Service Desk Почты России Алкалоиды и их классификация
Алкалоиды и их классификация Отчёт по производственной электромонтажной практике
Отчёт по производственной электромонтажной практике Информационная безопасность. Комплексная модель безопасности. (Лекция 14)
Информационная безопасность. Комплексная модель безопасности. (Лекция 14) Учимся рисовать. Воробей, цапля
Учимся рисовать. Воробей, цапля Лекция № 10. Элементы релятивистской механики
Лекция № 10. Элементы релятивистской механики Ваш ребенок пятиклассник
Ваш ребенок пятиклассник Обеспечение режимов работы усилительных каскадов на транзисторах (лекция 3)
Обеспечение режимов работы усилительных каскадов на транзисторах (лекция 3) Массивы. Сортировки
Массивы. Сортировки 1654847282750884
1654847282750884 Общий обзор организма человека
Общий обзор организма человека Методология педагогической науки
Методология педагогической науки Проект Ой вы, птицы, к нам летите, весну красную несите!
Проект Ой вы, птицы, к нам летите, весну красную несите!