- Transcriptional regualtion. Repression: Hypoxic Genes in Yeast

Содержание
- 2. Regulation of gene expression Almost as important as the genetic repertoire itself The chimp and human
- 3. A yeast model for repression of gene transcription The transcription of the yeast ANB1 gene is
- 4. Isolation of mutations affecting ANB1 repression Inversion of regulatory region Part of the regulatory region between
- 5. Characterizing mutations in ANB1 regulation cis-acting mutations (mutations on the plasmid in the regulatory region) were
- 6. Characterization of the rox1 mutation The initial rox1 mutant displayed de-repression of the ANB1 gene, as
- 7. Cloning of the rox1 mutation De-repression of hypoxic genes does not have a detectable phenotype Creation
- 8. Cloning of rox1 mutation (2) rox1 mutant cells with integrated ANB1-lacZ fusion on medium containing X-gal
- 9. Cloning of rox1 mutation (3) Grow rox1, ura3::ANB1-lacZ mutant cells Plate on SC- Ura, X-gal Screen
- 10. The Rox1 protein is the repressor of hypoxic genes Rox1p is a DNA – binding repressor
- 11. Rox1p requires Ssn6/Tup1 for repression In a similar screen, mutations in the genes for ROX4 and
- 12. Model of protein and nucleosome interactions at the RNR3 promoter. A, a schematic map of the
- 13. Ssn6/Tup1 recruit HDACs to establish a repressive chromatin structure Tup1 has been demonstrated to directly interact
- 14. URS HYPOXIC Genes
- 15. 2. Ssn6/Tup1 interacts with the RNA poymerase II mediator complex Figure 3. Interactions between Tup1 and
- 16. Oxygen regulation in yeast
- 17. Expression of Hypoxic genes ROX1 ANB1 O2
- 18. Promoter analysis What determines the efficiency of repression? - Sequence of repressor binding sites - Number
- 20. 3 .5 OpA in OpB site 0 .86 43 50 ANB1/lacZ OpA OpB TATA 31 bp
- 21. Role of position for repressor efficiency
- 22. through OpA
- 23. The sequence TGCCT is responsible for stronger repression from OpA
- 24. Insertion of the conserved sequence adjacent to the OpA 5’ Rox1 binding site improves repression from
- 25. MOT3 (Modulator Of Transcription): Mutant derepresses DAN1 (Delayed Anaerobic) and ANB1 Sertil O, Kapoor R, Cohen
- 26. Electrophoretic mobility shift assay (EMSA) Used in analysis of DNA binding properties of proteins Binding target
- 27. EMSA - Principle DNA with binding site DNA – protein complex (High molecular weight, bulky)
- 28. Rox1 Mot3 The Mot3 protein binds specifically to OpA in the ANB1 promoter - 1 5
- 29. A mot3 deletion causes mild derepression of ANB1 Northern blot probing for TIF51A/ANB1 transcripts in wild
- 30. How does Mot3p exert its effect on repression? 1. Interaction with Rox1p? (cooperative binding?) 2. Interaction
- 31. +R1 -R1 20ng Mot3 25ng Rox1 MBP Free DNA 20ngGST- Mot3 Mot3 and Rox1 do not
- 32. A micrococcal nuclease (MNase ) digest reveals chromatin structure of regulatory regions Operator ANB1 TATA Operator
- 33. Mot3 affects the chromatin structure of the ANB1 promoter in a similar manner as Tup1, Ssn6
- 34. MCNase generated digestion pattern is dependent on histone N-termini MNAse
- 35. Summary Operator efficiency: - operator orientation and position relative to the TATA box do only play
- 36. A Model Fungal Gene Regulatory Mechanism: The GAL genes of Saccharomyces cerevisiae GAL genes: involved in
- 37. GAL mutant phenotypes: GAL1, GAL7, GAL10, MEL1, (GAL5): If mutant, cells cannot utilize galactose; a specific
- 38. The GAL structural genes GAL1, GAL7, GAL10, induced >1000x on galactose MEL1 induced >100 x on
- 39. GAL4 and GAL80 are regulatory proteins gal4- : uninducible (recessive) gal80- : constitutive (recessive) Two (very
- 40. promoter Gal4p
- 41. promoter 2. Gal4p is the activator of the GAL genes; Gal80p is a repressor that disables
- 42. Galactose promoter
- 43. promoter Galactose
- 44. How can we distinguish between the two models? Epistasis analysis of pathway What would be the
- 45. Scenario 2 is correct: the gal4-/gal80- mutant is uninducible Gal4p is an activator protein, Gal80p inactivates
- 46. Cloning of the genes gal4- uninducible, cannot grow on plates with galactose as the sole carbon
- 47. The Gal4p Activator The Gal4 protein is a DNA - binding transcriptional activator protein and binds
- 48. Gal4p binds UAS sequences in the regulatory region of GAL structural genes GAL genes UAS UAS:
- 49. lacZ UAS Deletion analysis of promoter region identified Gal4p binding sites Gal4p binding site: 5’ –CGGAG/CGACA-3’
- 50. Gal4p is a modular protein H2N DNA bd Act Activ./Gal80 ia bd
- 51. lacZ UAS VP16 Activation domain (758-881)can activate independently of the rest of the protein if fused
- 52. Expression of GAL4 itself is regulated by glucose Under high glucose concentrations, the DNA – binding
- 53. The galactose sensor: Gal3p Gal3p is a protein with high similarity (homology) to galactokinase No enzymatic
- 54. In a nutshell…. Glucose (repressed): Mig1p Ssn6/Tup1 GAL genes Mig1p Ssn6/Tup1 UAS
- 55. other carbon source than Glucose (derepressed): GAL genes UAS
- 56. Galactose: UAS +Galactose GAL genes
- 57. What is the mechanism of transcriptional activation by Gal4p? A. Gal4p activates by perturbing positioned nucleosomes
- 58. UAS GAL genes TATA
- 59. Micrococcal nuclease digest of chromatin UAS GAL genes TATA UAS GAL genes Radioactive probe anneals to
- 60. Nucleosome Perturbation via recruitment of Histone Acetyl-transferases (HATs)? Histones have positively charged N-terminal tails (K/R –
- 61. B. Gal4p interacts directly with the TATA- binding protein or the polymerase II complex UAS GAL
- 62. Relevance of the Gal regulation research today? General understanding of basic molecular principles of gene activation
- 63. Galactose induction can be utilized to overexpress heterologous genes Genes of interest can be fused to
- 64. Three expression levels: Repressed (2% glucose) ? no expression Derepressed (2% Raffinose, 3% Glycerol) ? intermediate
- 65. Similar: Oleate induction: Oleate induced genes are involved in peroxisomal proliferation and in β-oxidation Activator is
- 67. Скачать презентацию

Regulation of gene expression
Almost as important as the genetic repertoire itself
The
Regulation of gene expression
Almost as important as the genetic repertoire itself
The

A yeast model for repression of gene transcription
The transcription of the
A yeast model for repression of gene transcription
The transcription of the
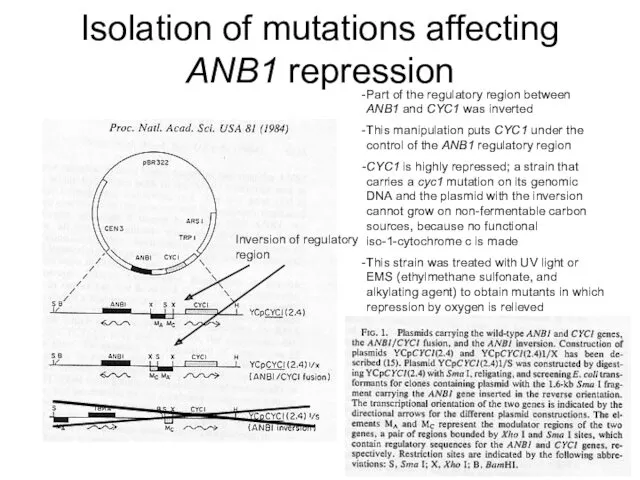
Isolation of mutations affecting ANB1 repression
Inversion of regulatory region
Part of the
Isolation of mutations affecting ANB1 repression
Inversion of regulatory region
Part of the

Characterizing mutations in ANB1 regulation
cis-acting mutations (mutations on the plasmid in
Characterizing mutations in ANB1 regulation
cis-acting mutations (mutations on the plasmid in

Characterization of the rox1 mutation
The initial rox1 mutant displayed de-repression of
Characterization of the rox1 mutation
The initial rox1 mutant displayed de-repression of
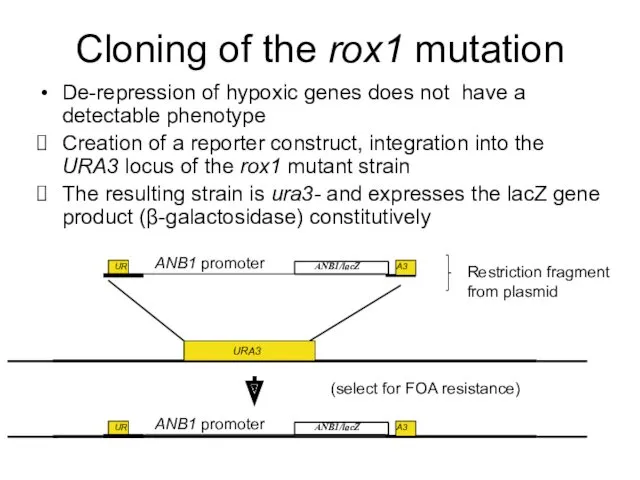
Cloning of the rox1 mutation
De-repression of hypoxic genes does not have
Cloning of the rox1 mutation
De-repression of hypoxic genes does not have
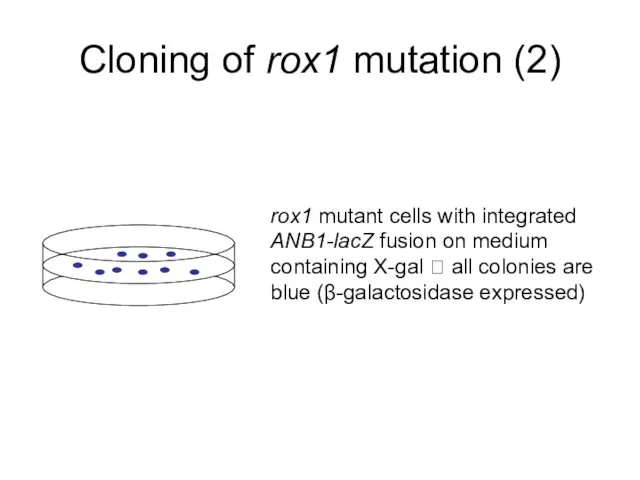
Cloning of rox1 mutation (2)
rox1 mutant cells with integrated ANB1-lacZ fusion
Cloning of rox1 mutation (2)
rox1 mutant cells with integrated ANB1-lacZ fusion
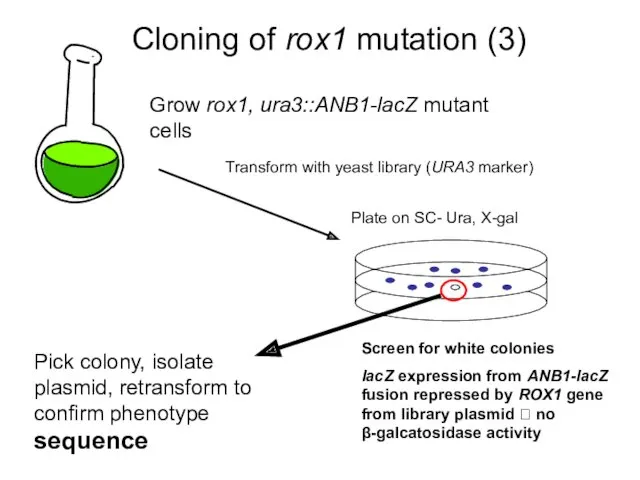
Cloning of rox1 mutation (3)
Grow rox1, ura3::ANB1-lacZ mutant cells
Plate on SC-
Cloning of rox1 mutation (3)
Grow rox1, ura3::ANB1-lacZ mutant cells
Plate on SC-

The Rox1 protein is the repressor of hypoxic genes
Rox1p is a
The Rox1 protein is the repressor of hypoxic genes
Rox1p is a

Rox1p requires Ssn6/Tup1 for repression
In a similar screen, mutations in the
Rox1p requires Ssn6/Tup1 for repression
In a similar screen, mutations in the
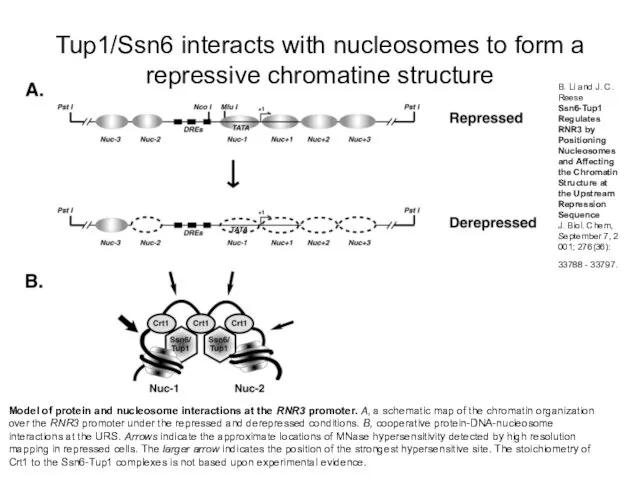
Model of protein and nucleosome interactions at the RNR3 promoter. A,
Model of protein and nucleosome interactions at the RNR3 promoter. A,

Ssn6/Tup1 recruit HDACs to establish a repressive chromatin structure
Tup1 has been
Ssn6/Tup1 recruit HDACs to establish a repressive chromatin structure
Tup1 has been
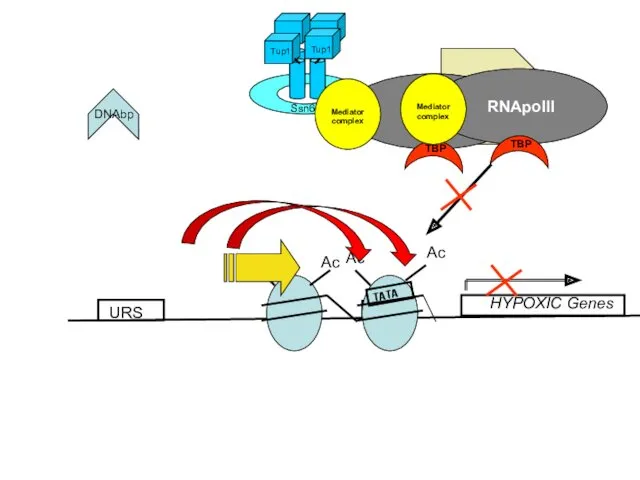
URS
HYPOXIC Genes
URS
HYPOXIC Genes
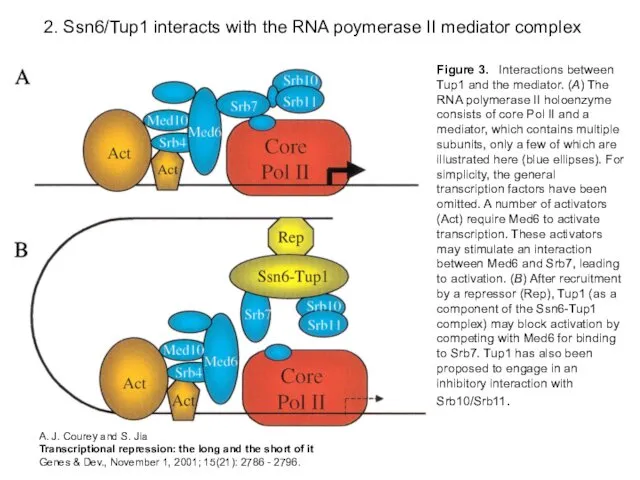
2. Ssn6/Tup1 interacts with the RNA poymerase II mediator complex
Figure 3.
2. Ssn6/Tup1 interacts with the RNA poymerase II mediator complex
Figure 3.
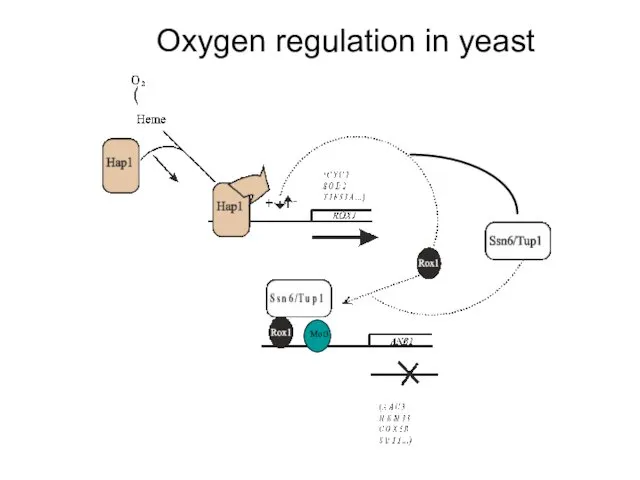
Oxygen regulation in yeast
Oxygen regulation in yeast
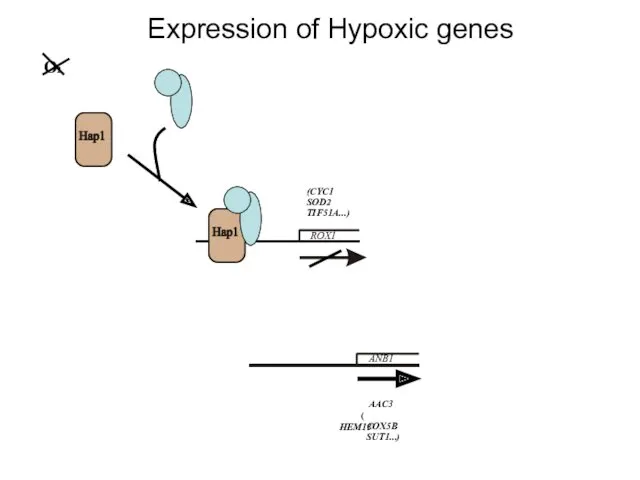
Expression of Hypoxic genes
ROX1
ANB1
O2
Expression of Hypoxic genes
ROX1
ANB1
O2

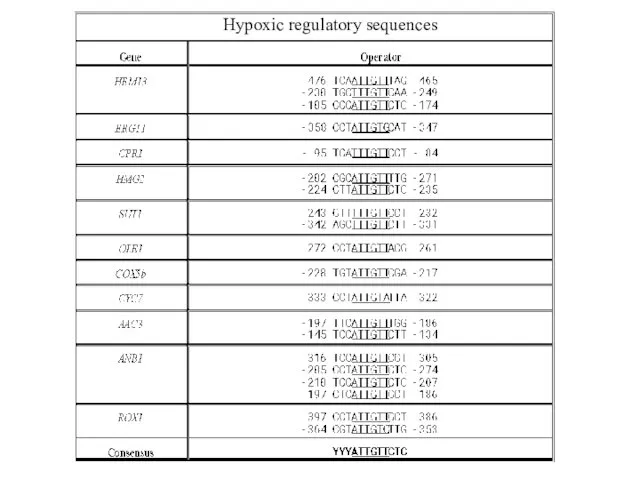
Promoter analysis
What determines the efficiency of repression?
- Sequence of repressor
Promoter analysis
What determines the efficiency of repression?
- Sequence of repressor
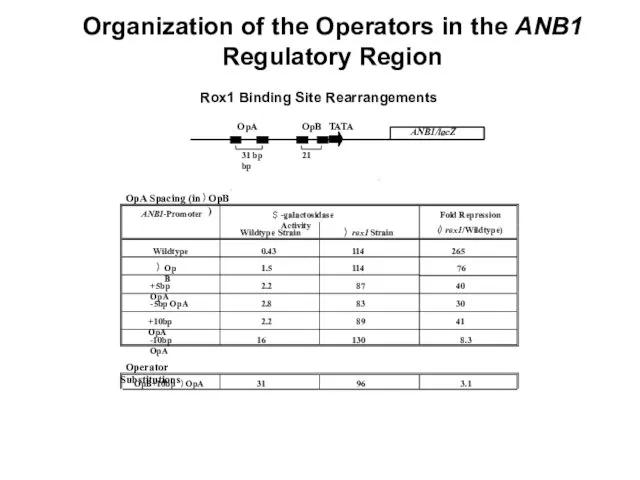
3
.5
OpA in OpB site
0
.86
43
50
ANB1/lacZ
OpA
OpB
TATA
31 bp 21 bp
)
)
)
$
)
)
Organization
3
.5
OpA in OpB site
0
.86
43
50
ANB1/lacZ
OpA
OpB
TATA
31 bp 21 bp
)
)
)
$
)
)
Organization
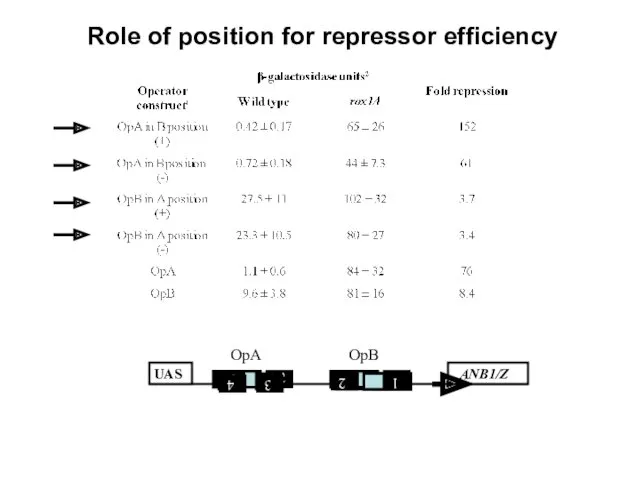
Role of position for repressor efficiency
Role of position for repressor efficiency

through OpA
through OpA
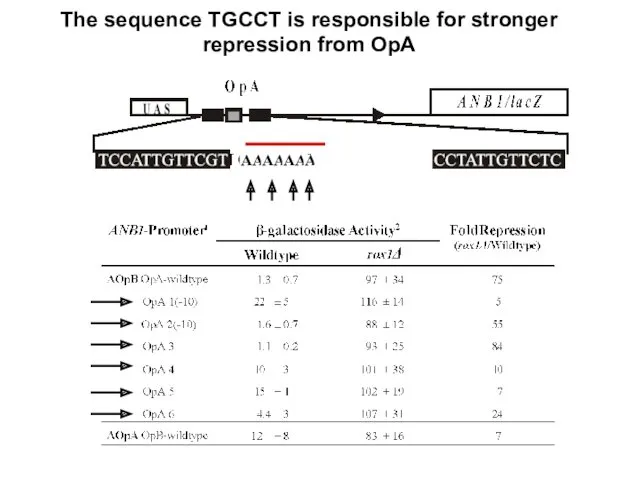
The sequence TGCCT is responsible for stronger repression from OpA
The sequence TGCCT is responsible for stronger repression from OpA

Insertion of the conserved sequence adjacent to the OpA 5’ Rox1
Insertion of the conserved sequence adjacent to the OpA 5’ Rox1

MOT3 (Modulator Of Transcription):
Mutant derepresses DAN1 (Delayed Anaerobic) and ANB1
Sertil O,
MOT3 (Modulator Of Transcription):
Mutant derepresses DAN1 (Delayed Anaerobic) and ANB1
Sertil O,

Electrophoretic mobility shift assay (EMSA)
Used in analysis of DNA binding properties
Electrophoretic mobility shift assay (EMSA)
Used in analysis of DNA binding properties
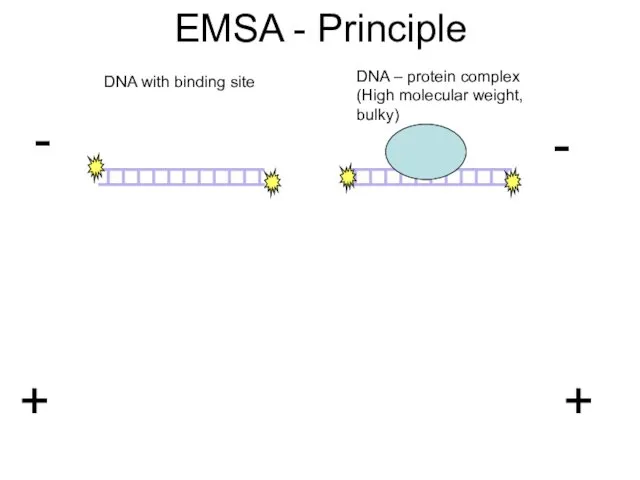
EMSA - Principle
DNA with binding site
DNA – protein complex (High molecular
EMSA - Principle
DNA with binding site
DNA – protein complex (High molecular
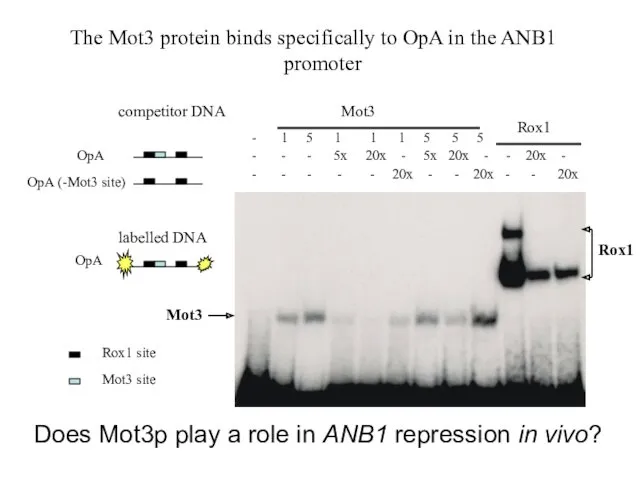
Rox1
Mot3
The Mot3 protein binds specifically to OpA in the
Rox1
Mot3
The Mot3 protein binds specifically to OpA in the
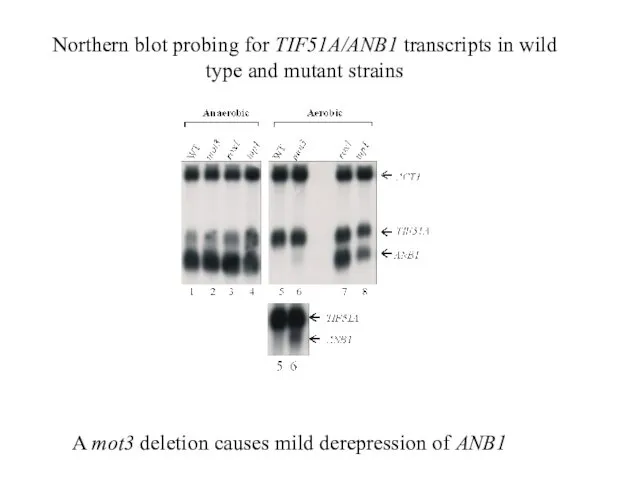
A mot3 deletion causes mild derepression of ANB1
Northern blot probing for
A mot3 deletion causes mild derepression of ANB1
Northern blot probing for

How does Mot3p exert its effect on repression?
1. Interaction with Rox1p?
How does Mot3p exert its effect on repression?
1. Interaction with Rox1p?
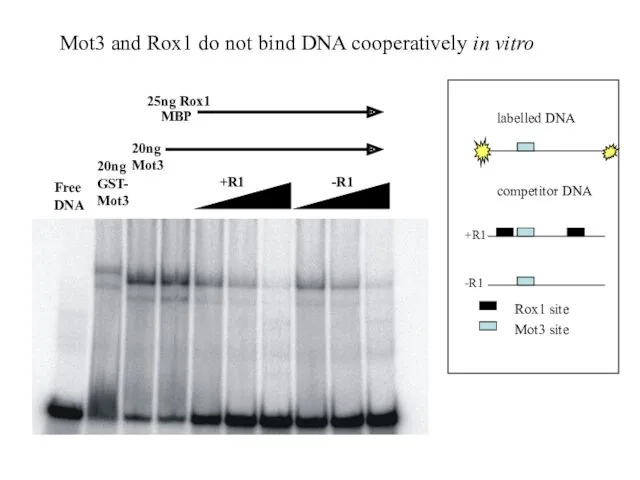
+R1
-R1
20ng
Mot3
25ng Rox1
MBP
Free
DNA
20ngGST-
Mot3
Mot3 and Rox1 do not bind DNA cooperatively
+R1
-R1
20ng
Mot3
25ng Rox1
MBP
Free
DNA
20ngGST-
Mot3
Mot3 and Rox1 do not bind DNA cooperatively
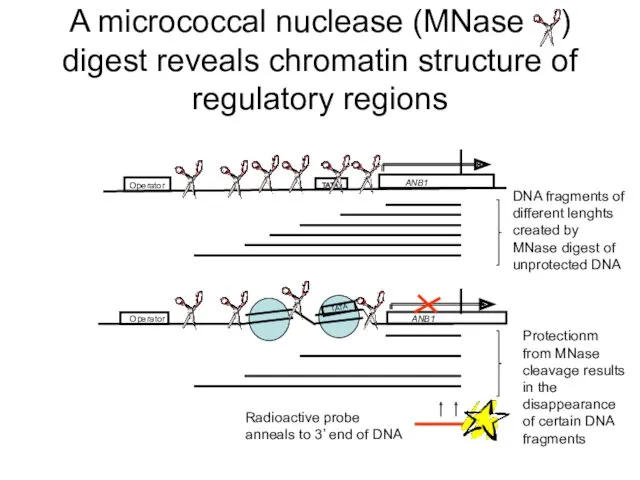
A micrococcal nuclease (MNase ) digest reveals chromatin structure of regulatory
A micrococcal nuclease (MNase ) digest reveals chromatin structure of regulatory
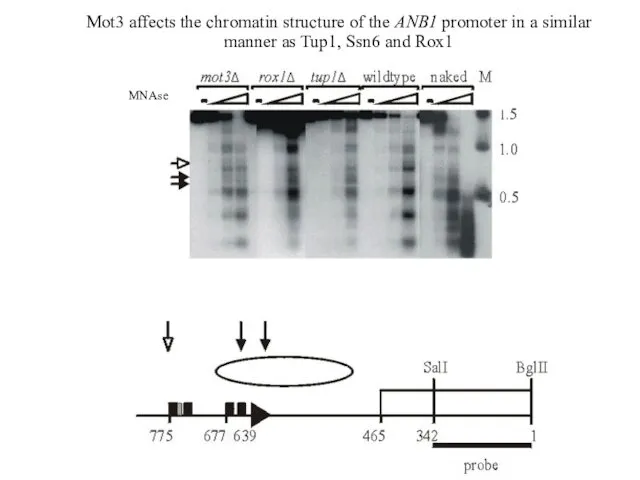
Mot3 affects the chromatin structure of the ANB1 promoter in a
Mot3 affects the chromatin structure of the ANB1 promoter in a
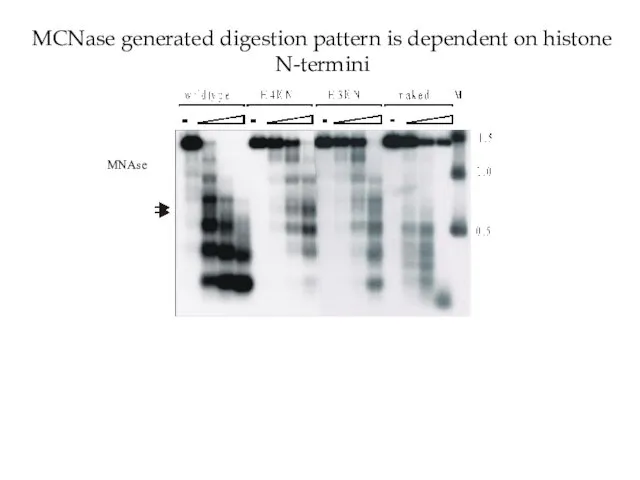
MCNase generated digestion pattern is dependent on histone N-termini
MNAse
MCNase generated digestion pattern is dependent on histone N-termini
MNAse

Summary
Operator efficiency:
- operator orientation and position relative to the TATA box
Summary
Operator efficiency:
- operator orientation and position relative to the TATA box

A Model Fungal Gene Regulatory Mechanism: The GAL genes of Saccharomyces
A Model Fungal Gene Regulatory Mechanism: The GAL genes of Saccharomyces

GAL mutant phenotypes:
GAL1, GAL7, GAL10, MEL1, (GAL5): If mutant, cells cannot
GAL mutant phenotypes:
GAL1, GAL7, GAL10, MEL1, (GAL5): If mutant, cells cannot
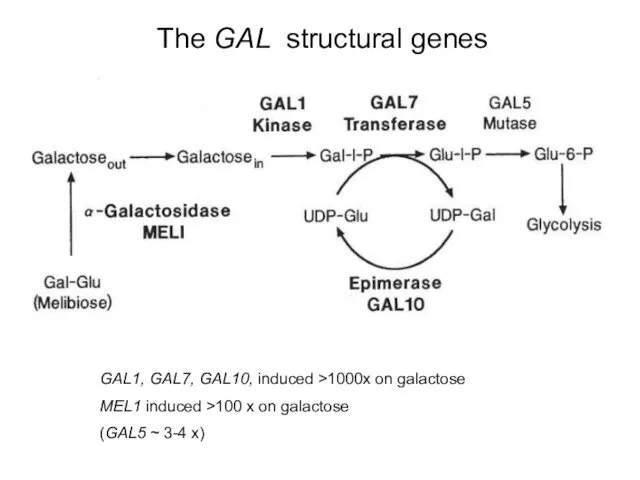
The GAL structural genes
GAL1, GAL7, GAL10, induced >1000x on galactose
MEL1 induced
The GAL structural genes
GAL1, GAL7, GAL10, induced >1000x on galactose
MEL1 induced
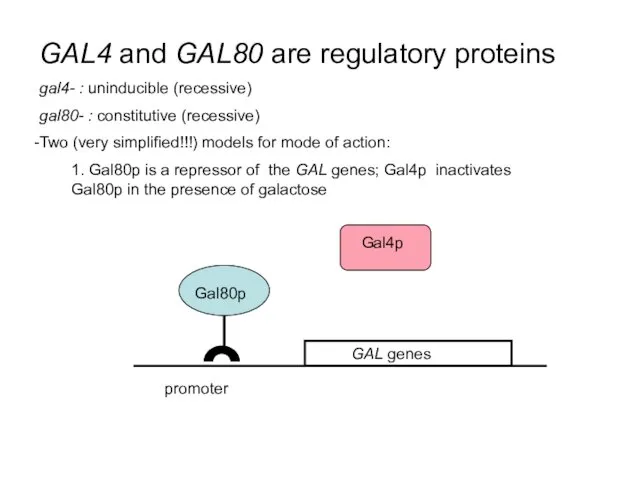
GAL4 and GAL80 are regulatory proteins
gal4- : uninducible (recessive)
gal80- : constitutive
GAL4 and GAL80 are regulatory proteins
gal4- : uninducible (recessive)
gal80- : constitutive
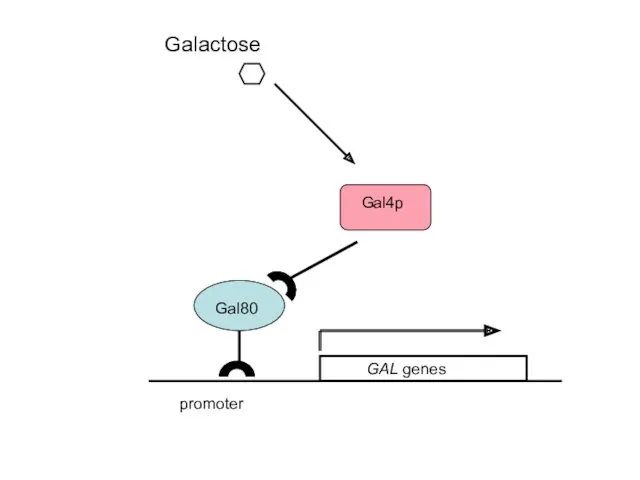
promoter
Gal4p
promoter
Gal4p
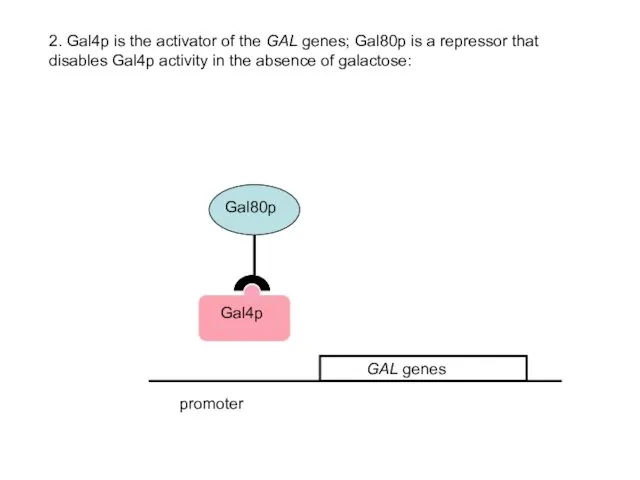
promoter
2. Gal4p is the activator of the GAL genes; Gal80p is
promoter
2. Gal4p is the activator of the GAL genes; Gal80p is
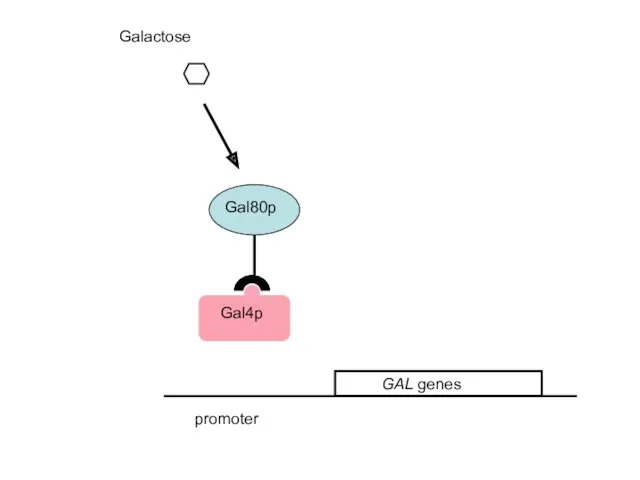
Galactose
promoter
Galactose
promoter
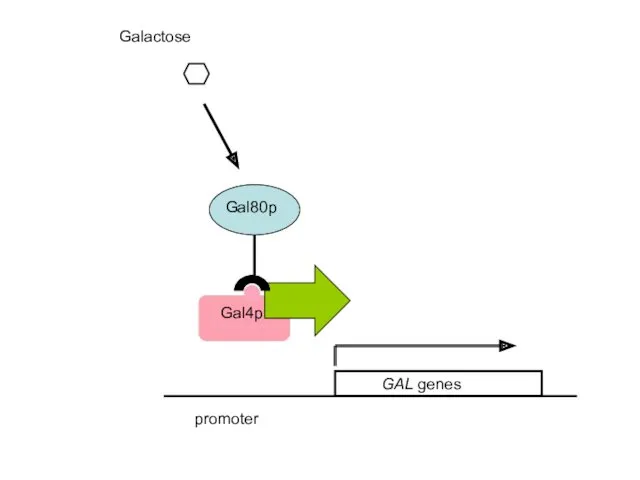
promoter
Galactose
promoter
Galactose
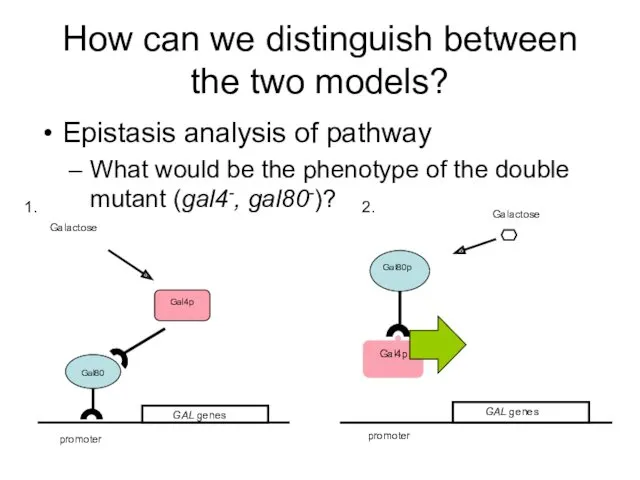
How can we distinguish between the two models?
Epistasis analysis of pathway
What
How can we distinguish between the two models?
Epistasis analysis of pathway
What
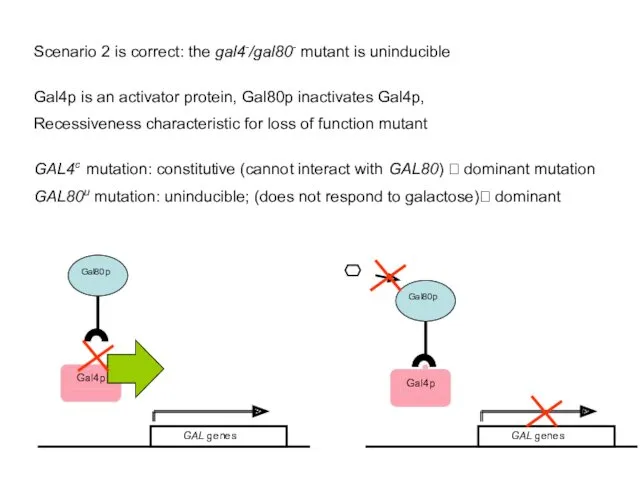
Scenario 2 is correct: the gal4-/gal80- mutant is uninducible
Gal4p is an
Scenario 2 is correct: the gal4-/gal80- mutant is uninducible
Gal4p is an

Cloning of the genes
gal4- uninducible, cannot grow on plates with galactose
Cloning of the genes
gal4- uninducible, cannot grow on plates with galactose
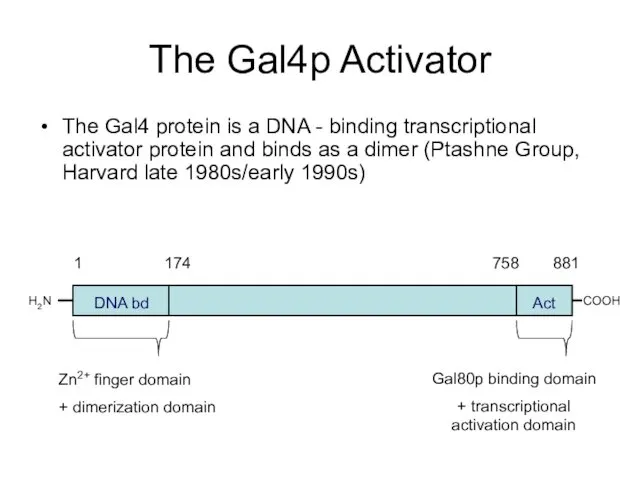
The Gal4p Activator
The Gal4 protein is a DNA - binding transcriptional
The Gal4p Activator
The Gal4 protein is a DNA - binding transcriptional
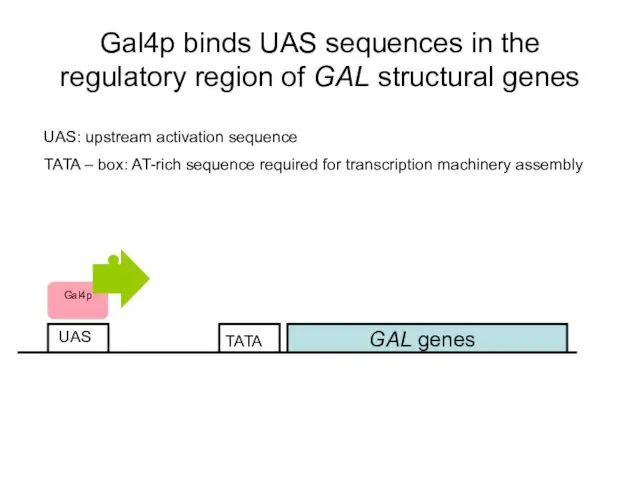
Gal4p binds UAS sequences in the regulatory region of GAL structural
Gal4p binds UAS sequences in the regulatory region of GAL structural
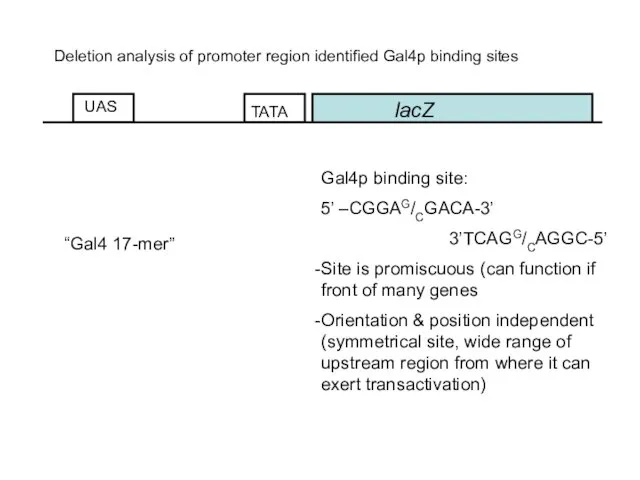
lacZ
UAS
Deletion analysis of promoter region identified Gal4p binding sites
Gal4p binding site:
5’
lacZ
UAS
Deletion analysis of promoter region identified Gal4p binding sites
Gal4p binding site:
5’
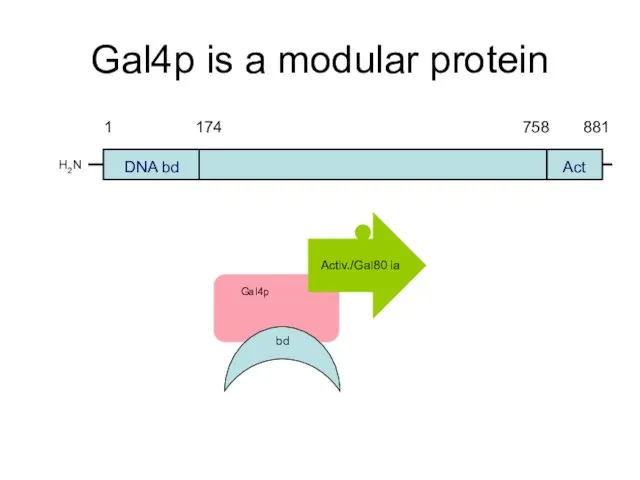
Gal4p is a modular protein
H2N
DNA bd
Act
Activ./Gal80 ia
bd
Gal4p is a modular protein
H2N
DNA bd
Act
Activ./Gal80 ia
bd
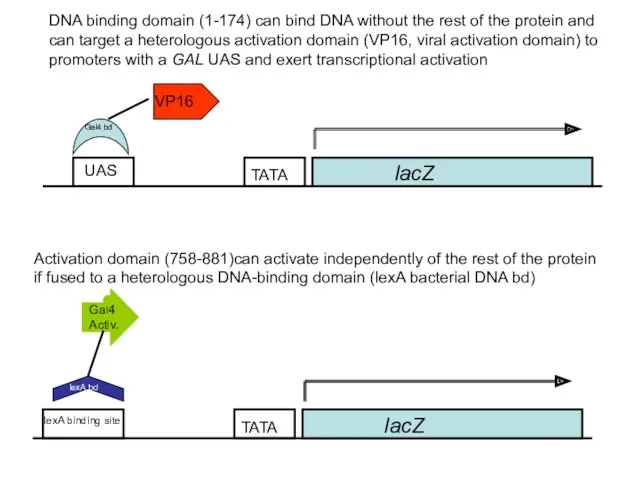
lacZ
UAS
VP16
Activation domain (758-881)can activate independently of the rest of the protein
lacZ
UAS
VP16
Activation domain (758-881)can activate independently of the rest of the protein
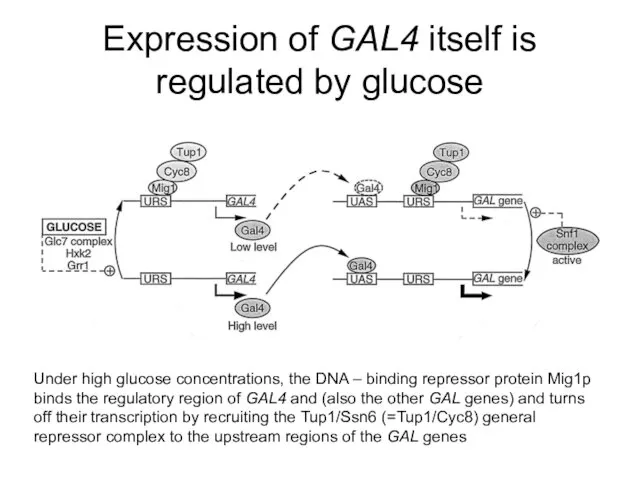
Expression of GAL4 itself is regulated by glucose
Under high glucose concentrations,
Expression of GAL4 itself is regulated by glucose
Under high glucose concentrations,

The galactose sensor: Gal3p
Gal3p is a protein with high similarity (homology)
The galactose sensor: Gal3p
Gal3p is a protein with high similarity (homology)
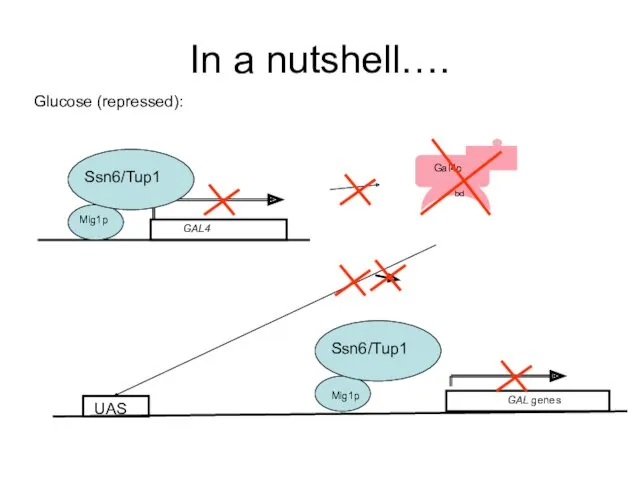
In a nutshell….
Glucose (repressed):
Mig1p
Ssn6/Tup1
GAL genes
Mig1p
Ssn6/Tup1
UAS
In a nutshell….
Glucose (repressed):
Mig1p
Ssn6/Tup1
GAL genes
Mig1p
Ssn6/Tup1
UAS
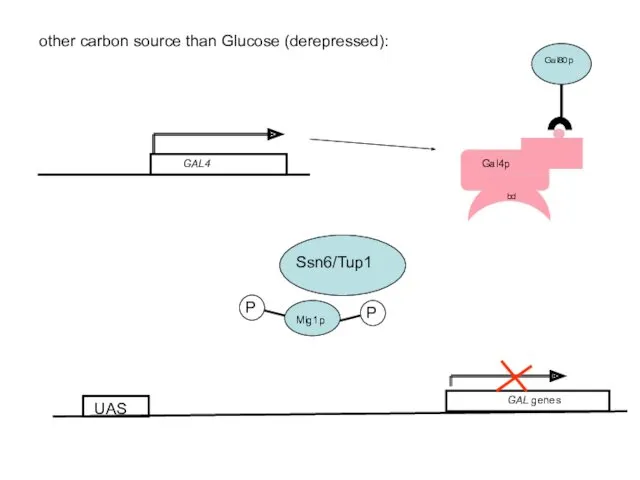
other carbon source than Glucose (derepressed):
GAL genes
UAS
other carbon source than Glucose (derepressed):
GAL genes
UAS
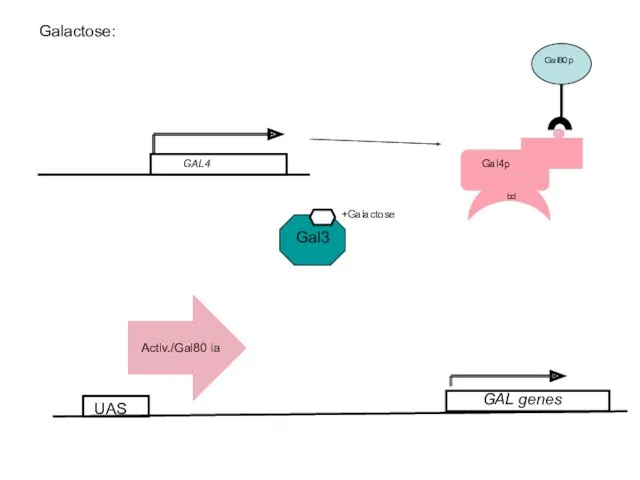
Galactose:
UAS
+Galactose
GAL genes
Galactose:
UAS
+Galactose
GAL genes
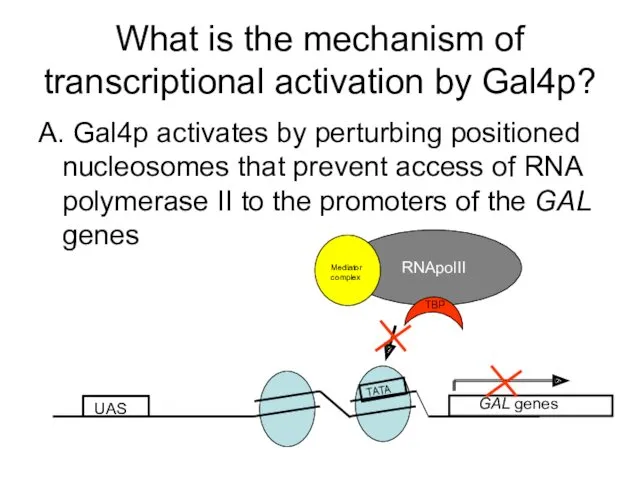
What is the mechanism of transcriptional activation by Gal4p?
A. Gal4p activates
What is the mechanism of transcriptional activation by Gal4p?
A. Gal4p activates
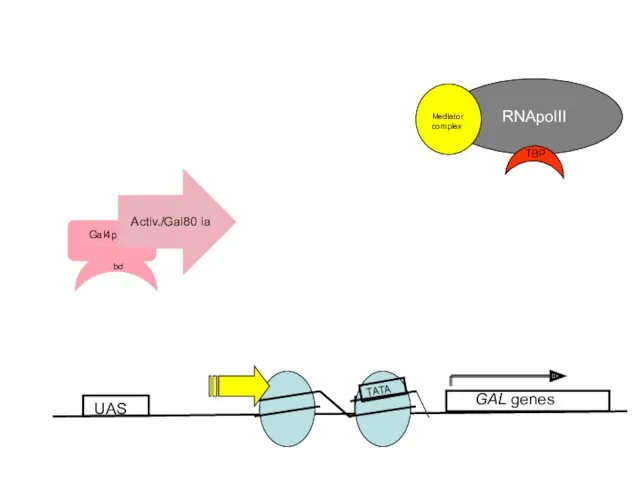
UAS
GAL genes
TATA
UAS
GAL genes
TATA
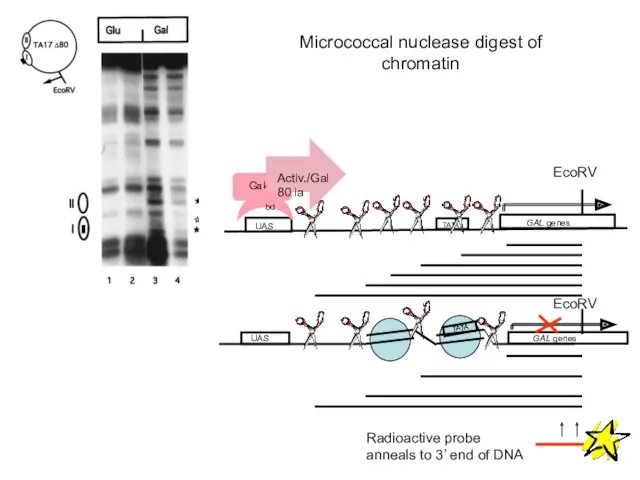
Micrococcal nuclease digest of chromatin
UAS
GAL genes
TATA
UAS
GAL genes
Radioactive probe anneals to 3’
Micrococcal nuclease digest of chromatin
UAS
GAL genes
TATA
UAS
GAL genes
Radioactive probe anneals to 3’
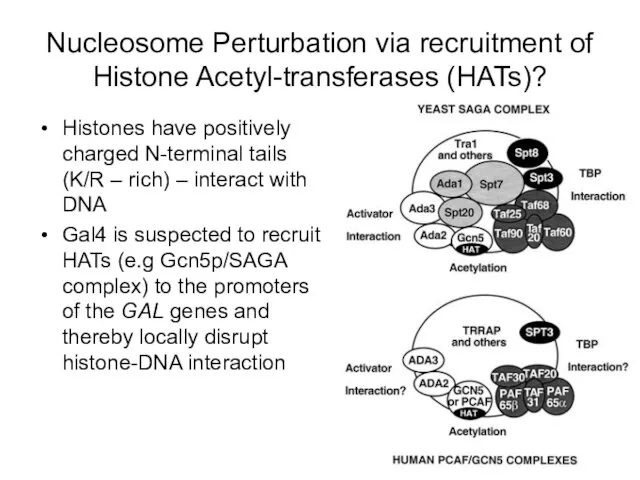
Nucleosome Perturbation via recruitment of Histone Acetyl-transferases (HATs)?
Histones have positively charged
Nucleosome Perturbation via recruitment of Histone Acetyl-transferases (HATs)?
Histones have positively charged
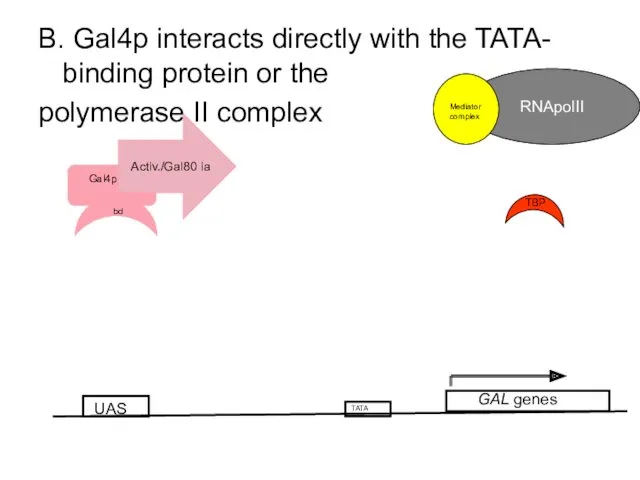
B. Gal4p interacts directly with the TATA- binding protein or the
B. Gal4p interacts directly with the TATA- binding protein or the

Relevance of the Gal regulation research today?
General understanding of basic molecular
Relevance of the Gal regulation research today?
General understanding of basic molecular
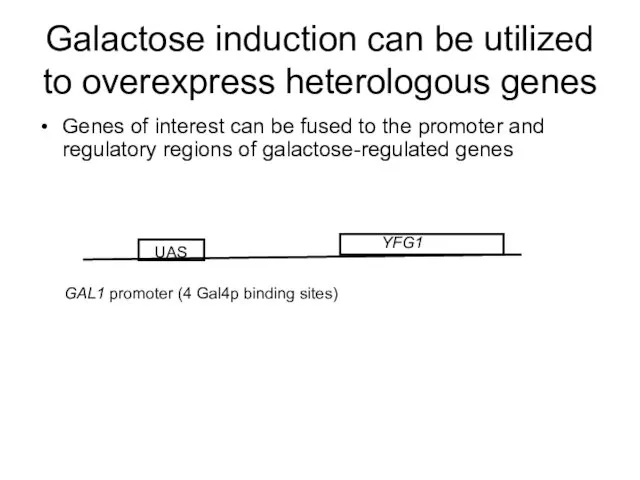
Galactose induction can be utilized to overexpress heterologous genes
Genes of interest
Galactose induction can be utilized to overexpress heterologous genes
Genes of interest

Three expression levels:
Repressed (2% glucose) ? no expression
Derepressed (2% Raffinose, 3%
Three expression levels:
Repressed (2% glucose) ? no expression
Derepressed (2% Raffinose, 3%

Similar: Oleate induction:
Oleate induced genes are involved in peroxisomal proliferation and
Similar: Oleate induction:
Oleate induced genes are involved in peroxisomal proliferation and
 Развитие фонематического слуха у детей с диагнозом: ТНР. (Методика по развитию фонематического слуха у детей с тяжёлым нарушением речи в детском саду.)
Развитие фонематического слуха у детей с диагнозом: ТНР. (Методика по развитию фонематического слуха у детей с тяжёлым нарушением речи в детском саду.) Organization moments first
Organization moments first Сборка ПК для работы с 3D графикой
Сборка ПК для работы с 3D графикой Реакция организма человека на физическое или психологическое воздействие, стресс
Реакция организма человека на физическое или психологическое воздействие, стресс конкурсная работа Я-учитель здоровья
конкурсная работа Я-учитель здоровья Мышление и деятельность. Потребности и интересы. Свобода и ответственность
Мышление и деятельность. Потребности и интересы. Свобода и ответственность Проект Волшебство своими руками
Проект Волшебство своими руками ЧЕТЫРЕ ЦВЕТА СВОБОДЫ: ограничения в жизни детей
ЧЕТЫРЕ ЦВЕТА СВОБОДЫ: ограничения в жизни детей Глубоководные экосистемы (экватор и южные широты)
Глубоководные экосистемы (экватор и южные широты) Кислород
Кислород Виды орнаментов (1 класс)
Виды орнаментов (1 класс) Проектная деятельность по теме: Вязание спицами
Проектная деятельность по теме: Вязание спицами Учебная практика. Общественные отношения, подпадающие под воздействие норм публичного и частного права
Учебная практика. Общественные отношения, подпадающие под воздействие норм публичного и частного права Тесты по органической химии
Тесты по органической химии Язык и речь. Типы речевых ситуаций
Язык и речь. Типы речевых ситуаций Оптика. Геометрическая оптика
Оптика. Геометрическая оптика Инженерные сети на строительной площадке
Инженерные сети на строительной площадке Цифрова система комутації EWSD
Цифрова система комутації EWSD Факторы риска, эпидемиология и профилактика важнейших неинфекционных болезней и их медико-социальные аспекты (БСК, ЗНО, БОД)
Факторы риска, эпидемиология и профилактика важнейших неинфекционных болезней и их медико-социальные аспекты (БСК, ЗНО, БОД) Продуктивное чтение - залог успешного обучения Диск
Продуктивное чтение - залог успешного обучения Диск Рождество Христово
Рождество Христово Теория легирования. Лекция 8
Теория легирования. Лекция 8 Основні вимоги до виробничого освітлення
Основні вимоги до виробничого освітлення Ответственность субъектов предпринимательской деятельности
Ответственность субъектов предпринимательской деятельности Минералы Урала
Минералы Урала Методическое и техническое обеспечение учебного процесса по информатике
Методическое и техническое обеспечение учебного процесса по информатике ВКР: Психологическое сопровождение семьи и школы в процессе профессионального ориентирования подростков
ВКР: Психологическое сопровождение семьи и школы в процессе профессионального ориентирования подростков КОММЕРЧЕСКОЕ ПРЕДЛОЖЕНИЕ
КОММЕРЧЕСКОЕ ПРЕДЛОЖЕНИЕ