- Углеводы. Лекция №10

Содержание
- 2. Содержание: 1.Пути обмена Гл-6-ф 2. Пентозный цикл ( ПФП) 3. Глюконеогенез ( ГНГ ) 4.Биосинтез глюкозаминогликанов
- 3. Пути метаболизма глюкозы С6Н12О6 + инсулиновый стимул Глюкозо 6 фосфат ПВК лактат ГНГ Гликоген, резерв ПФП
- 4. Общая схема ПФП содержит окислительную и неокислительную части
- 7. Пентозофосфатный путь Пентозофосфатный путь можно разделить 2 части: окислительную и неокислительную. В окислительной части, включающей 3
- 8. Пентозный цикл ( окислительная часть)
- 9. Вторая реакция – гидролиз 6-фосфоглюконолактона глюконолактонгидролазой. глюконолактонгидролаза 6-фосфоглюконолактон ------------------------------------? -----------------------? 6-фосфоглюконат -Н2О
- 10. СООН │ Н–С–ОН СН2ОН │ СО2 │ Н–С–ОН С=О │ │ Н–С–ОН 6-фосфоглюконатдегидрогеназа Н–С–ОН │ (декарбоксилирующая)
- 11. Неокислительная часть. В отличие от первой, окислительной, все реакции этой части пентозофосфатного пути обратимы. Рибулозо-5-фосфат может
- 12. В неокислительной части рибулозо-5-фосфат превращается в различные моносахариды с 3, 4, 5, 6, 7 и 8-ю
- 13. Транскетолаза (кофермент – тиаминпирофосфат) отщепляет 2С-фрагмент и переносит его на другие сахара (см. схему). Трансальдолаза способна
- 14. Рибозо-5-фосфат Ксилулозо-5-фосфат Транскетолаза (ТПФ) Седогептулозо-7- фосфат 3-ФГА
- 15. Затем два образовавшиеся соединения реагируют друг с другом в трансальдолазной реакции; при этом в результате переноса
- 16. Седогептулозо-7-ф 3-ФГА Эритрозо-4-ф Фруктозо-6-ф трансальдолаза
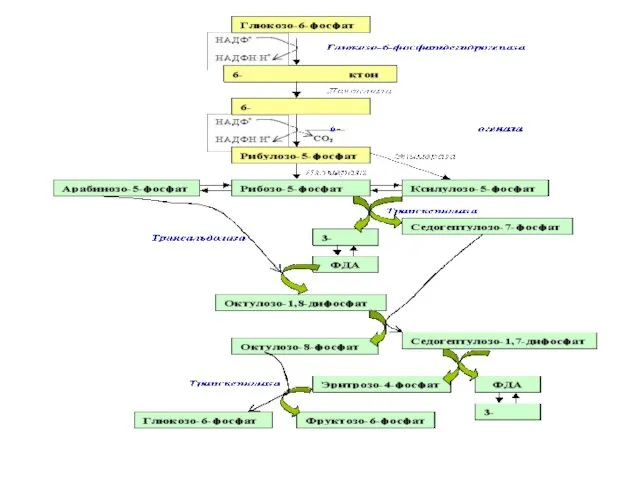
- 17. Однако реакция может идти и по другому пути. В этом случае в трансальдолазной реакции образуется октулозо-1,8-дифосфат.
- 19. Т.к.пентозный цикл протекает в цитоплазме и не может проникать в митохондрии, то он не имеет энергетического
- 20. 1. Биосинтез ХС 2. Синтез аминокислот, гормонов, биогенных аминов 3.Участвует в микросомальном окислении 4. Участвует в
- 21. 7. ПЦ поставляет СО2, который используется в реакциях биосинтеза ЖК, а также для создания щелочного резерва
- 22. 8. ПЦ имеет прямое отношение к механизму электрогенза в нейронах. Активность ПЦ зависит от концентрации Гл-6-ф.
- 23. Биосинтез ГАГ Синтез ГАГ протекает во всех тканях, в том числе и в хрящевой. ГАГ состоят
- 24. Глюкоза-6-ф Фруктозо-6-ф Гл-1-ф УДФ-галактоза УДФ-глюкоза Фруктозамин 6-ф ГЛН ГЛУ Фруктозамин-1-ф N-ацетилфруктозамин-1-ф УДФ-N-ацетилгалактозамин или УДФ-N- глюкозамин УДФ-глюкуроновая
- 25. Глюконеогенез - ГНГ ГНГ снабжает глюкозой прежде всего, конечно, мозг и эритроциты. ГНГ это синтетический процесс,
- 27. ГНГ протекает по общему метаболическому пути гллликолиза, только в обратном направлении: гексокиназа ФФК Гликолиз: Гл Гл-6ф
- 28. пируваткарбоксилаза ФЕП-карбоксикиназа ГНГ: ПВК ЩУК ФЕП ДАФ 2ФГК 3ФГК 1,3 ДФГК 3ФГА фруктозобисфосфатаза гл-6-фосфатаза ф-1,6 диф
- 29. Все реакции гликолиза, кроме гексокиназной, фосфофруктокиназной и пируваткиназной обратимы, поэтому в ГНГ они идут в обратном
- 30. 2 -я реакция ПВК---? ЩУК локализуется в митохондриях. Пируваткарбоксилаза- аллостерический, митохондриальный фермент,активируется ацетил-КоА
- 31. Мембрана митохондрий непроницаема для образовавшейся ЩУК, поэтому она восстанавливается в малат, для которого мембрана проницаема. Это
- 32. Регуляция ГНГ осуществляется теми же факторами,что и гликолиз, но с обратным знаком. Т.е.то, что активирует гликолиз,
- 33. ГНГ ингибируется АДФ, АМФ, Са++. NAD+, Рн, а активируется АТФ, цитратом, ЖК, глицерином, О2, NADH, глюкокортикоидами
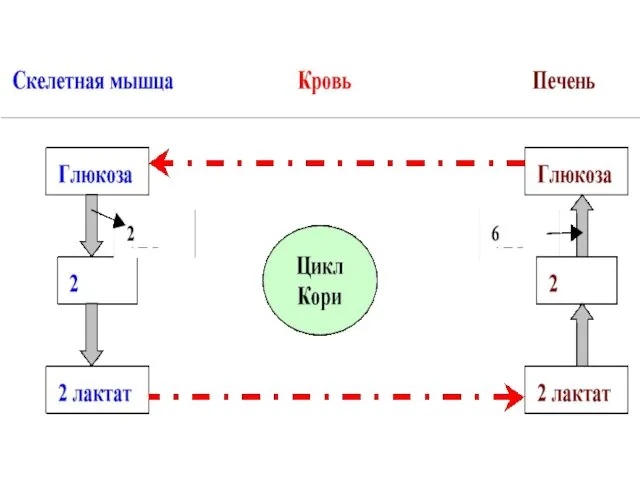
- 34. Между гликолизом интенсивно, протекающим в мышечной ткани при ее интенсивной деятельности, и ГНГ,протекающим в печени, существует
- 35. В печени лактат переходит ----?глюкозу путем ГНГ. Образовавшаяся в печени глюкоза опять с кровотоком потупает в
- 37. Регуляция уровня глюкозы в крови Нормальный уровень глюкозы в крови составляет 3.5-6.1 ммоль/л. Гипогликемия-снижение уровня Гл
- 38. Причины физиологической гипогликемии: 1.Физический труд ( повышенные затраты) 2.Беременность и лактация 3.Голодание
- 39. Причины патологической гипогликемии: 1.Нарушение депонирования Гл в печени 2.Нарушение всасывания углеводов в ЖКТ 3.Нарушение мобилизации гликогена(
- 40. Гиперинсулинизм возникает по двум причинам: а)опухоли ß-клеток островков Лангенгарса б)передозировки инсулина больным диабетом 7.Алкогольная интоксикация(этанол блокатор
- 41. Причины гипергликемии: 1.Переедание углеводов 2.Избыток континсулярных гормонов, которые препятствуют утилизации Гл мышечнй тканью и одновременно стимулируют
- 42. 4. Стрессовые воздействия(↑↑) уровень адреналина 5.Беременность 6.Расстройство мозгового кровобращения 7. Заболевания печени воспалительного или дегенеративного характера.
- 43. Регуляция уровня глюкозы в крови Уровень ГЛ в крови яв-ся обним из гомеостатических параметров. Регуляция ГЛ
- 44. Существуют 2 механизма регуляции: 1.Срочный( через ЦНС) 2.Постоянный ( через гормональное влияние) Срочный механизм срабатывает всегда
- 45. гипоталамус гипофиз СТГ ТТГ АКТГ протеолиз Т3,Т4 липолиз Кортизол. протеолиз
- 46. Он осуществляется по классической схеме:- жертва-хищник. -через зрительный анализатор воспринимается информация об опасности. Возбуждение из одного
- 47. Мобилизция может осуществляться через инозитол-3-фосфатный механизм ( посредством ионов Са++). Срочный механизм поддерживает стабильную гликемию на
- 48. После истощения гликогена, возбужденная кора продолжает посылать импульсы в гипоталамус. Гипоталамус –это гибрид нерной и эндокринной
- 49. Эти гормоны в свою очередь стимулируют выброс Т3, Т4, кортизола и кортизона. Эти же гормоны, в
- 50. Причем для протеолиза расходуются прежде всего дефектные белки, что имеет исключительное значение-гормоны блокируют воспалительные процессы. В
- 51. В условиях длительного воздействия отрицательных факторов на организм( постоянный стресс) может возникнуть дефицит ИНС, что и
- 53. Скачать презентацию

Содержание:
1.Пути обмена Гл-6-ф
2. Пентозный цикл ( ПФП)
3. Глюконеогенез ( ГНГ
Содержание:
1.Пути обмена Гл-6-ф
2. Пентозный цикл ( ПФП)
3. Глюконеогенез ( ГНГ
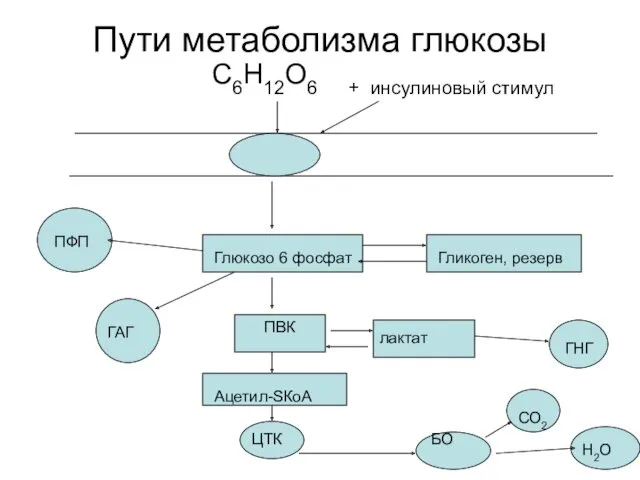
Пути метаболизма глюкозы
С6Н12О6 + инсулиновый стимул
Глюкозо 6 фосфат
ПВК
лактат
ГНГ
Гликоген,
Пути метаболизма глюкозы
С6Н12О6 + инсулиновый стимул
Глюкозо 6 фосфат
ПВК
лактат
ГНГ
Гликоген,

Общая схема ПФП
содержит окислительную и неокислительную части
Общая схема ПФП
содержит окислительную и неокислительную части
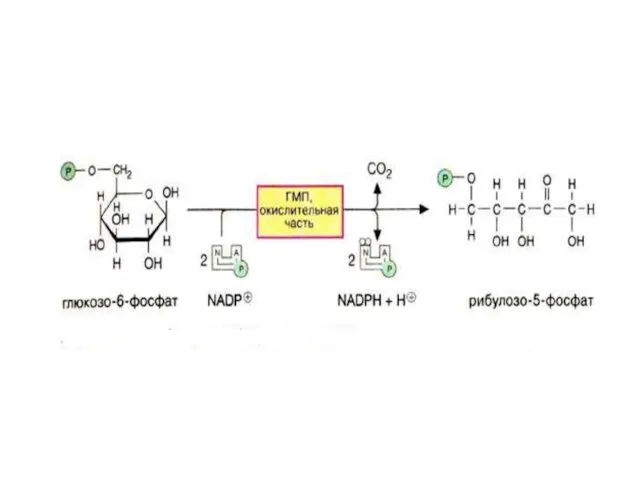
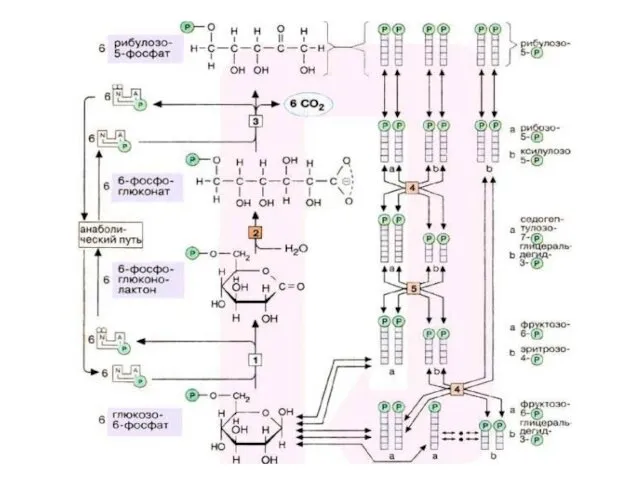

Пентозофосфатный путь
Пентозофосфатный путь можно разделить 2 части: окислительную и неокислительную.
Пентозофосфатный путь
Пентозофосфатный путь можно разделить 2 части: окислительную и неокислительную.
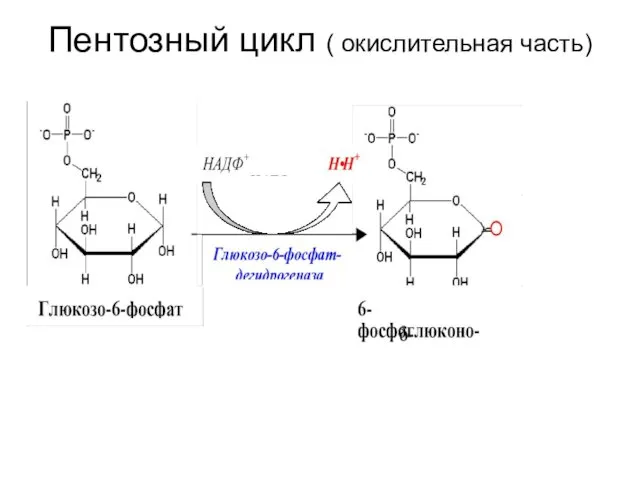
Пентозный цикл ( окислительная часть)
Пентозный цикл ( окислительная часть)

Вторая реакция – гидролиз 6-фосфоглюконолактона глюконолактонгидролазой.
глюконолактонгидролаза
6-фосфоглюконолактон ------------------------------------?
-----------------------? 6-фосфоглюконат
-Н2О
Вторая реакция – гидролиз 6-фосфоглюконолактона глюконолактонгидролазой.
глюконолактонгидролаза
6-фосфоглюконолактон ------------------------------------?
-----------------------? 6-фосфоглюконат
-Н2О
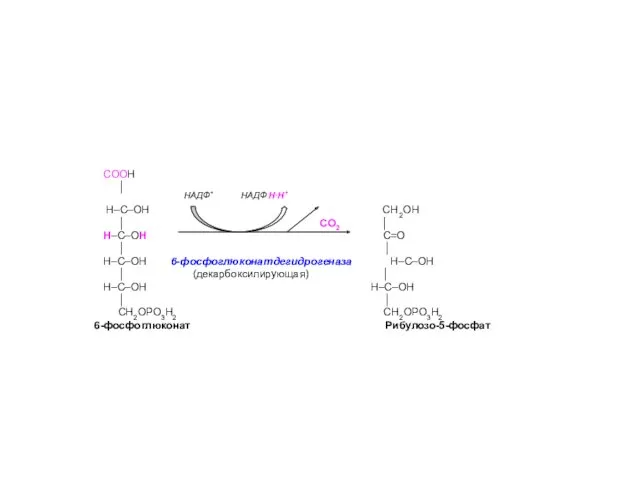
СООН
│
Н–С–ОН СН2ОН
│ СО2 │
Н–С–ОН С=О
СООН
│
Н–С–ОН СН2ОН
│ СО2 │
Н–С–ОН С=О

Неокислительная часть.
В отличие от первой, окислительной, все реакции этой
Неокислительная часть.
В отличие от первой, окислительной, все реакции этой

В неокислительной части рибулозо-5-фосфат превращается в различные моносахариды с 3,
В неокислительной части рибулозо-5-фосфат превращается в различные моносахариды с 3,

Транскетолаза (кофермент – тиаминпирофосфат) отщепляет 2С-фрагмент и переносит его на другие
Транскетолаза (кофермент – тиаминпирофосфат) отщепляет 2С-фрагмент и переносит его на другие
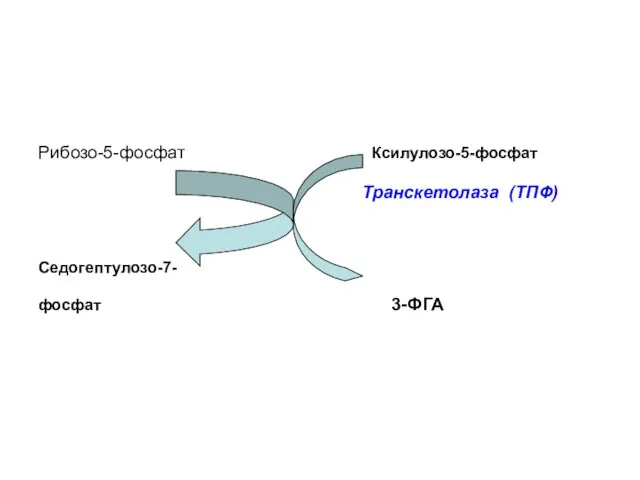
Рибозо-5-фосфат Ксилулозо-5-фосфат
Транскетолаза (ТПФ)
Седогептулозо-7-
фосфат 3-ФГА
Рибозо-5-фосфат Ксилулозо-5-фосфат
Транскетолаза (ТПФ)
Седогептулозо-7-
фосфат 3-ФГА

Затем два образовавшиеся соединения реагируют друг с другом в трансальдолазной реакции;
Затем два образовавшиеся соединения реагируют друг с другом в трансальдолазной реакции;
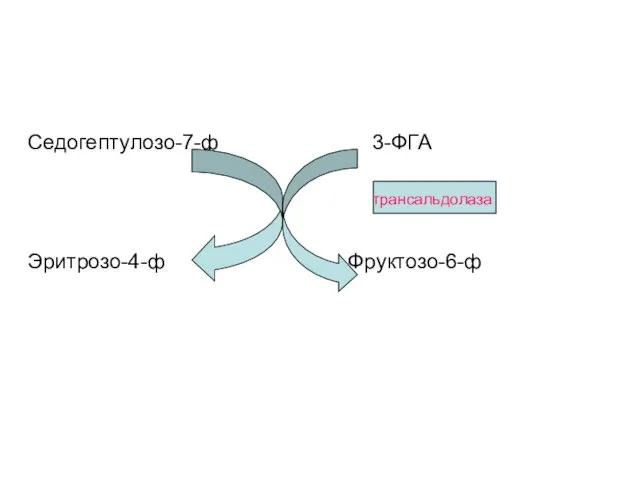
Седогептулозо-7-ф 3-ФГА
Эритрозо-4-ф Фруктозо-6-ф
трансальдолаза
Эритрозо-4-ф Фруктозо-6-ф
трансальдолаза

Однако реакция может идти и по другому пути. В этом случае

Т.к.пентозный цикл протекает в цитоплазме и не может проникать
Т.к.пентозный цикл протекает в цитоплазме и не может проникать

1. Биосинтез ХС
2. Синтез аминокислот, гормонов,
биогенных аминов
3.Участвует в микросомальном окислении
4.
1. Биосинтез ХС
2. Синтез аминокислот, гормонов,
биогенных аминов
3.Участвует в микросомальном окислении
4.

7. ПЦ поставляет СО2, который используется в реакциях биосинтеза ЖК, а
7. ПЦ поставляет СО2, который используется в реакциях биосинтеза ЖК, а

8. ПЦ имеет прямое отношение к механизму электрогенза в нейронах.
8. ПЦ имеет прямое отношение к механизму электрогенза в нейронах.

Биосинтез ГАГ
Синтез ГАГ протекает во всех тканях, в том числе и
Биосинтез ГАГ
Синтез ГАГ протекает во всех тканях, в том числе и
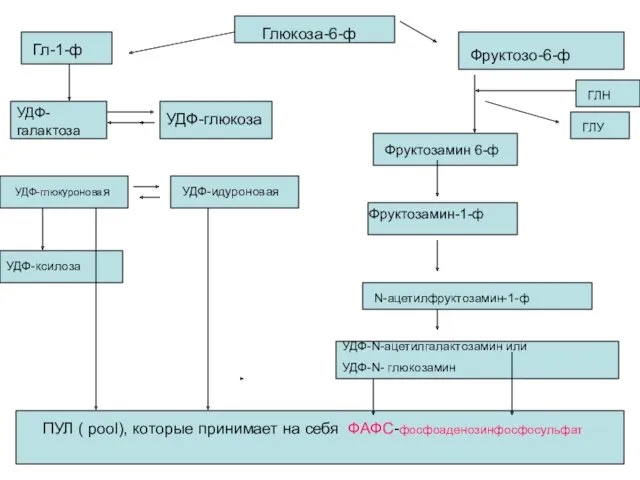
Глюкоза-6-ф
Фруктозо-6-ф
Гл-1-ф
УДФ-галактоза
УДФ-глюкоза
Фруктозамин 6-ф
ГЛН
ГЛУ
Фруктозамин-1-ф
N-ацетилфруктозамин-1-ф
УДФ-N-ацетилгалактозамин или
УДФ-N- глюкозамин
УДФ-глюкуроновая
УДФ-идуроновая
УДФ-ксилоза
ПУЛ ( pool), которые
Глюкоза-6-ф
Фруктозо-6-ф
Гл-1-ф
УДФ-галактоза
УДФ-глюкоза
Фруктозамин 6-ф
ГЛН
ГЛУ
Фруктозамин-1-ф
N-ацетилфруктозамин-1-ф
УДФ-N-ацетилгалактозамин или
УДФ-N- глюкозамин
УДФ-глюкуроновая
УДФ-идуроновая
УДФ-ксилоза
ПУЛ ( pool), которые

Глюконеогенез - ГНГ
ГНГ снабжает глюкозой прежде всего, конечно, мозг и эритроциты.
ГНГ
Глюконеогенез - ГНГ
ГНГ снабжает глюкозой прежде всего, конечно, мозг и эритроциты.
ГНГ
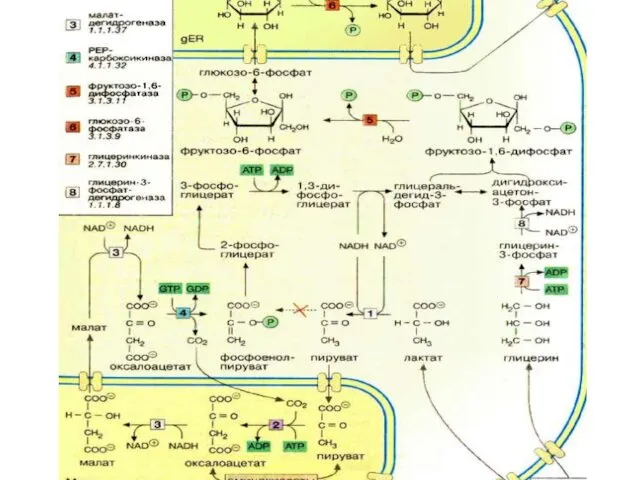

ГНГ протекает по общему метаболическому пути гллликолиза, только в обратном
ГНГ протекает по общему метаболическому пути гллликолиза, только в обратном
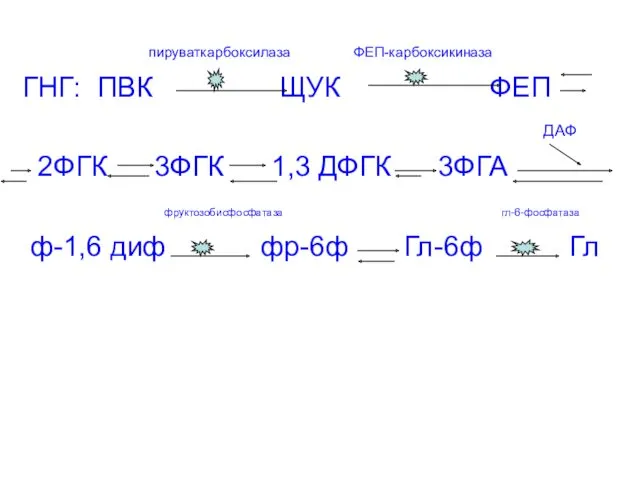
пируваткарбоксилаза ФЕП-карбоксикиназа
ГНГ: ПВК ЩУК ФЕП
ДАФ
2ФГК 3ФГК 1,3 ДФГК
пируваткарбоксилаза ФЕП-карбоксикиназа
ГНГ: ПВК ЩУК ФЕП
ДАФ
2ФГК 3ФГК 1,3 ДФГК

Все реакции гликолиза, кроме гексокиназной, фосфофруктокиназной и пируваткиназной обратимы, поэтому
Все реакции гликолиза, кроме гексокиназной, фосфофруктокиназной и пируваткиназной обратимы, поэтому
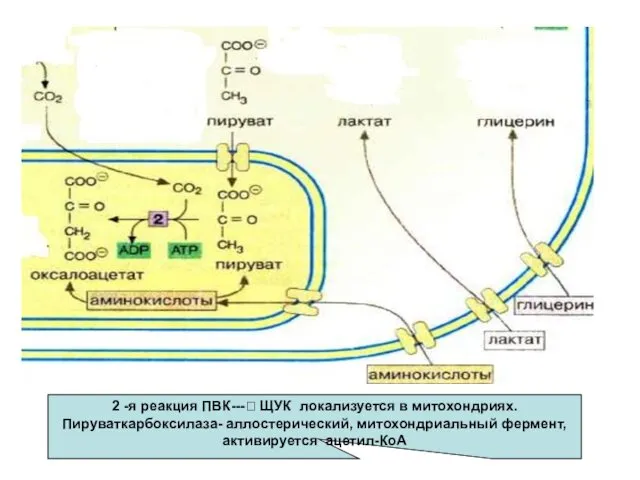
2 -я реакция ПВК---? ЩУК локализуется в митохондриях. Пируваткарбоксилаза- аллостерический, митохондриальный
2 -я реакция ПВК---? ЩУК локализуется в митохондриях. Пируваткарбоксилаза- аллостерический, митохондриальный
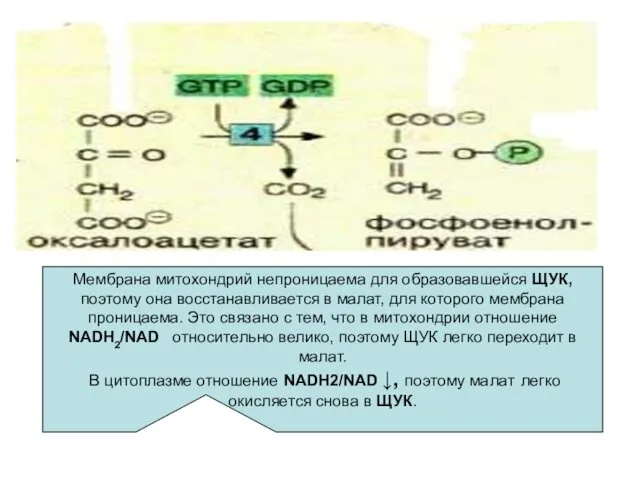
Мембрана митохондрий непроницаема для образовавшейся ЩУК, поэтому она восстанавливается в малат,
Мембрана митохондрий непроницаема для образовавшейся ЩУК, поэтому она восстанавливается в малат,

Регуляция ГНГ осуществляется теми же факторами,что и гликолиз, но
Регуляция ГНГ осуществляется теми же факторами,что и гликолиз, но

ГНГ ингибируется АДФ, АМФ, Са++. NAD+, Рн, а
активируется
ГНГ ингибируется АДФ, АМФ, Са++. NAD+, Рн, а
активируется

Между гликолизом интенсивно, протекающим в мышечной ткани при ее интенсивной
Между гликолизом интенсивно, протекающим в мышечной ткани при ее интенсивной

В печени лактат переходит ----?глюкозу путем ГНГ. Образовавшаяся в печени
В печени лактат переходит ----?глюкозу путем ГНГ. Образовавшаяся в печени

Регуляция уровня глюкозы в крови
Нормальный уровень глюкозы в крови составляет 3.5-6.1
Регуляция уровня глюкозы в крови
Нормальный уровень глюкозы в крови составляет 3.5-6.1

Причины физиологической гипогликемии:
1.Физический труд ( повышенные затраты)
2.Беременность и лактация
3.Голодание
1.Физический труд ( повышенные затраты)
2.Беременность и лактация
3.Голодание

Причины патологической гипогликемии:
1.Нарушение депонирования Гл в печени
2.Нарушение всасывания углеводов в ЖКТ
3.Нарушение
Причины патологической гипогликемии:
1.Нарушение депонирования Гл в печени
2.Нарушение всасывания углеводов в ЖКТ
3.Нарушение

Гиперинсулинизм возникает по двум причинам:
а)опухоли ß-клеток островков Лангенгарса
б)передозировки инсулина больным диабетом
7.Алкогольная
Гиперинсулинизм возникает по двум причинам:
а)опухоли ß-клеток островков Лангенгарса
б)передозировки инсулина больным диабетом
7.Алкогольная

Причины гипергликемии:
1.Переедание углеводов
2.Избыток континсулярных гормонов, которые препятствуют утилизации Гл мышечнй
Причины гипергликемии:
1.Переедание углеводов
2.Избыток континсулярных гормонов, которые препятствуют утилизации Гл мышечнй

4. Стрессовые воздействия(↑↑) уровень адреналина
5.Беременность
6.Расстройство мозгового кровобращения
7. Заболевания печени воспалительного или
5.Беременность
6.Расстройство мозгового кровобращения
7. Заболевания печени воспалительного или

Регуляция уровня глюкозы в крови
Уровень ГЛ в крови яв-ся обним
Регуляция уровня глюкозы в крови
Уровень ГЛ в крови яв-ся обним

Существуют 2 механизма регуляции:
1.Срочный( через ЦНС)
2.Постоянный ( через гормональное влияние)
Существуют 2 механизма регуляции:
1.Срочный( через ЦНС)
2.Постоянный ( через гормональное влияние)
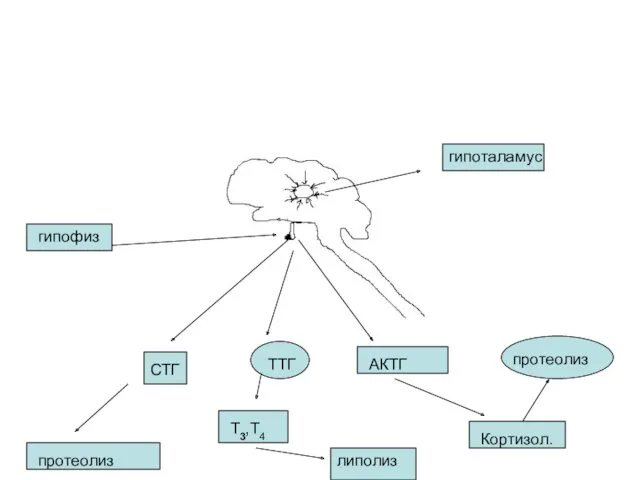
гипоталамус
гипофиз
СТГ
ТТГ
АКТГ
протеолиз
Т3,Т4
липолиз
Кортизол.
протеолиз
гипоталамус
гипофиз
СТГ
ТТГ
АКТГ
протеолиз
Т3,Т4
липолиз
Кортизол.
протеолиз

Он осуществляется по классической схеме:- жертва-хищник.
-через зрительный анализатор воспринимается
Он осуществляется по классической схеме:- жертва-хищник.
-через зрительный анализатор воспринимается

Мобилизция может осуществляться через инозитол-3-фосфатный механизм
( посредством ионов Са++).
Мобилизция может осуществляться через инозитол-3-фосфатный механизм
( посредством ионов Са++).

После истощения гликогена, возбужденная кора продолжает посылать импульсы в гипоталамус.
После истощения гликогена, возбужденная кора продолжает посылать импульсы в гипоталамус.

Эти гормоны в свою очередь стимулируют выброс Т3, Т4, кортизола
Эти гормоны в свою очередь стимулируют выброс Т3, Т4, кортизола

Причем для протеолиза расходуются прежде всего дефектные белки, что имеет
Причем для протеолиза расходуются прежде всего дефектные белки, что имеет

В условиях длительного воздействия отрицательных факторов на организм( постоянный стресс)
В условиях длительного воздействия отрицательных факторов на организм( постоянный стресс)
 Difference between a few and few
Difference between a few and few Организационный классный час. Начало 4 четверти
Организационный классный час. Начало 4 четверти Вещий сон: миф или реальность
Вещий сон: миф или реальность Проекты-презентации
Проекты-презентации Защитное вооружение русской дружины
Защитное вооружение русской дружины Mobile Phone
Mobile Phone Соборы Московского Кремля
Соборы Московского Кремля Микроконтроллеры PIC16R8x
Микроконтроллеры PIC16R8x Автомобильная промышленность Японии
Автомобильная промышленность Японии Современные образовательные технологии
Современные образовательные технологии История возникновения образа Золотой век Голливуда 40-х годов
История возникновения образа Золотой век Голливуда 40-х годов Аналитическая геометрия. Линии на плоскости и их уравнения
Аналитическая геометрия. Линии на плоскости и их уравнения 20231015_otkrytyy_urok_tsifry_anlanticheskiy_poligon
20231015_otkrytyy_urok_tsifry_anlanticheskiy_poligon 1С Парфюм Логистик
1С Парфюм Логистик Использование логических устройств в вычислительной технике
Использование логических устройств в вычислительной технике Обратные задачи вероятностных расчётов конструкций
Обратные задачи вероятностных расчётов конструкций Проект SDR приемника
Проект SDR приемника Комплексный подход к системе оценки результатов образования согласно ФГОС
Комплексный подход к системе оценки результатов образования согласно ФГОС Рождество Христово
Рождество Христово Модуль действительного числа
Модуль действительного числа Люди и роботы. Насколько они похожи
Люди и роботы. Насколько они похожи Тибетский тест личности
Тибетский тест личности Industry of fish and fish products
Industry of fish and fish products загадки о профессиях
загадки о профессиях Разработка бизнес-плана дизайн-агентства RedGrаy
Разработка бизнес-плана дизайн-агентства RedGrаy Урок-рефлексии по теме Соли
Урок-рефлексии по теме Соли Поурочные планы, внеклассные мероприятия
Поурочные планы, внеклассные мероприятия Созидательное и разрушительное общение. Конфликты. Модели поведения в конфликте
Созидательное и разрушительное общение. Конфликты. Модели поведения в конфликте