- Высшая нервная деятельность. Физиология сна. Лекция № 30

Содержание
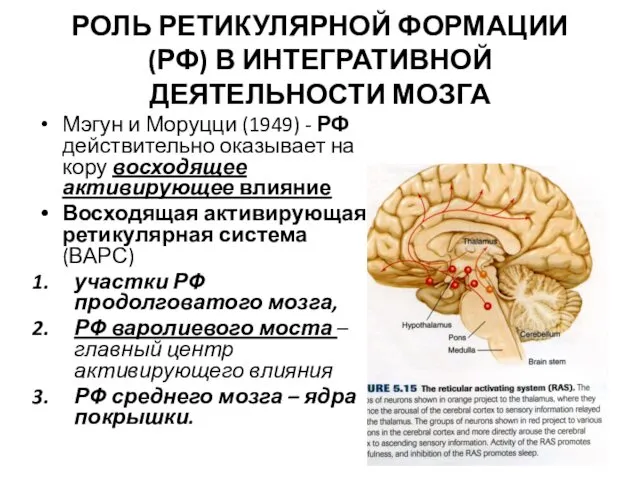
- 2. РОЛЬ РЕТИКУЛЯРНОЙ ФОРМАЦИИ (РФ) В ИНТЕГРАТИВНОЙ ДЕЯТЕЛЬНОСТИ МОЗГА Мэгун и Моруцци (1949) - РФ действительно оказывает
- 3. Неспецифическая проекционная таламо-кортикальная система составляющая неспецифическую проекционную таламо-кортикальную систему (НПТКС). Мэгун и Моруцци предположили, что одним
- 4. СОН Благослови, господь, изобретшего сон. М. Сервантес де Сааведра. "Дон Кихот" Сон — это чудо матери
- 5. Сон - активное физиологическое состояние, которое характеризуется потерей активных психических связей субъекта с окружающим его миром.
- 6. Выделяют несколько видов сна: физиологический периодический ежесуточный сон нефизиологический сон: - наркотический сон, вызываемый различными химическими
- 7. СТРУКТУРА СНА "Медленный" сон: 1. Стадия тета-волн. 2. Стадия «сонных веретен» и «К-комплексов». 3. Стадия дельта-волн
- 8. "Медленный" сон: Очень поверхностный сон. Высокоамплитудные медленные тета-волны (3-7 Гц). Поверхностный сон. Веретенообразные всплески бета-ритма (сонные
- 9. СТАДИИ СНА 1 стадия 2 стадия БДГ сон 4 стадия Стадии медленного сна
- 10. "Быстрый" сон (БДГ- сон) Основной отличительной особенностью "быстрого" сна является наличие быстрых движений глазных яблок (фаза
- 11. ТЕОРИИ СНА Переход от бодрствования ко сну предполагает два возможных пути: Пассивный, в результате утомления ЦНС
- 12. 1. Гипотеза гипнотоксинов или гуморальная теория Лежандр и Пьерон (1910) Сон обусловлен накоплением в крови особых
- 13. Традиционные обсуждаемые нейрохимические агенты, имеющие значение в организации цикла "сон- бодрствование": Фаза медленного сна: ГАМК, серотонин,
- 14. Однако многие факты и, в частности, то, что можно преодолеть сон интересным занятием или то, что
- 15. 2. Теория центров сна Фон Экономо (1930); Гесс (1933) передний гипоталамус
- 16. Однако в дальнейшем было показано, что состояние сна можно было получить при электростимуляции в определенном ритме
- 17. 3. Теория условного торможения И.П. Павлов даже сделал вывод о том, что условное торможение и сон
- 18. 4. Теории деафферентации сенсорных систем Бремер, конец 1930-х годов Для бодрствования необходим хотя бы минимальный уровень
- 19. Однако даже в изолированном переднем мозге появляются ритмичные колебания, характерные для цикла сна/бодрствования. Кроме того, лишение
- 20. 5. Теория неспецифических регуляторов сна Активирующие структуры: ВАРС Неспецифическая и специфическая таламическая система Ядра заднего гипоталамуса
- 21. Однако в ретикулярной формации существуют не только центры бодрствования, но и сна. Каудальные отделы оказывают тормозное
- 22. 6.Серотонинергическая теория М. Жуве - конец 60-х годов В верхних отделах ствола мозга есть две области
- 24. Скачать презентацию
РОЛЬ РЕТИКУЛЯРНОЙ ФОРМАЦИИ (РФ) В ИНТЕГРАТИВНОЙ ДЕЯТЕЛЬНОСТИ МОЗГА
Мэгун и Моруцци (1949)
РОЛЬ РЕТИКУЛЯРНОЙ ФОРМАЦИИ (РФ) В ИНТЕГРАТИВНОЙ ДЕЯТЕЛЬНОСТИ МОЗГА
Мэгун и Моруцци (1949)
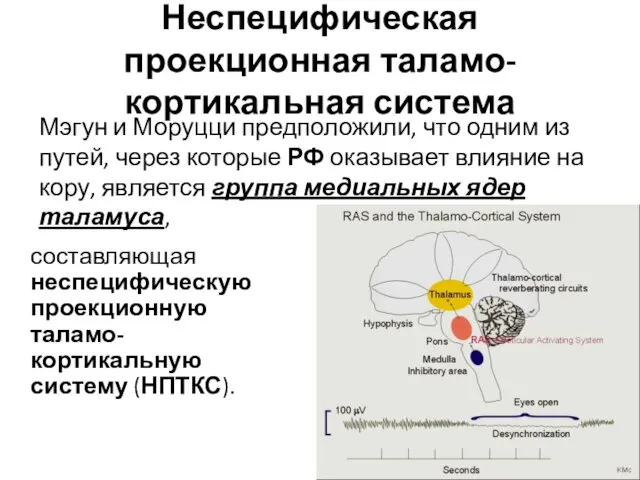
Неспецифическая проекционная таламо-кортикальная система
составляющая неспецифическую проекционную таламо-кортикальную систему (НПТКС).
Мэгун и
Неспецифическая проекционная таламо-кортикальная система
составляющая неспецифическую проекционную таламо-кортикальную систему (НПТКС).
Мэгун и

СОН
Благослови, господь, изобретшего сон.
М. Сервантес де Сааведра. "Дон Кихот"
Сон — это
СОН
Благослови, господь, изобретшего сон.
М. Сервантес де Сааведра. "Дон Кихот"
Сон — это

Сон - активное физиологическое состояние, которое характеризуется потерей активных психических связей
Сон - активное физиологическое состояние, которое характеризуется потерей активных психических связей

Выделяют несколько видов сна:
физиологический периодический ежесуточный сон
нефизиологический сон:
- наркотический сон, вызываемый
Выделяют несколько видов сна:
физиологический периодический ежесуточный сон
нефизиологический сон:
- наркотический сон, вызываемый

СТРУКТУРА СНА
"Медленный" сон:
1. Стадия тета-волн.
2. Стадия «сонных веретен» и «К-комплексов».
3. Стадия
СТРУКТУРА СНА
"Медленный" сон:
1. Стадия тета-волн.
2. Стадия «сонных веретен» и «К-комплексов».
3. Стадия

"Медленный" сон:
Очень поверхностный сон. Высокоамплитудные медленные тета-волны (3-7 Гц).
Поверхностный сон.
"Медленный" сон:
Очень поверхностный сон. Высокоамплитудные медленные тета-волны (3-7 Гц).
Поверхностный сон.
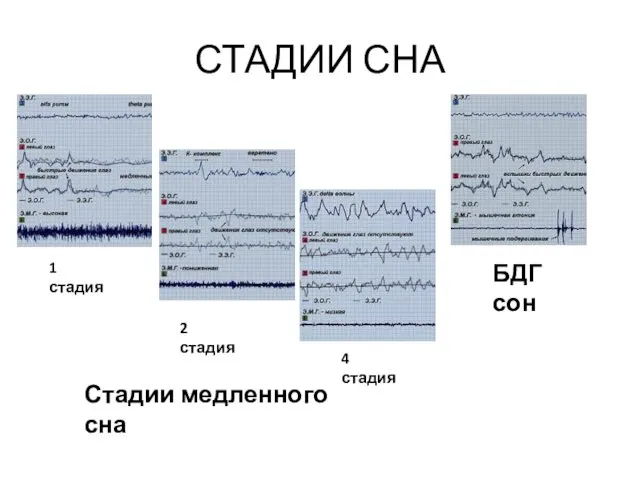
СТАДИИ СНА
1 стадия
2 стадия
БДГ сон
4 стадия
Стадии медленного сна
СТАДИИ СНА
1 стадия
2 стадия
БДГ сон
4 стадия
Стадии медленного сна

"Быстрый" сон (БДГ- сон)
Основной отличительной особенностью "быстрого" сна является наличие быстрых
"Быстрый" сон (БДГ- сон)
Основной отличительной особенностью "быстрого" сна является наличие быстрых

ТЕОРИИ СНА
Переход от бодрствования ко сну предполагает два возможных пути:
Пассивный, в
ТЕОРИИ СНА
Переход от бодрствования ко сну предполагает два возможных пути:
Пассивный, в

1. Гипотеза гипнотоксинов или гуморальная теория
Лежандр и Пьерон (1910)
Сон обусловлен накоплением
1. Гипотеза гипнотоксинов или гуморальная теория
Лежандр и Пьерон (1910)
Сон обусловлен накоплением

Традиционные обсуждаемые нейрохимические агенты, имеющие значение в организации цикла "сон- бодрствование":
Традиционные обсуждаемые нейрохимические агенты, имеющие значение в организации цикла "сон- бодрствование":

Однако
многие факты и, в частности, то, что можно преодолеть сон интересным
Однако
многие факты и, в частности, то, что можно преодолеть сон интересным

2. Теория центров сна
Фон Экономо (1930); Гесс (1933)
передний гипоталамус
2. Теория центров сна
Фон Экономо (1930); Гесс (1933)
передний гипоталамус

Однако
в дальнейшем было показано, что состояние сна можно было получить при
Однако
в дальнейшем было показано, что состояние сна можно было получить при

3. Теория условного торможения
И.П. Павлов даже сделал вывод о том, что
3. Теория условного торможения
И.П. Павлов даже сделал вывод о том, что

4. Теории деафферентации сенсорных систем
Бремер, конец 1930-х годов
Для бодрствования необходим
4. Теории деафферентации сенсорных систем
Бремер, конец 1930-х годов
Для бодрствования необходим

Однако
даже в изолированном переднем мозге появляются ритмичные колебания, характерные для цикла
Однако
даже в изолированном переднем мозге появляются ритмичные колебания, характерные для цикла

5. Теория неспецифических регуляторов сна
Активирующие структуры:
ВАРС
Неспецифическая и специфическая таламическая система
Ядра заднего
5. Теория неспецифических регуляторов сна
Активирующие структуры:
ВАРС
Неспецифическая и специфическая таламическая система
Ядра заднего

Однако
в ретикулярной формации существуют не только центры бодрствования, но и сна.
Однако
в ретикулярной формации существуют не только центры бодрствования, но и сна.

6.Серотонинергическая теория
М. Жуве - конец 60-х годов
В верхних отделах ствола
6.Серотонинергическая теория
М. Жуве - конец 60-х годов
В верхних отделах ствола
 Тренинг на сплочение коллектива
Тренинг на сплочение коллектива Игра - презентация Бабушка - Загадушка
Игра - презентация Бабушка - Загадушка Шаблон Фракталы-22
Шаблон Фракталы-22 Интеграция ФЭМП в разных образовательных областях
Интеграция ФЭМП в разных образовательных областях Презентация к открытому уроку по теме Фриволите
Презентация к открытому уроку по теме Фриволите Витамин С в сокосодержащих продуктах
Витамин С в сокосодержащих продуктах разноцветный детский сад
разноцветный детский сад Классный час Конституция РФ
Классный час Конституция РФ История моей библиотеки
История моей библиотеки Компетентностный подход в дополнительном образовании детей
Компетентностный подход в дополнительном образовании детей Правописание мягкого знака в числительных
Правописание мягкого знака в числительных Урок по географии в 6 классе Озёра. Происхождение озёр. Значение озёр
Урок по географии в 6 классе Озёра. Происхождение озёр. Значение озёр Изготовление двусторонних гибких печатных плат с металлизацией отверстий
Изготовление двусторонних гибких печатных плат с металлизацией отверстий Движение воздушных масс. Атмосферный фронт. Циклоны и антициклоны
Движение воздушных масс. Атмосферный фронт. Циклоны и антициклоны Метод проектов
Метод проектов Створення датчику руху та дослідження його характеристик
Створення датчику руху та дослідження його характеристик Задачи на изменение массы пластинки
Задачи на изменение массы пластинки Байкал
Байкал Arduino. Язык программирования Arduino
Arduino. Язык программирования Arduino Машины для свайных работ
Машины для свайных работ Святыни земли Ровеньской
Святыни земли Ровеньской Управление пользователями. Процессы. Права доступа. Репозитории. Урок 2
Управление пользователями. Процессы. Права доступа. Репозитории. Урок 2 Поражение сердечно-сосудистой системы при антифосфолипидном синдроме
Поражение сердечно-сосудистой системы при антифосфолипидном синдроме Релейная защита и автоматизация ЭЭС
Релейная защита и автоматизация ЭЭС Античная цивилизация, или античный мир
Античная цивилизация, или античный мир Игорь Иванович Сикорский
Игорь Иванович Сикорский Налаштовування параметрів безпеки в середовищі браузера
Налаштовування параметрів безпеки в середовищі браузера Fish Phonics Game
Fish Phonics Game