- Активный транспорт ионов

Содержание
- 2. Ионные токи, протекающие через отдельные каналы Открытие и закрытие ионных каналов отражаются в виде прямоугольных токовых
- 3. Потенциал-зависимые и другие ионные каналы Каналы, образующие водную пору, включают потенциал-зависимые: K+-, Ca2+- и Na+-селективные каналы
- 4. Зачем нужны системы активного транспорта?
- 5. Зачем нужны системы активного транспорта? Для поддержания нормального функционального состояния нейронов в их мембранах локализованы транспортные
- 6. Два механизма активного транспорта Известны два механизма активного транспорта: системы первичного активного транспорта (ионные насосы) с
- 7. Механизмы первичного активного транспорта Системы первичного активного транспорта используют энергию гидролиза АТФ. В настоящее время известно
- 8. Натрий-калиевый насос В начале 1950-х г.г., исследуя действие локальных анестетиков на ионную проводимость нерва краба, Йенс
- 9. Натрий-калиевый насос Позже Р. Кейнс (Richard Keynes) с соавторами, используя радиоактивный 24Na+, впервые непосредственно измерил энергозависимый
- 10. Натрий-калиевый насос После длительной стимуляции аксона, погруженного в раствор с радиоактивным 24Na+, измеряли выход этого иона
- 11. Натрий-калиевый насос Затем в экспериментах с использованием радиоактивного К+, было показано, что выходящий Na+-поток ассоциирован с
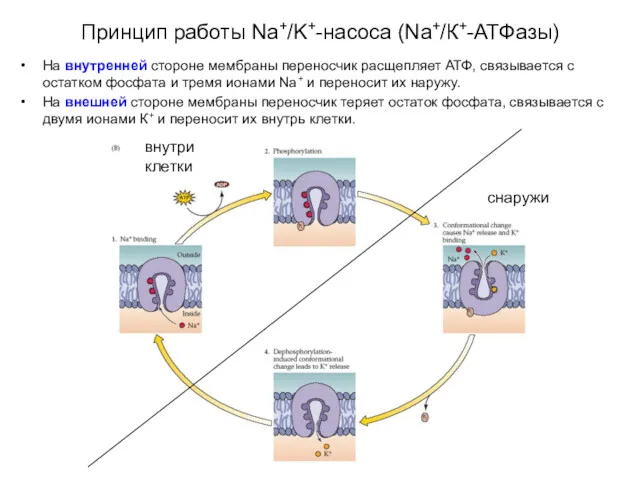
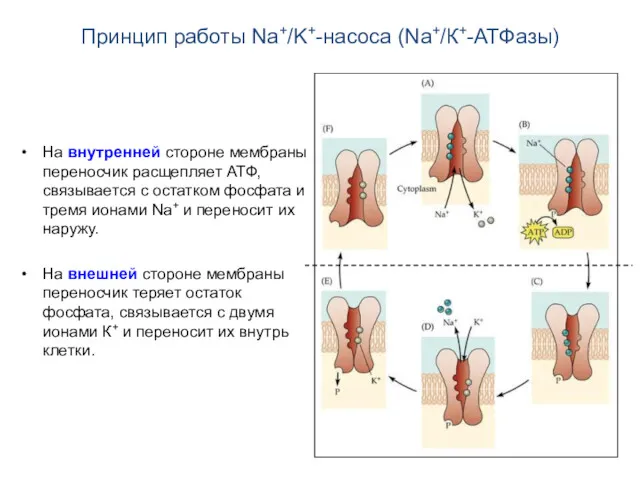
- 12. Принцип работы Na+/K+-насоса (Na+/К+-АТФазы) Переносчик (Na+/K+-насос) выводит из клетки три иона Na+ и вводит в нее
- 13. Принцип работы Na+/K+-насоса (Na+/К+-АТФазы) На внутренней стороне мембраны переносчик расщепляет АТФ, связывается с остатком фосфата и
- 14. Принцип работы Na+/K+-насоса (Na+/К+-АТФазы) На внутренней стороне мембраны переносчик расщепляет АТФ, связывается с остатком фосфата и
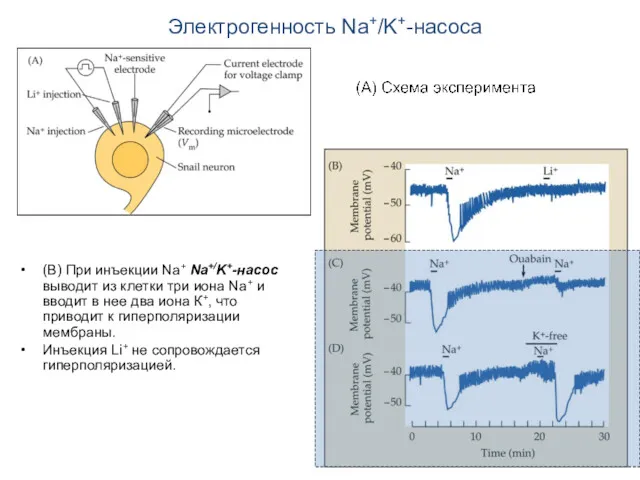
- 15. Электрогенность Na+/K+-насоса (В) При инъекции Na+ Na+/K+-насос выводит из клетки три иона Na+ и вводит в
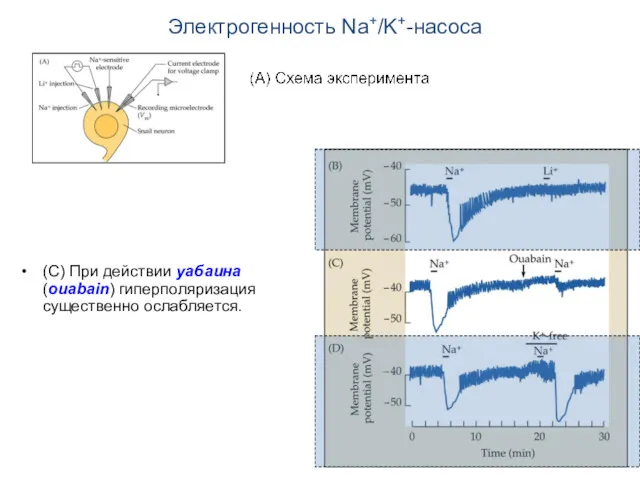
- 16. Электрогенность Na+/K+-насоса (С) При действии уабаина (ouabain) гиперполяризация существенно ослабляется.
- 17. Электрогенность Na+/K+-насоса (D) Удаление К+ блокирует насос, и гиперполяризации не наблюдается до тех пор, пока уровень
- 18. Na+/К+-АТФаза относится к группе P-АТФаз (от Phosphorylation), у которых специальный регуляторный участок (остаток аспарагиновой кислоты) активируется
- 19. Молекулярная структура Na+/K+-АТФазы
- 21. Na+/К+-АТФаза состоит из двух субъединиц - α- (с молекулярной массой около 100 кД) и β- (с
- 22. включает несколько мест связывания с субстратом: участок фосфорилирования, участок связывания АТФ, участок связывания одновалентных катионов, участок
- 23. Са2+-насос (Са2+-АТФаза) является АТФазой P-типа, выводящей Са2+ из цитоплазмы. Эта АТФаза (наряду с Na+/Ca2+-обменником) выполняют главную
- 24. Са2+-насос (Са2+-АТФаза) Существуют две разновидности Са2+-АТФаз Са2+-АТФазы эндоплазматического (в нейронах) и саркоплазматического (в мышечных клетках) ретикулумов,
- 25. Са2+-насос (Са2+-АТФаза) Са2+-АТФазы эндоплазматического (в нейронах) и саркоплазматического (в мышечных клетках) ретикулумов, а также митохондрий при
- 26. Са2+-насос (Са2+-АТФаза) Существуют две разновидности Са2+-АТФаз ! Еще одно отличие. Са2+-АТФаза плазматической мембраны регулируется внутриклеточным комплексом
- 27. Са2+-насос (Са2+-АТФаза) Обе формы Са2+-АТФазы (плазматическая и «органельная») представлены одиночной полипептидной цепочкой с молекулярной массой около
- 28. Протонный насос В мембранах клеточных органелл (лизосом, эндосом, а также синаптических везикул) протонный насос Н+-АТФаза V-типа
- 29. Протонный насос Кроме того в мембранах митохондрий содержится Н+-АТФаза F-типа, работающая в «обратном» направлении: этот фермент
- 30. АТФазы в других клетках Магниевый насос В мембранах эритроцитов выделена Mg2+-АТФаза P-типа, которая с использованием энергии
- 31. АТФазы в других клетках АТФаза F-типа в плазматической мембране бактерий является аналогом Н+-АТФазы F-типа, которая в
- 32. АТФазы в других клетках Хлорный насос Хлорная АТФаза, переносящая ионы Cl- из цитоплазмы во внеклеточное пространство,
- 33. Транспорт других катионов Среди группы P-АТФаз также известны насосы, переносящие через мембраны клеток другие катионы: Ag+
- 34. Механизмы вторичного активного транспорта Переносят некоторые ионы и низкомолекулярные соединения (например, медиаторы) против их концентрационных градиентов,
- 35. Ко-транспортеры и ионные обменники Ко-транспортеры (cotransporter) обеспечивают перенос одних ионов против их концентрационного градиента в том
- 36. Ко-транспортеры и ионные обменники Ко-транспортеры (cotransporter) обеспечивают перенос одних ионов против их концентрационного градиента в том
- 37. Ко-транспортеры и ионные обменники Обменники (exchanger) обеспечивают перенос одних ионов против их концентрационного градиента за счет
- 38. Регуляция внутриклеточной концентрации Са2+ Na+/Ca2+-обменник при значительной деполяризации может работать в противоположном направлении 3 Na+ 1
- 39. Регуляция внутриклеточной концентрации Са2+ Na+/Ca2+-K+-обменник в мембранах фоторецепторов сетчатки Благодаря дополнительной энергии градиента для К+ Na+/Ca2+-К+-обменник
- 40. Регуляция внутриклеточной концентрации Са2+ Система вторичного активного транспорта Са2+ является низкоаффинной (низкая степень связывания катиона), но
- 41. Регуляция внутриклеточной концентрации Сl- К+/Cl--ко-транспортная система выводит Cl- из клетки за счет градиента ионов К+ и
- 42. Регуляция внутриклеточной концентрации Сl- Высокая концентрация внутриклеточного Cl- при развитии нейронов поддерживается благодаря работе одной из
- 43. Регуляция внутриклеточного уровня pH Электрическая активность нейронов и внутриклеточный pH находятся в реципрокных отношениях: увеличение электрической
- 44. Регуляция внутриклеточного уровня pH 1) Внутриклеточный уровень pН поддерживается благодаря Na+/Н+-обменнику, транспортирующего через мембрану один ион
- 45. Регуляция внутриклеточного уровня pH Внутриклеточный уровень pН поддерживается: 2) Na+-зависимым Cl-/HCO3- -обменником, который выводит ионы Cl-
- 46. Регуляция внутриклеточного уровня pH Schematic presentation of the sodium-dependent Cl-/HCO3- exchanger and the sodium-independent Cl-/HCO3-exchanger and
- 47. Регуляция внутриклеточного уровня pH Ион HCO3- выполняет в клетке роль буфера протонов. 4) HCO3- транспортируется посредством
- 48. Транспорт медиаторов из внеклеточного пространства в цитоплазму Транспорт медиаторов через мембраны нейронов и глиальных клеток, а
- 49. Транспорт медиаторов из внеклеточного пространства в цитоплазму Транспортеры медиаторов подразделяются на два семейства (см. табл.): 1)
- 50. Транспортеры медиаторов плазматических мембран
- 51. Транспорт медиаторов из внеклеточного пространства в цитоплазму Схематичная структурная организация медиаторных транспортеров А - схематичная топология
- 52. Транспорт медиаторов из внеклеточного пространства в цитоплазму Схематичная структурная организация медиаторных транспортеров Б - схематичная топология
- 53. Транспорт медиаторов из цитоплазмы в везикулы Транспорт медиаторов в секреторные везикулы обеспечивается специальными системами вторичного активного
- 54. Везикулярные транспортеры медиаторов Везикулярные транспортеры медиаторов включают три семейства (см. табл.), переносящие 1) ГАМК и глицин,
- 55. Везикулярные транспортеры медиаторов
- 56. Везикулярные транспортеры медиаторов
- 57. Везикулярные транспортеры медиаторов Транспортеры (B) ГАМК и глицина и (A) аминов (включая ацетилхолин) являются обменниками и
- 58. Везикулярные транспортеры медиаторов В отличие от транспортеров двух первых семейств переносчики третьего семейства используют только электрический
- 59. Транспортеры могут выступать в роли ионных каналов, а ионные каналы могут выступать в роли транспортеров !?
- 60. Транспортеры могут выступать в роли ионных каналов Обнаружен токсин, который предположительно блокирует Na+/K+-АТФазу, превращая ее в
- 62. Скачать презентацию
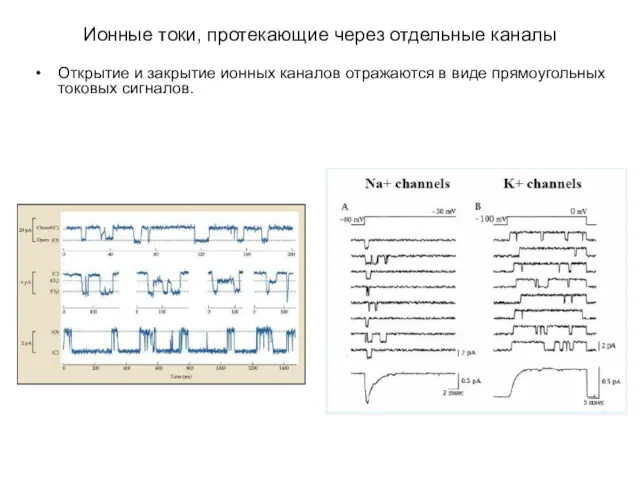
Ионные токи, протекающие через отдельные каналы
Открытие и закрытие ионных каналов отражаются
Ионные токи, протекающие через отдельные каналы
Открытие и закрытие ионных каналов отражаются

Потенциал-зависимые и другие ионные каналы
Каналы, образующие водную пору, включают потенциал-зависимые:
K+-, Ca2+-
Потенциал-зависимые и другие ионные каналы
Каналы, образующие водную пору, включают потенциал-зависимые:
K+-, Ca2+-

Зачем нужны системы активного транспорта?
Зачем нужны системы активного транспорта?

Зачем нужны системы активного транспорта?
Для поддержания нормального функционального состояния нейронов в
Зачем нужны системы активного транспорта?
Для поддержания нормального функционального состояния нейронов в

Два механизма активного транспорта
Известны два механизма активного транспорта:
системы первичного активного транспорта
Два механизма активного транспорта
Известны два механизма активного транспорта:
системы первичного активного транспорта

Механизмы первичного активного транспорта
Системы первичного активного транспорта используют энергию гидролиза АТФ.
В
Механизмы первичного активного транспорта
Системы первичного активного транспорта используют энергию гидролиза АТФ.
В

Натрий-калиевый насос
В начале 1950-х г.г., исследуя действие локальных анестетиков на ионную
Натрий-калиевый насос
В начале 1950-х г.г., исследуя действие локальных анестетиков на ионную
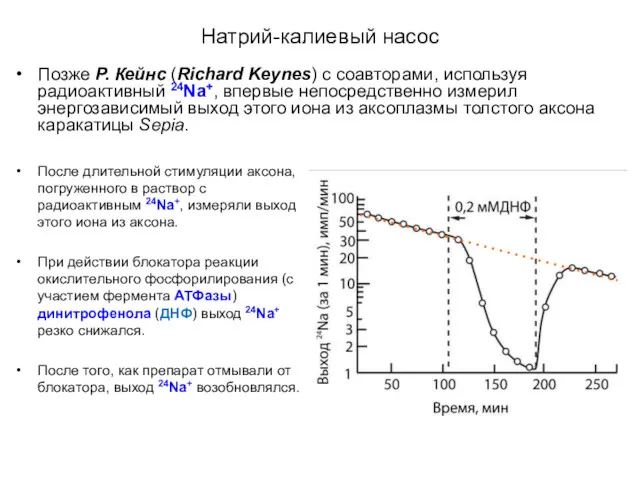
Натрий-калиевый насос
Позже Р. Кейнс (Richard Keynes) с соавторами, используя радиоактивный 24Na+,
Натрий-калиевый насос
Позже Р. Кейнс (Richard Keynes) с соавторами, используя радиоактивный 24Na+,
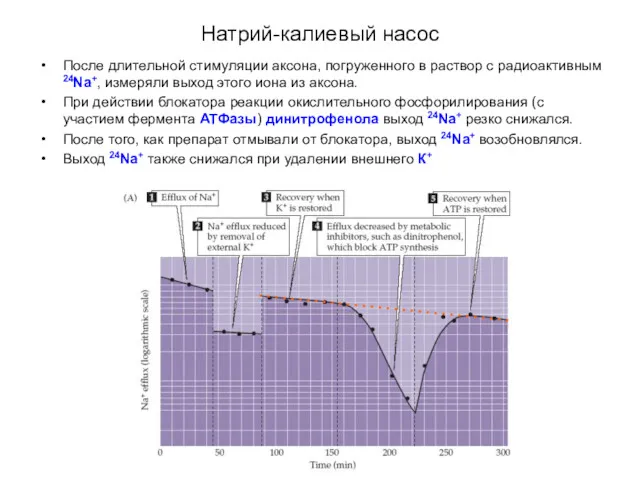
Натрий-калиевый насос
После длительной стимуляции аксона, погруженного в раствор с радиоактивным 24Na+,
Натрий-калиевый насос
После длительной стимуляции аксона, погруженного в раствор с радиоактивным 24Na+,

Натрий-калиевый насос
Затем в экспериментах с использованием радиоактивного К+, было показано, что
Натрий-калиевый насос
Затем в экспериментах с использованием радиоактивного К+, было показано, что
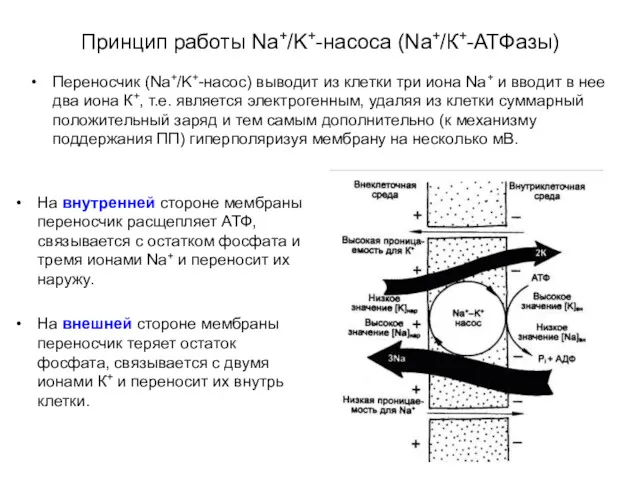
Принцип работы Na+/K+-насоса (Na+/К+-АТФазы)
Переносчик (Na+/K+-насос) выводит из клетки три иона
Принцип работы Na+/K+-насоса (Na+/К+-АТФазы)
Переносчик (Na+/K+-насос) выводит из клетки три иона
Принцип работы Na+/K+-насоса (Na+/К+-АТФазы)
На внутренней стороне мембраны переносчик расщепляет АТФ,
Принцип работы Na+/K+-насоса (Na+/К+-АТФазы)
На внутренней стороне мембраны переносчик расщепляет АТФ,
Принцип работы Na+/K+-насоса (Na+/К+-АТФазы)
На внутренней стороне мембраны переносчик расщепляет АТФ,
Принцип работы Na+/K+-насоса (Na+/К+-АТФазы)
На внутренней стороне мембраны переносчик расщепляет АТФ,
Электрогенность Na+/K+-насоса
(В) При инъекции Na+ Na+/K+-насос выводит из клетки три иона
Электрогенность Na+/K+-насоса
(В) При инъекции Na+ Na+/K+-насос выводит из клетки три иона
Электрогенность Na+/K+-насоса
(С) При действии уабаина (ouabain) гиперполяризация существенно ослабляется.
Электрогенность Na+/K+-насоса
(С) При действии уабаина (ouabain) гиперполяризация существенно ослабляется.
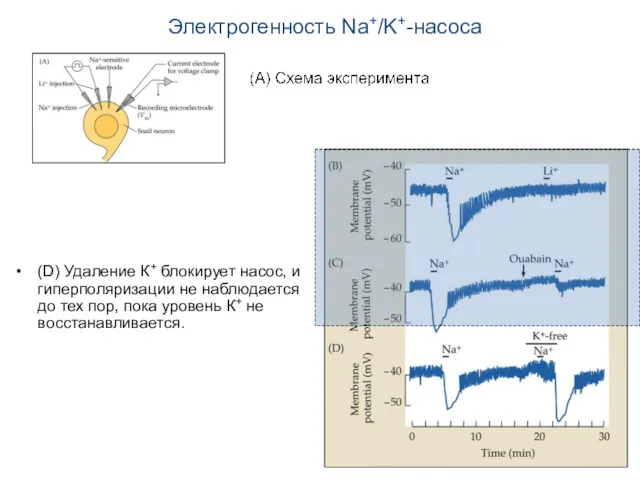
Электрогенность Na+/K+-насоса
(D) Удаление К+ блокирует насос, и гиперполяризации не наблюдается до
Электрогенность Na+/K+-насоса
(D) Удаление К+ блокирует насос, и гиперполяризации не наблюдается до
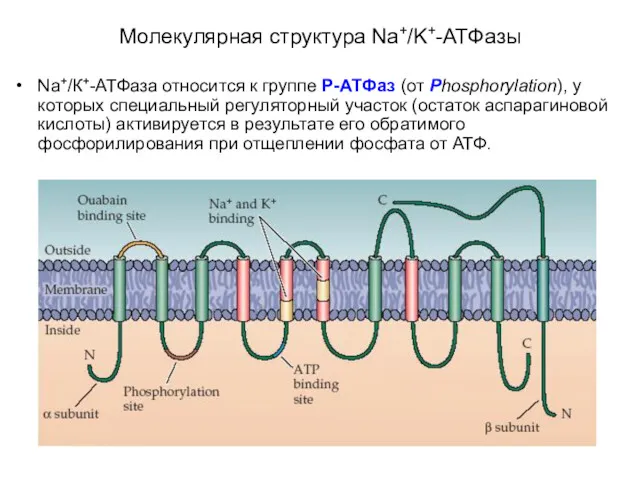
Na+/К+-АТФаза относится к группе P-АТФаз (от Phosphorylation), у которых специальный регуляторный
Na+/К+-АТФаза относится к группе P-АТФаз (от Phosphorylation), у которых специальный регуляторный
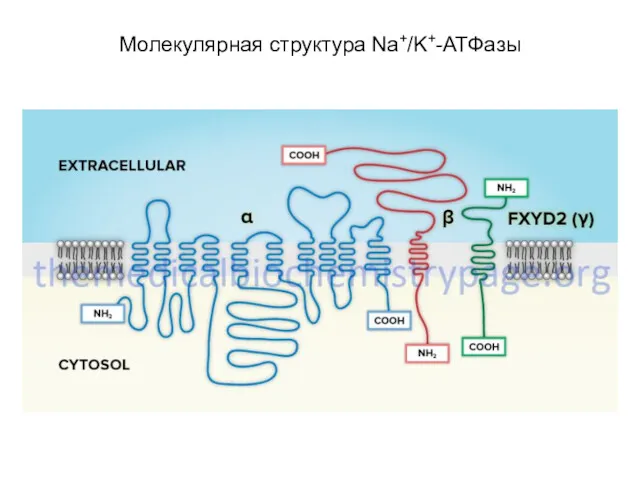
Молекулярная структура Na+/K+-АТФазы
Молекулярная структура Na+/K+-АТФазы
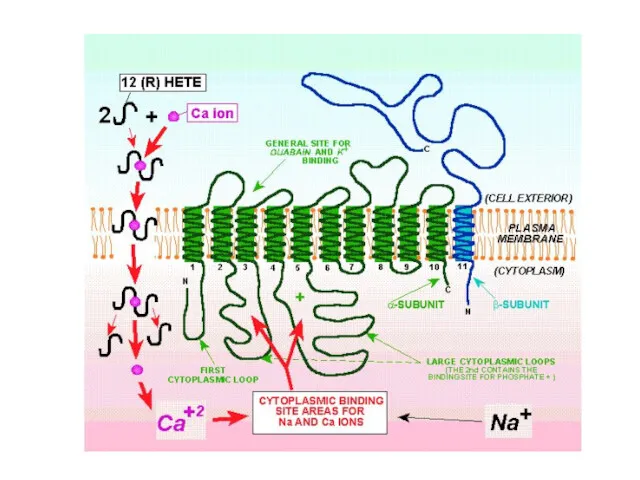
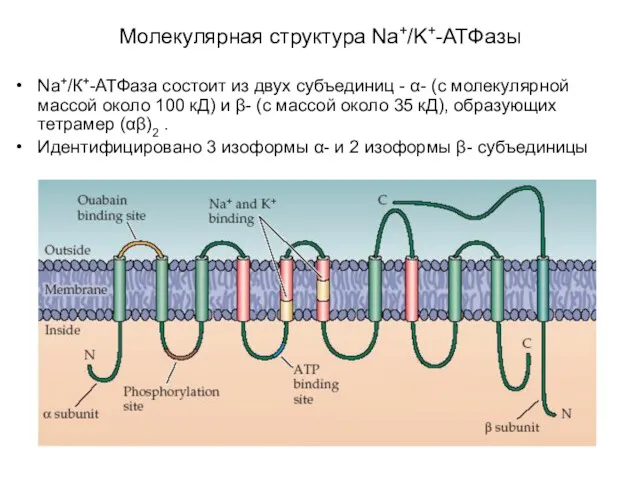
Na+/К+-АТФаза состоит из двух субъединиц - α- (с молекулярной массой около
Na+/К+-АТФаза состоит из двух субъединиц - α- (с молекулярной массой около
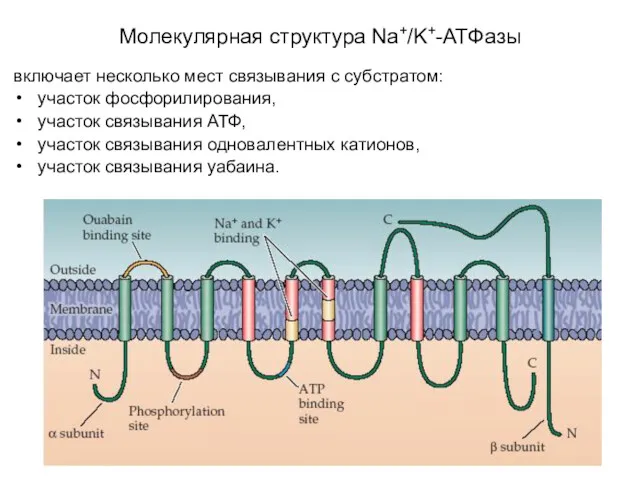
включает несколько мест связывания с субстратом:
участок фосфорилирования,
участок связывания АТФ,
участок связывания одновалентных
включает несколько мест связывания с субстратом:
участок фосфорилирования,
участок связывания АТФ,
участок связывания одновалентных

Са2+-насос (Са2+-АТФаза)
является АТФазой P-типа, выводящей Са2+ из цитоплазмы. Эта АТФаза (наряду
Са2+-насос (Са2+-АТФаза)
является АТФазой P-типа, выводящей Са2+ из цитоплазмы. Эта АТФаза (наряду

Са2+-насос (Са2+-АТФаза)
Существуют две разновидности Са2+-АТФаз
Са2+-АТФазы эндоплазматического (в нейронах) и саркоплазматического (в
Са2+-насос (Са2+-АТФаза)
Существуют две разновидности Са2+-АТФаз
Са2+-АТФазы эндоплазматического (в нейронах) и саркоплазматического (в
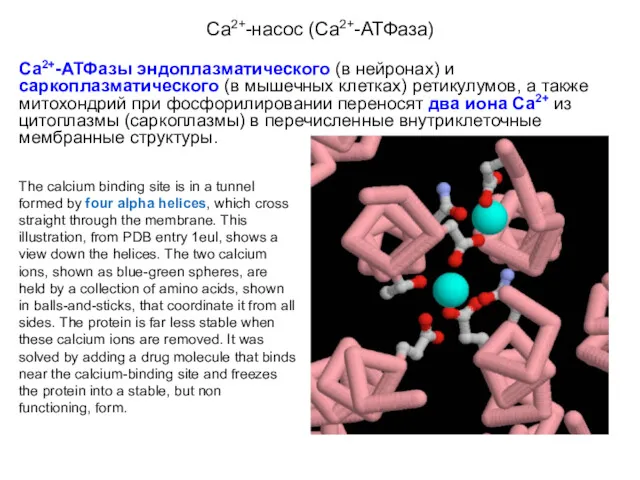
Са2+-насос (Са2+-АТФаза)
Са2+-АТФазы эндоплазматического (в нейронах) и саркоплазматического (в мышечных клетках) ретикулумов,
Са2+-насос (Са2+-АТФаза)
Са2+-АТФазы эндоплазматического (в нейронах) и саркоплазматического (в мышечных клетках) ретикулумов,

Са2+-насос (Са2+-АТФаза)
Существуют две разновидности Са2+-АТФаз
! Еще одно отличие.
Са2+-АТФаза плазматической мембраны регулируется
Са2+-насос (Са2+-АТФаза)
Существуют две разновидности Са2+-АТФаз
! Еще одно отличие.
Са2+-АТФаза плазматической мембраны регулируется

Са2+-насос (Са2+-АТФаза)
Обе формы Са2+-АТФазы (плазматическая и «органельная») представлены одиночной полипептидной цепочкой
Са2+-насос (Са2+-АТФаза)
Обе формы Са2+-АТФазы (плазматическая и «органельная») представлены одиночной полипептидной цепочкой
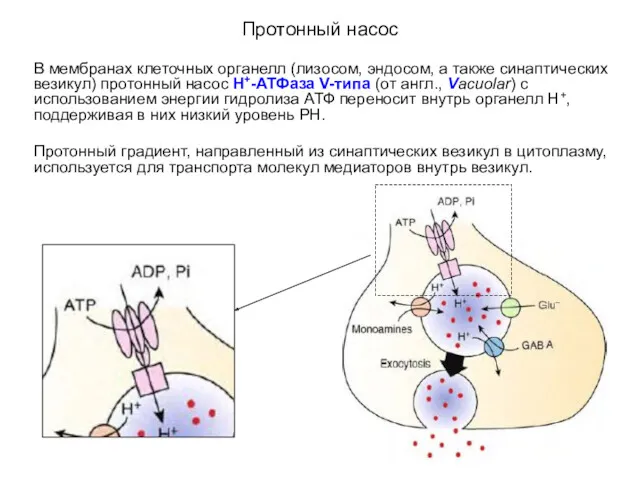
Протонный насос
В мембранах клеточных органелл (лизосом, эндосом, а также синаптических везикул)
Протонный насос
В мембранах клеточных органелл (лизосом, эндосом, а также синаптических везикул)
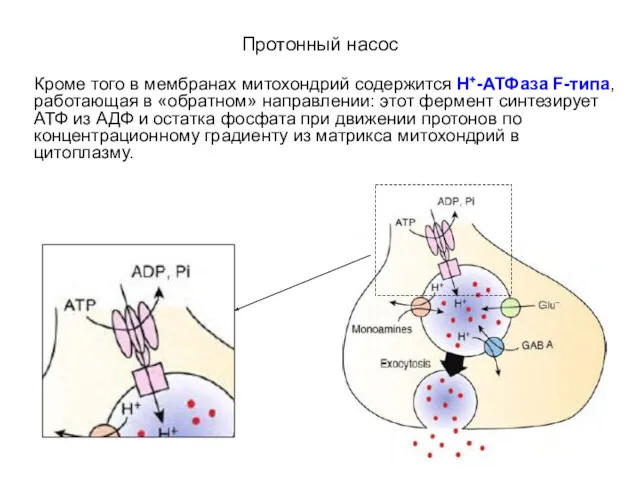
Протонный насос
Кроме того в мембранах митохондрий содержится Н+-АТФаза F-типа, работающая в
Протонный насос
Кроме того в мембранах митохондрий содержится Н+-АТФаза F-типа, работающая в

АТФазы в других клетках
Магниевый насос
В мембранах эритроцитов выделена Mg2+-АТФаза P-типа, которая
АТФазы в других клетках
Магниевый насос
В мембранах эритроцитов выделена Mg2+-АТФаза P-типа, которая

АТФазы в других клетках
АТФаза F-типа в плазматической мембране бактерий
является аналогом Н+-АТФазы
АТФазы в других клетках
АТФаза F-типа в плазматической мембране бактерий
является аналогом Н+-АТФазы

АТФазы в других клетках
Хлорный насос
Хлорная АТФаза, переносящая ионы Cl- из цитоплазмы
АТФазы в других клетках
Хлорный насос
Хлорная АТФаза, переносящая ионы Cl- из цитоплазмы

Транспорт других катионов
Среди группы P-АТФаз также известны насосы, переносящие через мембраны
Транспорт других катионов
Среди группы P-АТФаз также известны насосы, переносящие через мембраны

Механизмы вторичного активного транспорта
Переносят некоторые ионы и низкомолекулярные соединения (например, медиаторы)
Механизмы вторичного активного транспорта
Переносят некоторые ионы и низкомолекулярные соединения (например, медиаторы)
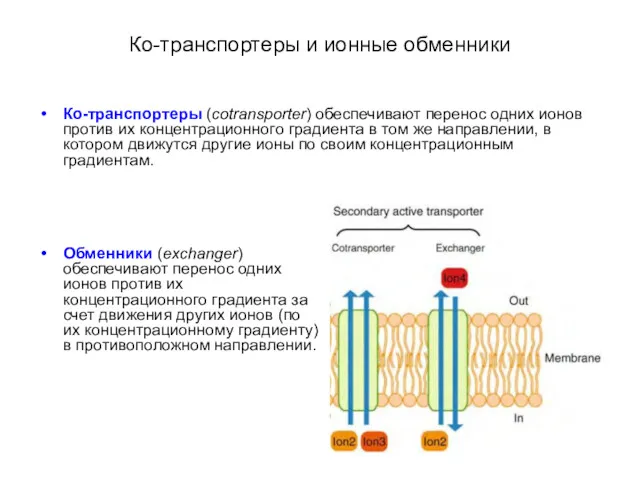
Ко-транспортеры и ионные обменники
Ко-транспортеры (cotransporter) обеспечивают перенос одних ионов против их
Ко-транспортеры и ионные обменники
Ко-транспортеры (cotransporter) обеспечивают перенос одних ионов против их
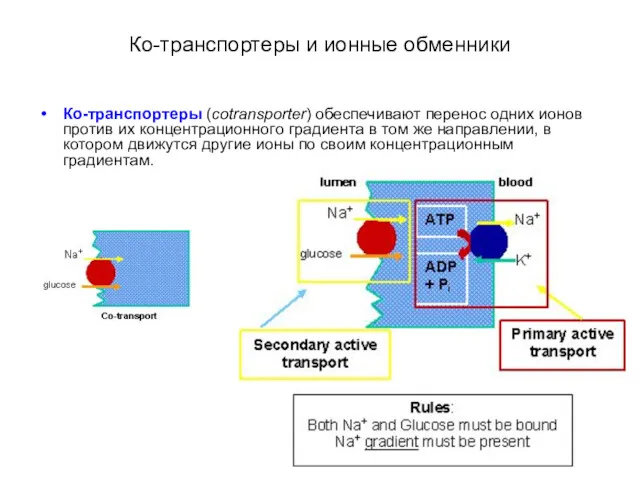
Ко-транспортеры и ионные обменники
Ко-транспортеры (cotransporter) обеспечивают перенос одних ионов против их
Ко-транспортеры и ионные обменники
Ко-транспортеры (cotransporter) обеспечивают перенос одних ионов против их
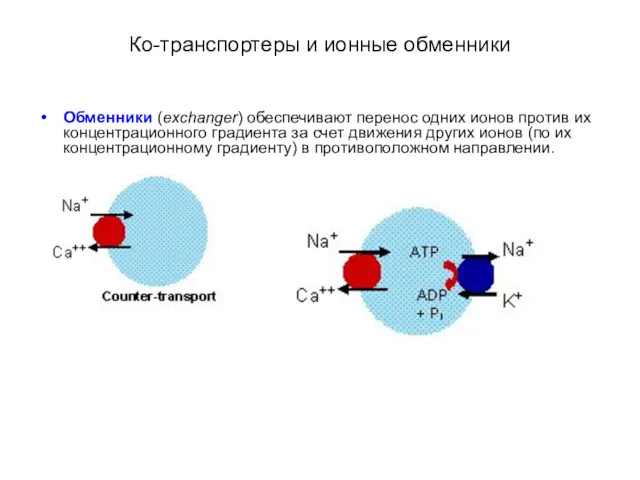
Ко-транспортеры и ионные обменники
Обменники (exchanger) обеспечивают перенос одних ионов против их
Ко-транспортеры и ионные обменники
Обменники (exchanger) обеспечивают перенос одних ионов против их
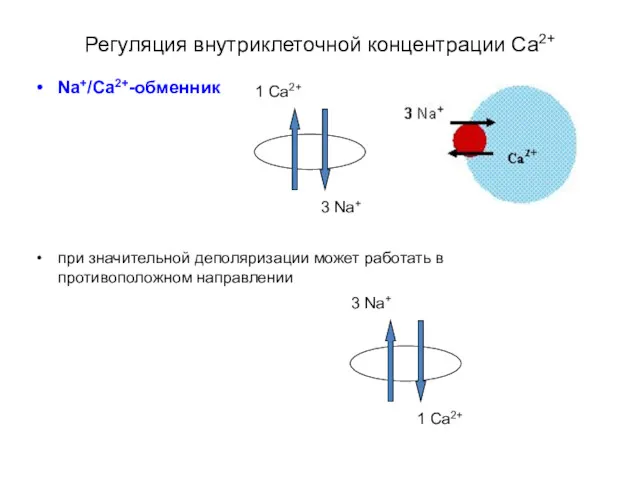
Регуляция внутриклеточной концентрации Са2+
Na+/Ca2+-обменник
при значительной деполяризации может работать в противоположном направлении
Регуляция внутриклеточной концентрации Са2+
Na+/Ca2+-обменник
при значительной деполяризации может работать в противоположном направлении
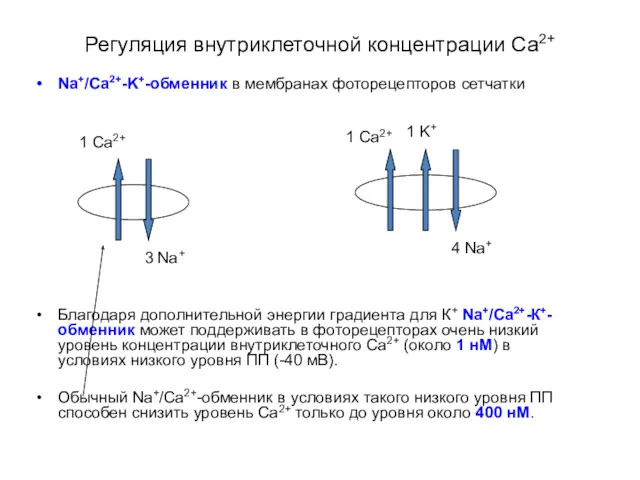
Регуляция внутриклеточной концентрации Са2+
Na+/Ca2+-K+-обменник в мембранах фоторецепторов сетчатки
Благодаря дополнительной энергии градиента
Регуляция внутриклеточной концентрации Са2+
Na+/Ca2+-K+-обменник в мембранах фоторецепторов сетчатки
Благодаря дополнительной энергии градиента

Регуляция внутриклеточной концентрации Са2+
Система вторичного активного транспорта Са2+ является низкоаффинной (низкая
Регуляция внутриклеточной концентрации Са2+
Система вторичного активного транспорта Са2+ является низкоаффинной (низкая
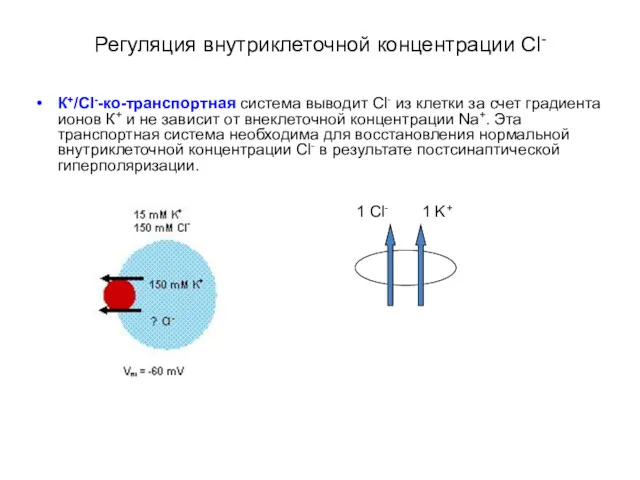
Регуляция внутриклеточной концентрации Сl-
К+/Cl--ко-транспортная система выводит Cl- из клетки за счет
Регуляция внутриклеточной концентрации Сl-
К+/Cl--ко-транспортная система выводит Cl- из клетки за счет
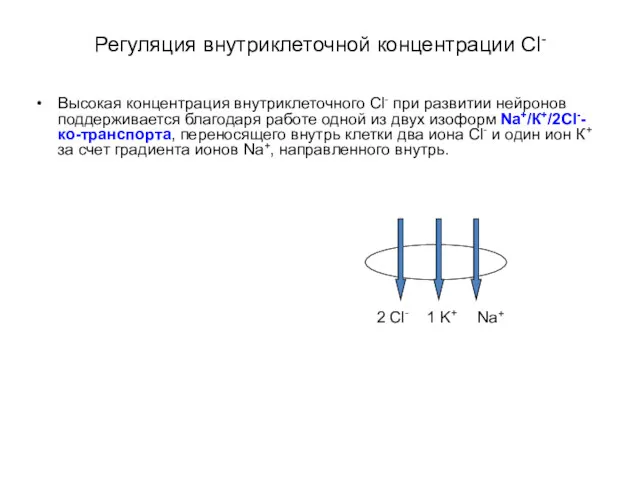
Регуляция внутриклеточной концентрации Сl-
Высокая концентрация внутриклеточного Cl- при развитии нейронов поддерживается
Регуляция внутриклеточной концентрации Сl-
Высокая концентрация внутриклеточного Cl- при развитии нейронов поддерживается

Регуляция внутриклеточного уровня pH
Электрическая активность нейронов и внутриклеточный pH находятся в
Регуляция внутриклеточного уровня pH
Электрическая активность нейронов и внутриклеточный pH находятся в
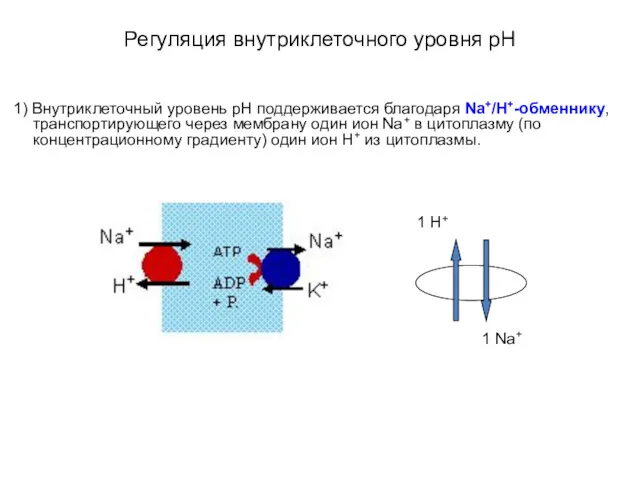
Регуляция внутриклеточного уровня pH
1) Внутриклеточный уровень pН поддерживается благодаря Na+/Н+-обменнику, транспортирующего
Регуляция внутриклеточного уровня pH
1) Внутриклеточный уровень pН поддерживается благодаря Na+/Н+-обменнику, транспортирующего
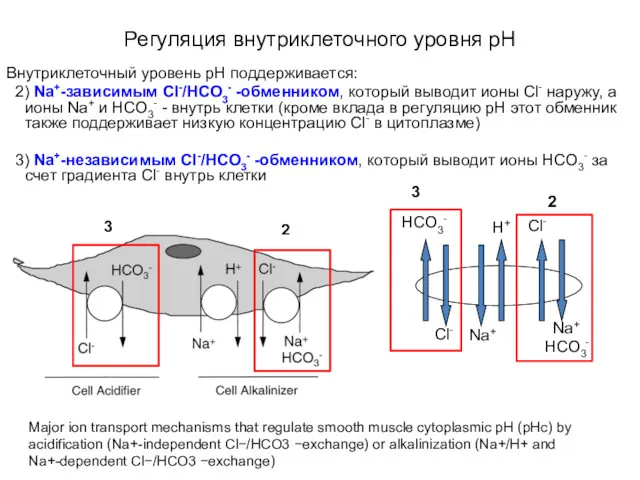
Регуляция внутриклеточного уровня pH
Внутриклеточный уровень pН поддерживается:
2) Na+-зависимым Cl-/HCO3- -обменником,
Регуляция внутриклеточного уровня pH
Внутриклеточный уровень pН поддерживается:
2) Na+-зависимым Cl-/HCO3- -обменником,
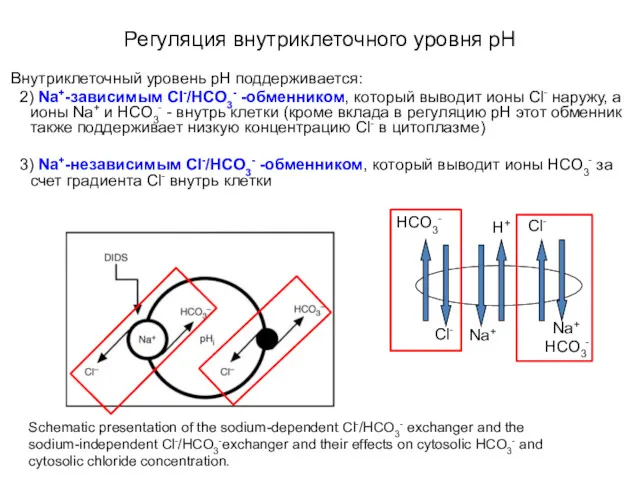
Регуляция внутриклеточного уровня pH
Schematic presentation of the sodium-dependent Cl-/HCO3- exchanger and
Регуляция внутриклеточного уровня pH
Schematic presentation of the sodium-dependent Cl-/HCO3- exchanger and
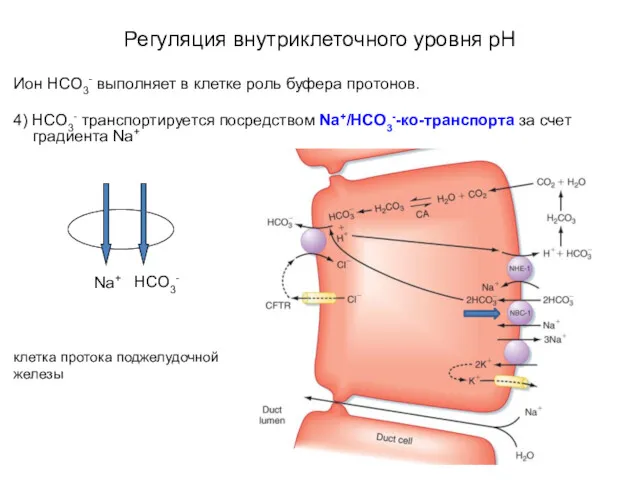
Регуляция внутриклеточного уровня pH
Ион HCO3- выполняет в клетке роль буфера протонов.
4)
Регуляция внутриклеточного уровня pH
Ион HCO3- выполняет в клетке роль буфера протонов.
4)
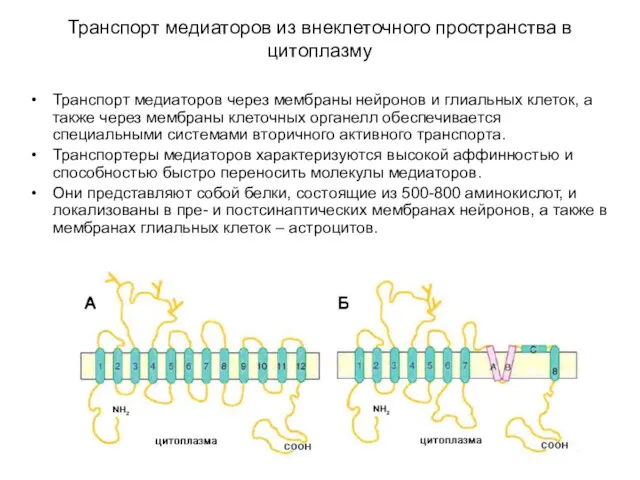
Транспорт медиаторов из внеклеточного пространства в цитоплазму
Транспорт медиаторов через мембраны нейронов
Транспорт медиаторов из внеклеточного пространства в цитоплазму
Транспорт медиаторов через мембраны нейронов
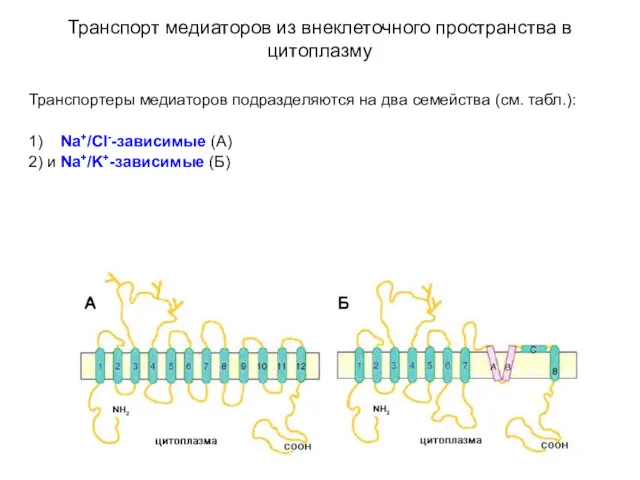
Транспорт медиаторов из внеклеточного пространства в цитоплазму
Транспортеры медиаторов подразделяются на два
Транспорт медиаторов из внеклеточного пространства в цитоплазму
Транспортеры медиаторов подразделяются на два
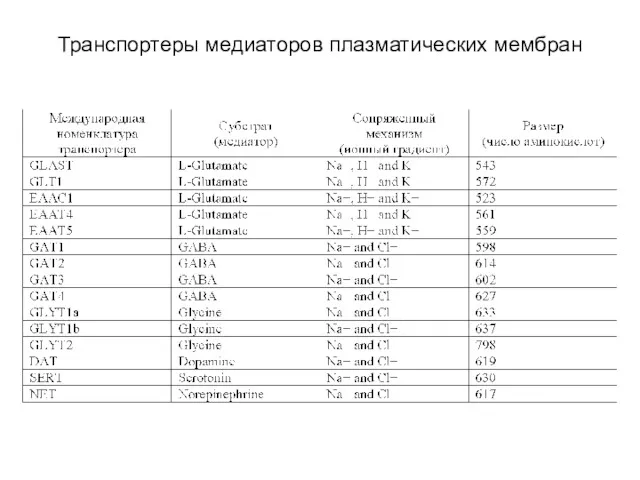
Транспортеры медиаторов плазматических мембран
Транспортеры медиаторов плазматических мембран
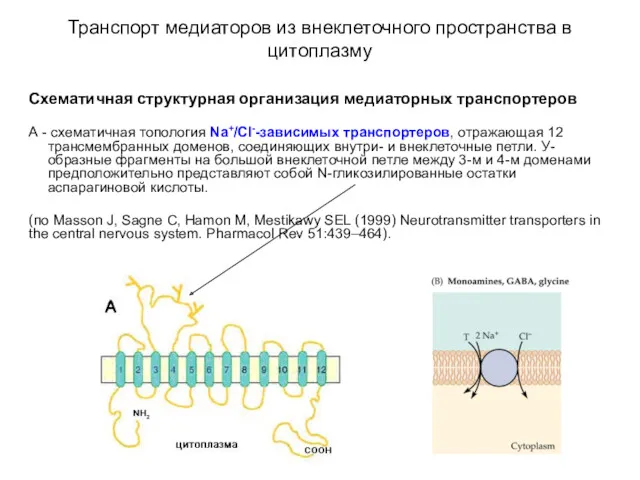
Транспорт медиаторов из внеклеточного пространства в цитоплазму
Схематичная структурная организация медиаторных транспортеров
А
Транспорт медиаторов из внеклеточного пространства в цитоплазму
Схематичная структурная организация медиаторных транспортеров
А
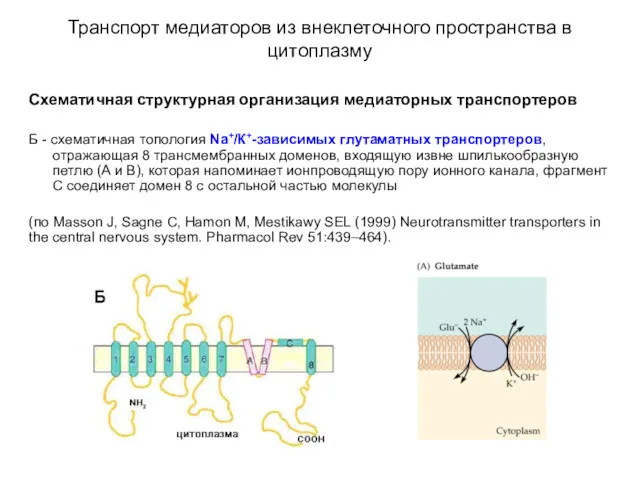
Транспорт медиаторов из внеклеточного пространства в цитоплазму
Схематичная структурная организация медиаторных транспортеров
Б
Транспорт медиаторов из внеклеточного пространства в цитоплазму
Схематичная структурная организация медиаторных транспортеров
Б

Транспорт медиаторов из цитоплазмы в везикулы
Транспорт медиаторов в секреторные везикулы
Транспорт медиаторов из цитоплазмы в везикулы
Транспорт медиаторов в секреторные везикулы

Везикулярные транспортеры медиаторов
Везикулярные транспортеры медиаторов включают три семейства (см. табл.), переносящие
1)
Везикулярные транспортеры медиаторов
Везикулярные транспортеры медиаторов включают три семейства (см. табл.), переносящие
1)
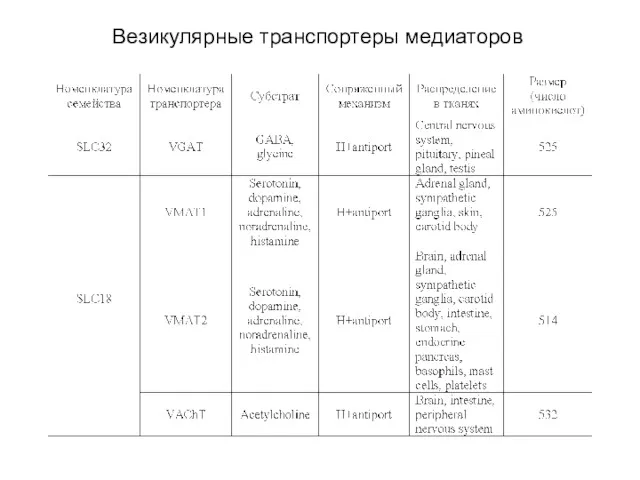
Везикулярные транспортеры медиаторов
Везикулярные транспортеры медиаторов
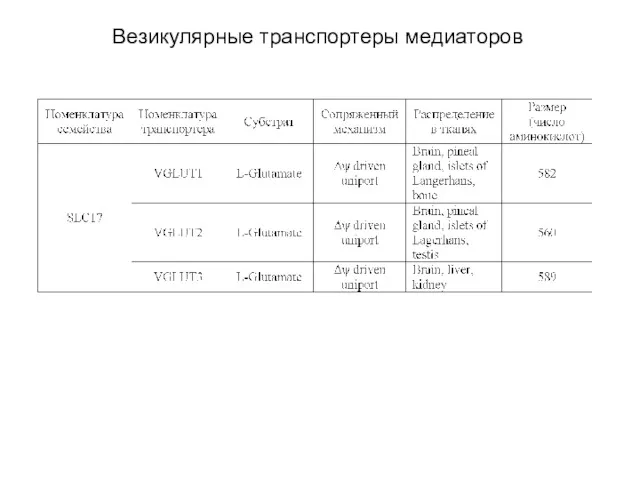
Везикулярные транспортеры медиаторов
Везикулярные транспортеры медиаторов
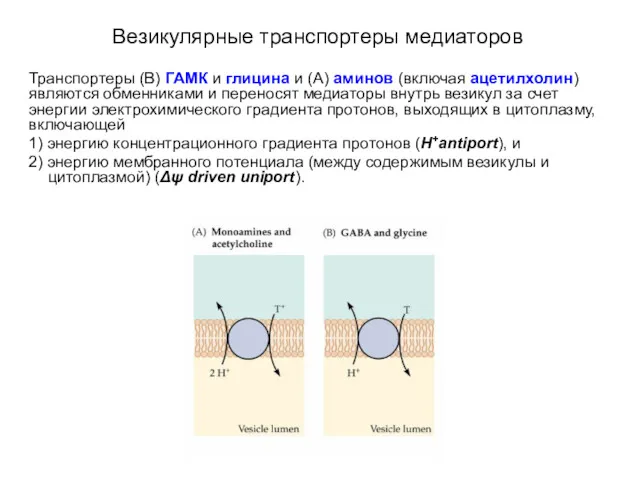
Везикулярные транспортеры медиаторов
Транспортеры (B) ГАМК и глицина и (A) аминов (включая
Везикулярные транспортеры медиаторов
Транспортеры (B) ГАМК и глицина и (A) аминов (включая

Везикулярные транспортеры медиаторов
В отличие от транспортеров двух первых семейств переносчики третьего
Везикулярные транспортеры медиаторов
В отличие от транспортеров двух первых семейств переносчики третьего

Транспортеры могут выступать в роли ионных каналов, а ионные каналы могут
Транспортеры могут выступать в роли ионных каналов, а ионные каналы могут

Транспортеры могут выступать в роли ионных каналов
Обнаружен токсин, который предположительно блокирует
Транспортеры могут выступать в роли ионных каналов
Обнаружен токсин, который предположительно блокирует
 Речные рыбы
Речные рыбы Растительная пища
Растительная пища Биогеохимический цикл фосфора
Биогеохимический цикл фосфора Ризосферные плазмидосодержащие бактерии рода Pseudomonas для био/фиторемедиации загрязненных почв
Ризосферные плазмидосодержащие бактерии рода Pseudomonas для био/фиторемедиации загрязненных почв Место человека в живой природе
Место человека в живой природе Кровообращение плода
Кровообращение плода Зоология беспозвоночных. Учебная программа
Зоология беспозвоночных. Учебная программа Задачи на биосинтез со штрихами
Задачи на биосинтез со штрихами Клиническая анатомия носа и околоносовых пазух
Клиническая анатомия носа и околоносовых пазух Хвойные растения
Хвойные растения Презентация к уроку биологии 7 класс
Презентация к уроку биологии 7 класс Как устроен организм человека
Как устроен организм человека Физиология центральной нервной системы
Физиология центральной нервной системы Презентация Весна
Презентация Весна Голоса птиц и животных леса
Голоса птиц и животных леса Механизмы репарации
Механизмы репарации Флокуляційне очищення питної води за допомогою катіонних та аніонних флокулянтів
Флокуляційне очищення питної води за допомогою катіонних та аніонних флокулянтів Корень. Значение корня. Виды корней. Корневые системы. Микроскопическое строение корня. Зоны корня. Корневой волосок
Корень. Значение корня. Виды корней. Корневые системы. Микроскопическое строение корня. Зоны корня. Корневой волосок Нейрофизиология эмоций
Нейрофизиология эмоций КВН. Прокариоты
КВН. Прокариоты Учение Ч. Дарвина об естественном отборе
Учение Ч. Дарвина об естественном отборе Водоросли – низшие растения
Водоросли – низшие растения Насекомые красной книги РФ
Насекомые красной книги РФ Органы пищеварения
Органы пищеварения Развитие жизни на земле
Развитие жизни на земле Разнообразие животных
Разнообразие животных Биологические шарады
Биологические шарады Рецепторы клетки
Рецепторы клетки