- Биологиялық тотығу электрондардың тасымалдану тізбегі тотығудан фосфорлану

Содержание
- 2. 7. Субстраттан фосфорлану реакциясы. Сукцинаттиокиназа әсерінен Сукцинил-КоА-дағы макроэргиялық байланыс үзіліп Сукцинат(янтарь қышқылы) түзіледі. Бөлінген энергия арқылы
- 3. Түзілген ГТФ нуклеозиддифосфат− киназа көмегімен АТФ-ке айналады: ГТФ + АДФ → АТФ + ГДФ
- 4. 8. 3−ші тотығу-тотықсыздану реакциясы. ФАД−тәуелді сукцинатдегидрогеназа(СДГ) әсерінен Сукцинаттан фумарат және тотықсызданған ФАДН2 түзіледі.
- 5. 9. Фумараза фумаратты гидратациялап малат түзіледі.
- 6. 10. 4−ші тотығу-тотықсыздану реакциясы . НАД−тәуелді малатдегидрогеназа (МДГ) әсерінен Малаттан ҚСҚ және НАДН2 түзіледі.
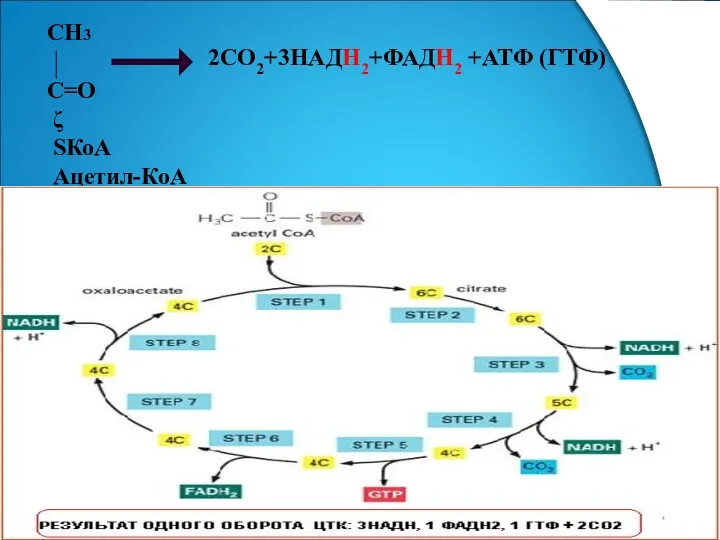
- 7. СН3 С=О ζ SКоА Ацетил-КоА 2СО2+3НАДН2+ФАДН2 +АТФ (ГТФ)
- 8. ДӘРІС N 8 БИОЛОГИЯЛЫҚ ТОТЫҒУ, ЭЛЕКТРОНДАРДЫҢ ТАСЫМАЛДАНУ ТІЗБЕГІ. ТОТЫҒУДАН ФОСФОРЛАНУ
- 9. ДӘРІС ЖОСПАРЫ: БИОЛОГИЯЛЫҚ ТОТЫҒУ, ЭЛЕКТРОНДАРДЫҢ ТАСЫМАЛДАНУ ТІЗБЕГІ БИОЛОГИЯЛЫҚ ТОТЫҒУ САТЫЛАРЫ ТОТЫҒУДАН ФОСФОРЛАНУ Р\О КОЭФФИЦИЕНТІ ТУРАЛЫ ТҮСІНІК
- 10. 3 САТЫ. БИОЛОГИЯЛЫҚ ТОТЫҒУ (БТ ) БТ— ЖАСУША МИТОХОНДРИЯСЫНДА ӨТЕТІН ТОТЫҒУ–ТОТЫҚСЫЗДАНУ (Т-Т) РЕАКЦИЯЛАРЫНЫҢ ЖИЫНЫ. Т-Т РЕАКЦИЯЛАРЫНЫҢ
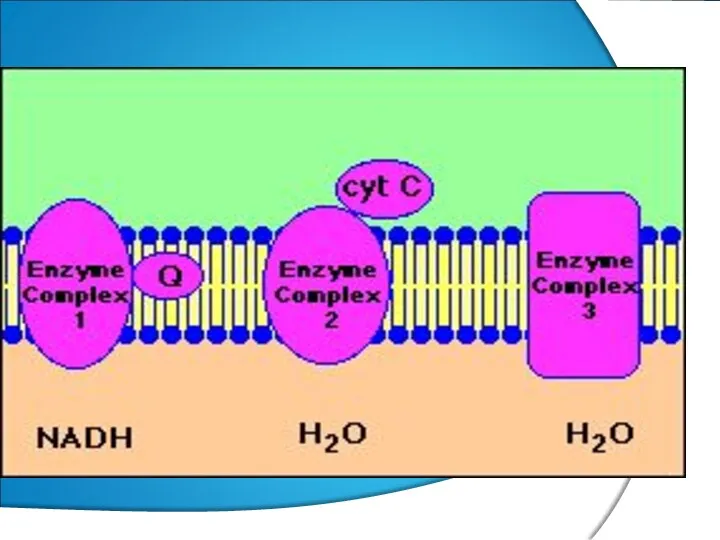
- 12. ПФ(НАД) → (ФП (ФМН)→КоQ →Цхb → с1→с→ а→а3 →О2 . SН2
- 13. БИОЛОГИЯЛЫҚ ТОТЫҒУ(БТ) МАҢЫЗЫ: 1. ЭНЕРГИЯ (Е) БӨЛІНЕДІ. (Е=40-48% АТФ +52-60% ЖЫЛУ) 2. ЭНДОГЕНДІ СУ ТҮЗІЛЕДІ. SН2+
- 14. БТ ОКСИДОРЕДУКТАЗАЛАР ҚАТЫСУЫМЕН МИТОХОНДРИЯДА ЖҮРЕДІ: 11 РЕАКЦИЯДАН ТҰРАДЫ.
- 15. БТ тізбегіндегі ферменттердің орналасу тәртібі неге тәуелді? Е , В Н/ Н2 -0.41 НАД\НАДН2 -0,32 ФМН\ФМНН2
- 16. Тотығатын зат әртүрлі зат алмасуында түзілген органикалық қышқылдар, негізінен ҮКЦ-де түзілген орг. қышқылдар болып табылады.
- 17. Субстрат (лактат) коферменті НАД болатын пиридинферментпен (ПФ) тотығады (дегидрлену жолымен). НАД НАДН2-ге тотықсызданады. Апофермент коферменттен бөлінеді.Бұл
- 18. II. НАДН2 ФП(ФМН) флавопротеидпен тотығады, ал ФП тотықсызданады. Бұл сатыда 46кДж энергия бөлінеді. Осы энергияның шамамен
- 19. III. Тотықсызданған ФПН2 екі сутегін убихинонға (КоQ) беріп тотығады, ал убихинон тотықсызданады (КоQН2).
- 20. IV. КоQН2 тотыққан кезде 2Н0→2Н++2е-ге ыдырайды. Н+ ерітіндіге ауысады, ал е- цитохром жүйесі арқылы оттекке беріледі:
- 21. Тоттыққан ж\е тотықсызданған цитохром гемдері цитохромдар электрондарды тасымалдау жолымен субстратты тотықтырады. Ол кезде цх геміндегі Ғе
- 22. Убихиноннан бөлінген 2е- 2 ферри цхb-ға беріліп оны ферро формаға айналдырады. 2е־ 3, 4, 5 САТЫДА
- 23. VI. 2ферро цхb 2e־ 2ферри цхс1-ге беріп тотығады, ал 2Цхс1 (Fe2+ ) ауысады. +43кДж
- 24. VII. 2 ферро цх с1 2e־ 2ферри цхс-ға беріп тотығады, ал 2Цхс тотықсызданады.
- 25. VIII. 2ферроЦхс 2e־ 2ферри цха–ға беріп тотығады, ал ферри 2Цха тотықсызданады, 7,8-сатыда аз энергия бөлініп, жылу
- 26. 9- және 10-сатылар тығыз байланысты, себебі цитохромоксидазды мультиферменттік комплекс (2Цха+4Цха3 +6Сu) қатысады. IX. 2ферро Цх а
- 27. X. 2ферро Цха3 2е־ О2 беріп тотығады, ал оттек ионданады(тотықсызданады). О2 тотықсыздану үшін 4е ־керек. Оттекпен
- 28. XI. Эндогенді судың түзілуі. Ионданған оттек КоQН2 тотыққанда (IV пункт) түзілген 2 H+ мен әрекеттесіп эндогенді
- 29. Қорытынды: БТ-ның 3 нүктесінде (сатысында) АТФ түзуге жеткілікті энергия бөлінеді. Осы сатылар БТ мен ТФ-ның қабысу
- 31. ЭНЕРГИЯ АЛМАСУЫНЫҢ 4 САТЫСЫ: ТОТЫҒУДАН ФОСФОРЛАНУ (ТФ). ТФ − БИОЛОГИЯЛЫҚ ТОТЫҒУ ЭНЕРГИЯСЫН ПАЙДАЛАНЫП АДФ ПЕН БЕЙОРГАНИКАЛЫҚ
- 32. АТФ-синтаза
- 33. Р/О−ТОТЫҒУДАН ФОСФОРЛАНУ КОЭФФИЦИЕНТІ. Р/О − АТФ־ ТІ ТҮЗУГЕ КЕТКЕН БЕЙОРГАНИКАЛЫҚ ФОСФАТТЫҢ МОЛЬ САНЫН КӨРСЕТЕДІ (СІҢІРІЛГЕН ОТТЕКТІҢ
- 34. ЕГЕР СУБСТРАТ ПИРИДИН ФЕРМЕНТІ ПФ(НАД) АРҚЫЛЫ ТОТЫҚСА: Р/О = 3; себебі бұл кезде бөлінген энергия 3АТФ
- 35. 1МОЛЕКУЛ АСҚ ТОТЫҚҚАНДА 12АТФ ТҮЗІЛЕДІ. АСҚ 2СО2+3НАДН2+ФПН2 +АТФ БТ БТ ЭНЕРГИЯ + Н2О ЭНЕРГИЯ+Н2О ТФ ЖЫЛУ
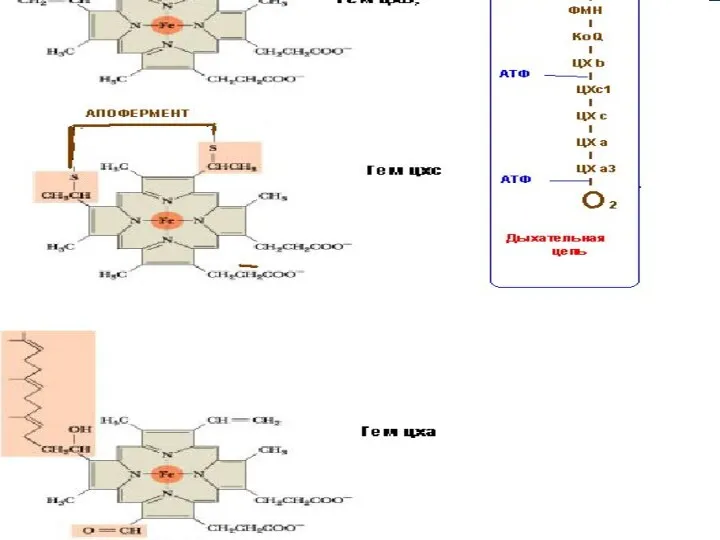
- 36. Ферменттік ансамбль. БТ тізбегіне қатысатын ферменттер сызықты түрде орналаспайды, олар 4 комплекске (тыныс алу ансамблі) бірігіп
- 37. ФП(ФМН) Цх b-Цхс1 2Цха +4Цха3 6Сu ФП(ФАД) S Н2 НАД Н2 НАД S КоQ Цхс О2
- 38. I ж/е II комплекс КоQ арқылы III –комплекспен байланысады, ал III ж/е IV–комплекс Цхс арқылы бір
- 39. БТ МЕН ТФ ӨЗАРА ТЫҒЫЗ БАЙЛАНЫСТЫ. БТ ЖЫЛДАМДЫҒЫН ТФ БАҚЫЛАЙДЫ, МҰНЫ ТЫНЫС АЛУДЫ БАҚЫЛАУ ДЕЙДІ. ЕГЕР:
- 41. ЭНЕРГИЯ АЛМАСУЫ САТЫЛАРЫ БІР-БІРІМЕН ТЫҒЫЗ БАЙЛАНЫСТЫ. 1-2 САТЫСЫ: АСҚ АРҚЫЛЫ БАЙЛАНЫСАДЫ. 1 САТЫДА ТҮЗІЛГЕН АСҚ 2САТЫ
- 42. ГИПОЭНЕРГЕТИКАЛЫҚ КҮЙ ־АТФ СИНТЕЗІНІҢ ТӨМЕНДЕУІ: АШТЫҚ О2 ЖАСУШАНЫ ОТТЕКПЕН ҚАМТАМАСЫЗ ЕТЕТІН ЖҮРЕК־ҚАН ТАМЫРЛАРЫ МЕН ТЫНЫС АЛУ
- 43. Ротенон Барбитурат прогестерон
- 44. БИОЛОГИЯЛЫҚ ТОТЫҒУ ФЕРМЕНТТЕРІ == ТЫНЫС АЛУ ФЕРМЕНТТЕРІ = ЭЛЕКТРОН ТАСЫМАЛДАУШЫ ЖҮЙЕ
- 45. Митохондрия мембранасындағы тыныс алу комплексі
- 46. БТ ИНГИБИТОРЛАРЫ НАДН-ДЕГИДРОГЕНАЗА ИНГИБИТОРЛАРЫ: БАРБИТУРАТТАР (ВЕНОНАЛ, ГЕКСЕНАЛ, НЕМБУТАЛ, АМИТАЛ); ЦХв −с ИНГИБИТОРЛАРЫ- АНТИМИЦИН А; ЦИТОХРОМОКСИДАЗА ИНГИБИТОРЛАРЫ-
- 47. БТ АЖЫРАТҚЫШТАРЫ КЕЙБІР ЛИПОФИЛЬДІ ПРОТОНОФОРЛАР МИТОХОНДРИЯ МЕМБРАНАСЫНЫҢ ӨТКІЗГІШТІГІН БҰЗЫП ПРОТОНДАРДЫ ПРОТОНДЫҚ НАСОС АРҚЫЛЫ ЕМЕС МИТОХОНДРИЯ МЕМБРАНАСЫ
- 48. ЭКЗОГЕНДІ АЖЫРАТҚЫШТАР−2,4-ДИНИТРОФЕНОЛ, ДИКУМАРОЛ, СТРЕПТОМИЦИН; ЭНДОГЕНДІ АЖЫРАТҚЫШТАР−МАЙ ҚЫШҚЫЛДАРЫ, ҚАЛҚАНША БЕЗІНІҢ ГОРМОНЫ־ ТИРОКСИН, ӨТ ПИГМЕНТІ־БИЛИРУБИН, ТЕРМОГЕНИН БЕЛОГЫ. ФОСФОРЛАНУДЫҢ
- 49. ӘДЕБИЕТТЕР: Негізгі: Т.Ш.Шарманов, С.М.Плешкова «Метаболические основы питания с курсом общей биохимии», Алматы, 1998 г. С.Тапбергенов «Медицинская
- 50. НАЗАРЛАРЫҢЫЗҒА РАХМЕТ!
- 51. Цитохромная система Существует несколько групп цитохромов –А,В, С, Д, которые являются гемопротеидами ( дыхательными ферментами) и
- 52. ГЕМЫ ЦИТОХРОМОВ b c a
- 53. В цепь биологического окисления или тканевого дыхания вступают в основном НАДН2, который получается в результате бета-окисления
- 54. Взаимодействие дыхательных комплексов и роль при этом кофермента Q и цитохрома с Кофермент Q получает 2Н
- 55. ПЕРЕНОС ЭЛЕКТРОНОВ ЦХ С НА ЦИТОХРОМОКСИДАЗУ 1.2 Цх с(Fe2+) +2 Cu2+ → 2 цхс(Fe3+) + 2
- 56. Тыныс алу тіэбегіндегі электрондардың тасымалдануы
- 57. ТЫНЫС АЛУ КОМПЛЕКСІНІҢ ЖҰМЫСЫ
- 58. В результате работы ДК внутренняя мембрана Мх заряжается Она разряжается путем возвращения Н+ в матрикс Мх
- 59. АТФ-СИНТАЗА –КОМПЛЕКС V Комплекс, катализирующий образование АТФ=окислительное фосфорилирование, называется АТФ синтазой, или комплексом V; он содержит
- 60. Схема переноса электронов и работы АТФ-СИНТАЗЫ При работе дыхательных ферментов катионы водорода выталкиваются в межмембранное прост-ранство.Затем
- 61. ХЕМООСМОТИЧЕСКОЕ СОПРЯЖЕНИЕ
- 63. Скачать презентацию

7. Субстраттан фосфорлану реакциясы.
Сукцинаттиокиназа әсерінен
Сукцинил-КоА-дағы
макроэргиялық байланыс үзіліп
Сукцинат(янтарь
7. Субстраттан фосфорлану реакциясы.
Сукцинаттиокиназа әсерінен
Сукцинил-КоА-дағы
макроэргиялық байланыс үзіліп
Сукцинат(янтарь

Түзілген ГТФ нуклеозиддифосфат−
киназа көмегімен АТФ-ке
айналады:
ГТФ + АДФ → АТФ
киназа көмегімен АТФ-ке
айналады:
ГТФ + АДФ → АТФ
8. 3−ші тотығу-тотықсыздану реакциясы.
ФАД−тәуелді сукцинатдегидрогеназа(СДГ) әсерінен Сукцинаттан фумарат және тотықсызданған ФАДН2
8. 3−ші тотығу-тотықсыздану реакциясы.
ФАД−тәуелді сукцинатдегидрогеназа(СДГ) әсерінен Сукцинаттан фумарат және тотықсызданған ФАДН2

9.
Фумараза фумаратты
гидратациялап малат
түзіледі.
9.
Фумараза фумаратты
гидратациялап малат
түзіледі.

10.
4−ші тотығу-тотықсыздану реакциясы .
НАД−тәуелді малатдегидрогеназа (МДГ)
10.
4−ші тотығу-тотықсыздану реакциясы .
НАД−тәуелді малатдегидрогеназа (МДГ)
СН3
С=О
ζ
SКоА
Ацетил-КоА
2СО2+3НАДН2+ФАДН2 +АТФ (ГТФ)
СН3
С=О
ζ
SКоА
Ацетил-КоА
2СО2+3НАДН2+ФАДН2 +АТФ (ГТФ)

ДӘРІС N 8
БИОЛОГИЯЛЫҚ ТОТЫҒУ, ЭЛЕКТРОНДАРДЫҢ ТАСЫМАЛДАНУ ТІЗБЕГІ. ТОТЫҒУДАН ФОСФОРЛАНУ
ДӘРІС N 8
БИОЛОГИЯЛЫҚ ТОТЫҒУ, ЭЛЕКТРОНДАРДЫҢ ТАСЫМАЛДАНУ ТІЗБЕГІ. ТОТЫҒУДАН ФОСФОРЛАНУ

ДӘРІС ЖОСПАРЫ:
БИОЛОГИЯЛЫҚ ТОТЫҒУ, ЭЛЕКТРОНДАРДЫҢ ТАСЫМАЛДАНУ ТІЗБЕГІ
БИОЛОГИЯЛЫҚ ТОТЫҒУ САТЫЛАРЫ
ТОТЫҒУДАН ФОСФОРЛАНУ
Р\О КОЭФФИЦИЕНТІ ТУРАЛЫ
ДӘРІС ЖОСПАРЫ:
БИОЛОГИЯЛЫҚ ТОТЫҒУ, ЭЛЕКТРОНДАРДЫҢ ТАСЫМАЛДАНУ ТІЗБЕГІ
БИОЛОГИЯЛЫҚ ТОТЫҒУ САТЫЛАРЫ
ТОТЫҒУДАН ФОСФОРЛАНУ
Р\О КОЭФФИЦИЕНТІ ТУРАЛЫ

3 САТЫ. БИОЛОГИЯЛЫҚ ТОТЫҒУ (БТ )
БТ— ЖАСУША МИТОХОНДРИЯСЫНДА ӨТЕТІН ТОТЫҒУ–ТОТЫҚСЫЗДАНУ (Т-Т)
БТ— ЖАСУША МИТОХОНДРИЯСЫНДА ӨТЕТІН ТОТЫҒУ–ТОТЫҚСЫЗДАНУ (Т-Т)

ПФ(НАД) → (ФП (ФМН)→КоQ →Цхb → с1→с→ а→а3 →О2 .
ПФ(НАД) → (ФП (ФМН)→КоQ →Цхb → с1→с→ а→а3 →О2 .
БИОЛОГИЯЛЫҚ ТОТЫҒУ(БТ) МАҢЫЗЫ:
1. ЭНЕРГИЯ (Е) БӨЛІНЕДІ. (Е=40-48% АТФ
БИОЛОГИЯЛЫҚ ТОТЫҒУ(БТ) МАҢЫЗЫ:
1. ЭНЕРГИЯ (Е) БӨЛІНЕДІ. (Е=40-48% АТФ

БТ ОКСИДОРЕДУКТАЗАЛАР ҚАТЫСУЫМЕН МИТОХОНДРИЯДА ЖҮРЕДІ:
11 РЕАКЦИЯДАН ТҰРАДЫ.
БТ ОКСИДОРЕДУКТАЗАЛАР ҚАТЫСУЫМЕН МИТОХОНДРИЯДА ЖҮРЕДІ:
11 РЕАКЦИЯДАН ТҰРАДЫ.

БТ тізбегіндегі ферменттердің орналасу
тәртібі неге тәуелді?
Е , В
Н/ Н2
тәртібі неге тәуелді?
Е , В
Н/ Н2

Тотығатын зат әртүрлі зат алмасуында түзілген органикалық қышқылдар, негізінен ҮКЦ-де түзілген
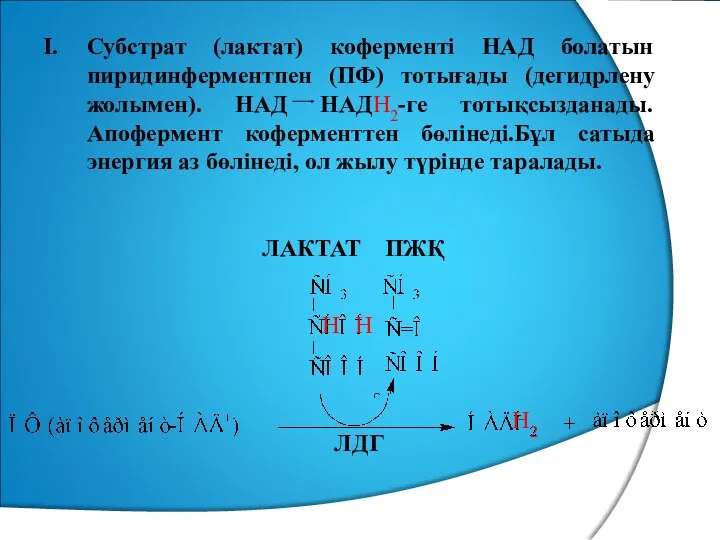
Субстрат (лактат) коферменті НАД болатын пиридинферментпен (ПФ) тотығады (дегидрлену жолымен). НАД
Субстрат (лактат) коферменті НАД болатын пиридинферментпен (ПФ) тотығады (дегидрлену жолымен). НАД
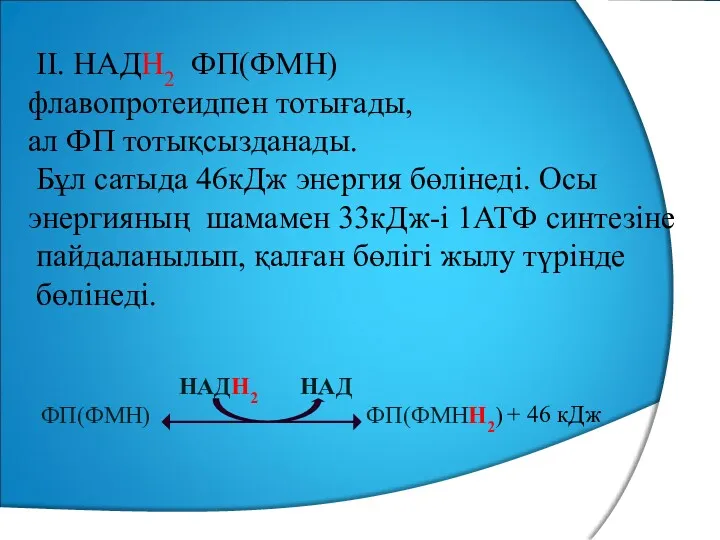
II. НАДН2 ФП(ФМН)
флавопротеидпен тотығады,
ал ФП тотықсызданады.
Бұл сатыда 46кДж энергия
флавопротеидпен тотығады,
ал ФП тотықсызданады.
Бұл сатыда 46кДж энергия
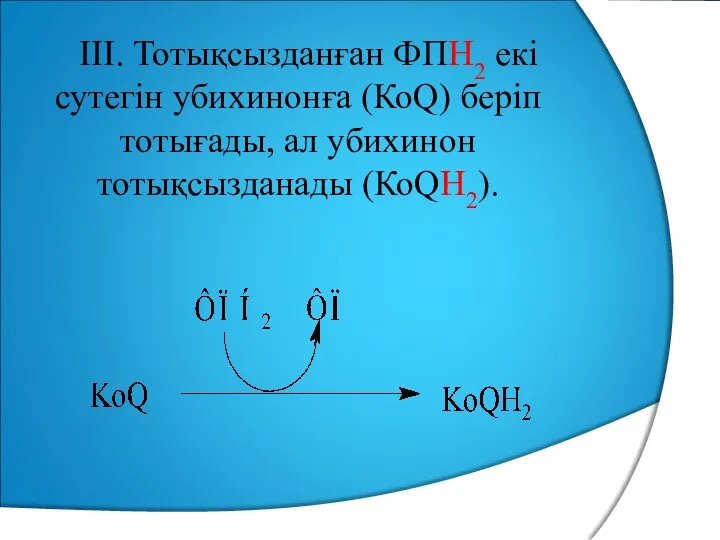
III. Тотықсызданған ФПН2 екі сутегін убихинонға (КоQ) беріп тотығады, ал убихинон
III. Тотықсызданған ФПН2 екі сутегін убихинонға (КоQ) беріп тотығады, ал убихинон
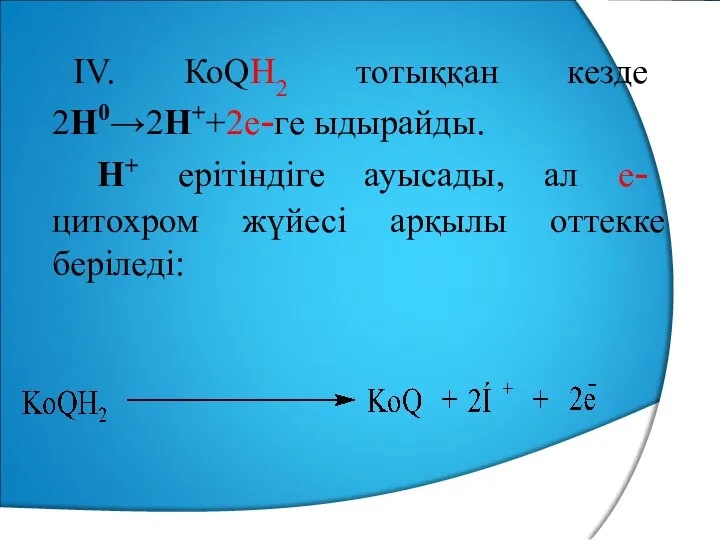
IV. КоQН2 тотыққан кезде 2Н0→2Н++2е-ге ыдырайды.
Н+ ерітіндіге ауысады, ал е-
Н+ ерітіндіге ауысады, ал е-
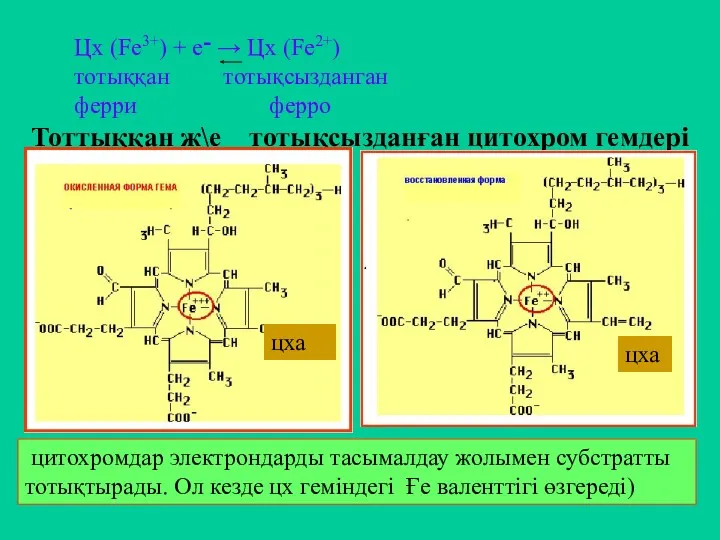
Тоттыққан ж\е тотықсызданған цитохром гемдері
цитохромдар электрондарды тасымалдау жолымен субстратты тотықтырады.
Тоттыққан ж\е тотықсызданған цитохром гемдері
цитохромдар электрондарды тасымалдау жолымен субстратты тотықтырады.

Убихиноннан бөлінген 2е-
2 ферри цхb-ға беріліп оны ферро формаға
Убихиноннан бөлінген 2е-
2 ферри цхb-ға беріліп оны ферро формаға

VI. 2ферро цхb 2e־ 2ферри цхс1-ге беріп тотығады, ал 2Цхс1 (Fe2+

VII. 2 ферро цх с1 2e־ 2ферри цхс-ға
беріп тотығады, ал
VII. 2 ферро цх с1 2e־ 2ферри цхс-ға
беріп тотығады, ал

VIII.
2ферроЦхс 2e־ 2ферри цха–ға беріп тотығады, ал ферри 2Цха
2ферроЦхс 2e־ 2ферри цха–ға беріп тотығады, ал ферри 2Цха

9- және 10-сатылар тығыз байланысты, себебі
цитохромоксидазды мультиферменттік
комплекс (2Цха+4Цха3 +6Сu) қатысады.
IX.
9- және 10-сатылар тығыз байланысты, себебі
цитохромоксидазды мультиферменттік
комплекс (2Цха+4Цха3 +6Сu) қатысады.
IX.

X.
2ферро Цха3 2е־ О2 беріп тотығады, ал
оттек ионданады(тотықсызданады).
О2
X.
2ферро Цха3 2е־ О2 беріп тотығады, ал
оттек ионданады(тотықсызданады).
О2

XI. Эндогенді судың түзілуі.
Ионданған оттек КоQН2 тотыққанда (IV пункт)
түзілген 2
Ионданған оттек КоQН2 тотыққанда (IV пункт)
түзілген 2

Қорытынды:
БТ-ның 3 нүктесінде (сатысында) АТФ түзуге жеткілікті энергия бөлінеді. Осы
БТ-ның 3 нүктесінде (сатысында) АТФ түзуге жеткілікті энергия бөлінеді. Осы

ЭНЕРГИЯ АЛМАСУЫНЫҢ 4 САТЫСЫ:
ТОТЫҒУДАН ФОСФОРЛАНУ (ТФ).
ТФ − БИОЛОГИЯЛЫҚ ТОТЫҒУ
ЭНЕРГИЯ АЛМАСУЫНЫҢ 4 САТЫСЫ:
ТОТЫҒУДАН ФОСФОРЛАНУ (ТФ).
ТФ − БИОЛОГИЯЛЫҚ ТОТЫҒУ
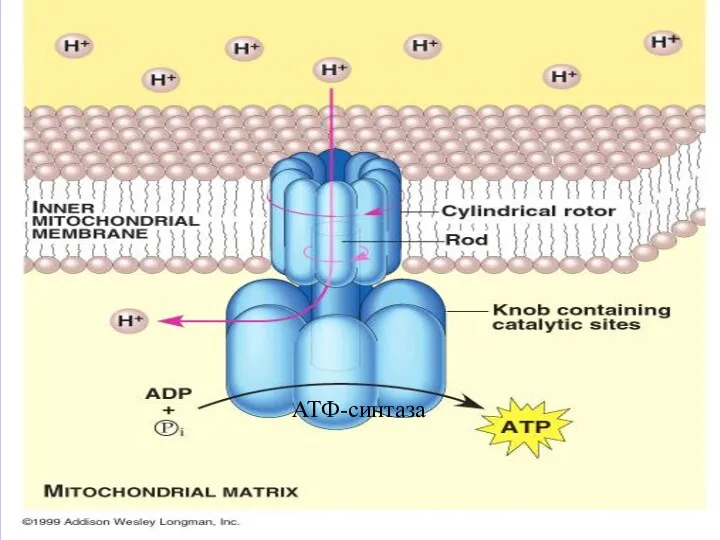
АТФ-синтаза
АТФ-синтаза

Р/О−ТОТЫҒУДАН ФОСФОРЛАНУ КОЭФФИЦИЕНТІ.
Р/О − АТФ־ ТІ ТҮЗУГЕ КЕТКЕН БЕЙОРГАНИКАЛЫҚ
Р/О−ТОТЫҒУДАН ФОСФОРЛАНУ КОЭФФИЦИЕНТІ.
Р/О − АТФ־ ТІ ТҮЗУГЕ КЕТКЕН БЕЙОРГАНИКАЛЫҚ

ЕГЕР СУБСТРАТ ПИРИДИН ФЕРМЕНТІ ПФ(НАД) АРҚЫЛЫ ТОТЫҚСА: Р/О = 3; себебі
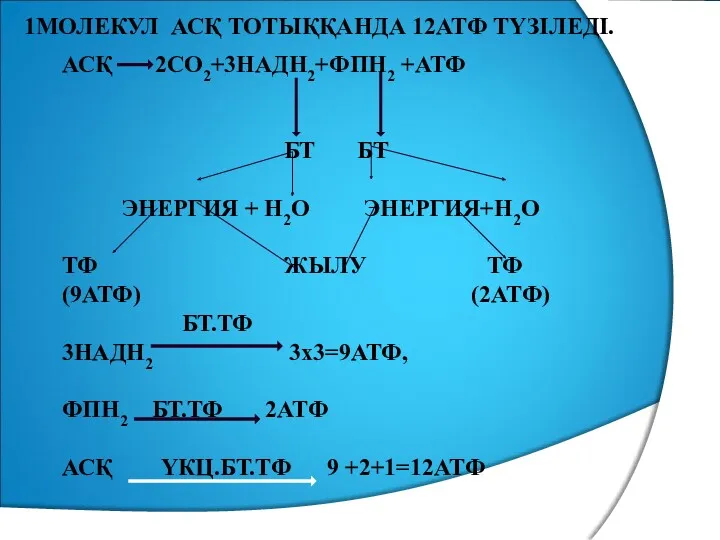
1МОЛЕКУЛ АСҚ ТОТЫҚҚАНДА 12АТФ ТҮЗІЛЕДІ.
АСҚ 2СО2+3НАДН2+ФПН2 +АТФ
БТ БТ
ЭНЕРГИЯ + Н2О
АСҚ 2СО2+3НАДН2+ФПН2 +АТФ
БТ БТ
ЭНЕРГИЯ + Н2О

Ферменттік ансамбль.
БТ тізбегіне қатысатын ферменттер сызықты
түрде орналаспайды, олар 4 комплекске
БТ тізбегіне қатысатын ферменттер сызықты
түрде орналаспайды, олар 4 комплекске
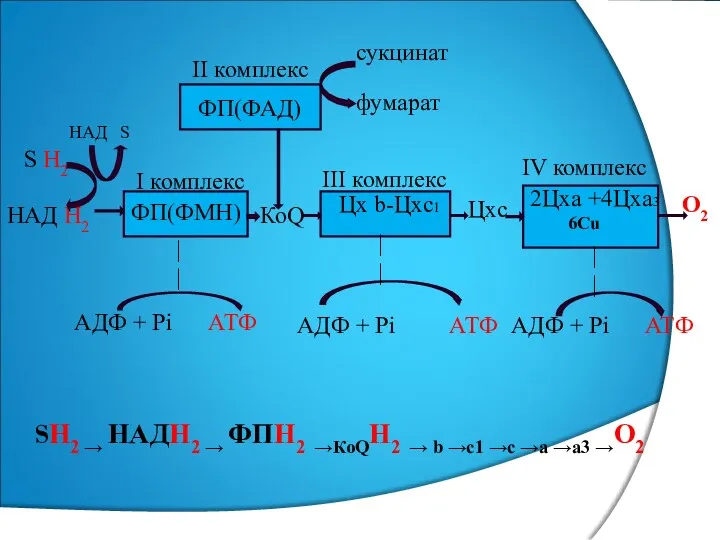
ФП(ФМН)
Цх b-Цхс1
2Цха +4Цха3
6Сu
ФП(ФАД)
S Н2
НАД Н2
НАД S
КоQ
Цхс
О2
II комплекс
I комплекс
III
ФП(ФМН)
Цх b-Цхс1
2Цха +4Цха3
6Сu
ФП(ФАД)
S Н2
НАД Н2
НАД S
КоQ
Цхс
О2
II комплекс
I комплекс
III

I ж/е II комплекс КоQ арқылы III –комплекспен байланысады,
ал III
I ж/е II комплекс КоQ арқылы III –комплекспен байланысады,
ал III

БТ МЕН ТФ ӨЗАРА ТЫҒЫЗ БАЙЛАНЫСТЫ.
БТ ЖЫЛДАМДЫҒЫН ТФ БАҚЫЛАЙДЫ, МҰНЫ ТЫНЫС
БТ МЕН ТФ ӨЗАРА ТЫҒЫЗ БАЙЛАНЫСТЫ.
БТ ЖЫЛДАМДЫҒЫН ТФ БАҚЫЛАЙДЫ, МҰНЫ ТЫНЫС

ЭНЕРГИЯ АЛМАСУЫ САТЫЛАРЫ БІР-БІРІМЕН ТЫҒЫЗ БАЙЛАНЫСТЫ.
1-2 САТЫСЫ: АСҚ АРҚЫЛЫ
ЭНЕРГИЯ АЛМАСУЫ САТЫЛАРЫ БІР-БІРІМЕН ТЫҒЫЗ БАЙЛАНЫСТЫ.
1-2 САТЫСЫ: АСҚ АРҚЫЛЫ

ГИПОЭНЕРГЕТИКАЛЫҚ КҮЙ ־АТФ СИНТЕЗІНІҢ
ТӨМЕНДЕУІ:
АШТЫҚ
О2
ЖАСУШАНЫ ОТТЕКПЕН ҚАМТАМАСЫЗ ЕТЕТІН
ЖҮРЕК־ҚАН ТАМЫРЛАРЫ
ГИПОЭНЕРГЕТИКАЛЫҚ КҮЙ ־АТФ СИНТЕЗІНІҢ
ТӨМЕНДЕУІ:
АШТЫҚ
О2
ЖАСУШАНЫ ОТТЕКПЕН ҚАМТАМАСЫЗ ЕТЕТІН
ЖҮРЕК־ҚАН ТАМЫРЛАРЫ
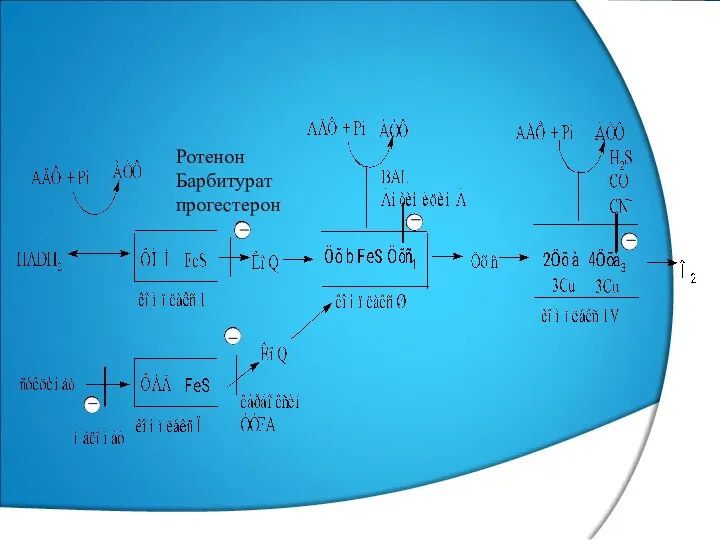
Ротенон
Барбитурат
прогестерон
Ротенон
Барбитурат
прогестерон
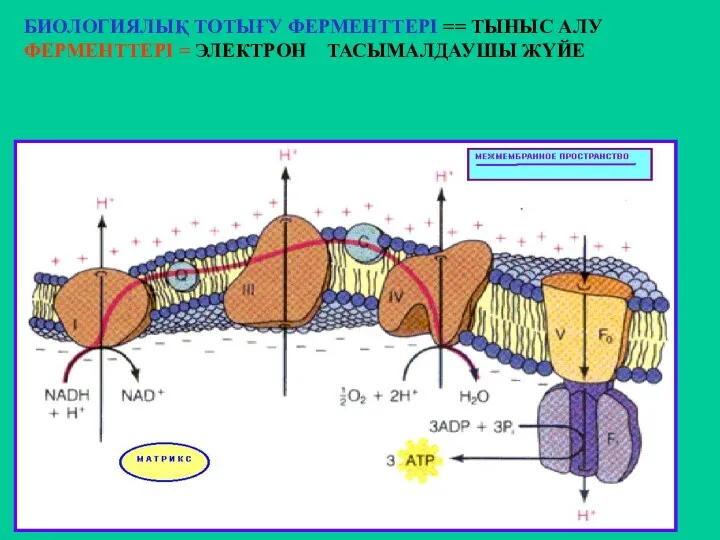
БИОЛОГИЯЛЫҚ ТОТЫҒУ ФЕРМЕНТТЕРІ == ТЫНЫС АЛУ ФЕРМЕНТТЕРІ = ЭЛЕКТРОН ТАСЫМАЛДАУШЫ ЖҮЙЕ
БИОЛОГИЯЛЫҚ ТОТЫҒУ ФЕРМЕНТТЕРІ == ТЫНЫС АЛУ ФЕРМЕНТТЕРІ = ЭЛЕКТРОН ТАСЫМАЛДАУШЫ ЖҮЙЕ
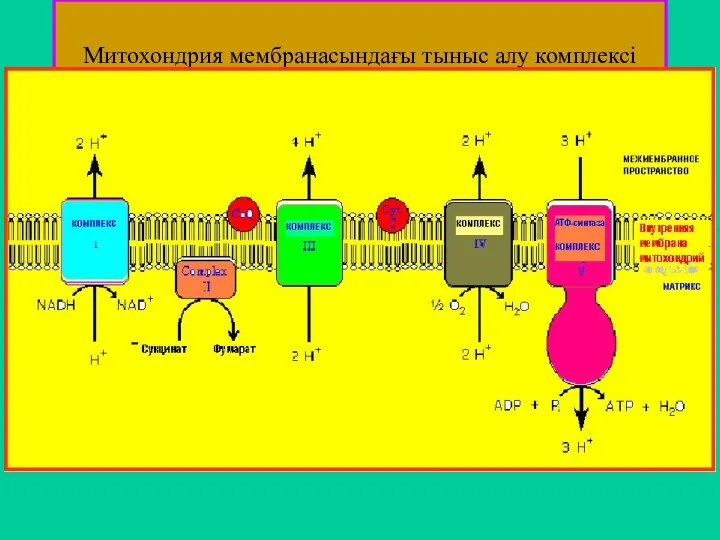
Митохондрия мембранасындағы тыныс алу комплексі
Митохондрия мембранасындағы тыныс алу комплексі

БТ ИНГИБИТОРЛАРЫ
НАДН-ДЕГИДРОГЕНАЗА ИНГИБИТОРЛАРЫ: БАРБИТУРАТТАР (ВЕНОНАЛ, ГЕКСЕНАЛ, НЕМБУТАЛ, АМИТАЛ);
ЦХв −с ИНГИБИТОРЛАРЫ- АНТИМИЦИН
БТ ИНГИБИТОРЛАРЫ
НАДН-ДЕГИДРОГЕНАЗА ИНГИБИТОРЛАРЫ: БАРБИТУРАТТАР (ВЕНОНАЛ, ГЕКСЕНАЛ, НЕМБУТАЛ, АМИТАЛ);
ЦХв −с ИНГИБИТОРЛАРЫ- АНТИМИЦИН

БТ АЖЫРАТҚЫШТАРЫ
КЕЙБІР ЛИПОФИЛЬДІ ПРОТОНОФОРЛАР МИТОХОНДРИЯ МЕМБРАНАСЫНЫҢ ӨТКІЗГІШТІГІН БҰЗЫП ПРОТОНДАРДЫ ПРОТОНДЫҚ
БТ АЖЫРАТҚЫШТАРЫ
КЕЙБІР ЛИПОФИЛЬДІ ПРОТОНОФОРЛАР МИТОХОНДРИЯ МЕМБРАНАСЫНЫҢ ӨТКІЗГІШТІГІН БҰЗЫП ПРОТОНДАРДЫ ПРОТОНДЫҚ

ЭКЗОГЕНДІ АЖЫРАТҚЫШТАР−2,4-ДИНИТРОФЕНОЛ, ДИКУМАРОЛ, СТРЕПТОМИЦИН;
ЭНДОГЕНДІ АЖЫРАТҚЫШТАР−МАЙ ҚЫШҚЫЛДАРЫ, ҚАЛҚАНША БЕЗІНІҢ ГОРМОНЫ־ ТИРОКСИН, ӨТ
ЭКЗОГЕНДІ АЖЫРАТҚЫШТАР−2,4-ДИНИТРОФЕНОЛ, ДИКУМАРОЛ, СТРЕПТОМИЦИН;
ЭНДОГЕНДІ АЖЫРАТҚЫШТАР−МАЙ ҚЫШҚЫЛДАРЫ, ҚАЛҚАНША БЕЗІНІҢ ГОРМОНЫ־ ТИРОКСИН, ӨТ

ӘДЕБИЕТТЕР:
Негізгі:
Т.Ш.Шарманов, С.М.Плешкова «Метаболические основы питания с курсом общей биохимии», Алматы,
ӘДЕБИЕТТЕР:
Негізгі:
Т.Ш.Шарманов, С.М.Плешкова «Метаболические основы питания с курсом общей биохимии», Алматы,

НАЗАРЛАРЫҢЫЗҒА
РАХМЕТ!
НАЗАРЛАРЫҢЫЗҒА
РАХМЕТ!

Цитохромная система
Существует несколько групп цитохромов –А,В, С, Д, которые являются гемопротеидами
Цитохромная система
Существует несколько групп цитохромов –А,В, С, Д, которые являются гемопротеидами
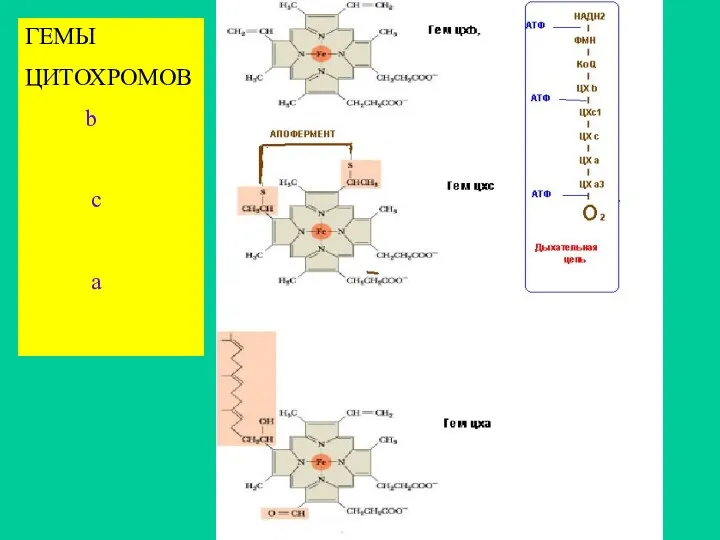
ГЕМЫ
ЦИТОХРОМОВ
b
c
a
ГЕМЫ
ЦИТОХРОМОВ
b
c
a
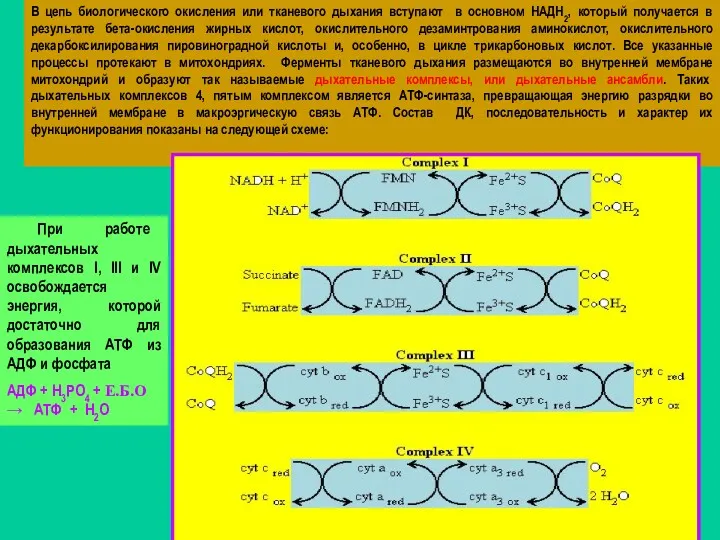
В цепь биологического окисления или тканевого дыхания вступают в основном
В цепь биологического окисления или тканевого дыхания вступают в основном
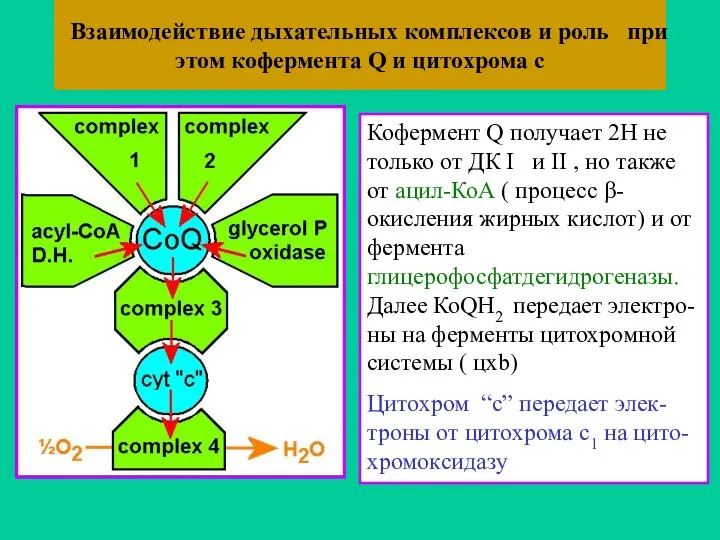
Взаимодействие дыхательных комплексов и роль при этом кофермента Q и цитохрома
Взаимодействие дыхательных комплексов и роль при этом кофермента Q и цитохрома
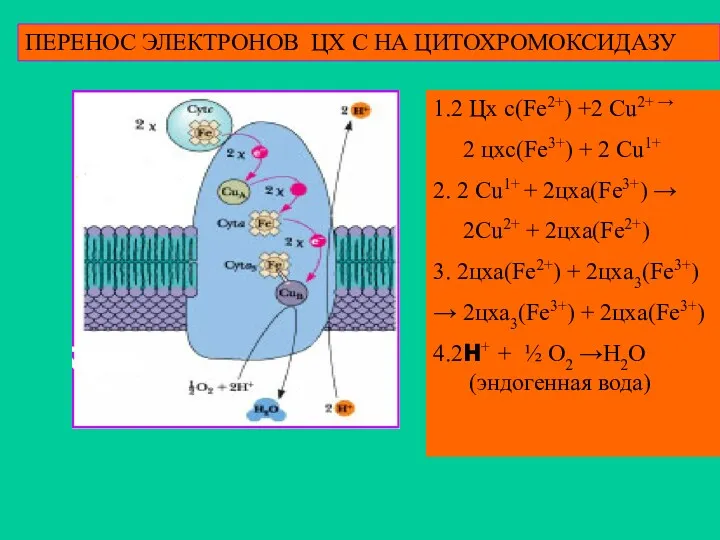
ПЕРЕНОС ЭЛЕКТРОНОВ ЦХ С НА ЦИТОХРОМОКСИДАЗУ
1.2 Цх с(Fe2+) +2 Cu2+ →
ПЕРЕНОС ЭЛЕКТРОНОВ ЦХ С НА ЦИТОХРОМОКСИДАЗУ
1.2 Цх с(Fe2+) +2 Cu2+ →
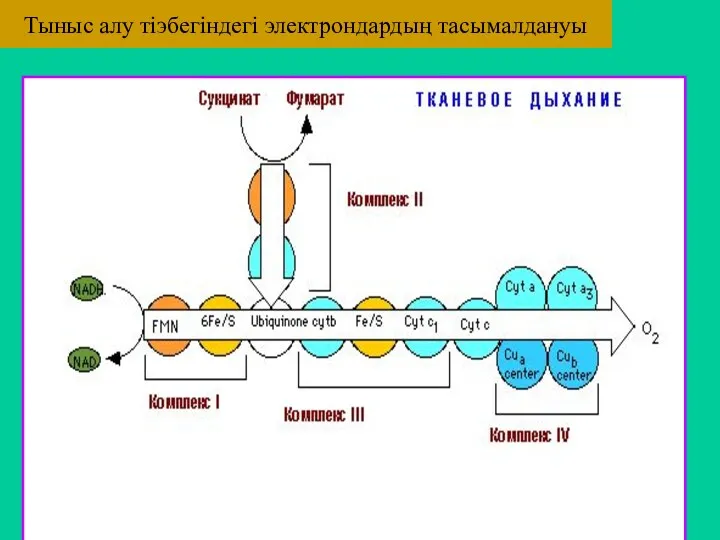
Тыныс алу тіэбегіндегі электрондардың тасымалдануы
Тыныс алу тіэбегіндегі электрондардың тасымалдануы
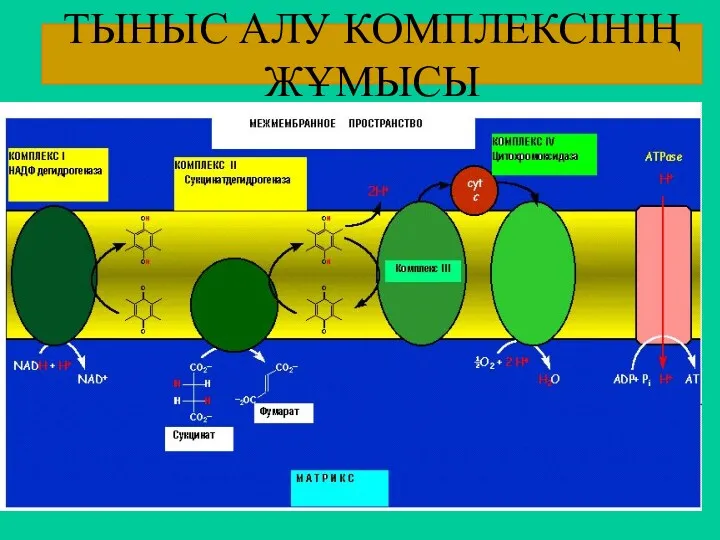
ТЫНЫС АЛУ КОМПЛЕКСІНІҢ ЖҰМЫСЫ
ТЫНЫС АЛУ КОМПЛЕКСІНІҢ ЖҰМЫСЫ
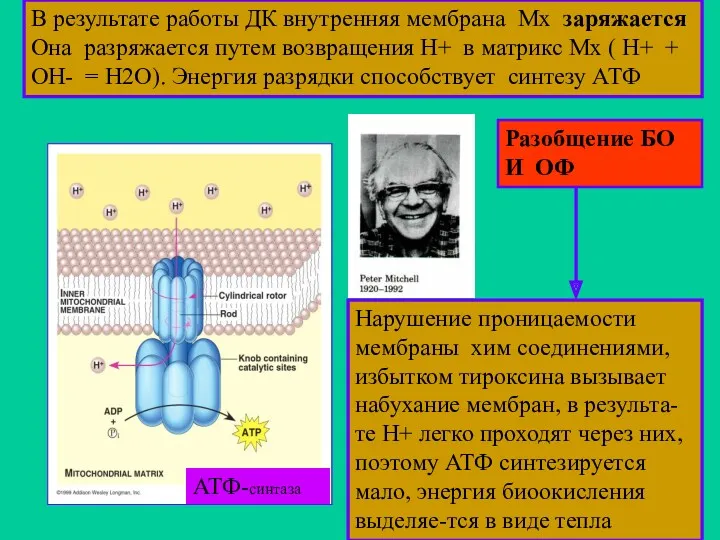
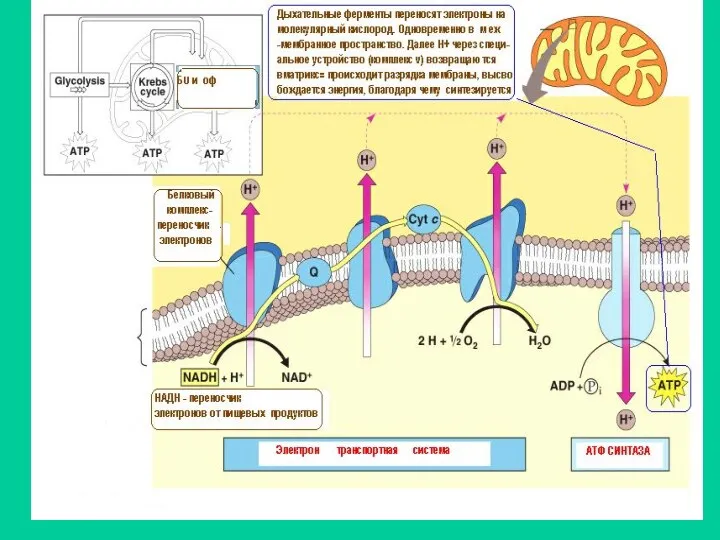
В результате работы ДК внутренняя мембрана Мх заряжается Она разряжается путем
В результате работы ДК внутренняя мембрана Мх заряжается Она разряжается путем

АТФ-СИНТАЗА –КОМПЛЕКС V
Комплекс, катализирующий образование АТФ=окислительное фосфорилирование, называется АТФ синтазой,
АТФ-СИНТАЗА –КОМПЛЕКС V
Комплекс, катализирующий образование АТФ=окислительное фосфорилирование, называется АТФ синтазой,
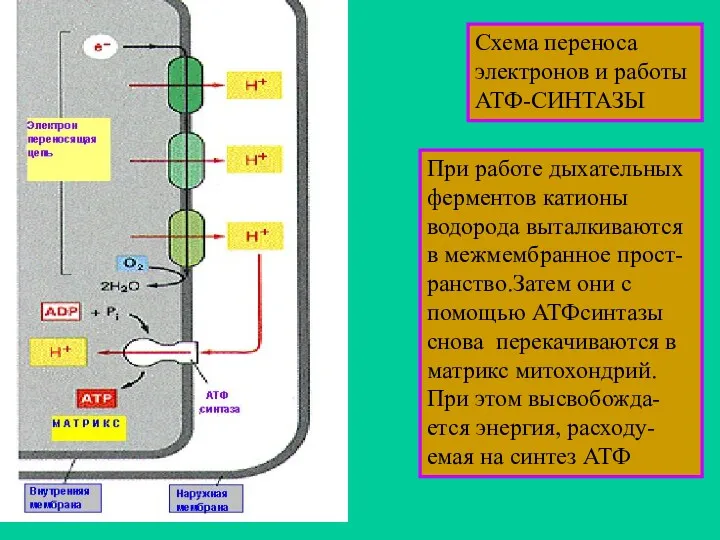
Схема переноса электронов и работы АТФ-СИНТАЗЫ
При работе дыхательных ферментов катионы
Схема переноса электронов и работы АТФ-СИНТАЗЫ
При работе дыхательных ферментов катионы

ХЕМООСМОТИЧЕСКОЕ СОПРЯЖЕНИЕ
ХЕМООСМОТИЧЕСКОЕ СОПРЯЖЕНИЕ
 Characteristic of families Rosaceae, Fabaceae, Linaceae, typical species and practical uses
Characteristic of families Rosaceae, Fabaceae, Linaceae, typical species and practical uses Соцветия
Соцветия Эволюция нервной системы
Эволюция нервной системы Осевой скелет
Осевой скелет Значення рослин для життя на планеті Земля
Значення рослин для життя на планеті Земля Хим.состав клетки
Хим.состав клетки Разработка урока на тему Организм и окружающая среда 11 класс
Разработка урока на тему Организм и окружающая среда 11 класс Стратегия, принципы и цели устойчивого развития
Стратегия, принципы и цели устойчивого развития Крупные пищеварительные железы. Печень, поджелудочная железа, желчный пузырь
Крупные пищеварительные железы. Печень, поджелудочная железа, желчный пузырь Членистоногие
Членистоногие Растения, которые нас лечат
Растения, которые нас лечат Взаимоотношения человека и природы, как фактор развития биосферы
Взаимоотношения человека и природы, как фактор развития биосферы Эволюция микроорганизмов
Эволюция микроорганизмов Как правильно построить научный эксперимент
Как правильно построить научный эксперимент Тип Круглые или Первичнополостные черви
Тип Круглые или Первичнополостные черви Почвоведение. Введение
Почвоведение. Введение Культуры эукариотических клеток
Культуры эукариотических клеток Открытый урок в 6 классеПоловое размножение растений
Открытый урок в 6 классеПоловое размножение растений Презентация Выделительная система
Презентация Выделительная система Ягоды. Кроссворд
Ягоды. Кроссворд Свойства ферментов. Механизм действия. Лекция 2
Свойства ферментов. Механизм действия. Лекция 2 Эволюция растительного мира на Земле. 6-7 классы
Эволюция растительного мира на Земле. 6-7 классы Животный мир. Тигр
Животный мир. Тигр Тип Членистоногие
Тип Членистоногие Биологическая роль и строение нуклеиновых кислот
Биологическая роль и строение нуклеиновых кислот Уход и содержание хомячков
Уход и содержание хомячков презентация по биологии Многообразие и значение корней
презентация по биологии Многообразие и значение корней Фитогормоны - гормоны растений
Фитогормоны - гормоны растений