- Детерминация пола у животных

Содержание
- 2. Первичные половые клетки (ППК) у млекопитающих, производные клеток внутренней массы, выявляются в пограничном районе желточного мешка
- 3. Маркёрными белками ППК млекопитающих являются фактор транскрипции Oct4 и щелочная фосфатаза
- 4. ППК дрозофилы обособляются в районе полярных гранул (на заднем конце зародыша) в ходе 9-го деления ядер
- 5. Миграция ППК дрозофилы из района полярной плазмы в районы латеральной мезодермы 10-12 парасегментов, где образуются гонады.
- 6. Регуляция пола может осуществляться у животных различным образом благодаря генетическим факторам или (и) факторам внешней среды
- 7. Хромосомное определение пола. У животных существуют половые хромосомы, так что пол определяется в зависимости от числа
- 11. У рыб наряду с раздельнополыми видами есть и гермафродиты. Гермафродиты могут быть : 1. Синхронные (одновременно
- 12. Изменения структуры гонад гермафродитной рыбы Sparus auratus
- 13. Наряду с генетическим факторами (наличие половых хромосом того или иного типа) на формирование пола у рыб,
- 14. Корреляция между сотношением полов F/ (F+M) и температурой у рыбы Menidia menidia появляется и нарастает в
- 15. Зависящее от температуры (инкубации яиц) определение пола у трех представителей класса пресмыкающихся: грифовая и красноухая черепахи
- 16. Температуро-зависимая детерминация пола у ящериц, черепах и крокодилов
- 17. Виды гормонов мужских и женских половых желез млекопитающих
- 18. Факторы внешней среды, определяющие формирование пола у позвоночных (пресмыкающихся), могут действовать, через изменение баланса половых гормонов.
- 19. Синтез стероидных половых гормонов у позвоночных
- 20. У эхиуриды Bonellia (организма, обитающего на морском каменистом грунте) формирование пола происходит у личинки в зависимости
- 21. У улитки- блюдечка Crepidula fornicata, новая особь, располагающаяся в колонии сверху - всегда самец. Затем мужская
- 22. Первичная и вторичная детерминация пола у млекопитающих Первичная детерминация пола – это детерминация гонад. Семенники и
- 23. Дифференцировка бипотенциальных гонад и образование половых структур из Вольфова и Мюллерова протоков
- 24. Дифференцировка гонад человека (начало)
- 25. Дифференцировка гонад человека (продолжение)
- 26. Дифференцировка гонад человека (окончание)
- 27. При анализе Y-хромосомы выяснилось, что гипотетический фактор, отвечающий за формирование пола, существует в действительности- это ген
- 29. Мышь с половым генотипом XX, трансгенная по гену SRY(справа), в норме локализованным на Y-хромосоме обладает фенотипом
- 30. Экспрессия гена Sry приводит к активации генов SF1 и Sox9 в тех же клетках. Взаимодействие генов
- 31. Роль антимюллерова гормона в дегенерации мюллерова протока (плод крысы). Слева - открыты оба протока, справа -через
- 32. Выявление гена DAX1 на X-хромосоме, ответственного за формирование женского пола. Ген был идентифицирован на X-хромосоме (в
- 33. Стероидогенный фактор SF1 в зачатках семенника активируется под контролем SRY и совместно с SOX9 активирует экспрессию
- 34. Каскады регуляции, паракринными и транскрипционными факторами, ведущие к образованию альтернативных половых фенотипов млекопитающих
- 35. Тестостерон и 5 α-дигидротестостерон - андрогены. Тестостерон синтезируется в клетках Лейдига и отвечает за образование эпидидимиса,
- 36. Районы мужской половой системы, формирующиеся под контролем тестостерона и дигидротестостерона.
- 37. Мужские и женские стероидные гормоны. Эстрогены (включая диэтилстилбестрол) важны не только для формирования женских половых органов
- 38. Виды женских половых гормонов и их функции
- 39. Хромосомная детерминация пола у Drosophila Особи генотипа XX -самки и генотипа XY -самцы. В случае генотипа
- 40. Гинандроморфы дрозофилы и мотылька, образовавшиеся в результате утраты в раннем развитии из ряда клеток одной их
- 41. Гены нумераторы (числителя), локализованные на X-хромосоме, кодируют факторы транскрипции sisterless-a, sisterless-b=scute (sis-a,sis-b), runt (runt), активирующие транскрипцию
- 42. Схематическое представление генетической программы детерминации пола Drosophila, определяемое соотношением X:A. Детерминация пола по мужскому или женскому
- 43. Каскады регуляций, контролирующих формирование половых структур самки и самца дрозофилы Различная активация гена sxl (sex-lethal) будущих
- 44. Различная активация гена sxl будущих самок и самцов дрозофилы. Деятельность генов нумераторов и деноминаторов определяет альтернативные
- 45. Альтернативный сплайсинг -основной путь реализации программы детерминации пола у дрозофилы (последовательность событий)
- 46. По ходу развития, когда sxl начинает транскрибироваться с позднего (дистального) промотора, альтернативный сплайсинг sxl при генотипе
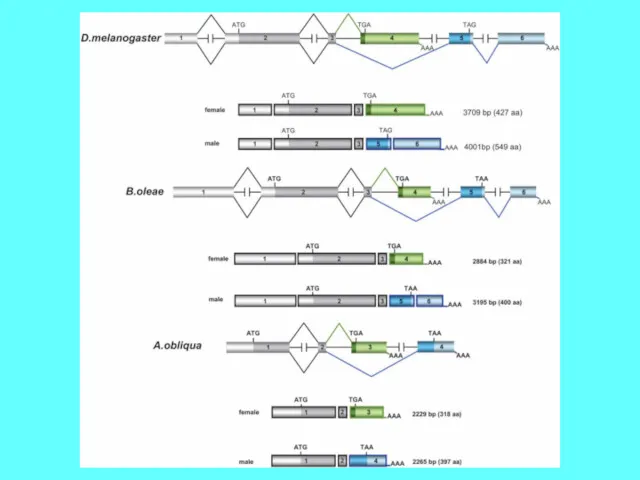
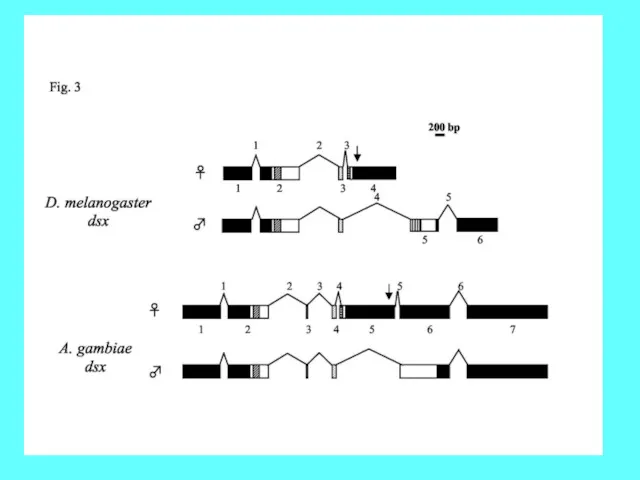
- 47. Формирование мужской и женской половых гонад у дрозофилы под контролем различных форм Dsx
- 48. Роли DsxM и DsxF в формировании пола у дрозофилы. Dsx- транскрипционный фактор, действующий как репрессор или
- 49. Нематода Caenorhabditis elegans имеет два пола: XX -гермафродиты (более 99% популяции) и X0 -самцы. У взрослой
- 50. Нематода Сaenorhabditis elegans, гермафродитная особь
- 51. Нематода Сaenorhabditis elegans, самец
- 52. В случае самооплодотворения 0,2% новых особей -самцы, остальные -гермафродиты, в случае спаривания особей XX и X0
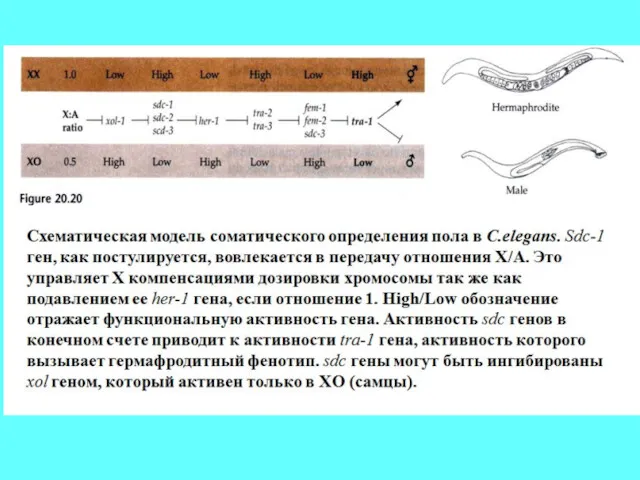
- 53. Детерминация пола у С. Elegans (также как и у Drosophila имеет значение отношение числа X-хромосом к
- 55. Сигнальная цепочка детерминации пола C. elegans. Перемещение в ядро транскрипционного фактора TRA-1 означает реализацию гермафродитного фенотипа.
- 56. Детерминация первичных половых клеток у гермафородитов по мужскому или женскому пути осуществляется в два этапа: 1этап
- 57. Детерминация первичных половых клеток по мужскому или женскому пути у гермафордитов осуществляется в два этапа: 2
- 58. Образование градиента Hunchback в бластодерме дрозофилы . Контроль трансляции мРНК hunchback белками Nanos и Pumilio
- 59. Эволюционные связи между генами, ответственными за детерминацию пола у разных животных В то время как Sry
- 60. Сколько раз в эволюции возникали механизмы детерминации пола?
- 61. Сколько раз в эволюции возникали механизмы детерминации пола? Гипотеза: Cуществуют две группы позвоночных (начиная с костистых
- 62. Cуществуют две группы позвоночных (начиная с костистых рыб), у одной группы –функционально активен ген –Sox9, у
- 63. Cуществуют две группы позвоночных (начиная с костистых рыб), у одной группы –функционально активен ген –Sox9, у
- 64. На 9-й хромосоме человека (короткое плечо) было идентифицировано 3 гена Dmrt (1-3), родственные генам Dsx (дрозофилы)
- 65. Ортологи генов Sox 9 активно экспрессируются в клетках Сертоли у птиц и пресмыкающихся (аллигаторы, ящерицы). Экспрессирующийся
- 66. Хотя млекопитающие, дрозофила и C. elegans имеют различные механизмы детерминации пола (основанные на различных способах передачи
- 67. Возможное участие гена DMRT в формировании семенников и сперматогенезе
- 68. Возможность существования консервативных факторов детерминации пола у разных животных
- 74. Скачать презентацию
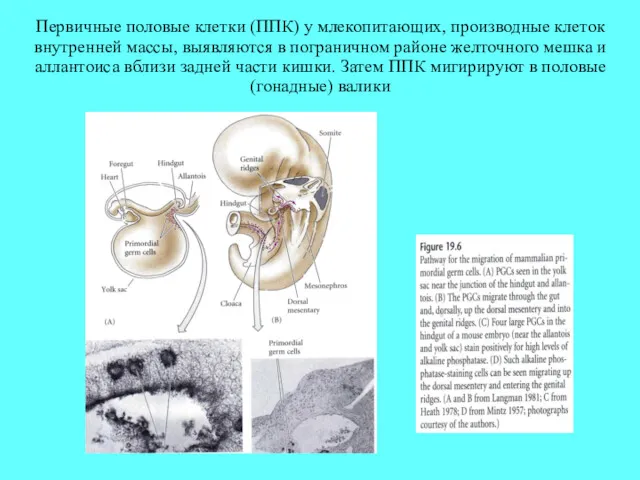
Первичные половые клетки (ППК) у млекопитающих, производные клеток внутренней массы, выявляются
Первичные половые клетки (ППК) у млекопитающих, производные клеток внутренней массы, выявляются
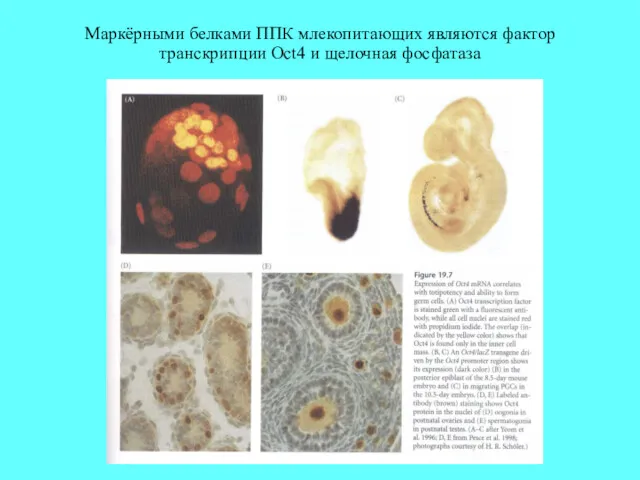
Маркёрными белками ППК млекопитающих являются фактор транскрипции Oct4 и щелочная фосфатаза
Маркёрными белками ППК млекопитающих являются фактор транскрипции Oct4 и щелочная фосфатаза
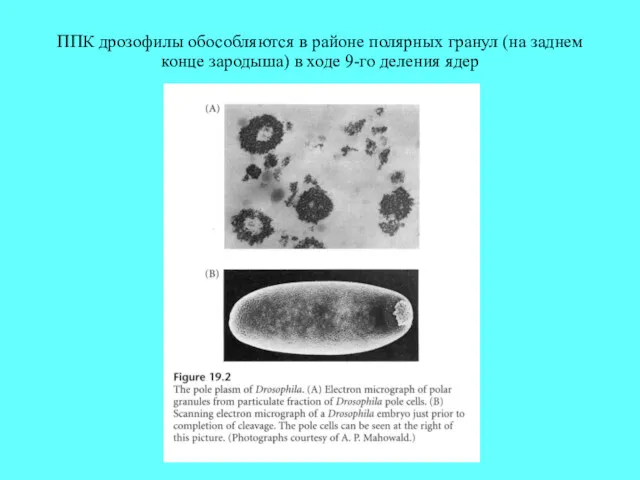
ППК дрозофилы обособляются в районе полярных гранул (на заднем конце зародыша)
ППК дрозофилы обособляются в районе полярных гранул (на заднем конце зародыша)
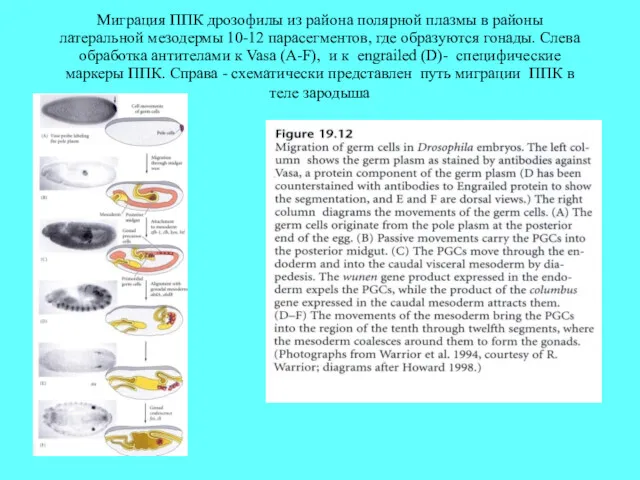
Миграция ППК дрозофилы из района полярной плазмы в районы латеральной мезодермы
Миграция ППК дрозофилы из района полярной плазмы в районы латеральной мезодермы

Регуляция пола может осуществляться у животных различным образом благодаря генетическим факторам
Регуляция пола может осуществляться у животных различным образом благодаря генетическим факторам

Хромосомное определение пола.
У животных существуют половые хромосомы, так что пол определяется
Хромосомное определение пола. У животных существуют половые хромосомы, так что пол определяется


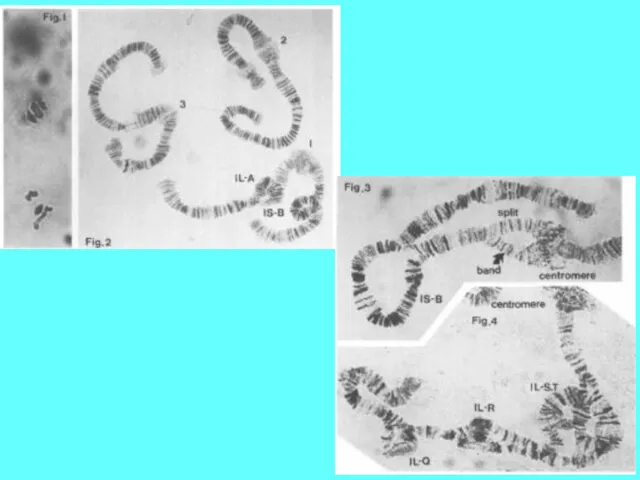

У рыб наряду с раздельнополыми видами есть и гермафродиты.
Гермафродиты могут быть
У рыб наряду с раздельнополыми видами есть и гермафродиты. Гермафродиты могут быть
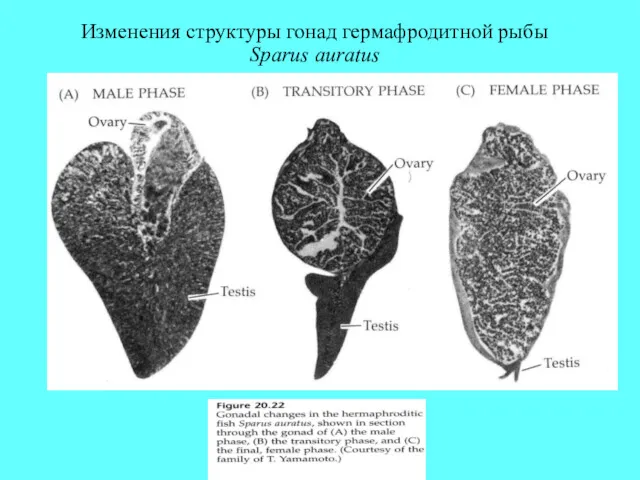
Изменения структуры гонад гермафродитной рыбы Sparus auratus
Изменения структуры гонад гермафродитной рыбы Sparus auratus

Наряду с генетическим факторами (наличие половых хромосом того или иного типа)
Наряду с генетическим факторами (наличие половых хромосом того или иного типа)
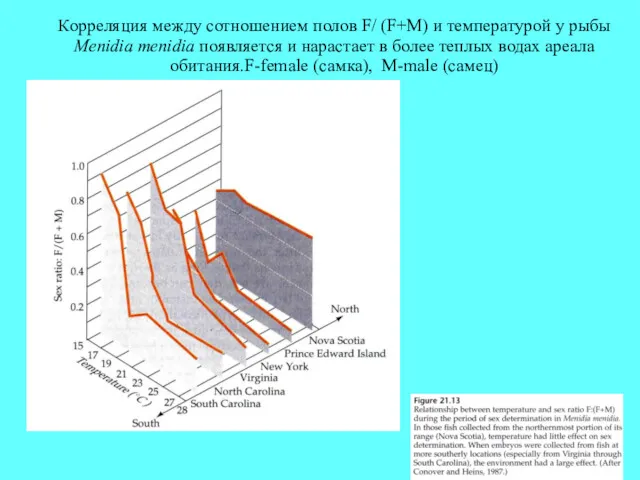
Корреляция между сотношением полов F/ (F+M) и температурой у рыбы Menidia
Корреляция между сотношением полов F/ (F+M) и температурой у рыбы Menidia
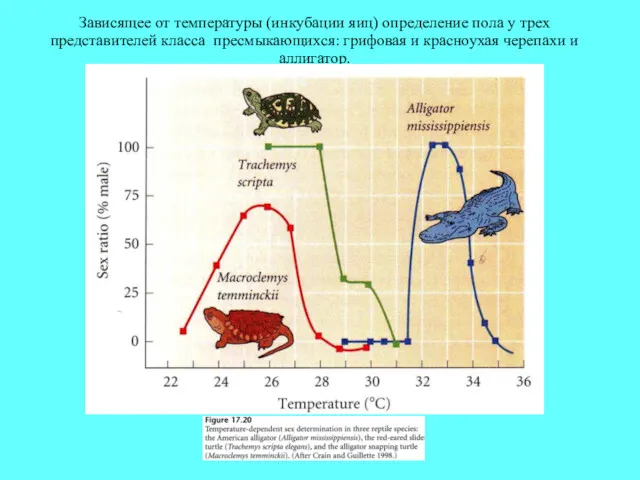
Зависящее от температуры (инкубации яиц) определение пола у трех представителей класса
Зависящее от температуры (инкубации яиц) определение пола у трех представителей класса
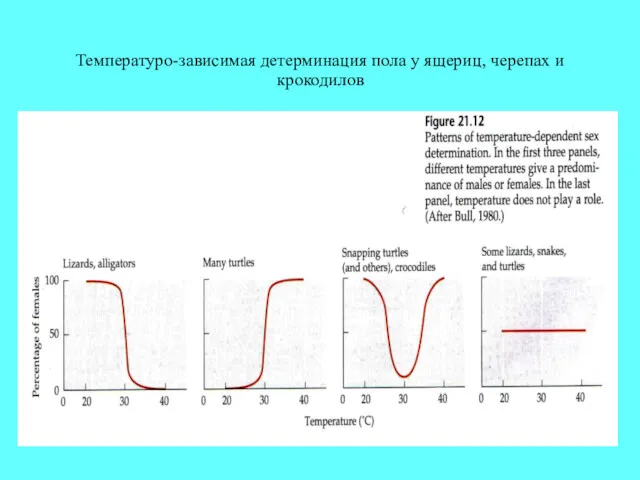
Температуро-зависимая детерминация пола у ящериц, черепах и крокодилов
Температуро-зависимая детерминация пола у ящериц, черепах и крокодилов

Виды гормонов мужских и женских половых желез млекопитающих
Виды гормонов мужских и женских половых желез млекопитающих

Факторы внешней среды, определяющие формирование пола у позвоночных (пресмыкающихся), могут действовать,
Факторы внешней среды, определяющие формирование пола у позвоночных (пресмыкающихся), могут действовать,
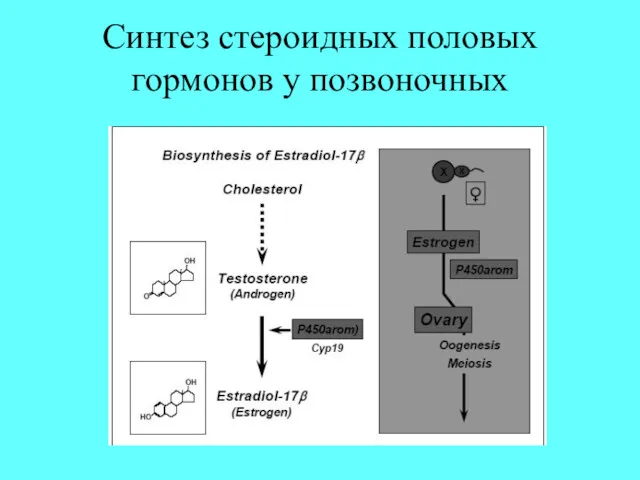
Синтез стероидных половых гормонов у позвоночных
Синтез стероидных половых гормонов у позвоночных
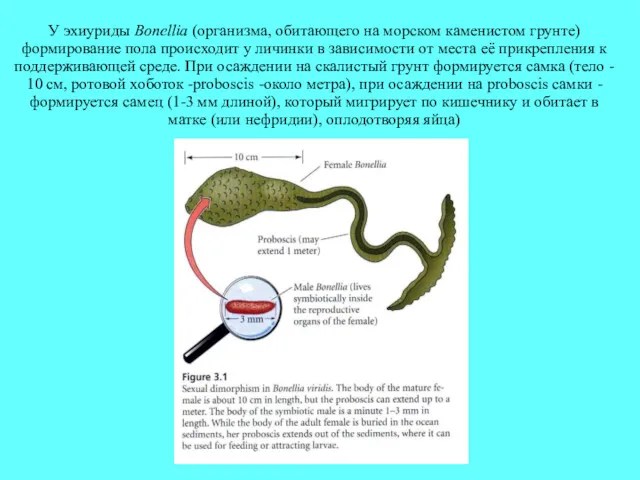
У эхиуриды Bonellia (организма, обитающего на морском каменистом грунте) формирование пола
У эхиуриды Bonellia (организма, обитающего на морском каменистом грунте) формирование пола
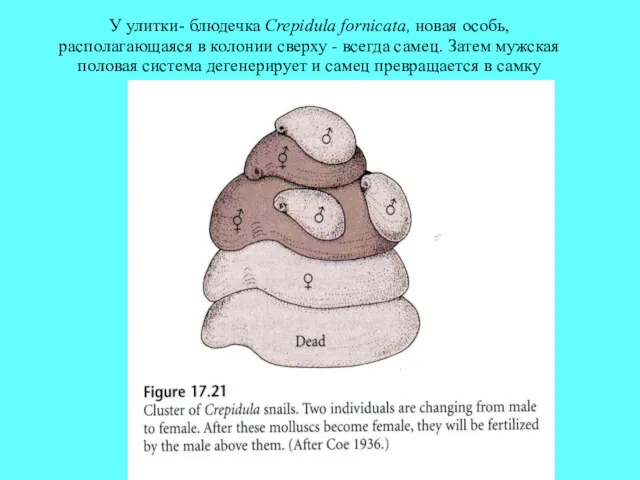
У улитки- блюдечка Crepidula fornicata, новая особь, располагающаяся в колонии сверху
У улитки- блюдечка Crepidula fornicata, новая особь, располагающаяся в колонии сверху

Первичная и вторичная детерминация пола у млекопитающих
Первичная детерминация пола – это
Первичная и вторичная детерминация пола у млекопитающих Первичная детерминация пола – это
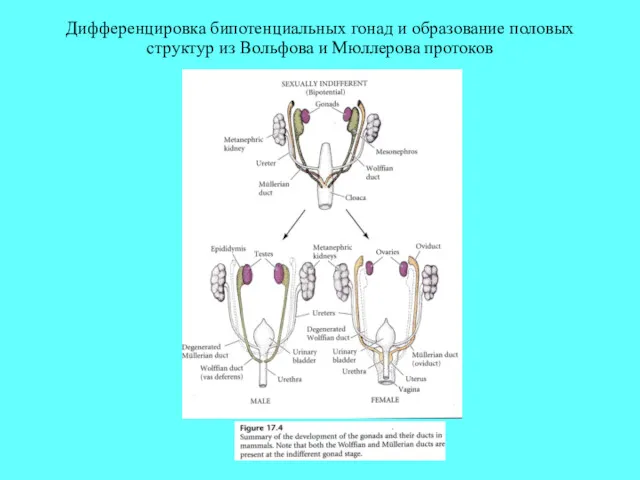
Дифференцировка бипотенциальных гонад и образование половых структур из Вольфова и Мюллерова
Дифференцировка бипотенциальных гонад и образование половых структур из Вольфова и Мюллерова
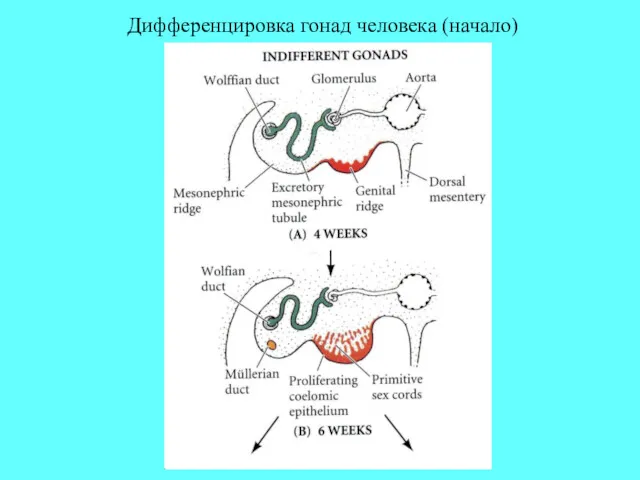
Дифференцировка гонад человека (начало)
Дифференцировка гонад человека (начало)
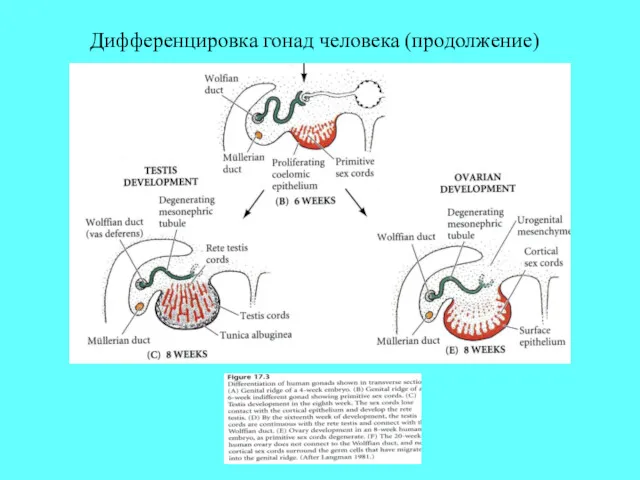
Дифференцировка гонад человека (продолжение)
Дифференцировка гонад человека (продолжение)
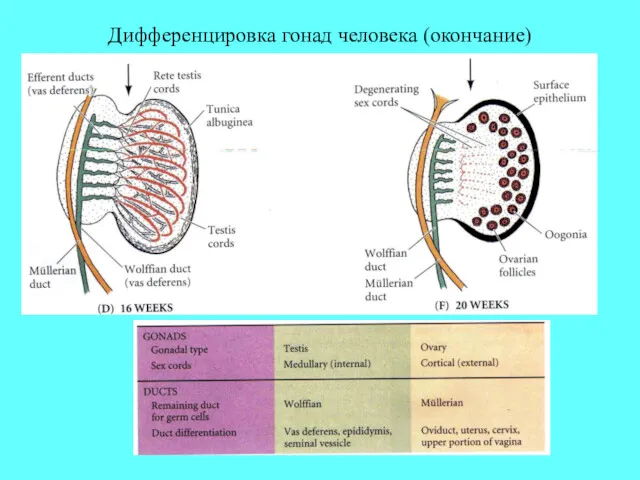
Дифференцировка гонад человека (окончание)
Дифференцировка гонад человека (окончание)

При анализе Y-хромосомы выяснилось, что гипотетический фактор, отвечающий за формирование пола,
При анализе Y-хромосомы выяснилось, что гипотетический фактор, отвечающий за формирование пола,

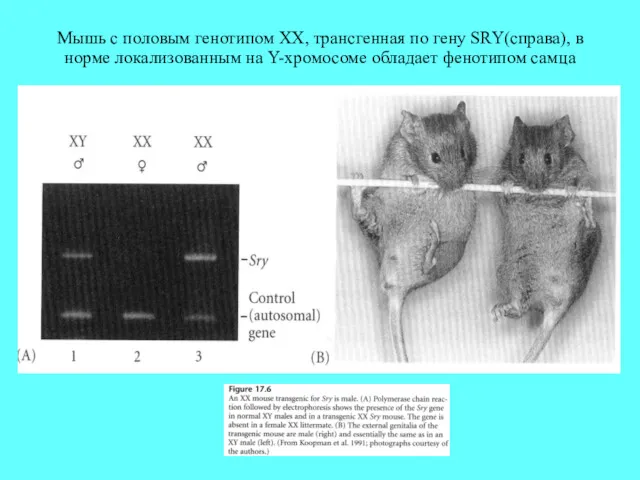
Мышь с половым генотипом XX, трансгенная по гену SRY(справа), в норме
Мышь с половым генотипом XX, трансгенная по гену SRY(справа), в норме
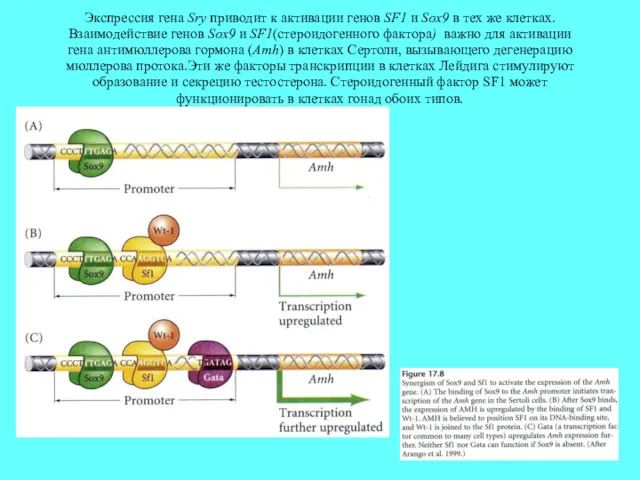
Экспрессия гена Sry приводит к активации генов SF1 и Sox9 в
Экспрессия гена Sry приводит к активации генов SF1 и Sox9 в
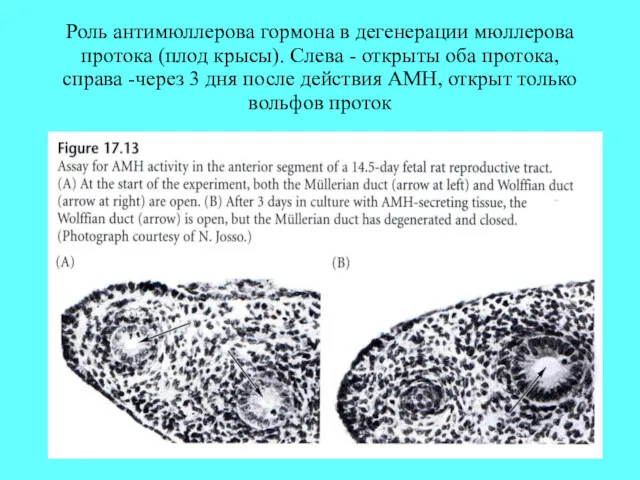
Роль антимюллерова гормона в дегенерации мюллерова протока (плод крысы). Слева -
Роль антимюллерова гормона в дегенерации мюллерова протока (плод крысы). Слева -
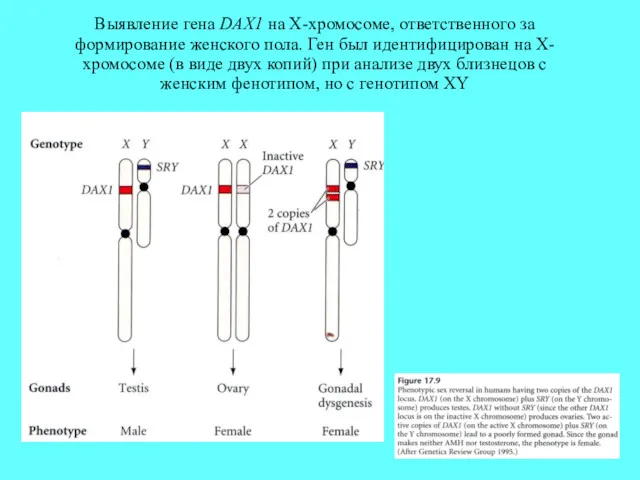
Выявление гена DAX1 на X-хромосоме, ответственного за формирование женского пола. Ген
Выявление гена DAX1 на X-хромосоме, ответственного за формирование женского пола. Ген
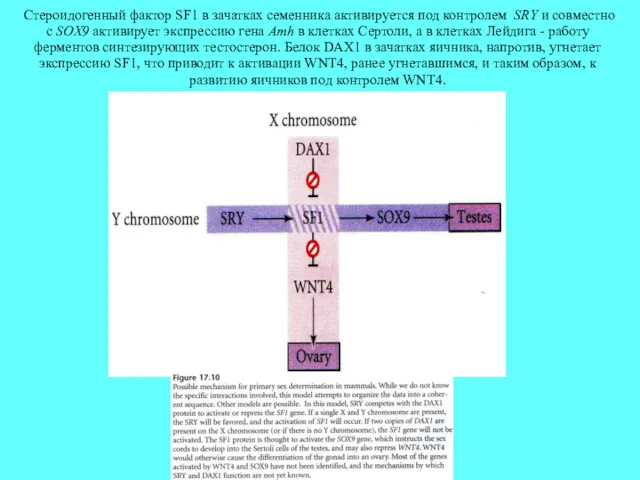
Стероидогенный фактор SF1 в зачатках семенника активируется под контролем SRY и
Стероидогенный фактор SF1 в зачатках семенника активируется под контролем SRY и
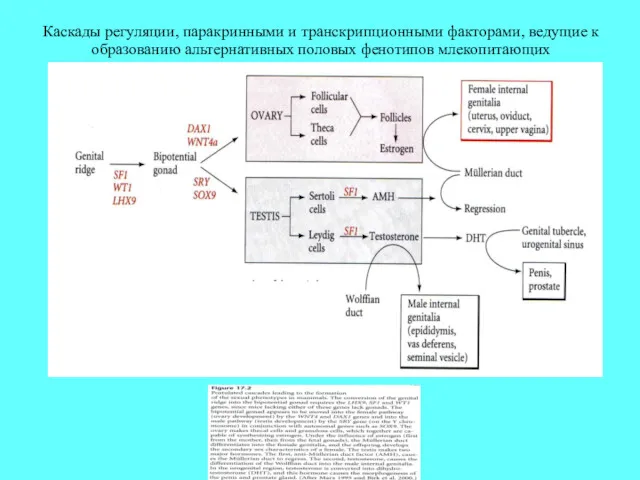
Каскады регуляции, паракринными и транскрипционными факторами, ведущие к образованию альтернативных половых
Каскады регуляции, паракринными и транскрипционными факторами, ведущие к образованию альтернативных половых

Тестостерон и 5 α-дигидротестостерон - андрогены. Тестостерон синтезируется в клетках Лейдига
Тестостерон и 5 α-дигидротестостерон - андрогены. Тестостерон синтезируется в клетках Лейдига
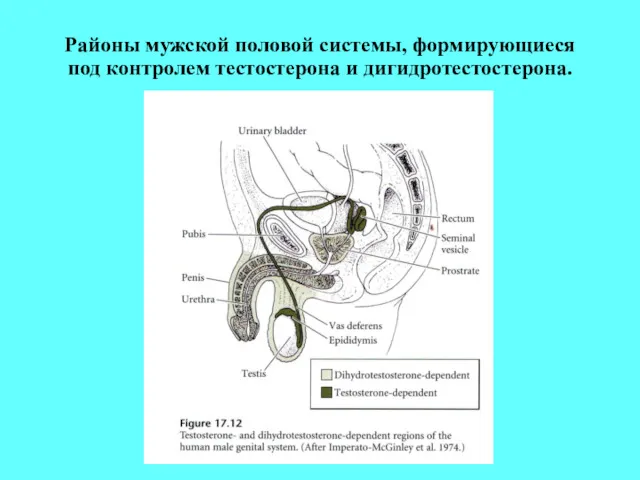
Районы мужской половой системы, формирующиеся под контролем тестостерона и дигидротестостерона.
Районы мужской половой системы, формирующиеся под контролем тестостерона и дигидротестостерона.

Мужские и женские стероидные гормоны. Эстрогены (включая диэтилстилбестрол) важны не только
Мужские и женские стероидные гормоны. Эстрогены (включая диэтилстилбестрол) важны не только

Виды женских половых гормонов и их функции
Виды женских половых гормонов и их функции

Хромосомная детерминация пола у Drosophila
Особи генотипа XX -самки и генотипа XY
Хромосомная детерминация пола у Drosophila Особи генотипа XX -самки и генотипа XY
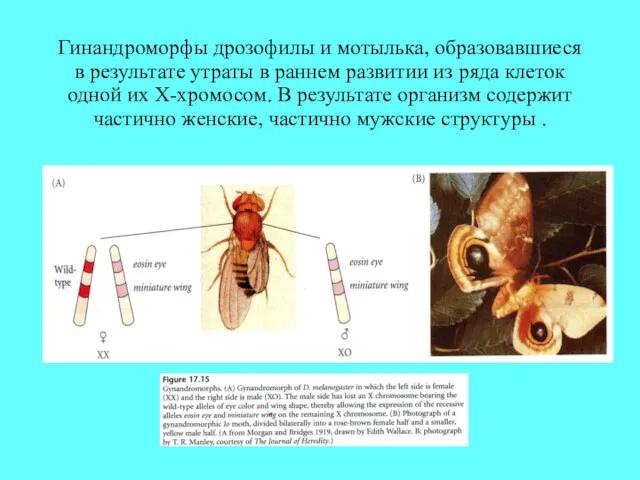
Гинандроморфы дрозофилы и мотылька, образовавшиеся в результате утраты в раннем развитии
Гинандроморфы дрозофилы и мотылька, образовавшиеся в результате утраты в раннем развитии

Гены нумераторы (числителя), локализованные на X-хромосоме, кодируют факторы транскрипции sisterless-a, sisterless-b=scute
Гены нумераторы (числителя), локализованные на X-хромосоме, кодируют факторы транскрипции sisterless-a, sisterless-b=scute
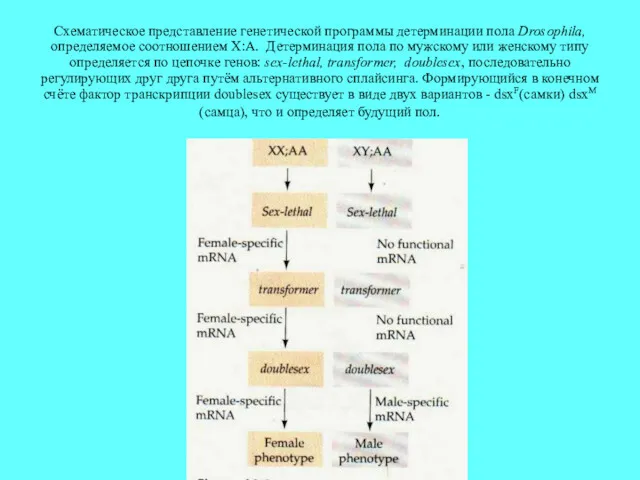
Схематическое представление генетической программы детерминации пола Drosophila, определяемое соотношением X:A. Детерминация
Схематическое представление генетической программы детерминации пола Drosophila, определяемое соотношением X:A. Детерминация
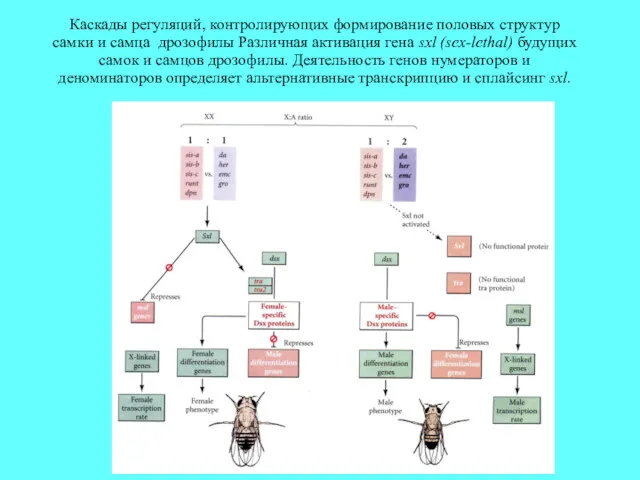
Каскады регуляций, контролирующих формирование половых структур самки и самца дрозофилы Различная
Каскады регуляций, контролирующих формирование половых структур самки и самца дрозофилы Различная
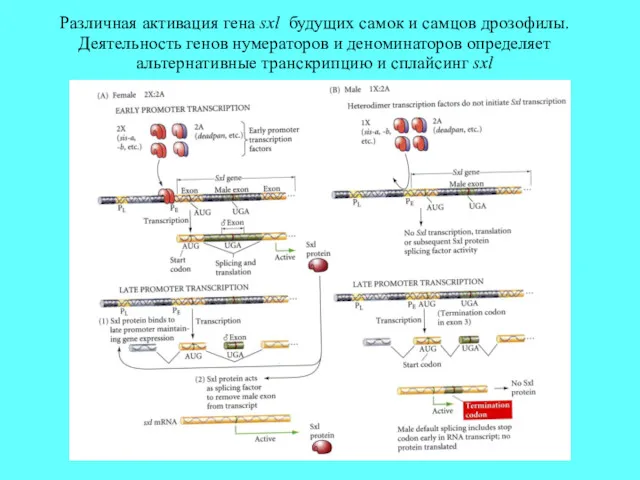
Различная активация гена sxl будущих самок и самцов дрозофилы. Деятельность генов
Различная активация гена sxl будущих самок и самцов дрозофилы. Деятельность генов
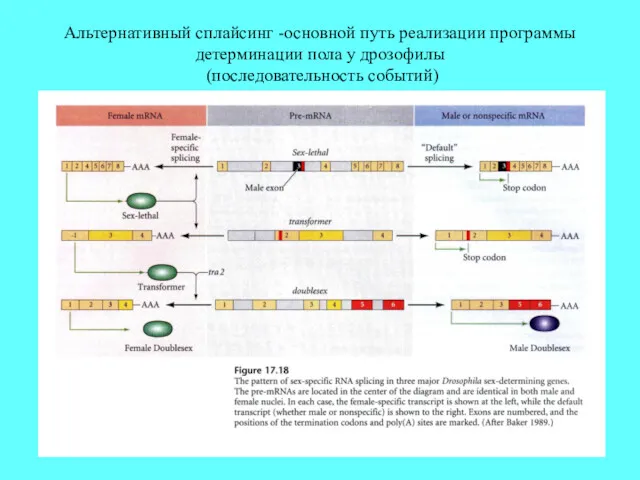
Альтернативный сплайсинг -основной путь реализации программы детерминации пола у дрозофилы (последовательность
Альтернативный сплайсинг -основной путь реализации программы детерминации пола у дрозофилы (последовательность

По ходу развития, когда sxl начинает транскрибироваться с позднего (дистального) промотора,
По ходу развития, когда sxl начинает транскрибироваться с позднего (дистального) промотора,
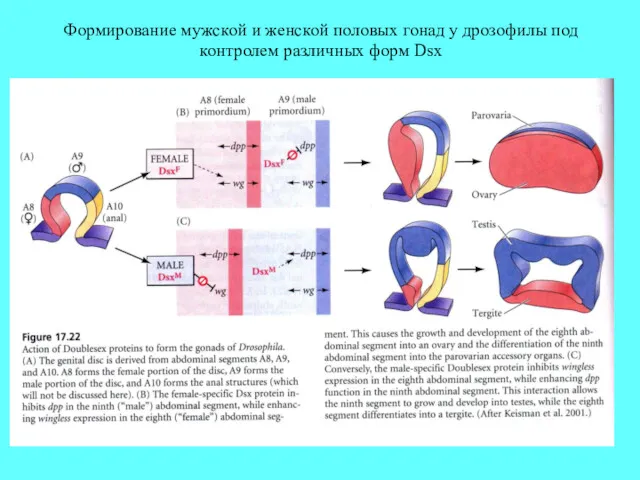
Формирование мужской и женской половых гонад у дрозофилы под контролем различных
Формирование мужской и женской половых гонад у дрозофилы под контролем различных
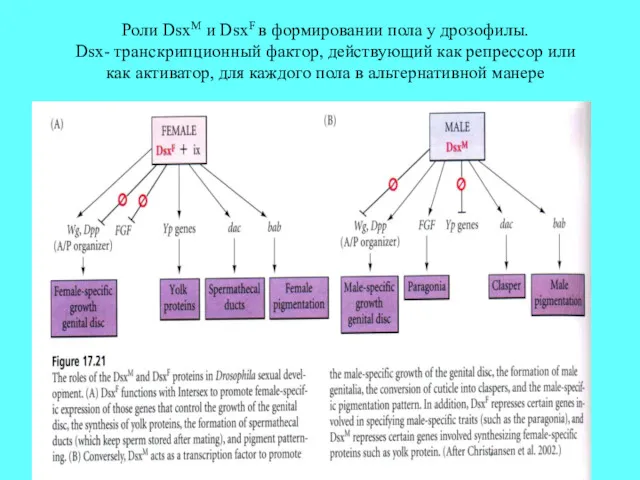
Роли DsxM и DsxF в формировании пола у дрозофилы.
Dsx- транскрипционный фактор,
Роли DsxM и DsxF в формировании пола у дрозофилы. Dsx- транскрипционный фактор,
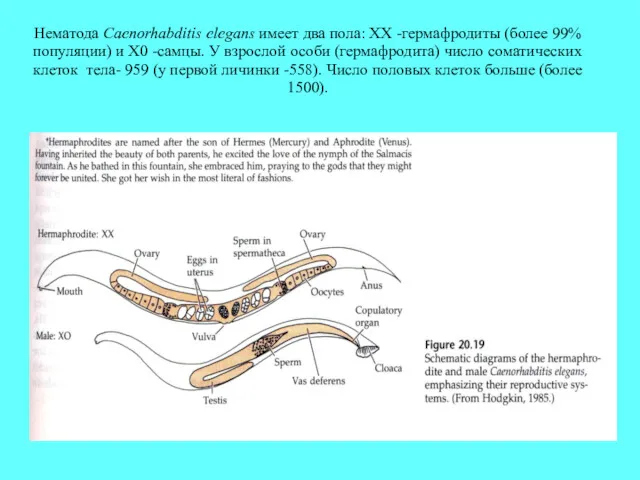
Нематода Caenorhabditis elegans имеет два пола: XX -гермафродиты (более 99% популяции)
Нематода Caenorhabditis elegans имеет два пола: XX -гермафродиты (более 99% популяции)
Нематода Сaenorhabditis elegans, гермафродитная особь
Нематода Сaenorhabditis elegans, гермафродитная особь

Нематода Сaenorhabditis elegans, самец
Нематода Сaenorhabditis elegans, самец

В случае самооплодотворения 0,2% новых особей -самцы, остальные -гермафродиты, в случае
В случае самооплодотворения 0,2% новых особей -самцы, остальные -гермафродиты, в случае

Детерминация пола у С. Elegans (также как и у Drosophila имеет
Детерминация пола у С. Elegans (также как и у Drosophila имеет
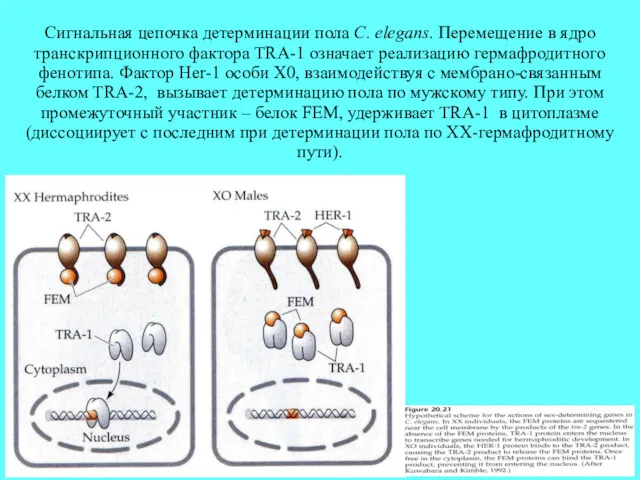
Сигнальная цепочка детерминации пола C. elegans. Перемещение в ядро транскрипционного фактора
Сигнальная цепочка детерминации пола C. elegans. Перемещение в ядро транскрипционного фактора
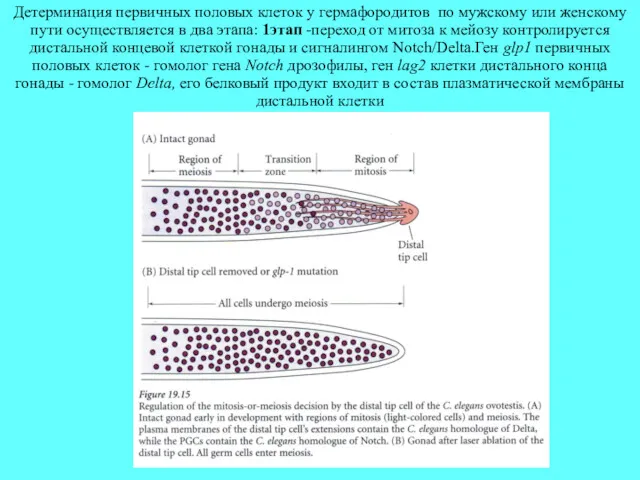
Детерминация первичных половых клеток у гермафородитов по мужскому или женскому пути
Детерминация первичных половых клеток у гермафородитов по мужскому или женскому пути
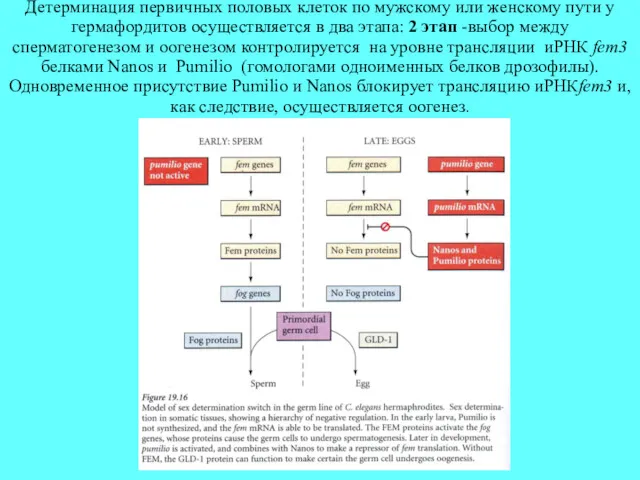
Детерминация первичных половых клеток по мужскому или женскому пути у гермафордитов
Детерминация первичных половых клеток по мужскому или женскому пути у гермафордитов
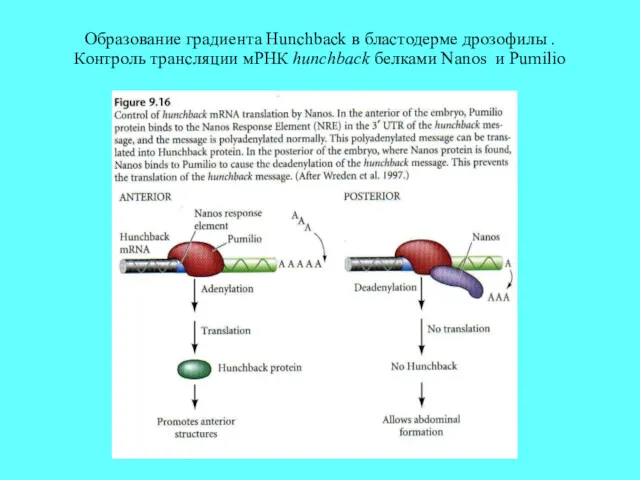
Образование градиента Hunchback в бластодерме дрозофилы . Контроль трансляции мРНК hunchback
Образование градиента Hunchback в бластодерме дрозофилы . Контроль трансляции мРНК hunchback

Эволюционные связи между генами, ответственными за детерминацию пола у разных животных
В
Эволюционные связи между генами, ответственными за детерминацию пола у разных животных
В
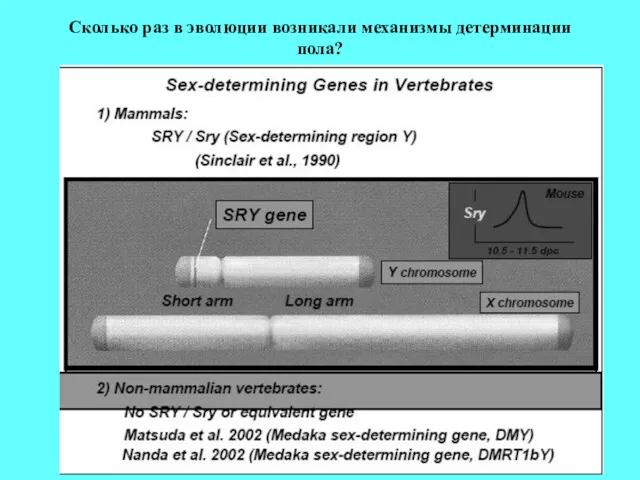
Сколько раз в эволюции возникали механизмы детерминации пола?
Сколько раз в эволюции возникали механизмы детерминации пола?

Сколько раз в эволюции возникали механизмы детерминации пола?
Гипотеза:
Cуществуют две группы позвоночных
Сколько раз в эволюции возникали механизмы детерминации пола? Гипотеза: Cуществуют две группы позвоночных
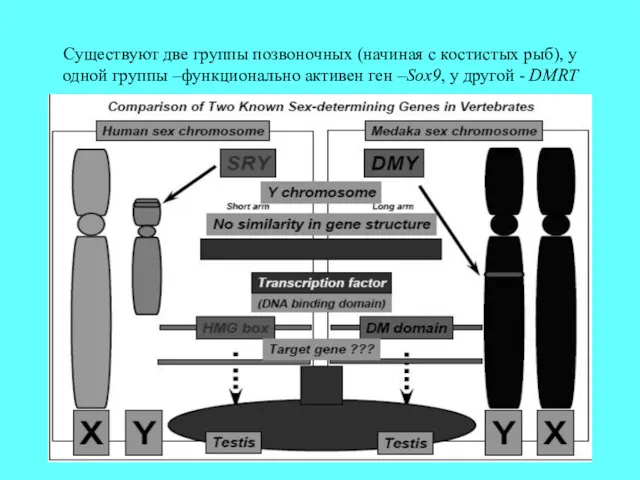
Cуществуют две группы позвоночных (начиная с костистых рыб), у одной группы
Cуществуют две группы позвоночных (начиная с костистых рыб), у одной группы
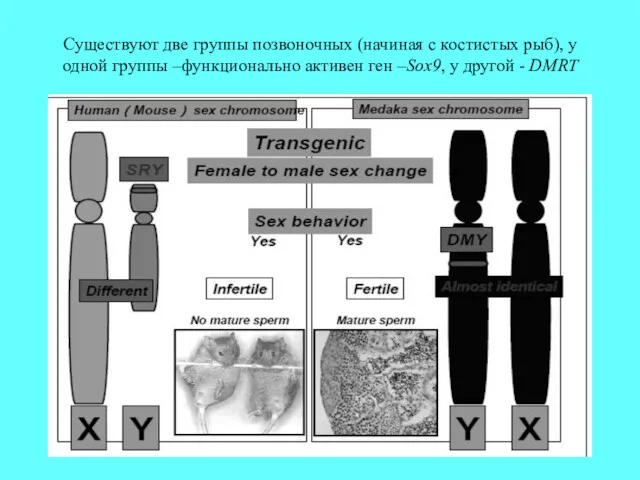
Cуществуют две группы позвоночных (начиная с костистых рыб), у одной группы
Cуществуют две группы позвоночных (начиная с костистых рыб), у одной группы

На 9-й хромосоме человека (короткое плечо) было идентифицировано 3 гена Dmrt
На 9-й хромосоме человека (короткое плечо) было идентифицировано 3 гена Dmrt

Ортологи генов Sox 9 активно экспрессируются в клетках Сертоли у птиц
Ортологи генов Sox 9 активно экспрессируются в клетках Сертоли у птиц
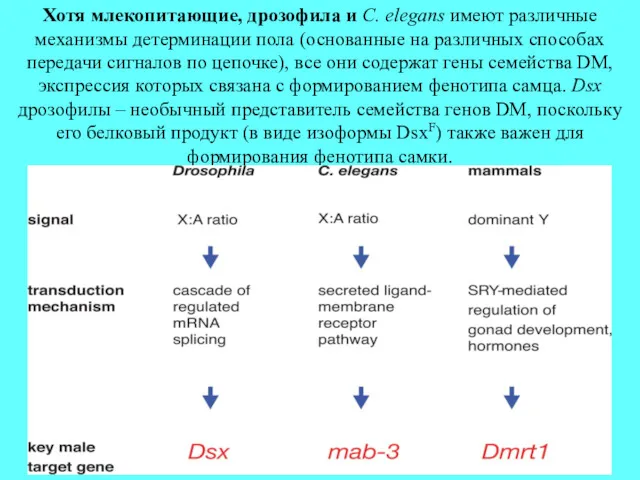
Хотя млекопитающие, дрозофила и C. elegans имеют различные механизмы детерминации пола
Хотя млекопитающие, дрозофила и C. elegans имеют различные механизмы детерминации пола
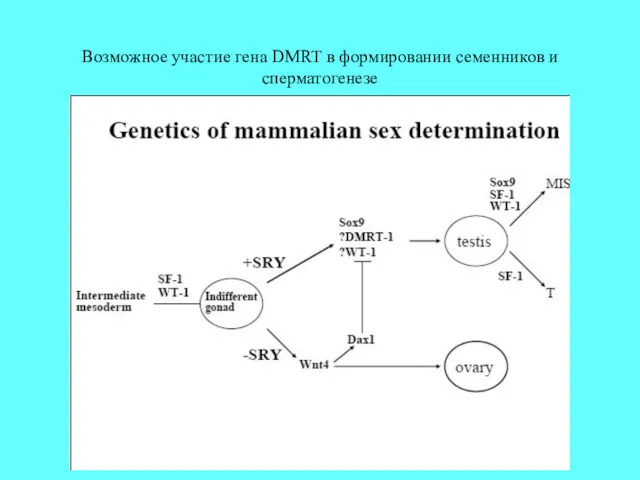
Возможное участие гена DMRT в формировании семенников и сперматогенезе
Возможное участие гена DMRT в формировании семенников и сперматогенезе
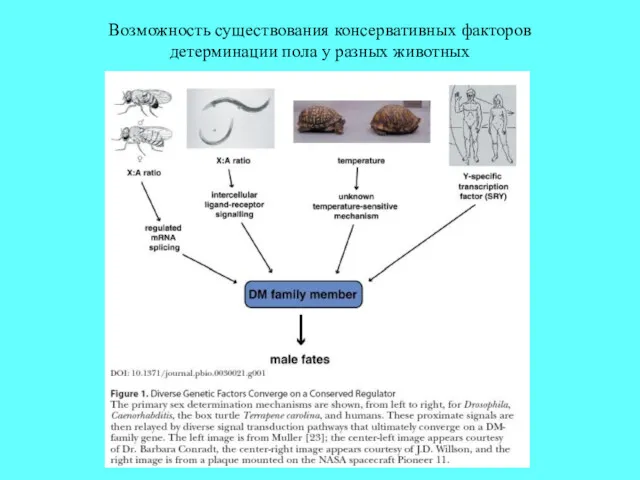
Возможность существования консервативных факторов детерминации пола у разных животных
Возможность существования консервативных факторов детерминации пола у разных животных


 Көбею. Өсу және даму
Көбею. Өсу және даму Генетика людини
Генетика людини Семейство Розоцветные
Семейство Розоцветные Мышечная система человека
Мышечная система человека Что такое адаптации? Какие бывают адаптации?
Что такое адаптации? Какие бывают адаптации? Физиология пищеварения. Пищеварение в полости рта и желудка
Физиология пищеварения. Пищеварение в полости рта и желудка Урок на тему Приспособленность организмов к среде обитания и ее относительный характер. Общая биология. 9 класс.
Урок на тему Приспособленность организмов к среде обитания и ее относительный характер. Общая биология. 9 класс. Влияние галогенов на организм человека
Влияние галогенов на организм человека Памятники животным
Памятники животным междисциплинарное обучение
междисциплинарное обучение Тройничный нерв
Тройничный нерв Вегетативтік әрекеттердің гуморалдық реттелуі. Гипоталамо-гипофизарлық жүйе
Вегетативтік әрекеттердің гуморалдық реттелуі. Гипоталамо-гипофизарлық жүйе Поведінка тварин
Поведінка тварин Роль металлов в организме человека
Роль металлов в организме человека Особенности обмена веществ у детей дошкольного возраста
Особенности обмена веществ у детей дошкольного возраста Молекулярная структура фотосинтетического аппарата и регуляция экспрессии генов фотосинтеза
Молекулярная структура фотосинтетического аппарата и регуляция экспрессии генов фотосинтеза Механика и человек. Законы физики на службе у человека. Материалы и методы исследования
Механика и человек. Законы физики на службе у человека. Материалы и методы исследования Дрожалка оранжевая (tremella mesenterica)
Дрожалка оранжевая (tremella mesenterica) Соль и сахар. (3 класс)
Соль и сахар. (3 класс) презентация к уроку биологии по теме Клетка
презентация к уроку биологии по теме Клетка Презентация Рыбы часть1,2
Презентация Рыбы часть1,2 ОГЭ по обществознанию, 9 класс. Человек и общество
ОГЭ по обществознанию, 9 класс. Человек и общество Жизненный цикл растений. Вопросы
Жизненный цикл растений. Вопросы Классификация эпителиев
Классификация эпителиев Зоология – наука о животных подготовка к ОГЭ и ЕГЭ по биологии
Зоология – наука о животных подготовка к ОГЭ и ЕГЭ по биологии Съедобные и ядовитые дикорастущие растения
Съедобные и ядовитые дикорастущие растения Звёздный час развития и эволюции растительноядных. 1 часть
Звёздный час развития и эволюции растительноядных. 1 часть Многообразие пресноводных рыб в водоемах
Многообразие пресноводных рыб в водоемах