Электрогенез клетки. Мембранный потенциал. Электрические сигналы (локальный потенциал и потенциал действия) презентация
- Электрогенез клетки. Мембранный потенциал. Электрические сигналы (локальный потенциал и потенциал действия)

Содержание
- 2. 1. Электрогенез клетки
- 3. Схема строения клеточной мембраны Мембрана – крепостная стена и одновременно ворота для входа в клетку и
- 4. Frederick George Donnan (September 6, 1870 – December 16, 1956) was an Irish physical chemist who
- 5. FIGURE 3 The equilibrium potential is influenced by the concentration gradient and the voltage difference across
- 6. Равновесие Доннана
- 7. Концентрация ионов снаружи и внутри клетки, мМ/л Таблица 2.2. Концентрация ионов снаружи и внутри клетки, ммоль/л
- 8. Органические 15 11 137 анионы Концентрация ионов в цитозоле и внеклеточной среде нейрона млекопитающего (в мМ/л)
- 9. FIGURE 2 Differential distribution of ions inside and outside plasma membrane of neurons and neuronal processes,
- 10. Схема опыта по измерению мембранного потенциала клетки K+= 140 mM K+= 4 mM Уравнение Нернста «Если
- 11. Вальтер Герман Нернст — немецкий химик, лауреат Нобелевской премии по химии в 1920 году «в признание
- 12. Ионные потоки через мембрану клетки Снаружи Внутри Мало калия Много натрия Много калия Мало натрия Высокая
- 13. FIGURE 5 The voltage-clamp technique keeps the voltage across the membrane constant so that the amplitude
- 14. 2. Электрические сигналы, возникающие в нервной клетке (локальный потенциал и потенциал действия)
- 15. Что вызывает локальные потенциалы?
- 16. Типы локальных потенциалов Локальные потенциалы градуальны: 1 и 2 гиперполяризующие (тормозящие) 3 деполяризующий (возбуждающий) 4 если
- 17. Пассивное распространение электротонического потенциала вдоль нервного проводника (с декрементом, т.е. затуханием) Большинство нервных сигналов локальны, т.е.
- 18. Локальный потенциал - локальный ответ распространяется декрементно, т. е. по мере удаления от места раздражения его
- 19. Если локальный деполяризующий стимул достиг уровня порога, то появляются условия для генерации потенциала действия Порог Потенциал
- 20. FIGURE 1 Intracellular recording of the membrane potential and action potential generation in the squid giant
- 21. Родился: 5 февраля 1914 г., Банбери, Великобритания Умер: 20 декабря 1998 г., Кембридж, Великобритания Алан Ходжкин
- 22. Мембранный потенциал и потенциал действия Все начинается с появления локального потенциала. Если его амплитуда достигает определенного
- 23. Начальный этап генерации потенциала действия Все начинается с появления локального потенциала. Если его амплитуда достигает определенного
- 24. Локальный потенциал активирует сначала небольшое число самых чувствительных Na-каналов, но входящий через них Na+ дополнительно деполяризует
- 25. Когда большинство натриевых каналов открыты – потенциал на мембране во время пика достигает уровень натриевого равновесного
- 26. Натриевые каналы могут находиться в открытом состоянии в течение короткого периода – происходит их инактивация По
- 27. А — в покое m-активационные ворота («m-ворота») закрыты; Б — при возбуждении «m-ворота» открыты; В —
- 28. Уже в фазу роста ПД по мере быстро нарастающего натриевого тока начинают медленно открываться калиевые каналы,
- 29. Ток через открытые калиевые каналы сдвигает мембранный потенциал к уровню калиевого равновесного потенциала (следовая гиперполяризация) Na+
- 30. Конечный этап генерации потенциала действия Часть калиевых каналов закрывается и мембранный потенциал медленно возвращается к исходному
- 31. Потенциал действия – результат суммирования разнонаправленных (натриевый ток внутрь, калиевый ток наружу) и сдвинутых по временной
- 32. Последовательность открывания-закрывания натриевых и калиевых каналов во время генерации потенциала действия
- 33. FIGURE 7 Generation of the action potential is associated with an increase in membrane Na+ conductance
- 34. Различия в кинетике процессов активации и инактивации натриевых и калиевых каналов предопределяют феномен существование рефрактерных периодов
- 35. FIGURE 6 Voltage-clamp analysis reveals ionic currents underlying action potential generation
- 36. 3. Проведение нервного импульса по аксону
- 37. Структура нейрона
- 38. Почему у беспозвоночных есть гигантские аксоны?
- 39. Миелинизированный (мякотный) нерв
- 40. FIGURE 8 Propagation of the action potential in unmyelinated and myelinated axons. (A) Action potentials propagate
- 41. Стимул Миелиновая оболочка Аксон Проведение локального потенциала под оболочкой (декремент) Генерация потенциала действия (ПД) в перехвате
- 42. Потенциал действия будет распространяться по нерву быстро и без потерь на значительные расстояния, если Нерв имеет
- 43. Сравнение локального потенциала и потенциала действия
- 44. 4. Ионные каналы и рецепторы – молекулярная основа генерации клеточных сигналов
- 45. Ионные каналы и другие интегральные белки в фосфолипидной мембране Общая схема строения ионного канала
- 46. Ацетилхолиновый рецептор в липидном бислое мембраны
- 47. Субъединичное строение ацетилхолинового рецептора
- 48. Потенциалозависимые Закрытый Открытый Механочувствительные Лигандуправляемые Снаружи Изнутри Основные типы ионных каналов
- 49. Регистрация одиночных ионных каналов (patch clamp)
- 50. Регистрация одиночных ионных каналов (patch clamp)
- 51. Примеры одиночных ионных каналов, зарегистрированных методом пэтч-клемп Глутаматный рецептор Ацетилхолиновый рецептор Глициновый рецептор
- 52. Инъекция мРНК, выделяемой из мозговой ткани в ооциты лягушки Xenopus и регистрация ответов экспрессированных рецепторов
- 53. Клонирование генов определенных каналов и рецепторов, создание библиотек генов и последующее их использование
- 54. Ионные каналы клонированы, изучены аминокислотные последовательности их полипептидных цепей и топография в мембране
- 55. Структура потенциалзависимых натриевых и кальциевых каналов Натриевый канал – одна полипептидная цепь с четырьмя доменами, соединенными
- 56. Субъединица калиевого канала похожа на домен натриевого канала Трехмерная структура калиевого канала Структура потенциалзависимого калиевого канала
- 57. Селективный фильтр Na+ канала Узкий участок водной поры канала, определяющий тип иона, способного пройти через канал,
- 58. Поперечный срез трехмерной структуры калиевого канала
- 59. K+ канал в мембране бактерии (структура по данным кристаллографии) Doyle et al, 1998
- 60. AMPA рецептор Молекулярные модели селективных фильтров открытых каналов глутматных рецепторов NMDA и AMPA типов Первые, помимо
- 61. Токсины, действующие на Na+ канал Рыба кузовок содержит tetrodotoxin, сильнейший яд. Batrachotoxin содержится в коже колумбийских
- 62. Разделение натриевых и калиевых токов, лежащих в основе потенциала действия Суммарный ток Натриевые каналы заблокированы, регистрируется
- 63. Ионотропные и метаботропные рецепторы
- 64. G-белок 1. Выключенное состояние: α‑СЕ связана с гуанозиндифосфатом (ГДФ) и не контактирует с рецептором; 2. При
- 65. Роль инозитолтрифосфата и диацилглицерола в реализации эффекта лигандов на клетку–мишень [11]. Образование комплекса лиганда с рецептором
- 66. Вопросы?
- 68. Химические структуры веществ, блокирующих натриевые (TTX, procaine) и калиевые (ТЕА) каналы возбудимой мембраны. С их помощью
- 69. Сравнение локального потенциала и потенциала действия Локальный потенциал Потенциал действия
- 70. Диффузионный потенциал, создаваемый разностью концентраций ионов калия (К+) в сосудах, разделенных полупроницаемой перегородкой, непропускающей анионы (А-)
- 71. FIGURE 8 Propagation of the action potential in unmyelinated and myelinated axons. (A) Action potentials propagate
- 72. FIGURE 9 Structure of the sodium channel. (A) Cross section of a hypothetical sodium channel consisting
- 73. Ионные каналы и рецепторы
- 74. FIGURE 4 Increases in K+ conductance can result in hyperpolarization, depolarization, or no change in membrane
- 76. Скачать презентацию

1. Электрогенез клетки
1. Электрогенез клетки
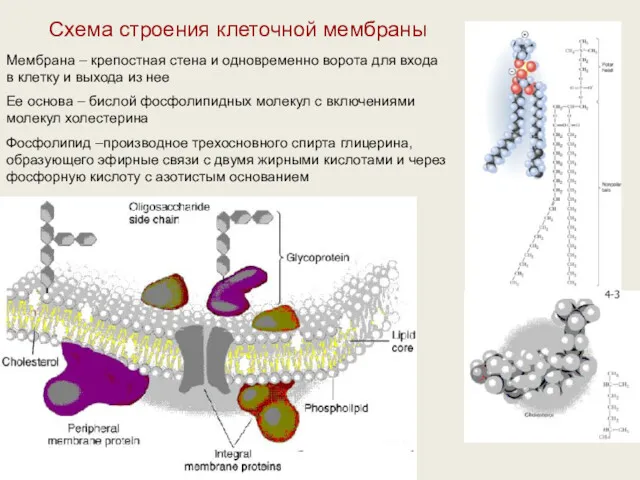
Схема строения клеточной мембраны
Мембрана – крепостная стена и одновременно ворота для
Схема строения клеточной мембраны
Мембрана – крепостная стена и одновременно ворота для

Frederick George Donnan (September 6, 1870 – December 16, 1956) was
Frederick George Donnan (September 6, 1870 – December 16, 1956) was
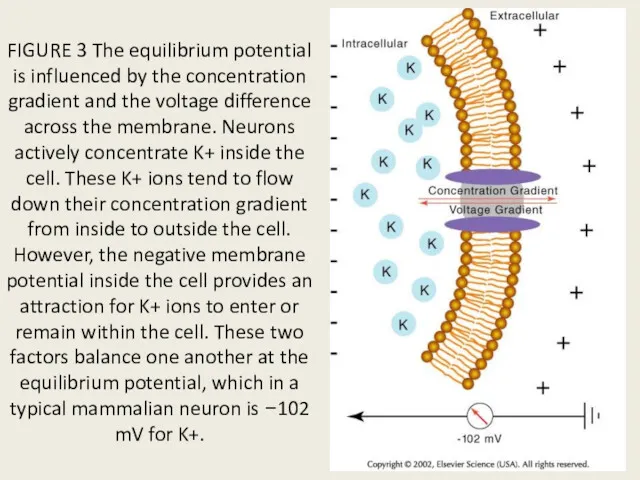
FIGURE 3 The equilibrium potential is influenced by the concentration gradient
FIGURE 3 The equilibrium potential is influenced by the concentration gradient

Равновесие Доннана
Равновесие Доннана

Концентрация ионов снаружи и внутри клетки, мМ/л
Таблица 2.2. Концентрация ионов снаружи
Концентрация ионов снаружи и внутри клетки, мМ/л
Таблица 2.2. Концентрация ионов снаружи
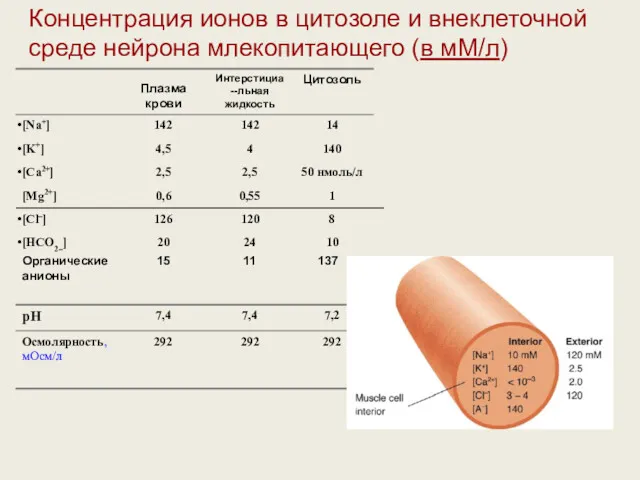
Органические 15 11 137
анионы
Концентрация ионов в цитозоле и внеклеточной
Органические 15 11 137
анионы
Концентрация ионов в цитозоле и внеклеточной
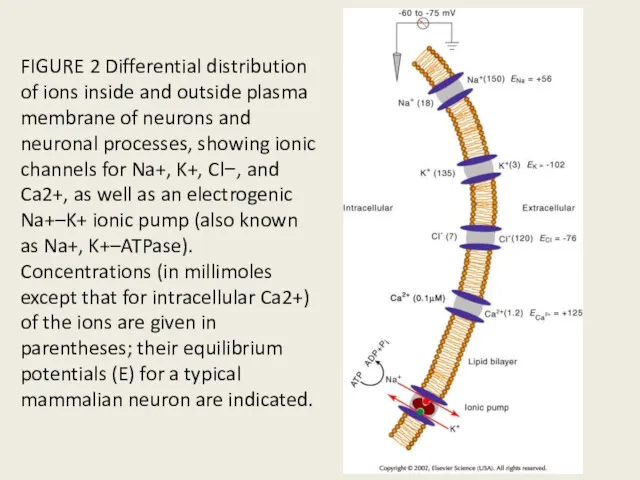
FIGURE 2 Differential distribution of ions inside and outside plasma membrane
FIGURE 2 Differential distribution of ions inside and outside plasma membrane
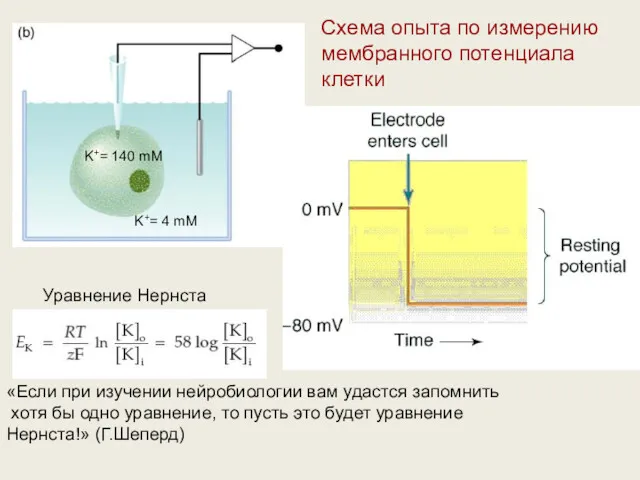
Схема опыта по измерению мембранного потенциала клетки
K+= 140 mM
K+= 4 mM
Уравнение
Схема опыта по измерению мембранного потенциала клетки
K+= 140 mM
K+= 4 mM
Уравнение

Вальтер Герман Нернст — немецкий химик, лауреат Нобелевской премии по химии
Вальтер Герман Нернст — немецкий химик, лауреат Нобелевской премии по химии
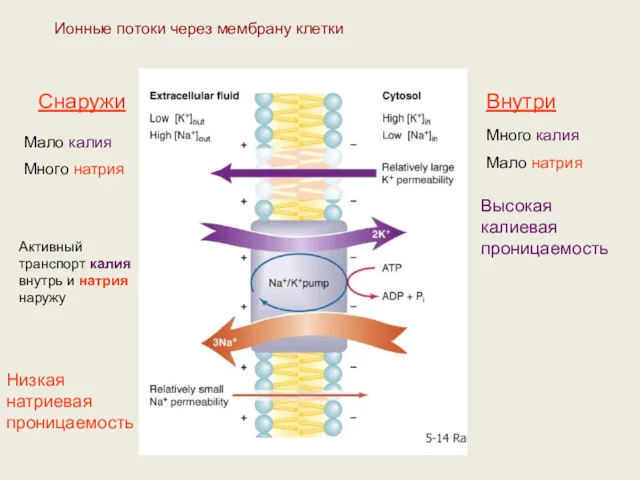
Ионные потоки через мембрану клетки
Снаружи
Внутри
Мало калия
Много натрия
Много калия
Мало натрия
Высокая калиевая проницаемость
Низкая
Ионные потоки через мембрану клетки
Снаружи
Внутри
Мало калия
Много натрия
Много калия
Мало натрия
Высокая калиевая проницаемость
Низкая
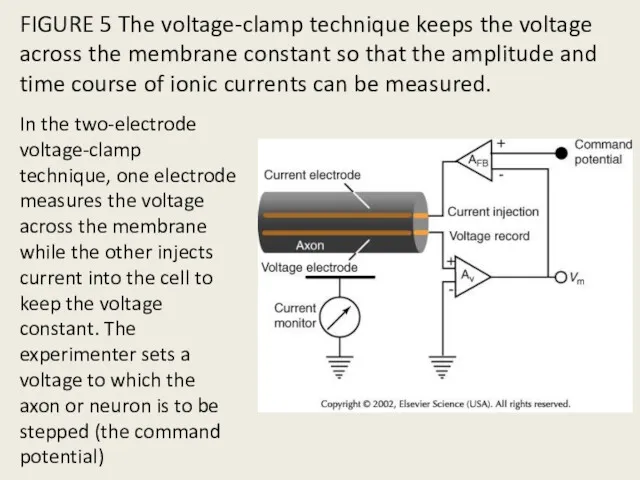
FIGURE 5 The voltage-clamp technique keeps the voltage across the membrane
FIGURE 5 The voltage-clamp technique keeps the voltage across the membrane

2. Электрические сигналы, возникающие в нервной клетке (локальный потенциал и потенциал
2. Электрические сигналы, возникающие в нервной клетке (локальный потенциал и потенциал

Что вызывает
локальные потенциалы?
Что вызывает
локальные потенциалы?
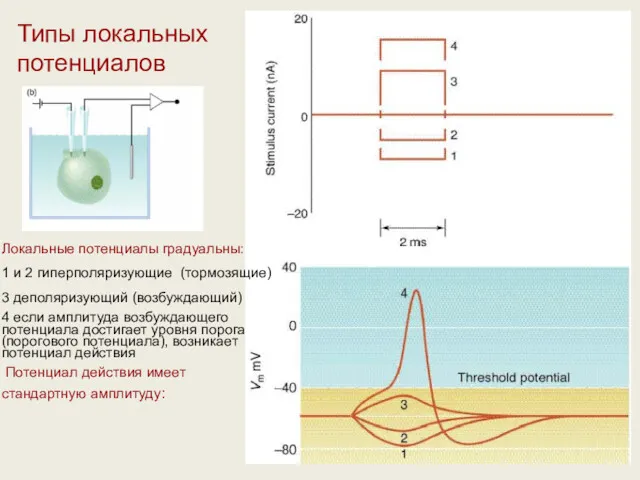
Типы локальных потенциалов
Локальные потенциалы градуальны:
1 и 2 гиперполяризующие (тормозящие)
3 деполяризующий (возбуждающий)
4
Типы локальных потенциалов
Локальные потенциалы градуальны:
1 и 2 гиперполяризующие (тормозящие)
3 деполяризующий (возбуждающий)
4
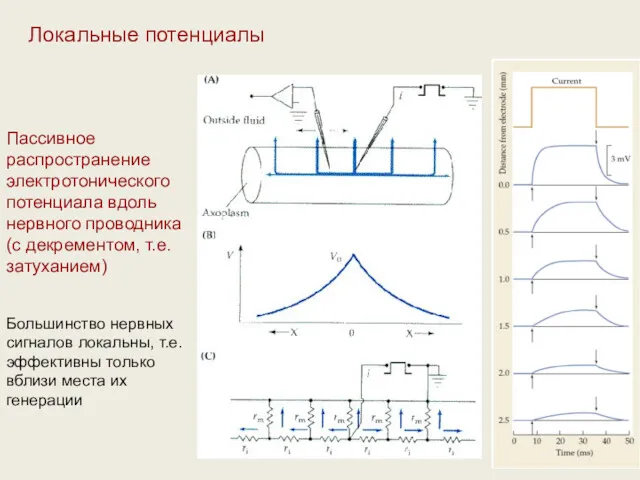
Пассивное распространение электротонического потенциала вдоль нервного проводника (с декрементом, т.е. затуханием)
Большинство
Пассивное распространение электротонического потенциала вдоль нервного проводника (с декрементом, т.е. затуханием)
Большинство

Локальный потенциал
- локальный ответ распространяется декрементно, т. е. по мере удаления
Локальный потенциал
- локальный ответ распространяется декрементно, т. е. по мере удаления

Если локальный деполяризующий стимул достиг уровня порога, то появляются условия для
Если локальный деполяризующий стимул достиг уровня порога, то появляются условия для
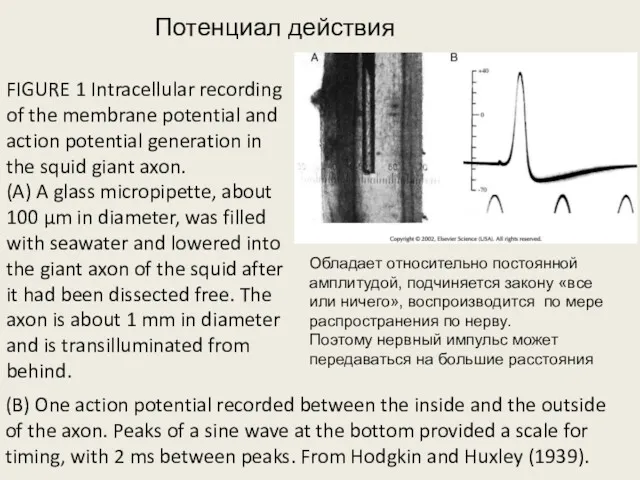
FIGURE 1 Intracellular recording of the membrane potential and action potential
FIGURE 1 Intracellular recording of the membrane potential and action potential

Родился: 5 февраля 1914 г., Банбери, Великобритания
Умер: 20 декабря 1998 г.,
Родился: 5 февраля 1914 г., Банбери, Великобритания
Умер: 20 декабря 1998 г.,
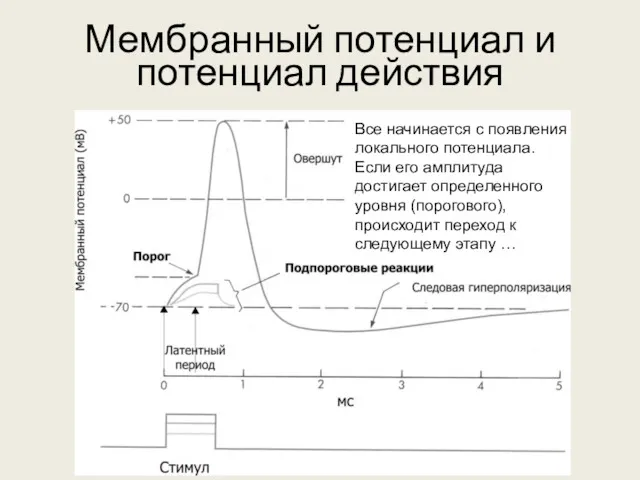
Мембранный потенциал и потенциал действия
Все начинается с появления локального потенциала. Если
Мембранный потенциал и потенциал действия
Все начинается с появления локального потенциала. Если
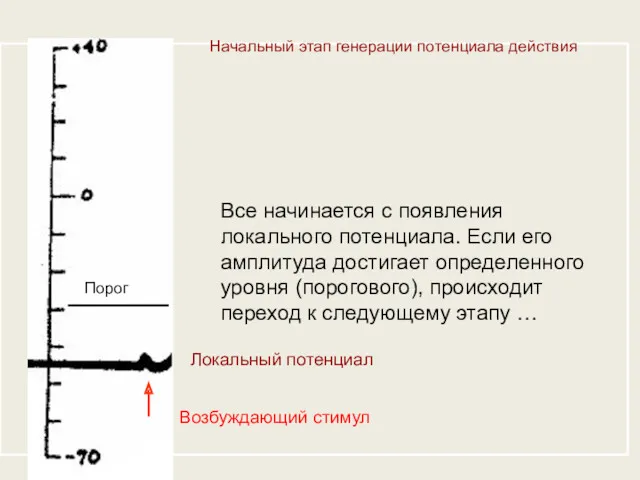
Начальный этап генерации потенциала действия
Все начинается с появления локального потенциала. Если
Начальный этап генерации потенциала действия
Все начинается с появления локального потенциала. Если

Локальный потенциал активирует сначала небольшое число самых чувствительных Na-каналов, но входящий
Локальный потенциал активирует сначала небольшое число самых чувствительных Na-каналов, но входящий

Когда большинство натриевых каналов открыты – потенциал на мембране во время
Когда большинство натриевых каналов открыты – потенциал на мембране во время
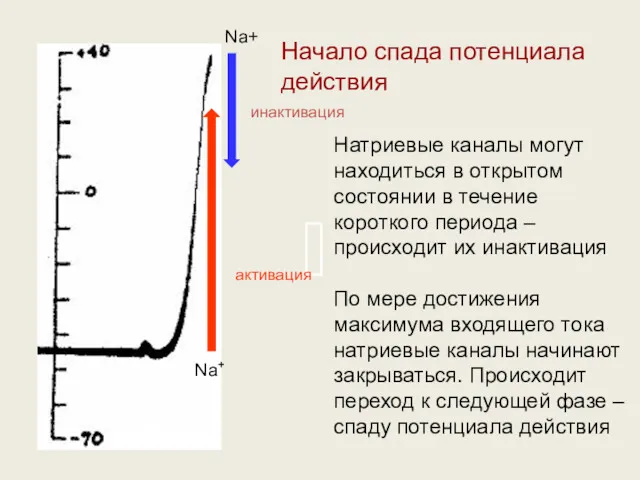
Натриевые каналы могут находиться в открытом состоянии в течение короткого периода
Натриевые каналы могут находиться в открытом состоянии в течение короткого периода
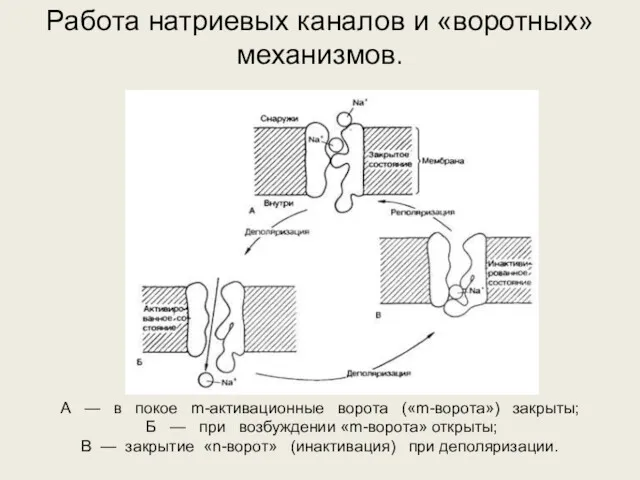
А — в покое m-активационные ворота («m-ворота») закрыты;
Б — при возбуждении «m-ворота»
А — в покое m-активационные ворота («m-ворота») закрыты;
Б — при возбуждении «m-ворота»
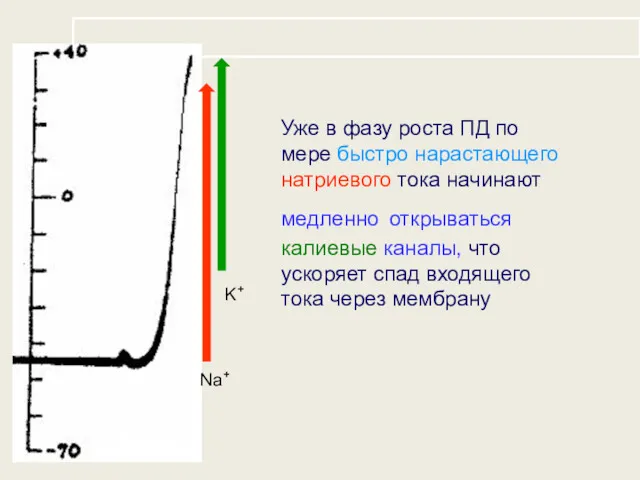
Уже в фазу роста ПД по мере быстро нарастающего натриевого тока
Уже в фазу роста ПД по мере быстро нарастающего натриевого тока

Ток через открытые калиевые каналы сдвигает мембранный потенциал к уровню калиевого
Ток через открытые калиевые каналы сдвигает мембранный потенциал к уровню калиевого
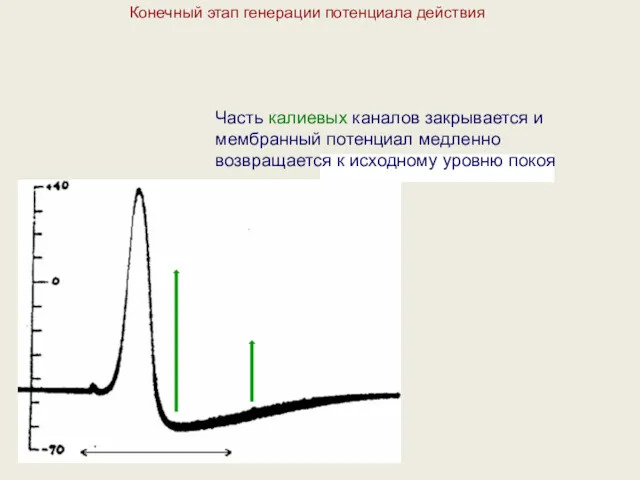
Конечный этап генерации потенциала действия
Часть калиевых каналов закрывается и мембранный потенциал
Конечный этап генерации потенциала действия
Часть калиевых каналов закрывается и мембранный потенциал

Потенциал действия – результат суммирования разнонаправленных (натриевый ток внутрь, калиевый ток
Потенциал действия – результат суммирования разнонаправленных (натриевый ток внутрь, калиевый ток
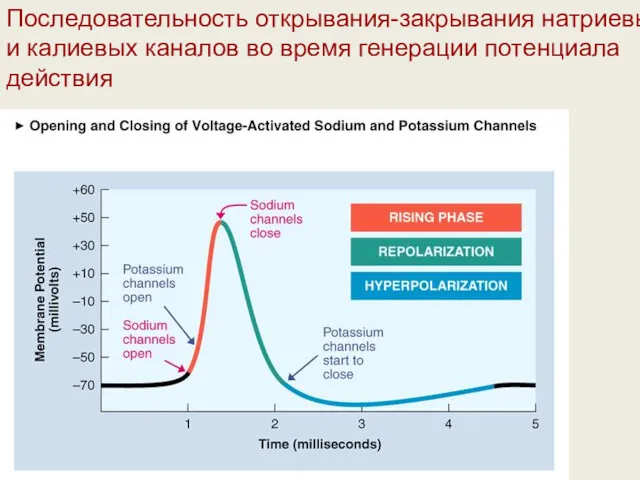
Последовательность открывания-закрывания натриевых и калиевых каналов во время генерации потенциала действия
Последовательность открывания-закрывания натриевых и калиевых каналов во время генерации потенциала действия

FIGURE 7 Generation of the action potential is associated with an
FIGURE 7 Generation of the action potential is associated with an
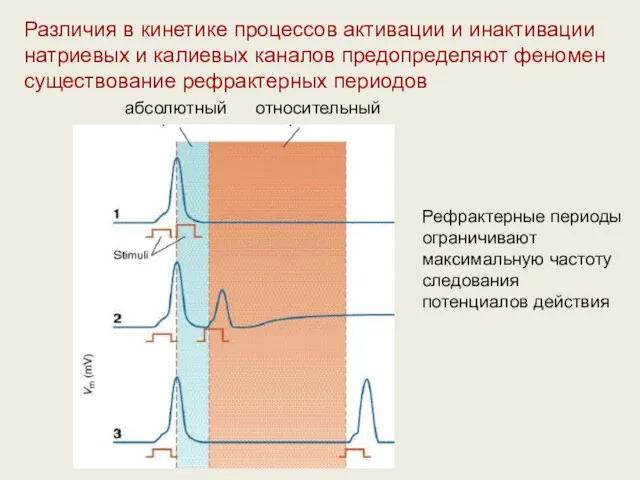
Различия в кинетике процессов активации и инактивации натриевых и калиевых каналов
Различия в кинетике процессов активации и инактивации натриевых и калиевых каналов

FIGURE 6 Voltage-clamp analysis reveals ionic currents underlying action potential generation
FIGURE 6 Voltage-clamp analysis reveals ionic currents underlying action potential generation

3. Проведение нервного импульса по аксону
3. Проведение нервного импульса по аксону
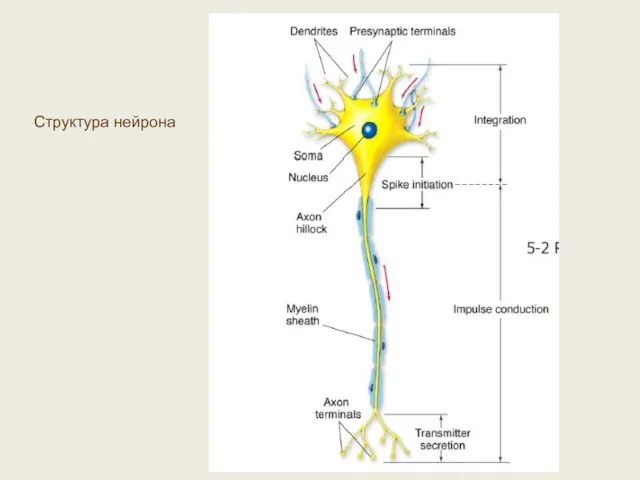
Структура нейрона
Структура нейрона

Почему у беспозвоночных
есть гигантские аксоны?
Почему у беспозвоночных
есть гигантские аксоны?
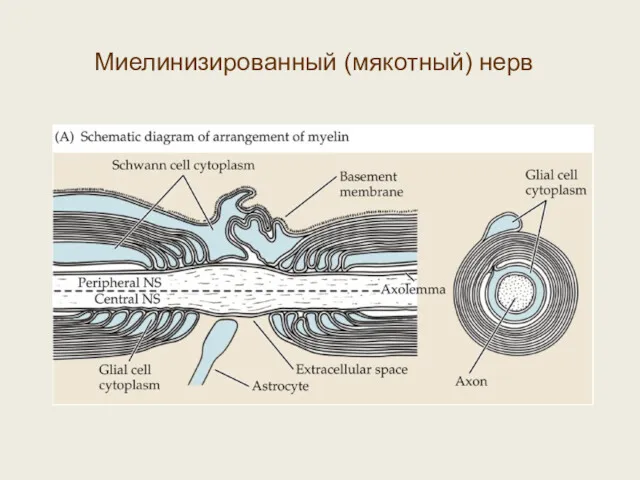
Миелинизированный (мякотный) нерв
Миелинизированный (мякотный) нерв
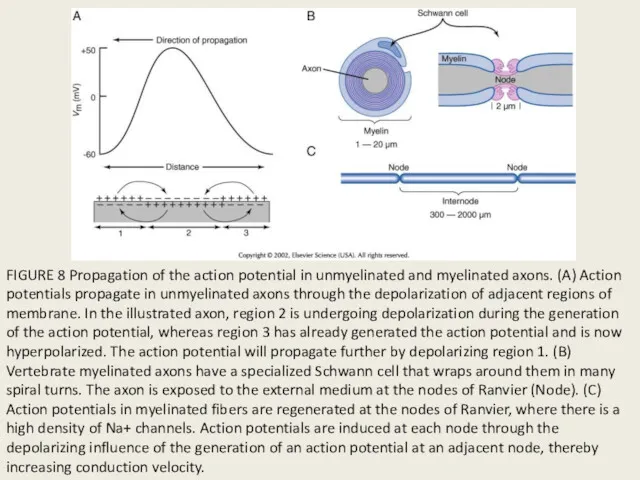
FIGURE 8 Propagation of the action potential in unmyelinated and myelinated
FIGURE 8 Propagation of the action potential in unmyelinated and myelinated

Стимул
Миелиновая оболочка
Аксон
Проведение локального потенциала под оболочкой (декремент)
Генерация потенциала действия (ПД) в
Стимул
Миелиновая оболочка
Аксон
Проведение локального потенциала под оболочкой (декремент)
Генерация потенциала действия (ПД) в

Потенциал действия будет распространяться по нерву быстро и без потерь на
Потенциал действия будет распространяться по нерву быстро и без потерь на
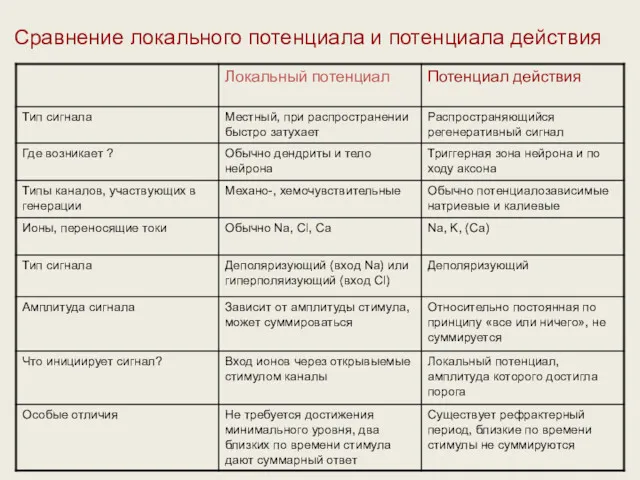
Сравнение локального потенциала и потенциала действия
Сравнение локального потенциала и потенциала действия

4. Ионные каналы и рецепторы – молекулярная основа генерации клеточных сигналов
4. Ионные каналы и рецепторы – молекулярная основа генерации клеточных сигналов
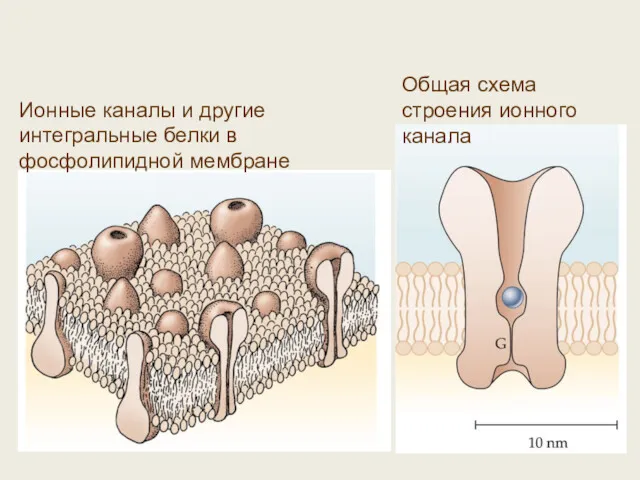
Ионные каналы и другие интегральные белки в фосфолипидной мембране
Общая схема строения
Ионные каналы и другие интегральные белки в фосфолипидной мембране
Общая схема строения
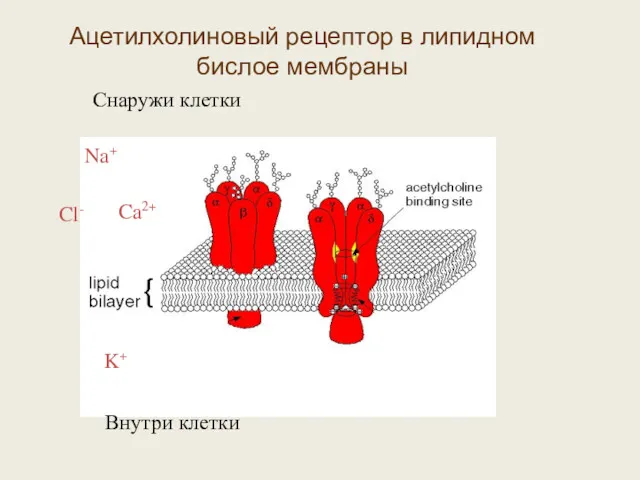
Ацетилхолиновый рецептор в липидном бислое мембраны
Ацетилхолиновый рецептор в липидном бислое мембраны
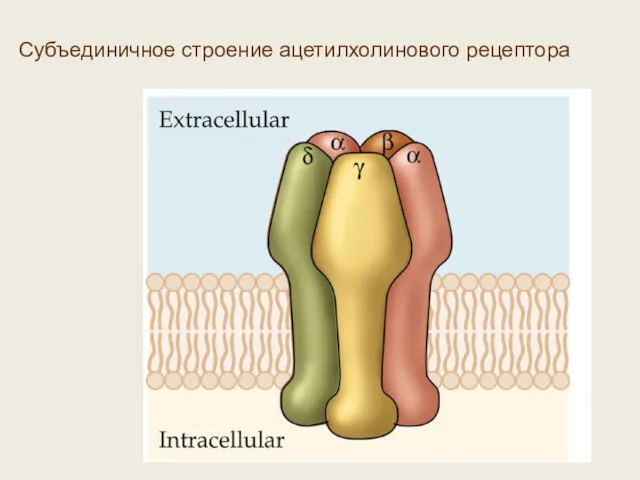
Субъединичное строение ацетилхолинового рецептора
Субъединичное строение ацетилхолинового рецептора
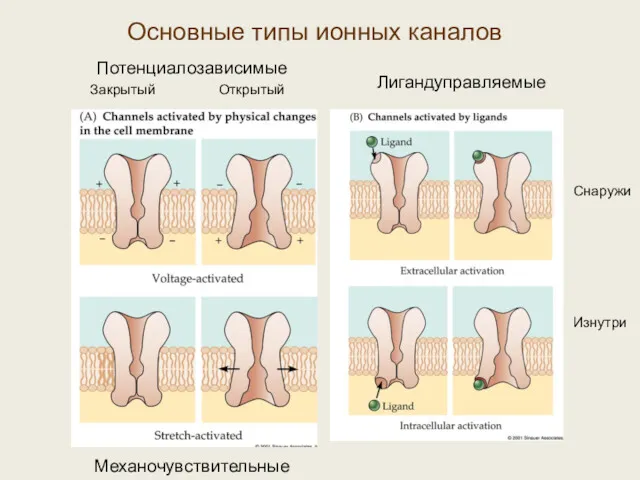
Потенциалозависимые
Закрытый Открытый
Механочувствительные
Лигандуправляемые
Снаружи
Изнутри
Основные типы ионных каналов
Потенциалозависимые
Закрытый Открытый
Механочувствительные
Лигандуправляемые
Снаружи
Изнутри
Основные типы ионных каналов
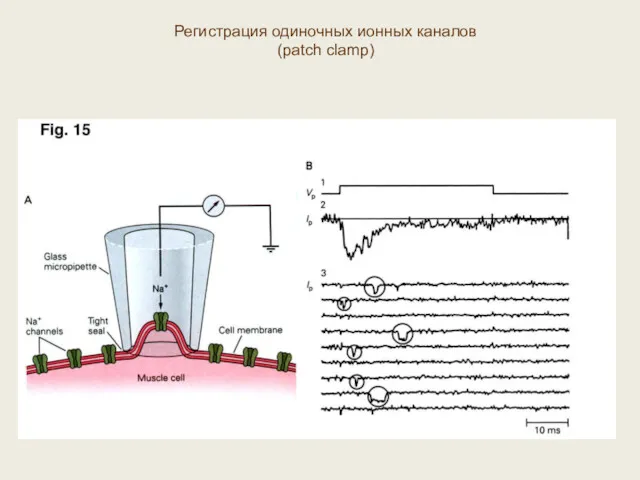
Регистрация одиночных ионных каналов
(patch clamp)
Регистрация одиночных ионных каналов
(patch clamp)
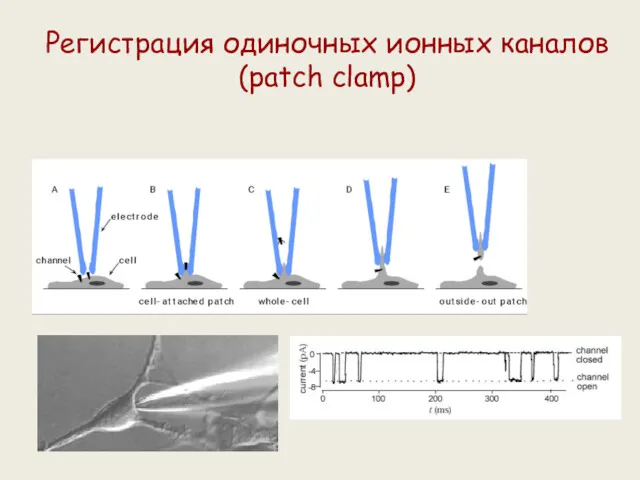
Регистрация одиночных ионных каналов (patch clamp)
Регистрация одиночных ионных каналов (patch clamp)
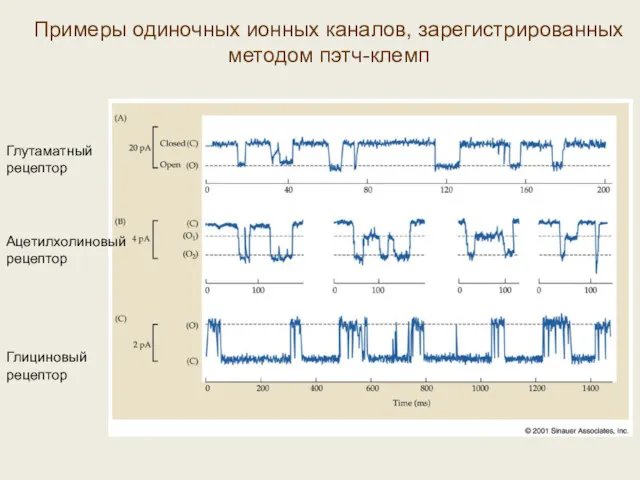
Примеры одиночных ионных каналов, зарегистрированных методом пэтч-клемп
Глутаматный рецептор
Ацетилхолиновый рецептор
Глициновый рецептор
Примеры одиночных ионных каналов, зарегистрированных методом пэтч-клемп
Глутаматный рецептор
Ацетилхолиновый рецептор
Глициновый рецептор
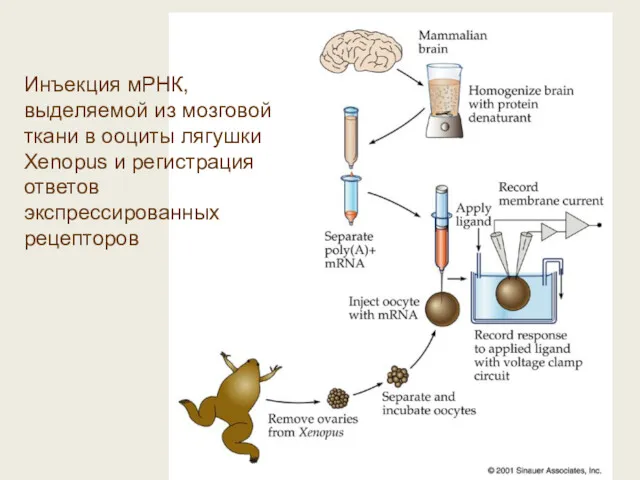
Инъекция мРНК, выделяемой из мозговой ткани в ооциты лягушки Xenopus и
Инъекция мРНК, выделяемой из мозговой ткани в ооциты лягушки Xenopus и
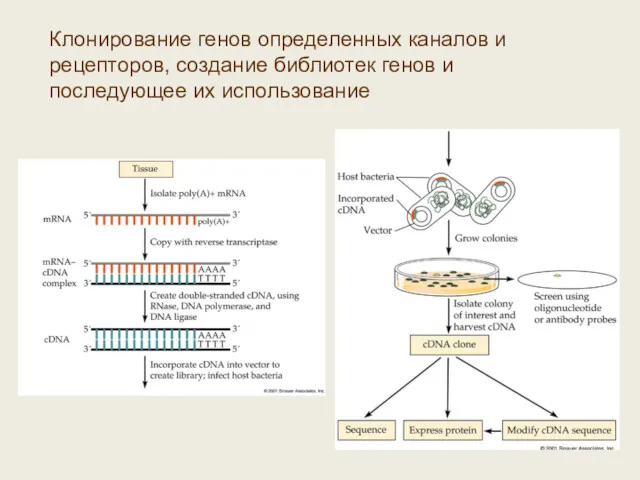
Клонирование генов определенных каналов и рецепторов, создание библиотек генов и последующее
Клонирование генов определенных каналов и рецепторов, создание библиотек генов и последующее
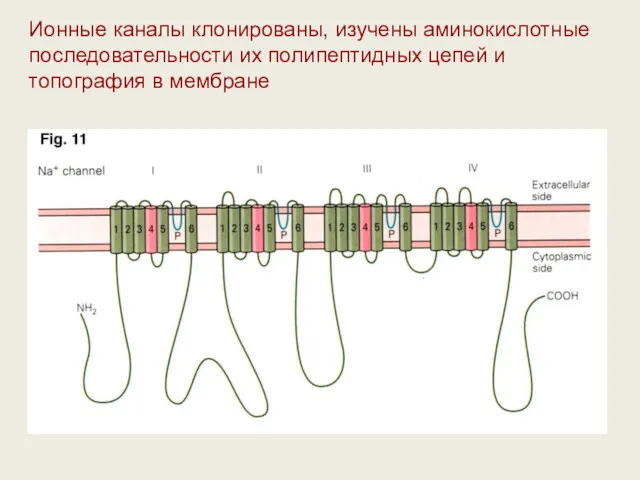
Ионные каналы клонированы, изучены аминокислотные последовательности их полипептидных цепей и топография
Ионные каналы клонированы, изучены аминокислотные последовательности их полипептидных цепей и топография
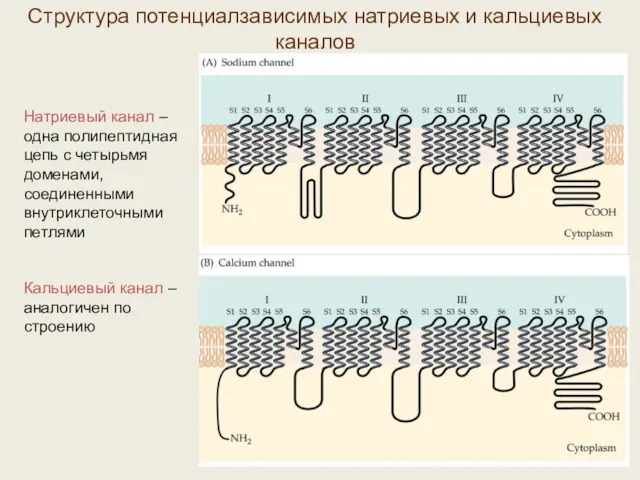
Структура потенциалзависимых натриевых и кальциевых каналов
Натриевый канал – одна полипептидная цепь
Структура потенциалзависимых натриевых и кальциевых каналов
Натриевый канал – одна полипептидная цепь
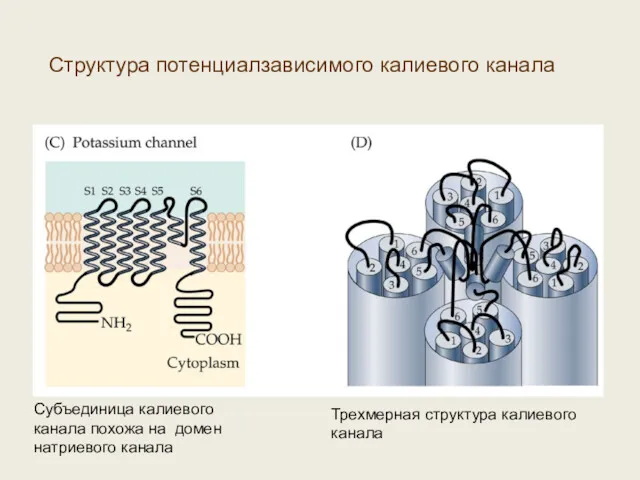
Субъединица калиевого канала похожа на домен натриевого канала
Трехмерная структура калиевого канала
Структура
Субъединица калиевого канала похожа на домен натриевого канала
Трехмерная структура калиевого канала
Структура
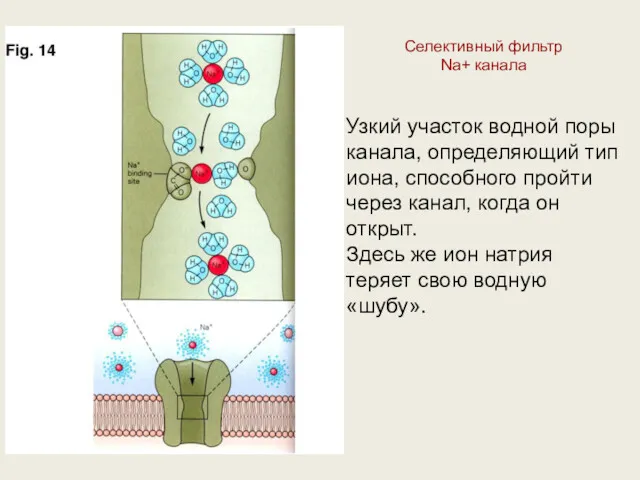
Селективный фильтр
Na+ канала
Узкий участок водной поры
канала, определяющий тип
иона,
Селективный фильтр
Na+ канала
Узкий участок водной поры
канала, определяющий тип
иона,
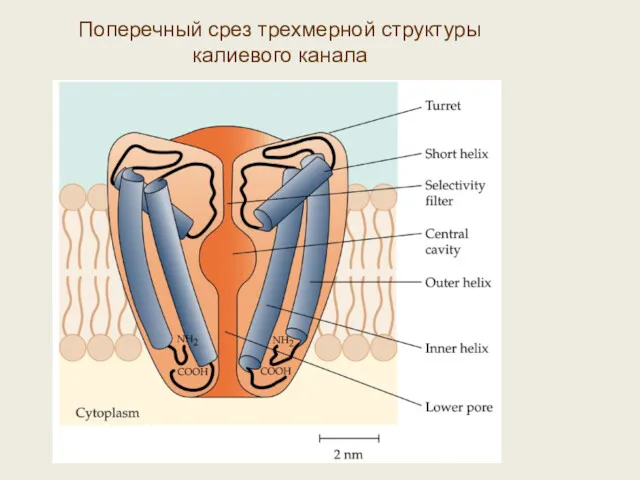
Поперечный срез трехмерной структуры калиевого канала
Поперечный срез трехмерной структуры калиевого канала
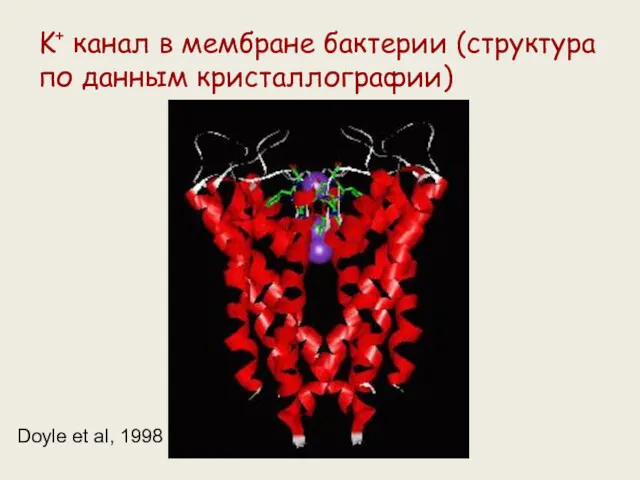
K+ канал в мембране бактерии (структура по данным кристаллографии)
Doyle et al,
K+ канал в мембране бактерии (структура по данным кристаллографии)
Doyle et al,
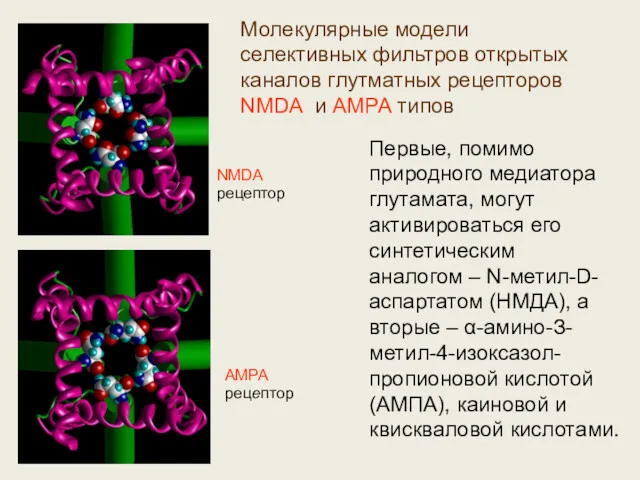
AMPA
рецептор
Молекулярные модели селективных фильтров открытых каналов глутматных рецепторов NMDA и
AMPA
рецептор
Молекулярные модели селективных фильтров открытых каналов глутматных рецепторов NMDA и
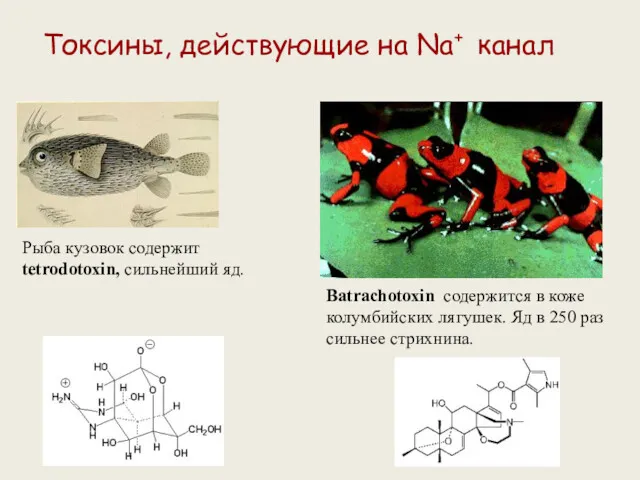
Токсины, действующие на Na+ канал
Рыба кузовок содержит tetrodotoxin, сильнейший яд.
Batrachotoxin содержится
Токсины, действующие на Na+ канал
Рыба кузовок содержит tetrodotoxin, сильнейший яд.
Batrachotoxin содержится

Разделение натриевых и калиевых токов, лежащих в основе потенциала действия
Суммарный ток
Натриевые
Разделение натриевых и калиевых токов, лежащих в основе потенциала действия
Суммарный ток
Натриевые
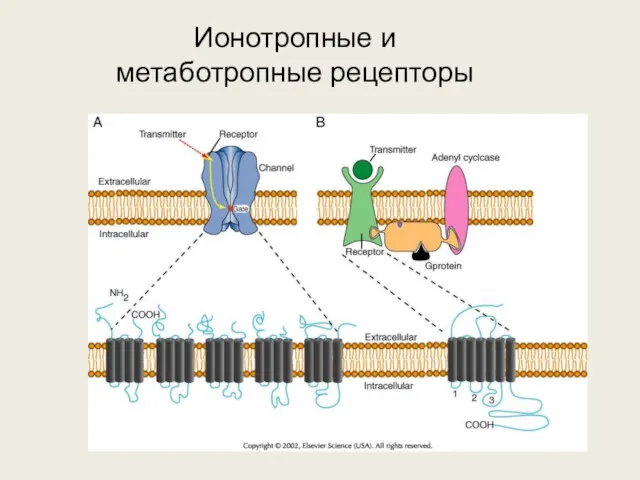
Ионотропные и метаботропные рецепторы
Ионотропные и метаботропные рецепторы
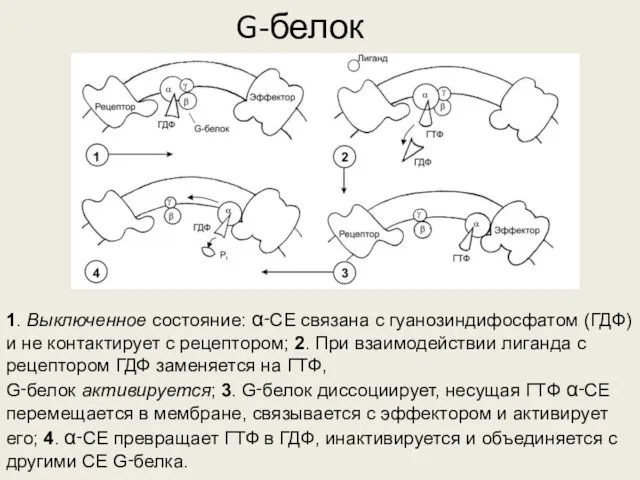
G-белок
1. Выключенное состояние: α‑СЕ связана с гуанозиндифосфатом (ГДФ) и не контактирует с рецептором; 2. При взаимодействии
G-белок
1. Выключенное состояние: α‑СЕ связана с гуанозиндифосфатом (ГДФ) и не контактирует с рецептором; 2. При взаимодействии
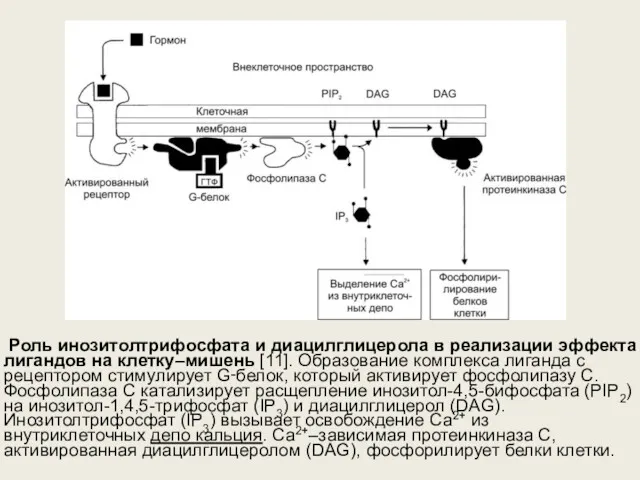
Роль инозитолтрифосфата и диацилглицерола в реализации эффекта лигандов на клетку–мишень [11]. Образование комплекса лиганда с рецептором стимулирует G‑белок, который активирует фосфолипазу С.
Роль инозитолтрифосфата и диацилглицерола в реализации эффекта лигандов на клетку–мишень [11]. Образование комплекса лиганда с рецептором стимулирует G‑белок, который активирует фосфолипазу С.

Вопросы?
Вопросы?

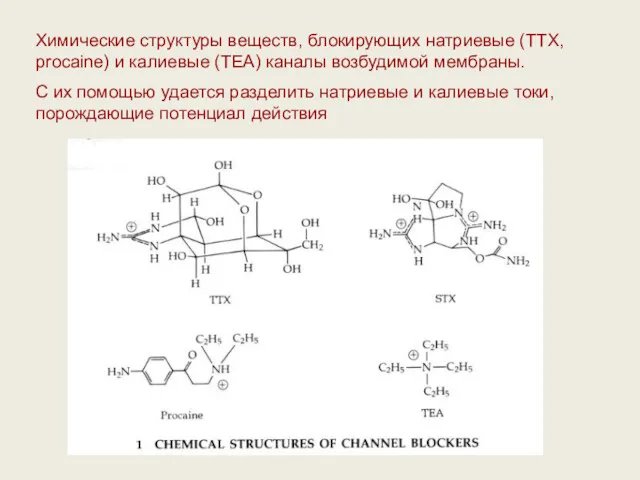
Химические структуры веществ, блокирующих натриевые (TTX, procaine) и калиевые (ТЕА) каналы
Химические структуры веществ, блокирующих натриевые (TTX, procaine) и калиевые (ТЕА) каналы
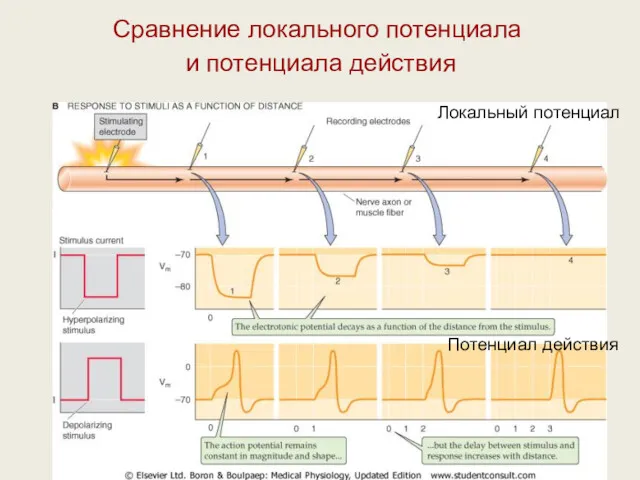
Сравнение локального потенциала
и потенциала действия
Локальный потенциал
Потенциал действия
Сравнение локального потенциала
и потенциала действия
Локальный потенциал
Потенциал действия
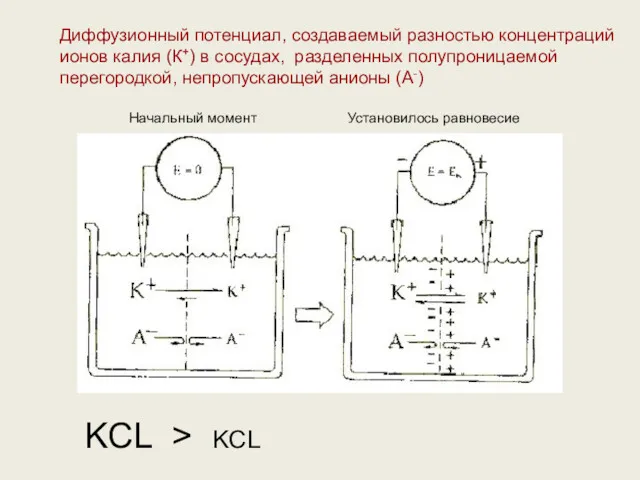
Диффузионный потенциал, создаваемый разностью концентраций ионов калия (К+) в сосудах, разделенных
Диффузионный потенциал, создаваемый разностью концентраций ионов калия (К+) в сосудах, разделенных
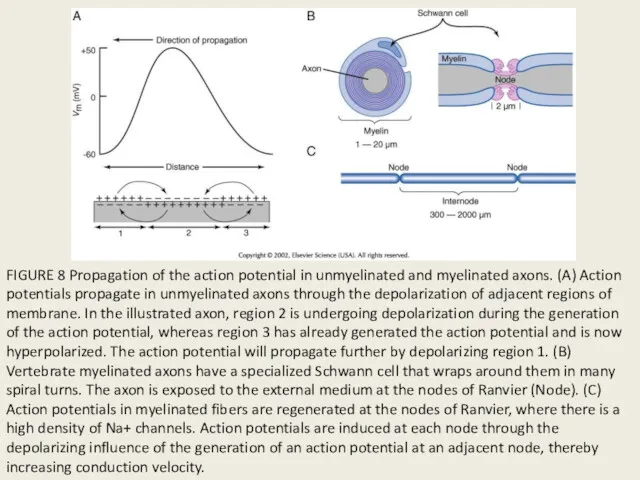
FIGURE 8 Propagation of the action potential in unmyelinated and myelinated
FIGURE 8 Propagation of the action potential in unmyelinated and myelinated
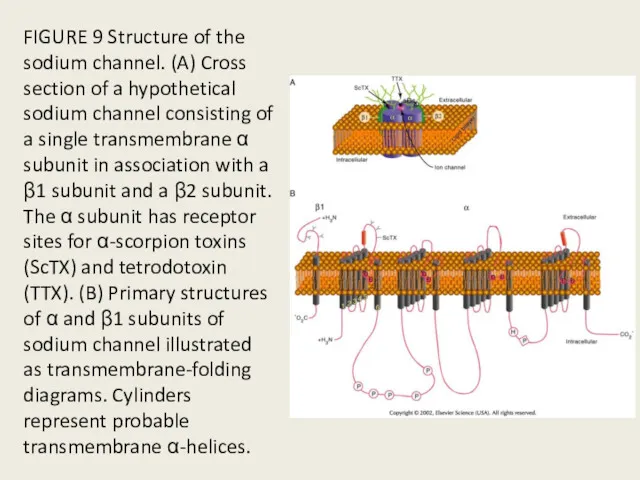
FIGURE 9 Structure of the sodium channel. (A) Cross section of
FIGURE 9 Structure of the sodium channel. (A) Cross section of

Ионные каналы и рецепторы
Ионные каналы и рецепторы
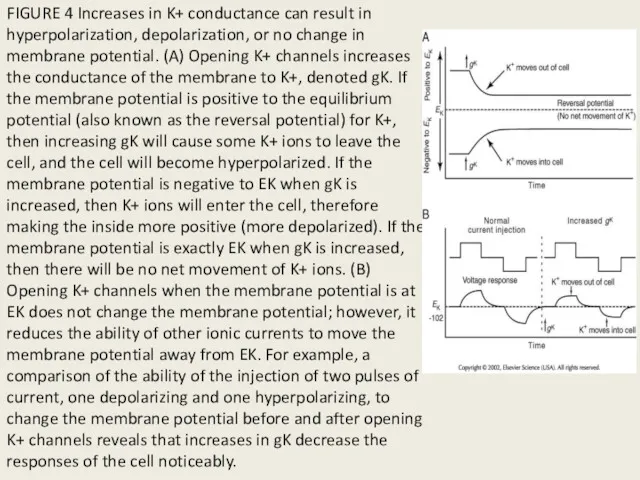
FIGURE 4 Increases in K+ conductance can result in hyperpolarization, depolarization,
FIGURE 4 Increases in K+ conductance can result in hyperpolarization, depolarization,
 Сходства и различия клеток растений, животных и грибов
Сходства и различия клеток растений, животных и грибов Использование дрожжевых грибов для производства разных групп продуктов
Использование дрожжевых грибов для производства разных групп продуктов Открытое мероприятие по профилактике вредных привычек
Открытое мероприятие по профилактике вредных привычек Бактерії - прокаріотичні одноклітинні мікроорганізми
Бактерії - прокаріотичні одноклітинні мікроорганізми Влияние ГМО на организм человека
Влияние ГМО на организм человека Общая физиология сенсорных систем
Общая физиология сенсорных систем Половое поведение животных
Половое поведение животных Дыхание растений
Дыхание растений Круговорот кислорода в природе
Круговорот кислорода в природе Звёздный час. Загадки о животных и растениях
Звёздный час. Загадки о животных и растениях Популяция. Популяционная генетика
Популяция. Популяционная генетика Сахароза. Дисахариды (олигосахариды)
Сахароза. Дисахариды (олигосахариды) Клетки и их разнообразие в многоклеточном организме
Клетки и их разнообразие в многоклеточном организме Основы гистологии. Виды тканей
Основы гистологии. Виды тканей An introduction to metabolism
An introduction to metabolism Глюкоза. Молекулярна формула. Фізичні та хімічні властивості глюкози. Поширення в природі
Глюкоза. Молекулярна формула. Фізичні та хімічні властивості глюкози. Поширення в природі 20231101_vestniki_vesny_viktorina
20231101_vestniki_vesny_viktorina Глаз
Глаз Cattle breeds
Cattle breeds Как зимой помочь птицам (урок окружающего мира, 1 класс)
Как зимой помочь птицам (урок окружающего мира, 1 класс) Эффективный способ посадки картофеля
Эффективный способ посадки картофеля Мы говорим Нет алкоголю!
Мы говорим Нет алкоголю! Физиология растений. Механизмы поступления воды в растительную клетку. Дальний транспорт воды в растении
Физиология растений. Механизмы поступления воды в растительную клетку. Дальний транспорт воды в растении Презентация по биологии для 9 класса по теме:Метаболизм. Энергетический обмен.
Презентация по биологии для 9 класса по теме:Метаболизм. Энергетический обмен. Животные Антарктиды
Животные Антарктиды Красная книга Удмуртии. Животные
Красная книга Удмуртии. Животные Кератин в волосах и восстановление волос
Кератин в волосах и восстановление волос Катархей. Архей
Катархей. Архей