Физиология растений. Механизмы поступления воды в растительную клетку. Дальний транспорт воды в растении презентация
- Физиология растений. Механизмы поступления воды в растительную клетку. Дальний транспорт воды в растении

Содержание
- 2. Механизмы поступления воды в растительную клетку. Дальний транспорт воды в растении. План лекции: - понятия осмоса,
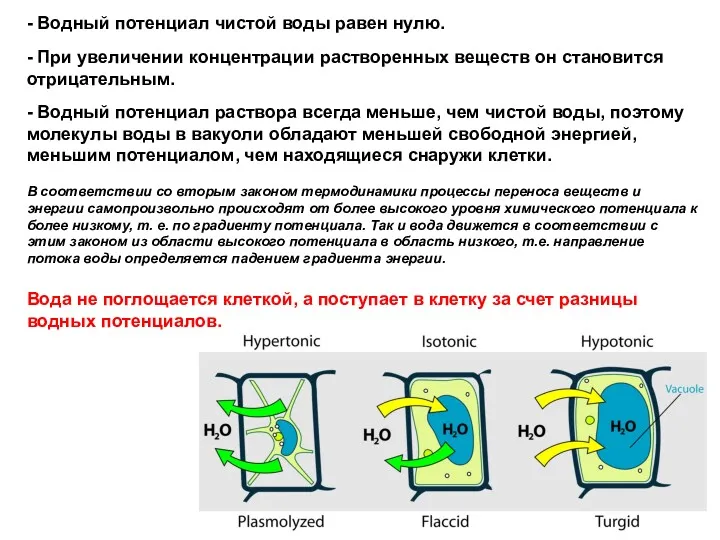
- 3. Гипертонический раствор Состояние протоплазмы зависит от активности воды Изотонический раствор Гипотонический раствор Плазмолиз Нормальное состояние Тургесцентное
- 4. Гипертонический раствор Состояние протоплазмы зависит от активности воды НОРМА: Гипотонический раствор (ПОЧВА, АПОПЛАСТ) клетки лука (внешний
- 5. Гипертонический раствор Состояние протоплазмы зависит от активности воды Изотонический раствор Гипотонический раствор Плазмолиз Нормальное состояние Тургесцентное
- 6. Как движется вода? Каков механизм? Почему так быстро? Какой белок/ген регулирует этот процесс? Этим занимается раздел
- 7. эндоцитоз экзоцитоз Удивительно, но сейчас есть данные и об активном транспорте воды Это только эндоцитоз -
- 8. Некоторые фундаментальные физические основы процессов движения воды в клетке и организме. Химический потенциал воды μw -
- 9. Химический потенциал воды μw выражает максимальное количество внутренней энергии молекул воды, которое может быть превращено в
- 10. - Активность чистой воды равна единице, а внутри клетки в растворах aw меньше единицы, поэтому величина
- 11. Водный потенциал характеризует способность воды диффундировать, испаряться или поглощаться (и чем он выше, тем выше эта
- 12. - Водный потенциал чистой воды равен нулю. - При увеличении концентрации растворенных веществ он становится отрицательным.
- 13. Водный потенциал клетки, ткани или органа, целого растения – величина интегральная (комбинированная). Он состоит из осмотического
- 14. Матричный потенциал Ψm Связан со снижением активности воды за счет гидратации коллоидных веществ и адсорбции на
- 15. Осмотический потенциал Ψр обусловлен наличием в клетках осмотически активных веществ и отражает их влияние на активность
- 16. Тургорный потенциал Ψt обусловлен эластичным противодействием клеточной оболочки разбуханию и характеризует влияние на активность воды тургорного
- 17. Поскольку в большинстве случаев величины как матричного, так и гравитационного потенциалов малы, то Ψw = Ψр+
- 18. Транспорт воды через мембраны клетки осуществляют специальные мембранные белки аквапорины, формирующие в мембране селективные для воды
- 19. Транспорт воды через мембраны клетки осуществляют специальные мембранные белки аквапорины
- 20. снаружи цитоплазма Селективная к воде пора Одна субъединица 1 нм Аквапорин имеет тетрамерное строение, т.е. состоит
- 21. Современная модель строения аквапоринов:
- 22. Клеточная биология аквапоринов: Аквапорины у растений составляют до 2-3% от общего экстрагируемого белка. Принадлежат к так-называемой
- 23. MIPs - транспортируют Небольшие нейтральные молекулы – воду, глицерол, мочевину
- 24. Аквапорины регулируются фосфорилированием как минимум трех сайтов. Фосфорилирование приводит к активации аквапорина. Водная проницаместь увеличивается в
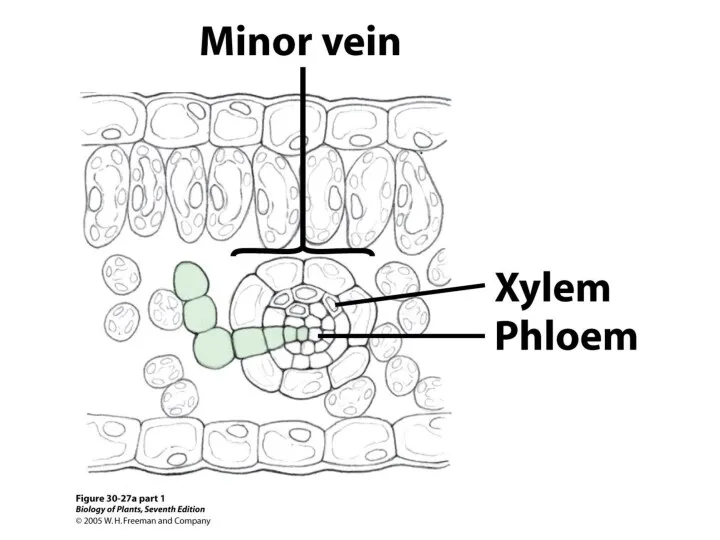
- 25. Как вода движется в надземную часть растения? Что является проводящей тканью?
- 26. Что случилось с этим насекомым?
- 27. Перфориро- ванная пластинка Сосудистый элемент Трахейные (сосудистые элементы с перфорированными кл. ст. На концах) трахеиды отверстия
- 28. Флоэма Ситовидно-трубочный элемент (слева) и клетка-спутник (Т.Э.М.) 3 мм Ситовидно-трубочные элементы: Продольный разрез (С.М.) Ситовидная пластинка
- 30. Между сосудами возможен обмен водой.
- 31. Листья (верхний концевой «двигатель») Градиент воды (направление осмоса и корневого давления) Градиент органических веществ (направление диффузии)
- 32. Радиальный транспорт между почвенным раствором и ксилемой происходит в корне двумя путями. Какова их доля? плазмодесмы
- 33. Апопласти-ческий путь Поясок Каспари Клетка эндодермы Симпласти-ческий путь Поясок Каспари Плазматическая мембрана Апопластический путь Симпласти-ческий путь
- 34. Системы, которые создают корневое давление и ответственны за поднятие корневого водного раствора (пасоки) вверх по сосудам
- 35. Верхний концевой двигатель – системы, обеспечивающие присасывающую силу листьев вследствие испарения воды через устьица и снижения
- 36. Транспирация - физиологический процесс испарения воды наземными органами растений Основным органом транспирации является лист. Сверху и
- 37. Транспирация. У большинства растений верхняя эпидерма имеет меньшее количество устьиц (20–100 шт. на 1 мм2 поверхности),
- 39. В испарении принимают участие три структуры: Устьица – поры, через которые диффундирует вода, испаряющаяся с поверхности
- 40. Интенсивность (скорость) транспирации определяется количеством граммов воды, испаренной с 1 м2 листовой поверхности за 1 час
- 41. Замыкающие клетки устьиц – главные регуляторы траспирации (присасывающего верхнего концевого двигателя) Около 95% воды выносится из
- 42. Воздух Ψ = −100.0 Mpa Лист (межклетник) Ψ = −7.0 Mpa Ксилемный сок (пасока) Клетки мезофилла
- 43. Корневые волоски - клеточные выросты, ответственные за всасывание воды
- 44. Эпидермальные клетки корня представлены (примерно 1:1) трихобластами (образующими корневые волоски) и атрихобластами (необразующими корневые волоски) 100
- 45. Корневые волоски - окраска клеточной стенки (Tinopal Blue)
- 46. Корневые волоски арабидопсиса могут разрастаться при недостатке влаги! Реакция в течение 1 часа!
- 48. Корневые волоски способны к поиску воды в почве!
- 49. Г у т т а ц и я. Физиологический процесс испарения воды наземными органами растений -
- 50. Гуттация, “слёзы растений”, это выделение воды из листьев при избыточном корневом давлении и низком уровне транспирации
- 52. Скачать презентацию

Механизмы поступления воды в растительную клетку. Дальний транспорт воды в растении.
План
Механизмы поступления воды в растительную клетку. Дальний транспорт воды в растении.
План
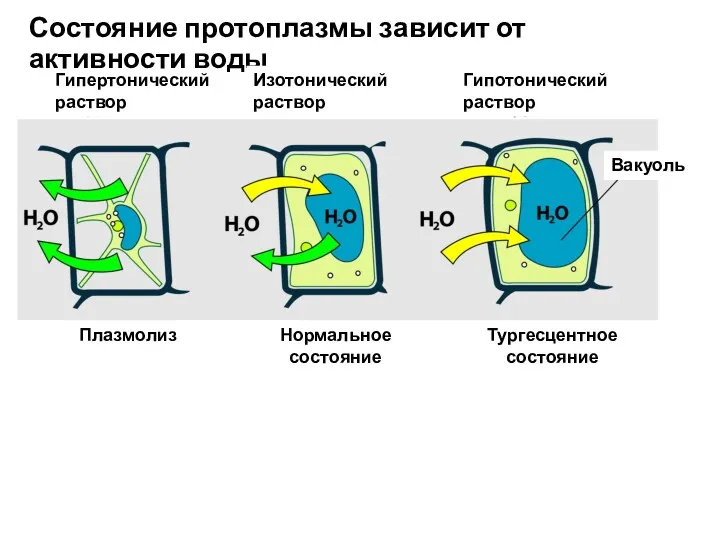
Гипертонический
раствор
Состояние протоплазмы зависит от активности воды
Изотонический
раствор
Гипотонический
раствор
Плазмолиз
Нормальное
состояние
Гипертонический
раствор
Состояние протоплазмы зависит от активности воды
Изотонический
раствор
Гипотонический
раствор
Плазмолиз
Нормальное
состояние
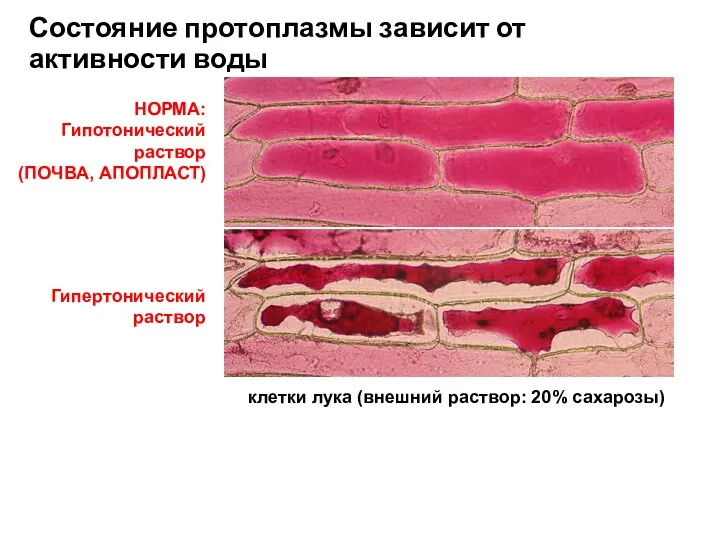
Гипертонический
раствор
Состояние протоплазмы зависит от активности воды
НОРМА:
Гипотонический
раствор
(ПОЧВА, АПОПЛАСТ)
клетки лука (внешний раствор: 20%
Гипертонический
раствор
Состояние протоплазмы зависит от активности воды
НОРМА:
Гипотонический
раствор
(ПОЧВА, АПОПЛАСТ)
клетки лука (внешний раствор: 20%
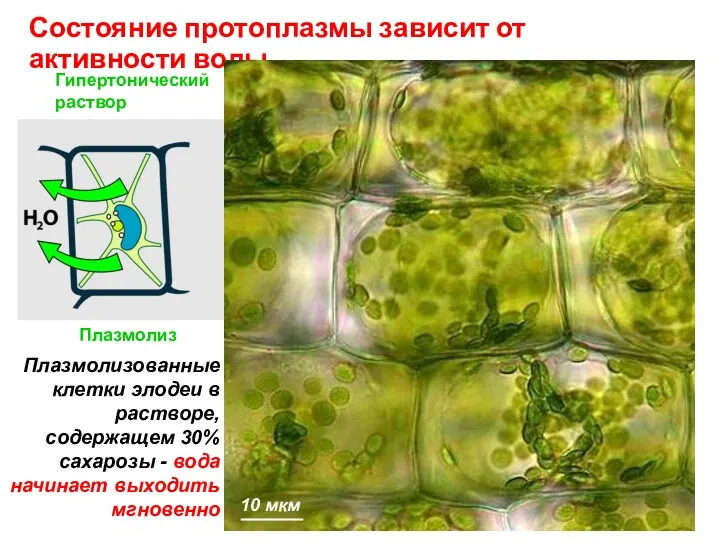
Гипертонический
раствор
Состояние протоплазмы зависит от активности воды
Изотонический
раствор
Гипотонический
раствор
Плазмолиз
Нормальное
состояние
Гипертонический
раствор
Состояние протоплазмы зависит от активности воды
Изотонический
раствор
Гипотонический
раствор
Плазмолиз
Нормальное
состояние
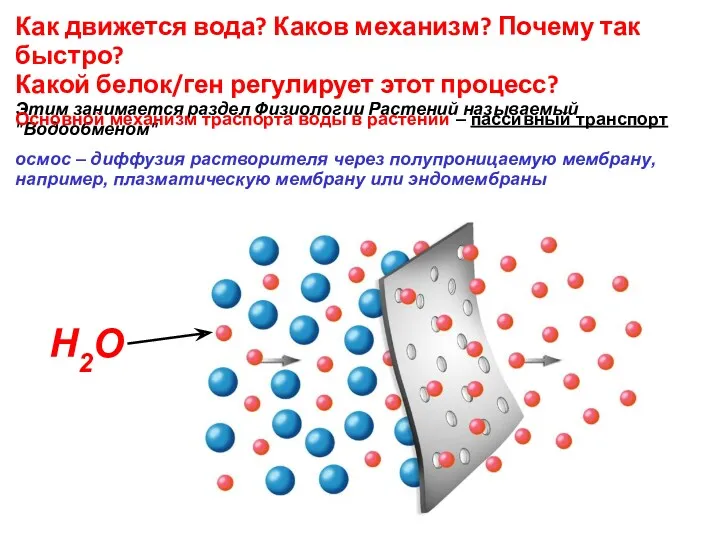
Как движется вода? Каков механизм? Почему так быстро?
Какой белок/ген регулирует
Как движется вода? Каков механизм? Почему так быстро?
Какой белок/ген регулирует
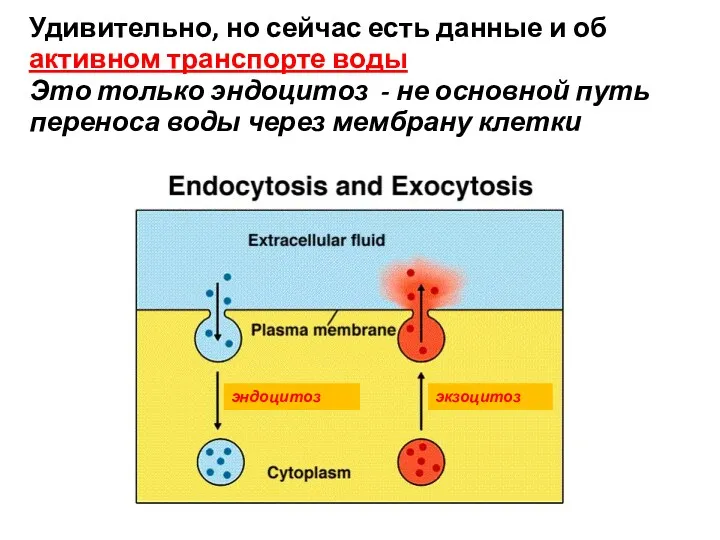
эндоцитоз
экзоцитоз
Удивительно, но сейчас есть данные и об
активном транспорте воды
Это только
эндоцитоз
экзоцитоз
Удивительно, но сейчас есть данные и об
активном транспорте воды
Это только

Некоторые фундаментальные физические основы процессов движения воды в клетке и организме.
Химический
Некоторые фундаментальные физические основы процессов движения воды в клетке и организме.
Химический

Химический потенциал воды μw
выражает максимальное количество внутренней энергии молекул воды, которое
Химический потенциал воды μw
выражает максимальное количество внутренней энергии молекул воды, которое

- Активность чистой воды равна единице, а внутри клетки в растворах
- Активность чистой воды равна единице, а внутри клетки в растворах

Водный потенциал характеризует способность воды диффундировать, испаряться или поглощаться (и чем
Водный потенциал характеризует способность воды диффундировать, испаряться или поглощаться (и чем
- Водный потенциал чистой воды равен нулю.
- При увеличении концентрации
- Водный потенциал чистой воды равен нулю.
- При увеличении концентрации

Водный потенциал клетки, ткани или органа, целого растения – величина интегральная
Водный потенциал клетки, ткани или органа, целого растения – величина интегральная
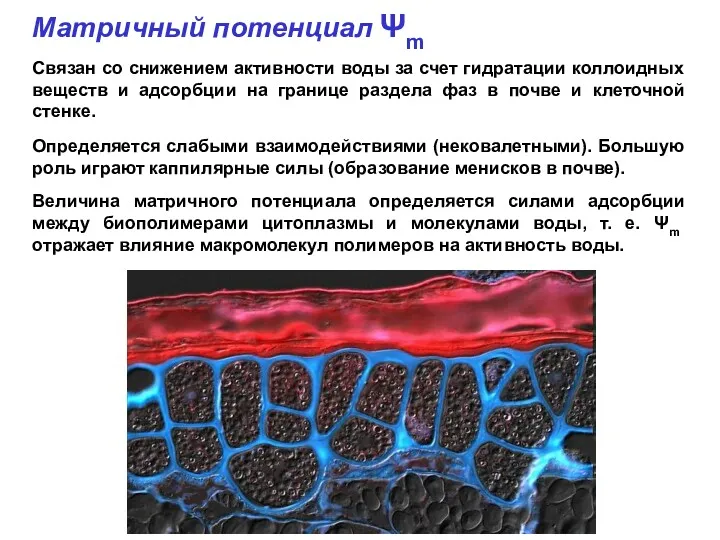
Матричный потенциал Ψm
Связан со снижением активности воды за счет гидратации
Матричный потенциал Ψm
Связан со снижением активности воды за счет гидратации

Осмотический потенциал Ψр обусловлен наличием в клетках осмотически активных веществ и
Осмотический потенциал Ψр обусловлен наличием в клетках осмотически активных веществ и

Тургорный потенциал Ψt обусловлен эластичным противодействием клеточной оболочки разбуханию и характеризует
Тургорный потенциал Ψt обусловлен эластичным противодействием клеточной оболочки разбуханию и характеризует

Поскольку в большинстве случаев величины как матричного, так и гравитационного потенциалов
Поскольку в большинстве случаев величины как матричного, так и гравитационного потенциалов

Транспорт воды через мембраны клетки осуществляют специальные мембранные белки аквапорины,
формирующие
Транспорт воды через мембраны клетки осуществляют специальные мембранные белки аквапорины,
формирующие

Транспорт воды через мембраны клетки осуществляют специальные мембранные белки аквапорины
Транспорт воды через мембраны клетки осуществляют специальные мембранные белки аквапорины
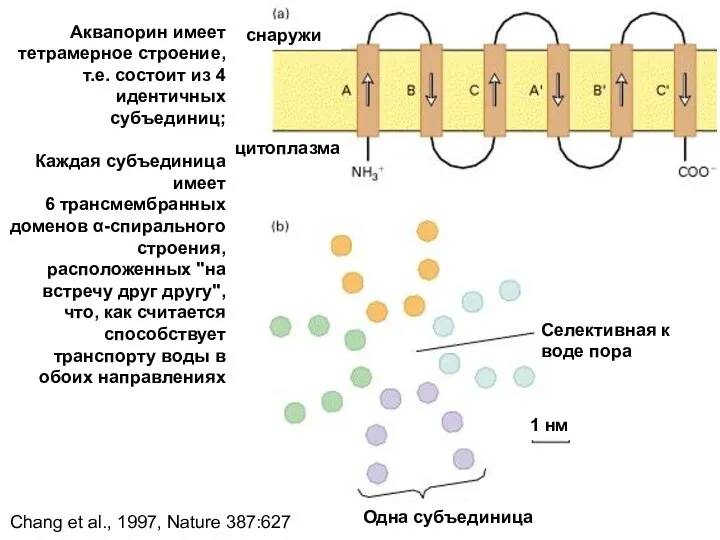
снаружи
цитоплазма
Селективная к воде пора
Одна субъединица
1 нм
Аквапорин имеет
тетрамерное строение, т.е. состоит
снаружи
цитоплазма
Селективная к воде пора
Одна субъединица
1 нм
Аквапорин имеет
тетрамерное строение, т.е. состоит
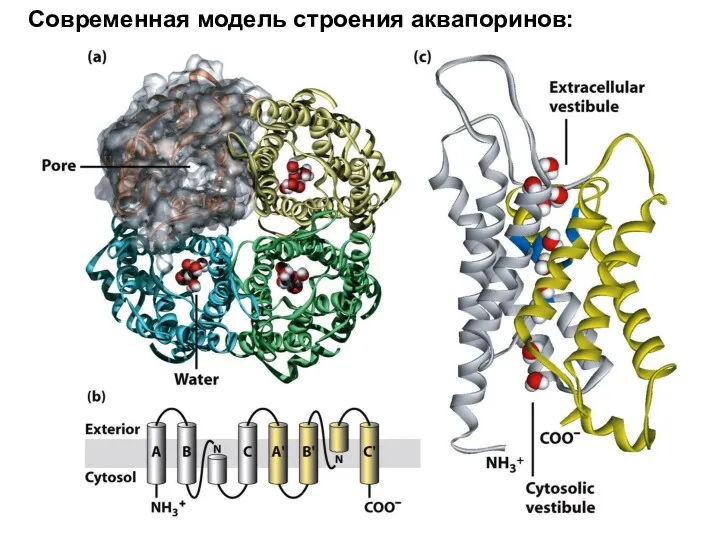
Современная модель строения аквапоринов:
Современная модель строения аквапоринов:

Клеточная биология аквапоринов:
Аквапорины у растений составляют до 2-3% от общего экстрагируемого
Клеточная биология аквапоринов:
Аквапорины у растений составляют до 2-3% от общего экстрагируемого
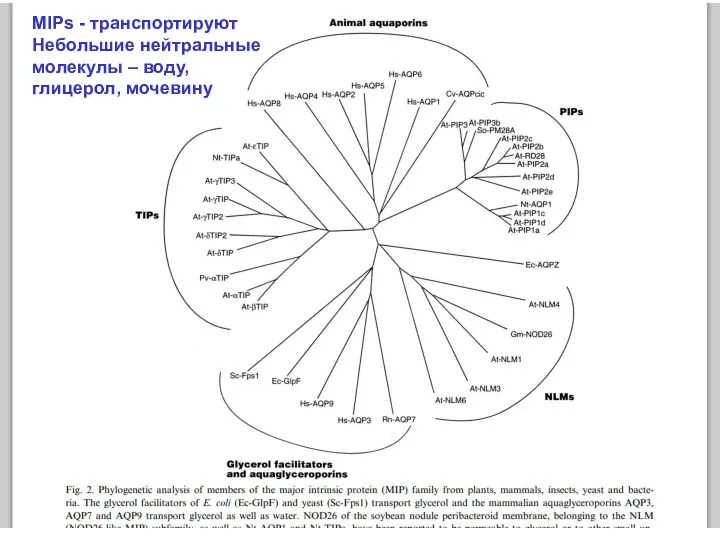
MIPs - транспортируют
Небольшие нейтральные
молекулы – воду,
глицерол, мочевину
MIPs - транспортируют
Небольшие нейтральные
молекулы – воду,
глицерол, мочевину
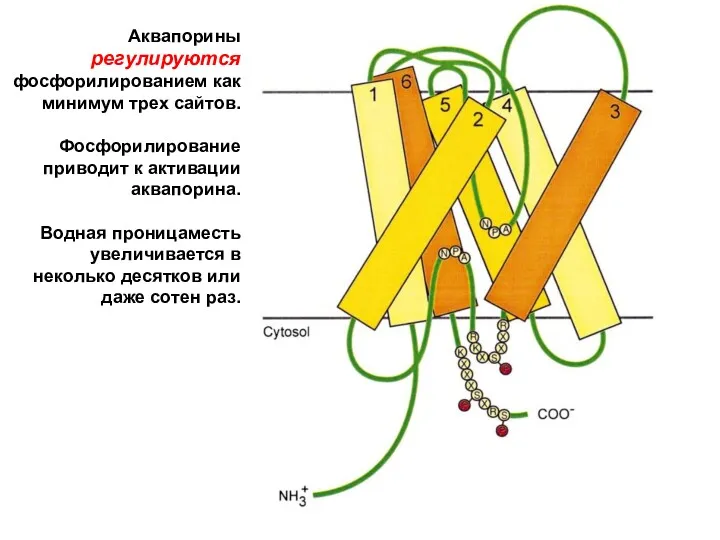
Аквапорины регулируются фосфорилированием как минимум трех сайтов.
Фосфорилирование приводит к активации аквапорина.
Фосфорилирование приводит к активации аквапорина.

Как вода движется в
надземную часть
растения?
Что является
проводящей
тканью?
Как вода движется в
надземную часть
растения?
Что является
проводящей
тканью?

Что случилось с
этим насекомым?
Что случилось с
этим насекомым?
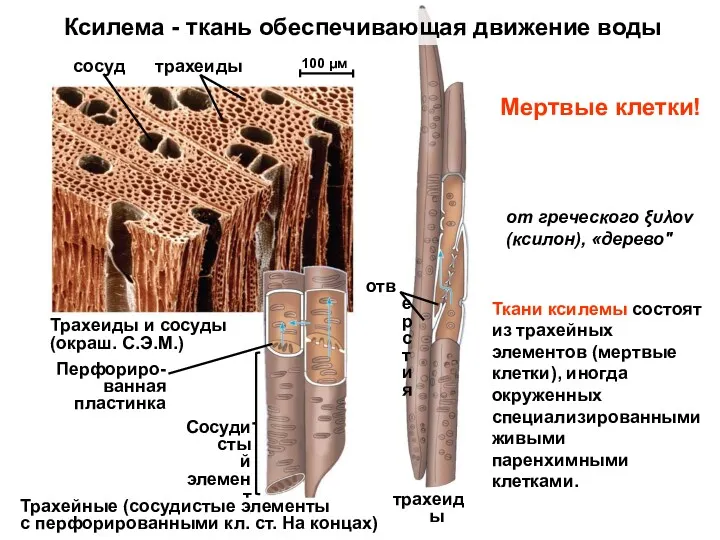
Перфориро-
ванная
пластинка
Сосудистый
элемент
Трахейные (сосудистые элементы
с перфорированными кл. ст. На концах)
трахеиды
отверстия
Трахеиды и
Перфориро-
ванная
пластинка
Сосудистый
элемент
Трахейные (сосудистые элементы
с перфорированными кл. ст. На концах)
трахеиды
отверстия
Трахеиды и
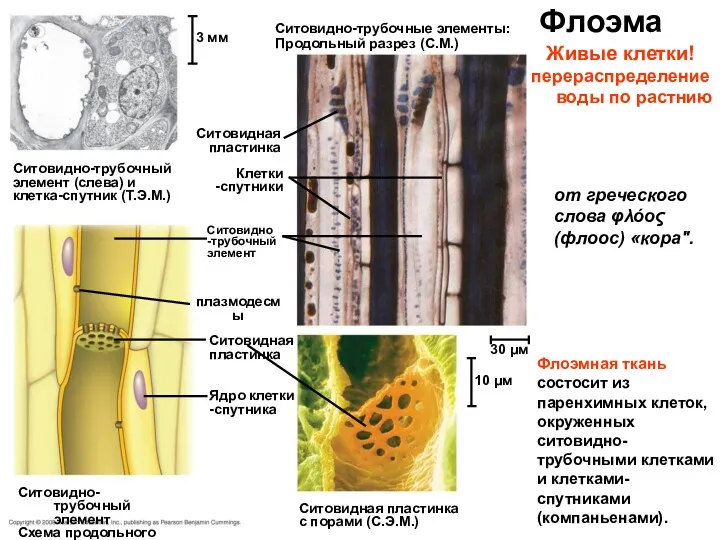
Флоэма
Ситовидно-трубочный
элемент (слева) и
клетка-спутник (Т.Э.М.)
3 мм
Ситовидно-трубочные элементы:
Продольный разрез (С.М.)
Ситовидная
пластинка
Клетки
-спутники
Ситовидно
-трубочный
элемент
плазмодесмы
Ситовидная
Флоэма
Ситовидно-трубочный
элемент (слева) и
клетка-спутник (Т.Э.М.)
3 мм
Ситовидно-трубочные элементы:
Продольный разрез (С.М.)
Ситовидная
пластинка
Клетки
-спутники
Ситовидно
-трубочный
элемент
плазмодесмы
Ситовидная
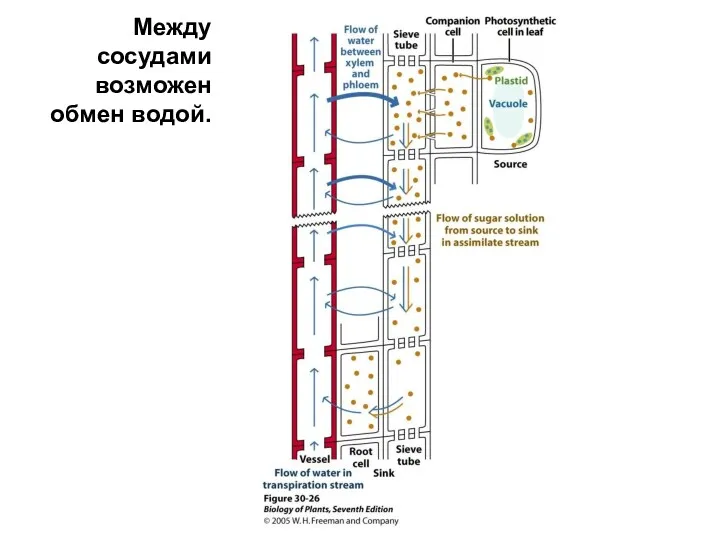
Между сосудами возможен обмен водой.
Между сосудами возможен обмен водой.
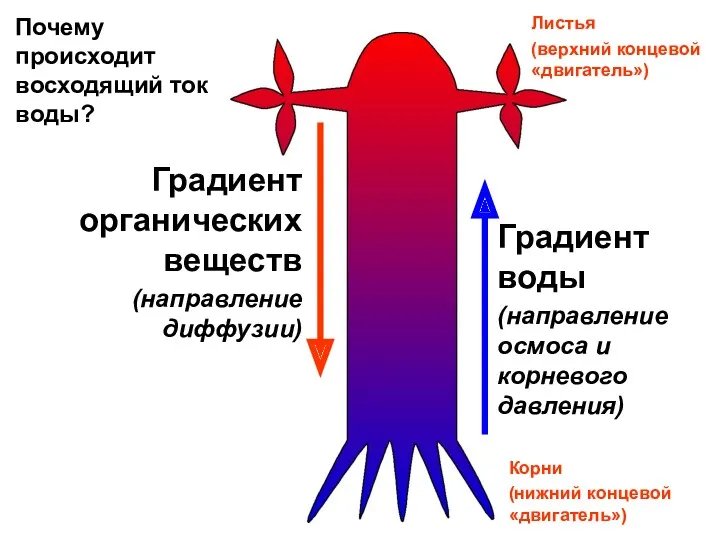
Листья
(верхний концевой «двигатель»)
Градиент воды
(направление осмоса и корневого давления)
Градиент органических веществ
(направление диффузии)
Почему
Листья
(верхний концевой «двигатель»)
Градиент воды
(направление осмоса и корневого давления)
Градиент органических веществ
(направление диффузии)
Почему
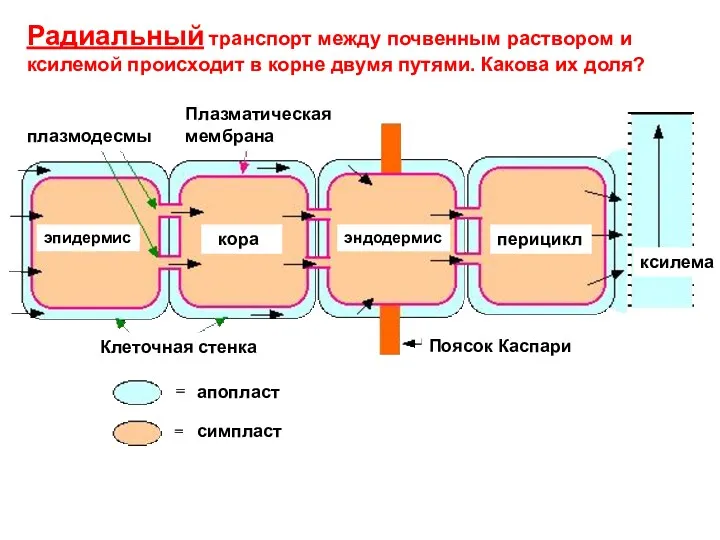
Радиальный транспорт между почвенным раствором и ксилемой происходит в корне двумя
Радиальный транспорт между почвенным раствором и ксилемой происходит в корне двумя
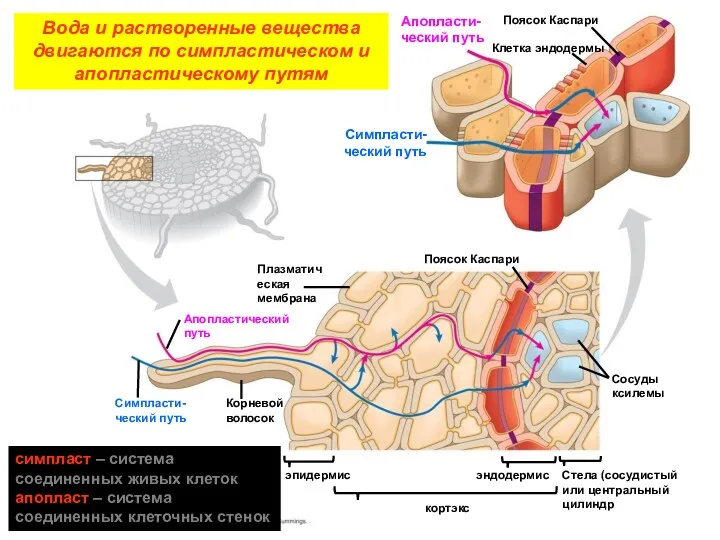
Апопласти-ческий путь
Поясок Каспари
Клетка эндодермы
Симпласти-ческий путь
Поясок Каспари
Плазматическая мембрана
Апопластический путь
Симпласти-ческий путь
Корневой волосок
эпидермис
кортэкс
эндодермис
Сосуды ксилемы
Стела
Апопласти-ческий путь
Поясок Каспари
Клетка эндодермы
Симпласти-ческий путь
Поясок Каспари
Плазматическая мембрана
Апопластический путь
Симпласти-ческий путь
Корневой волосок
эпидермис
кортэкс
эндодермис
Сосуды ксилемы
Стела

Системы, которые создают корневое давление и ответственны за поднятие корневого водного
Системы, которые создают корневое давление и ответственны за поднятие корневого водного
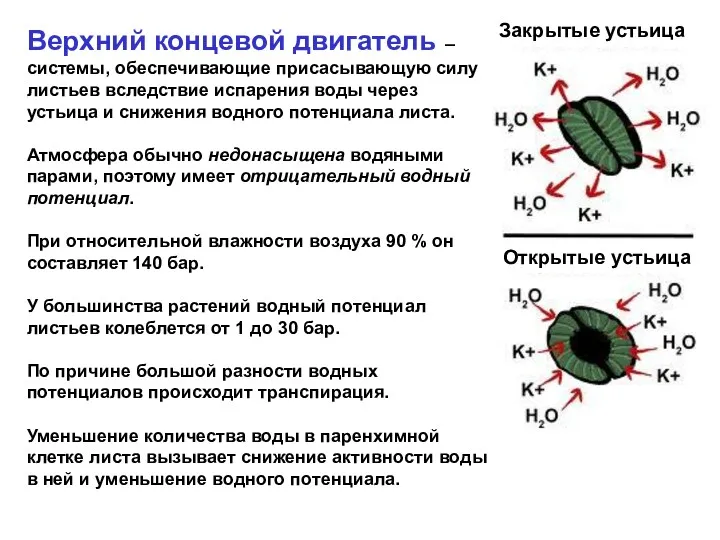
Верхний концевой двигатель – системы, обеспечивающие присасывающую силу листьев вследствие испарения
Верхний концевой двигатель – системы, обеспечивающие присасывающую силу листьев вследствие испарения
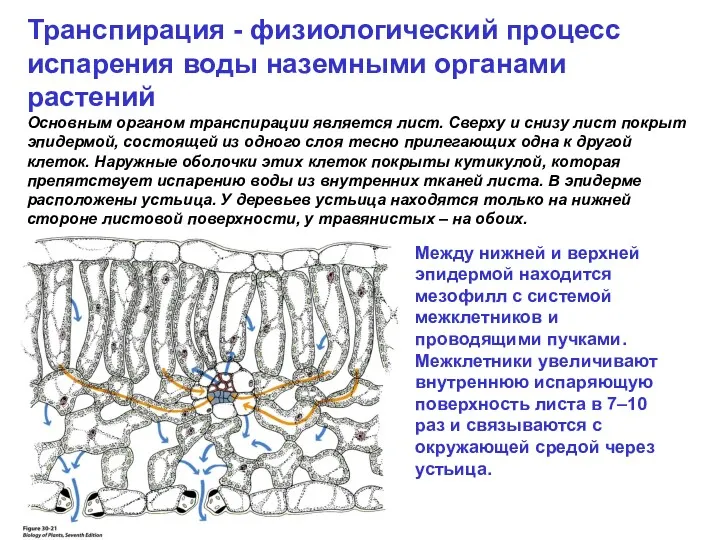
Транспирация - физиологический процесс испарения воды наземными органами растений
Основным органом транспирации
Транспирация - физиологический процесс испарения воды наземными органами растений
Основным органом транспирации

Транспирация.
У большинства растений верхняя эпидерма имеет меньшее количество устьиц (20–100
Транспирация.
У большинства растений верхняя эпидерма имеет меньшее количество устьиц (20–100


В испарении принимают участие три структуры:
Устьица – поры, через которые диффундирует
В испарении принимают участие три структуры:
Устьица – поры, через которые диффундирует

Интенсивность (скорость) транспирации определяется количеством граммов воды, испаренной с 1 м2
Интенсивность (скорость) транспирации определяется количеством граммов воды, испаренной с 1 м2

Замыкающие клетки устьиц – главные регуляторы траспирации (присасывающего верхнего концевого двигателя)
Около
Замыкающие клетки устьиц – главные регуляторы траспирации (присасывающего верхнего концевого двигателя)
Около
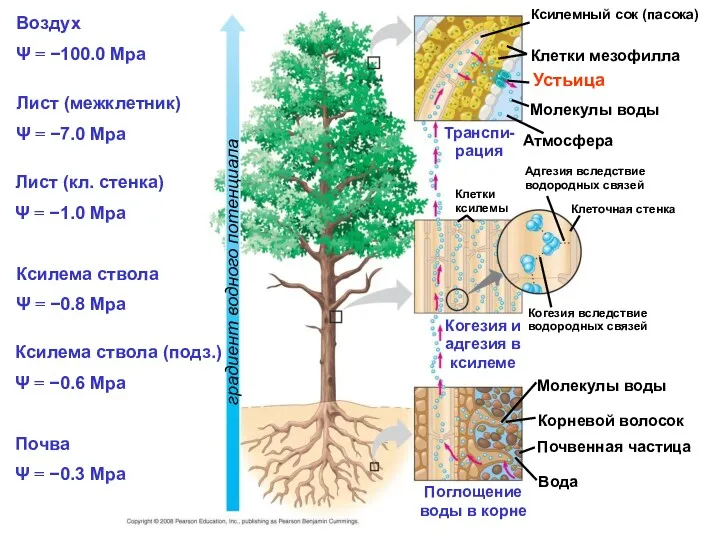
Воздух
Ψ = −100.0 Mpa
Лист (межклетник)
Ψ = −7.0 Mpa
Ксилемный сок (пасока)
Клетки
Воздух
Ψ = −100.0 Mpa
Лист (межклетник)
Ψ = −7.0 Mpa
Ксилемный сок (пасока)
Клетки
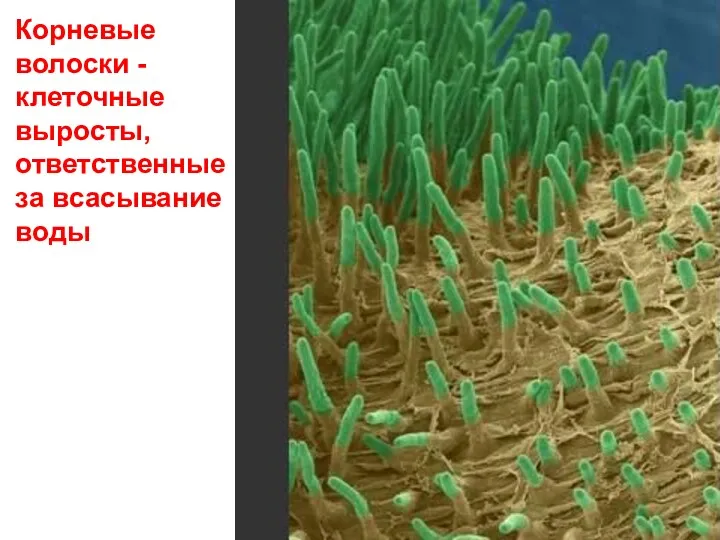
Корневые
волоски -
клеточные выросты, ответственные
за всасывание воды
Корневые
волоски -
клеточные выросты, ответственные
за всасывание воды
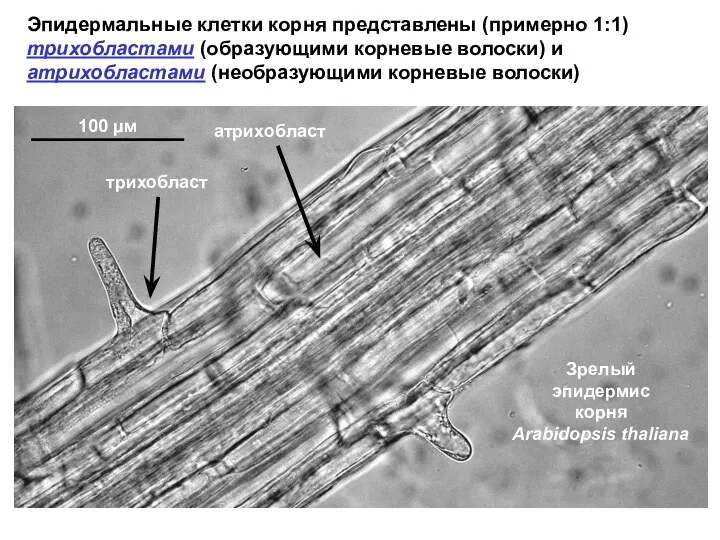
Эпидермальные клетки корня представлены (примерно 1:1)
трихобластами (образующими корневые волоски) и атрихобластами
Эпидермальные клетки корня представлены (примерно 1:1)
трихобластами (образующими корневые волоски) и атрихобластами

Корневые
волоски - окраска клеточной стенки (Tinopal Blue)
Корневые
волоски - окраска клеточной стенки (Tinopal Blue)

Корневые волоски арабидопсиса могут разрастаться при недостатке влаги!
Реакция в течение 1
Корневые волоски арабидопсиса могут разрастаться при недостатке влаги!
Реакция в течение 1

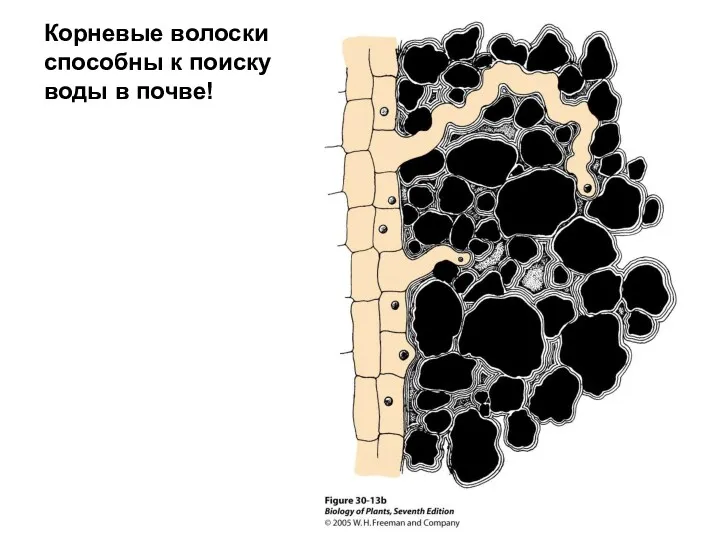
Корневые волоски способны к поиску воды в почве!
Корневые волоски способны к поиску воды в почве!

Г у т т а ц и я.
Физиологический процесс испарения воды
Г у т т а ц и я.
Физиологический процесс испарения воды

Гуттация, “слёзы растений”, это выделение воды из листьев
при избыточном корневом давлении
Гуттация, “слёзы растений”, это выделение воды из листьев
при избыточном корневом давлении
 Световая микроскопия
Световая микроскопия Генерация ПД в нейронах
Генерация ПД в нейронах Семейство лошадиные (equidae)
Семейство лошадиные (equidae) Одноклеточные животные
Одноклеточные животные Иондаушы сәулелер көздерімен жұмыстың санитарлық ережелері
Иондаушы сәулелер көздерімен жұмыстың санитарлық ережелері Cellular neurophysiology
Cellular neurophysiology презентация к уроку биологии в 5 классе по теме Покрытосеменные
презентация к уроку биологии в 5 классе по теме Покрытосеменные Соцветия
Соцветия Роль покрытосеменных в жизни человека
Роль покрытосеменных в жизни человека Анатомия нервной системы
Анатомия нервной системы Генетика – наука о закономерностях наследственности и изменчивости
Генетика – наука о закономерностях наследственности и изменчивости Дихальна система
Дихальна система Вітаміни та ферменти
Вітаміни та ферменти Птахи, народжені взимку
Птахи, народжені взимку Животное, занесённое в красную книгу – манул
Животное, занесённое в красную книгу – манул Что такое адаптации? Какие бывают адаптации?
Что такое адаптации? Какие бывают адаптации? Сохранение и восстановление биоразнообразия на популяционно-видовом уровне
Сохранение и восстановление биоразнообразия на популяционно-видовом уровне Неорганические вещества клетки
Неорганические вещества клетки Факторы устойчивости инфекционных заболеваний пчел
Факторы устойчивости инфекционных заболеваний пчел Ритмы здоровья. Витаминно-минеральный комплекс
Ритмы здоровья. Витаминно-минеральный комплекс Органы чувств. Анализаторы. Как действуют органы чувств и анализаторы. 8 класс
Органы чувств. Анализаторы. Как действуют органы чувств и анализаторы. 8 класс Valerian. Description
Valerian. Description Развитие жизни на Земле
Развитие жизни на Земле Физиологические особенности дошкольного возраста
Физиологические особенности дошкольного возраста Проблемы Антропогенеза (фрагмент курса Эволюция Биосферы)
Проблемы Антропогенеза (фрагмент курса Эволюция Биосферы) Нарушение кожных покровов и их причины.8 класс
Нарушение кожных покровов и их причины.8 класс Открытый урок на тему: Многообразие земноводных.
Открытый урок на тему: Многообразие земноводных. Вид. Критерии вида
Вид. Критерии вида