- Элементы физиологии клетки

Содержание
- 2. ВОПРОСЫ Функции клеточных органелл Компоненты мембраны возбудимой клетки как объект изучения физиологии Белки – насосы и
- 3. ОСНОВНЫЕ ПОНЯТИЯ Клетка – это наименьшая функциональная единица тканей и органов животного организма, располагающая специальным набором
- 4. Электровозбудимые клетки: нейроны, миоциты, гландоциты
- 5. Системы Под физиологической системой понимают анатомически детерминированную совокупность органов и тканей, выполняющую определенные функции.
- 6. Интегративные системы нервная, эндокринная, иммунная
- 7. Функциональные системы Функциональная система, согласно теории функциональных систем П.К.Анохина, — динамическая совокупность различных органов и физиологических
- 8. Функциональные системы Функциональные системы поддерживают оптимальные физиологические показатели, обеспечивающие достижение полезного результата — удовлетворение биологических и
- 9. 1.Функции клеточных органелл
- 10. Структура везикулы
- 11. Функции клетки 1. Базовая основа жизни. Мельчайшая часть организма, способная сохранять свойство жизни (в культуре живут
- 12. Основные органеллы и включения
- 13. Структуры клетки человека
- 14. 2. Компоненты мембраны возбудимой клетки как объект изучения физиологии.
- 15. МНОГО ОБЩИХ ЧЕРТ ЛЕЖАТ В ОСНОВЕ РАЗНООБРАЗИЯ БИОЛОГИЧЕСКИХ МЕМБРАН Мембраны столь же разнообразны по структуре, как
- 16. МНОГО ОБЩИХ ЧЕРТ ЛЕЖАТ В ОСНОВЕ РАЗНООБРАЗИЯ БИОЛОГИЧЕСКИХ МЕМБРАН 4. Отличительные функции мембран зависят от набора
- 17. МНОГО ОБЩИХ ЧЕРТ ЛЕЖАТ В ОСНОВЕ РАЗНООБРАЗИЯ БИОЛОГИЧЕСКИХ МЕМБРАН 7. Мембраны - жидкие структуры. Молекулы липида
- 18. Жидкостно-мозаичная модель
- 19. Клетки всех живых организмов окружены внешней мембраной – плазмолеммой, цитолеммой. Большинство клеток содержат мембраны и в
- 20. Липиды Структурную матрицу мембраны составляют липиды, на долю которых приходится от 15 до 50% сухой массы.
- 21. Белки мембраны условно разделяют на периферические и собственные (интегральные). Периферические белки расположены на поверхности липидного бислоя,
- 22. Рецептор связанный с G-белком
- 23. Мембранно связанный энзим
- 24. Внутриклеточный рецептор
- 25. Углеводы Углеводы клеточных мембран присутствуют в плазмолемме в виде соединений с белками (гликопротеиды) и липидами (гликолипиды)
- 26. Транспорт веществ: аквапорин
- 27. Транспорт веществ: диффузия
- 28. Осмос и осмотические нарушения
- 29. Фильтрация и реабсорбция
- 30. Na+К+ помпа: активный транспорт
- 31. Вторично-активный транспорт: симпорт и антипорт, GLUT1-12
- 33. Относительное значение механизмов транспорта веществ
- 34. 3. Белки – насосы и белки – ионные каналы. Насосы
- 35. Классы ионных насосов
- 36. Разные формы цитоза: эндоцитоз
- 37. пиноцитоз
- 38. Рецептор-опосредованный эндоцитоз ЛПНП
- 39. экзоцитоз
- 40. Обобщение видов транспорта через мембрану
- 41. Обобщение видов транспорта через мембрану
- 42. 4.Белки-рецепторы химических и физических сигналов
- 43. Классификация рецепторов Мембранные рецепторы регистрируют наличие лиганда (пептидного гормона, производных аминокислот, нейромедиаторов, цитокинов) и передают сигнал
- 44. Классификация рецепторов 1) рецепторы, трансмембранный сегмент которых состоит из семи фрагментов (петель); 2) рецепторы, трансмембранный сегмент
- 45. К гормонам, рецептор которых состоит из семи трансмембранных фрагментов, относятся: АКТГ, ТТГ, ФСГ, ЛГ, хорионический гонадотропин,
- 46. Ко второй группе относятся гормоны, имеющие один трансмембранный фрагмент: СТГ, пролактин, инсулин, соматомаммотропин, или плацентарный лактоген,
- 47. К гормонам третьей группы, рецептор которых имеет четыре трансмембранных фрагмента, относятся ацетилхолин (никотиновые мышечные и нервные
- 48. Количество имеет значение Количество клеточных рецепторов, как и других компонентов клетки, постоянно изменяется, отражая процессы их
- 49. В представлениях современной молекулярной физиологии клетки, основные процессы в воспринимающих сигнал клетках связаны с фосфорилированием и
- 50. Среди подвергаемых фосфорилированию и дефосфорилированию ферментов имеется набор протеинкиназ, осуществляющих регуляцию функций разнообразных белковых молекул. Обычно
- 51. Поэтому различают сериновые/треониновые протеинкиназы и тирозиновые протеинкиназы. Обратный процесс – дефосфолрилирование обеспечивается тирозиновыми или сериновыми фосфатазами,
- 52. Другой путь, который используется при передаче сигналов между клетками, связан с активацией процессов в клетке, вовлекающих
- 53. Мембранные рецепторы подразделяют на: каталитические, связанные с ионными каналами, являющиеся ионными каналами, оперирующие через G‑белки освобождающие
- 54. Классификация рецепторов
- 55. Каталитические 1. Рецепторные гуанилатциклазы катализируют образование цГМФ из ГТФ. К таким мембранным рецепторным белкам-ферментам относятся 2
- 56. Каталитические 2.Рецепторные серин/треониновые киназы фосфорилируют остатки серина и/или треонина как самого рецептора (автофосфорилирование), так и внутриклеточных
- 57. Каталитические 3.Рецепторные тирозинкиназы фосфорилируют остатки тирозина как самого белка-рецептора (автофосфорилирование), так и разных внутриклеточных белков. К
- 58. Рецептор к инсулину - тирозинкиназа
- 59. Каталитические 4.Рецепторы, связанные с тирозинкиназами, взаимодействуют с цитоплазматическими тирозинкиназами, многие из которых являются протоонкогенами. К этой
- 60. Каталитические 5.Рецепторные тирозинфосфатазы освобождают (дефосфорилируют) фосфатные группировки из фосфорилированных тирозилов внутриклеточных белков. К таким рецепторам относится
- 61. Рецепторы, связанные с ионными каналами, влияют на проницаемость ионных каналов через вторые посредники. К ним относятся
- 62. Управляемые лигандами ионные каналы — интегральные мембранные белки — сочетают свойства рецептора и ионного канала и
- 63. Классификация синаптических рецепторов: ионотропные и метаботропные
- 64. Рецепторы, связанные с G‑белком — полипептидные цепочки, расположенные в плазматической мембране клетки–мишени таким образом, что полипептид
- 66. Лиганды связанных с G‑белком мембранных рецепторов ангиотензин II, АТФ, ацетилхолин (мускариновые рецепторы), нейромедин B, гастрин–рилизинг гормон,
- 67. Ионы асимметрично распределены во внутренней среде
- 68. ИОННЫЕ ГРАДИЕНТЫ НУЖНЫ ДЛЯ: ЗАПАСАНИЯ ЭНЕРГИИ ГЕНЕРАЦИИ ВОЗБУЖДЕНИЯ ТРАНСПОРТА ГЛЮКОЗЫ И АМИНОКИСЛОТ
- 69. Для создания потенциалов нужны ионы Na+, K+, Ca++, Cl- Na+ и K+ существенно отличаются по физико-химическим
- 70. физико-химические отличия ионов 1.Для создания электрохимического потенциала ионов K+ нужно на порядок меньше, чем Na+ 2.У
- 71. физико-химические отличия ионов Из-за высокой гидрофобности K+ проникает через мембрану в 5 раз легче, чем гидрофильный
- 72. физико-химические отличия ионов
- 74. Стехиометрия Для встречного перемещения (антипорта) натрия и калия Na-K-АТФазой характерна стехиометрия, для аксона кальмара равная 3/2.
- 75. 30% метаболизма Калий-натриевая помпа локализована в мембранах практически всех клеток организма, но относительно много молекул –
- 76. Са-АТФазы Кальциевый насос поддерживает содержание ионов Са++ в цитозоле на низком уровне. В качестве депо кальция
- 77. Са-АТФазы В саркоплазматической сети на долю Са-АТФазы приходится 60% общего мембранного белка. По-видимому, в мембране саркоплазматической
- 78. Ионные каналы Некоторые из них являются высокоизбирательными для определенных ионов, их относят к классу селективных (например,
- 79. потенциал-активируемые ионные каналы (переход из закрытого в открытое состояние и обратно осуществляется конформацией белковой молекулы при
- 80. Структура ионного канала
- 81. Потенциал - управляемые каналы – воротный механизм Сенсор напряжения Шар на веревочке
- 82. Холинорецептор – лиганд-управляемый канал
- 83. Токи в одном канале
- 84. ПП При потенциале покоя внутренняя сторона клеточной мембраны имеет заряд, знак которого (отрицательность) определяется наличием в
- 85. ПП Из-за этого в клетке наблюдается избыток отрицательных ионов, и, следовательно, отрицательных зарядов, а в интерстиции
- 86. Механизм электрогенеза Уравнение Гольдмана Здесь R – газовая постоянная, T –температура, z – заряд иона, а
- 87. РП Равновесный потенциал – такой потенциал плазмолеммы клетки, при котором суммарный ток конкретного иона через мембрану
- 88. Равновесный потенциал Ек = –75 мВ; ЕNa = +55 мВ; ЕCa = +150 мВ; ЕCl =
- 89. Закон Ома I=U/R iNa = gNa (Vm – ENa), где gNa = 1/R – проводимость, величина,
- 90. Направление ионного тока iК = gК(Vm–EК) При Vm = EК ток равен 0 При Vm если
- 91. 12.1 MANY COMMON FEATURES UNDERLIE THE DIVERSITY OF BIOLOGICAL MEMBRANES Membranes are as diverse in structure
- 95. Скачать презентацию

ВОПРОСЫ
Функции клеточных органелл
Компоненты мембраны возбудимой клетки как объект изучения физиологии
Белки
ВОПРОСЫ
Функции клеточных органелл
Компоненты мембраны возбудимой клетки как объект изучения физиологии
Белки

ОСНОВНЫЕ ПОНЯТИЯ
Клетка – это наименьшая функциональная единица тканей и органов животного
ОСНОВНЫЕ ПОНЯТИЯ
Клетка – это наименьшая функциональная единица тканей и органов животного
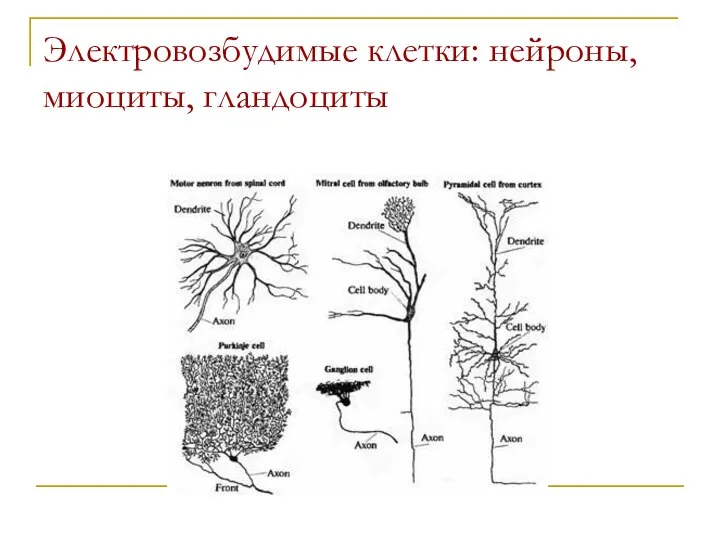
Электровозбудимые клетки: нейроны, миоциты, гландоциты
Электровозбудимые клетки: нейроны, миоциты, гландоциты

Системы
Под физиологической системой понимают анатомически детерминированную совокупность органов и тканей, выполняющую
Системы
Под физиологической системой понимают анатомически детерминированную совокупность органов и тканей, выполняющую

Интегративные системы
нервная,
эндокринная,
иммунная
Интегративные системы
нервная,
эндокринная,
иммунная

Функциональные системы
Функциональная система, согласно теории функциональных систем П.К.Анохина, — динамическая совокупность
Функциональные системы
Функциональная система, согласно теории функциональных систем П.К.Анохина, — динамическая совокупность

Функциональные системы
Функциональные системы поддерживают оптимальные физиологические показатели, обеспечивающие достижение полезного результата
Функциональные системы
Функциональные системы поддерживают оптимальные физиологические показатели, обеспечивающие достижение полезного результата

1.Функции клеточных органелл
1.Функции клеточных органелл

Структура везикулы
Структура везикулы

Функции клетки
1. Базовая основа жизни. Мельчайшая часть организма, способная сохранять свойство
Функции клетки
1. Базовая основа жизни. Мельчайшая часть организма, способная сохранять свойство
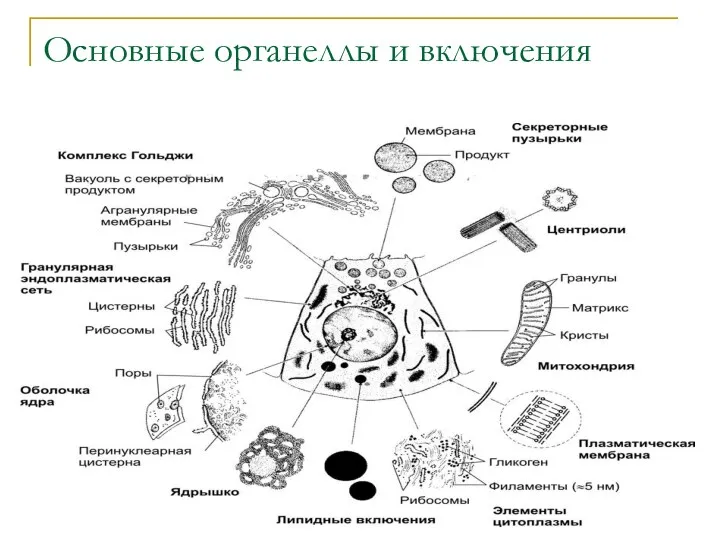
Основные органеллы и включения
Основные органеллы и включения

Структуры клетки человека
Структуры клетки человека

2. Компоненты мембраны возбудимой клетки как объект изучения физиологии.
2. Компоненты мембраны возбудимой клетки как объект изучения физиологии.

МНОГО ОБЩИХ ЧЕРТ ЛЕЖАТ В ОСНОВЕ
РАЗНООБРАЗИЯ БИОЛОГИЧЕСКИХ МЕМБРАН
Мембраны столь же разнообразны
МНОГО ОБЩИХ ЧЕРТ ЛЕЖАТ В ОСНОВЕ
РАЗНООБРАЗИЯ БИОЛОГИЧЕСКИХ МЕМБРАН
Мембраны столь же разнообразны

МНОГО ОБЩИХ ЧЕРТ ЛЕЖАТ В ОСНОВЕ
РАЗНООБРАЗИЯ БИОЛОГИЧЕСКИХ МЕМБРАН
4. Отличительные функции мембран
МНОГО ОБЩИХ ЧЕРТ ЛЕЖАТ В ОСНОВЕ
РАЗНООБРАЗИЯ БИОЛОГИЧЕСКИХ МЕМБРАН
4. Отличительные функции мембран

МНОГО ОБЩИХ ЧЕРТ ЛЕЖАТ В ОСНОВЕ
РАЗНООБРАЗИЯ БИОЛОГИЧЕСКИХ МЕМБРАН
7. Мембраны - жидкие
МНОГО ОБЩИХ ЧЕРТ ЛЕЖАТ В ОСНОВЕ
РАЗНООБРАЗИЯ БИОЛОГИЧЕСКИХ МЕМБРАН
7. Мембраны - жидкие

Жидкостно-мозаичная модель
Жидкостно-мозаичная модель

Клетки всех живых организмов окружены внешней мембраной – плазмолеммой, цитолеммой. Большинство
Клетки всех живых организмов окружены внешней мембраной – плазмолеммой, цитолеммой. Большинство

Липиды
Структурную матрицу мембраны составляют липиды, на долю которых приходится от 15
Липиды
Структурную матрицу мембраны составляют липиды, на долю которых приходится от 15

Белки мембраны условно разделяют на периферические и собственные (интегральные).
Периферические белки расположены
Белки мембраны условно разделяют на периферические и собственные (интегральные).
Периферические белки расположены

Рецептор связанный с G-белком
Рецептор связанный с G-белком

Мембранно связанный энзим
Мембранно связанный энзим

Внутриклеточный рецептор
Внутриклеточный рецептор

Углеводы
Углеводы клеточных мембран присутствуют в плазмолемме в виде соединений с белками
Углеводы
Углеводы клеточных мембран присутствуют в плазмолемме в виде соединений с белками

Транспорт веществ: аквапорин
Транспорт веществ: аквапорин

Транспорт веществ: диффузия
Транспорт веществ: диффузия

Осмос и осмотические нарушения
Осмос и осмотические нарушения

Фильтрация и реабсорбция
Фильтрация и реабсорбция

Na+К+ помпа: активный транспорт
Na+К+ помпа: активный транспорт

Вторично-активный транспорт: симпорт и антипорт, GLUT1-12
Вторично-активный транспорт: симпорт и антипорт, GLUT1-12


Относительное значение механизмов транспорта веществ
Относительное значение механизмов транспорта веществ

3. Белки – насосы и белки – ионные каналы.
Насосы
3. Белки – насосы и белки – ионные каналы.
Насосы

Классы ионных насосов
Классы ионных насосов

Разные формы цитоза: эндоцитоз
Разные формы цитоза: эндоцитоз

пиноцитоз
пиноцитоз

Рецептор-опосредованный эндоцитоз ЛПНП
Рецептор-опосредованный эндоцитоз ЛПНП

экзоцитоз
экзоцитоз

Обобщение видов транспорта через мембрану
Обобщение видов транспорта через мембрану

Обобщение видов транспорта через мембрану
Обобщение видов транспорта через мембрану

4.Белки-рецепторы химических и физических сигналов
4.Белки-рецепторы химических и физических сигналов

Классификация рецепторов
Мембранные рецепторы регистрируют наличие лиганда (пептидного гормона, производных аминокислот, нейромедиаторов,
Классификация рецепторов
Мембранные рецепторы регистрируют наличие лиганда (пептидного гормона, производных аминокислот, нейромедиаторов,

Классификация рецепторов
1) рецепторы, трансмембранный сегмент которых состоит из семи фрагментов (петель);
2)
Классификация рецепторов
1) рецепторы, трансмембранный сегмент которых состоит из семи фрагментов (петель);
2)

К гормонам, рецептор которых состоит из семи трансмембранных фрагментов, относятся:
АКТГ, ТТГ,
К гормонам, рецептор которых состоит из семи трансмембранных фрагментов, относятся:
АКТГ, ТТГ,

Ко второй группе относятся гормоны, имеющие один трансмембранный фрагмент:
СТГ, пролактин, инсулин,
Ко второй группе относятся гормоны, имеющие один трансмембранный фрагмент:
СТГ, пролактин, инсулин,

К гормонам третьей группы, рецептор которых имеет четыре трансмембранных фрагмента, относятся
ацетилхолин
К гормонам третьей группы, рецептор которых имеет четыре трансмембранных фрагмента, относятся
ацетилхолин

Количество имеет значение
Количество клеточных рецепторов, как и других компонентов клетки, постоянно
Количество имеет значение
Количество клеточных рецепторов, как и других компонентов клетки, постоянно

В представлениях современной молекулярной физиологии клетки, основные процессы в воспринимающих сигнал
В представлениях современной молекулярной физиологии клетки, основные процессы в воспринимающих сигнал

Среди подвергаемых фосфорилированию и дефосфорилированию ферментов имеется набор протеинкиназ, осуществляющих регуляцию
Среди подвергаемых фосфорилированию и дефосфорилированию ферментов имеется набор протеинкиназ, осуществляющих регуляцию

Поэтому различают сериновые/треониновые протеинкиназы
и тирозиновые протеинкиназы.
Обратный процесс – дефосфолрилирование обеспечивается
Поэтому различают сериновые/треониновые протеинкиназы
и тирозиновые протеинкиназы.
Обратный процесс – дефосфолрилирование обеспечивается

Другой путь, который используется при передаче сигналов между клетками, связан с
Другой путь, который используется при передаче сигналов между клетками, связан с

Мембранные рецепторы подразделяют на:
каталитические,
связанные с ионными каналами,
являющиеся ионными каналами,
Мембранные рецепторы подразделяют на:
каталитические,
связанные с ионными каналами,
являющиеся ионными каналами,
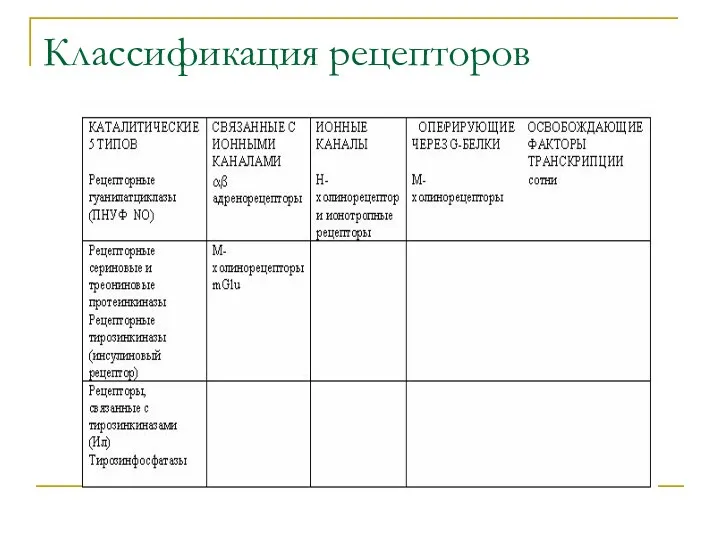
Классификация рецепторов
Классификация рецепторов

Каталитические
1. Рецепторные гуанилатциклазы катализируют образование цГМФ из ГТФ.
К таким мембранным
Каталитические
1. Рецепторные гуанилатциклазы катализируют образование цГМФ из ГТФ.
К таким мембранным

Каталитические
2.Рецепторные серин/треониновые киназы
фосфорилируют остатки серина и/или треонина как самого рецептора
Каталитические
2.Рецепторные серин/треониновые киназы
фосфорилируют остатки серина и/или треонина как самого рецептора

Каталитические
3.Рецепторные тирозинкиназы фосфорилируют остатки тирозина как самого белка-рецептора (автофосфорилирование), так и
Каталитические
3.Рецепторные тирозинкиназы фосфорилируют остатки тирозина как самого белка-рецептора (автофосфорилирование), так и

Рецептор к инсулину - тирозинкиназа
Рецептор к инсулину - тирозинкиназа

Каталитические
4.Рецепторы, связанные с тирозинкиназами, взаимодействуют с цитоплазматическими тирозинкиназами, многие из которых
Каталитические
4.Рецепторы, связанные с тирозинкиназами, взаимодействуют с цитоплазматическими тирозинкиназами, многие из которых

Каталитические
5.Рецепторные тирозинфосфатазы освобождают (дефосфорилируют) фосфатные группировки из фосфорилированных тирозилов внутриклеточных белков.
Каталитические
5.Рецепторные тирозинфосфатазы освобождают (дефосфорилируют) фосфатные группировки из фосфорилированных тирозилов внутриклеточных белков.

Рецепторы, связанные с ионными каналами, влияют на проницаемость ионных каналов через
Рецепторы, связанные с ионными каналами, влияют на проницаемость ионных каналов через

Управляемые лигандами ионные каналы — интегральные мембранные белки — сочетают свойства рецептора и
Управляемые лигандами ионные каналы — интегральные мембранные белки — сочетают свойства рецептора и

Классификация синаптических рецепторов: ионотропные и метаботропные
Классификация синаптических рецепторов: ионотропные и метаботропные

Рецепторы, связанные с G‑белком — полипептидные цепочки, расположенные в плазматической мембране клетки–мишени
Рецепторы, связанные с G‑белком — полипептидные цепочки, расположенные в плазматической мембране клетки–мишени


Лиганды связанных с G‑белком мембранных рецепторов
ангиотензин II, АТФ, ацетилхолин (мускариновые рецепторы),
Лиганды связанных с G‑белком мембранных рецепторов
ангиотензин II, АТФ, ацетилхолин (мускариновые рецепторы),
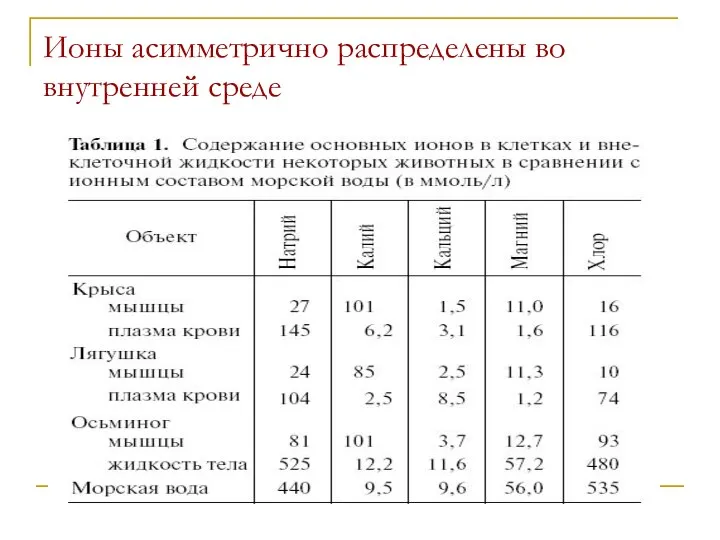
Ионы асимметрично распределены во внутренней среде
Ионы асимметрично распределены во внутренней среде

ИОННЫЕ ГРАДИЕНТЫ НУЖНЫ ДЛЯ:
ЗАПАСАНИЯ ЭНЕРГИИ
ГЕНЕРАЦИИ ВОЗБУЖДЕНИЯ
ТРАНСПОРТА ГЛЮКОЗЫ И АМИНОКИСЛОТ
ИОННЫЕ ГРАДИЕНТЫ НУЖНЫ ДЛЯ:
ЗАПАСАНИЯ ЭНЕРГИИ
ГЕНЕРАЦИИ ВОЗБУЖДЕНИЯ
ТРАНСПОРТА ГЛЮКОЗЫ И АМИНОКИСЛОТ

Для создания потенциалов нужны ионы Na+, K+, Ca++, Cl-
Na+ и K+
Для создания потенциалов нужны ионы Na+, K+, Ca++, Cl-
Na+ и K+

физико-химические отличия ионов
1.Для создания электрохимического потенциала ионов K+ нужно на порядок
физико-химические отличия ионов
1.Для создания электрохимического потенциала ионов K+ нужно на порядок

физико-химические отличия ионов
Из-за высокой гидрофобности K+ проникает через мембрану в 5
физико-химические отличия ионов
Из-за высокой гидрофобности K+ проникает через мембрану в 5
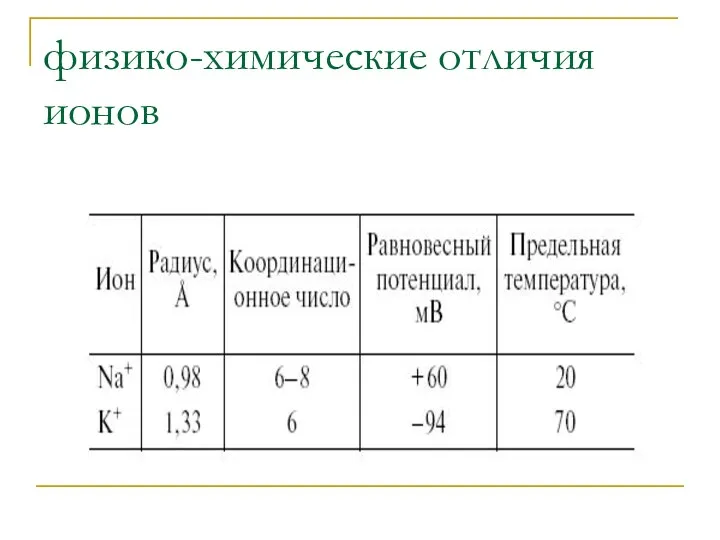
физико-химические отличия ионов
физико-химические отличия ионов
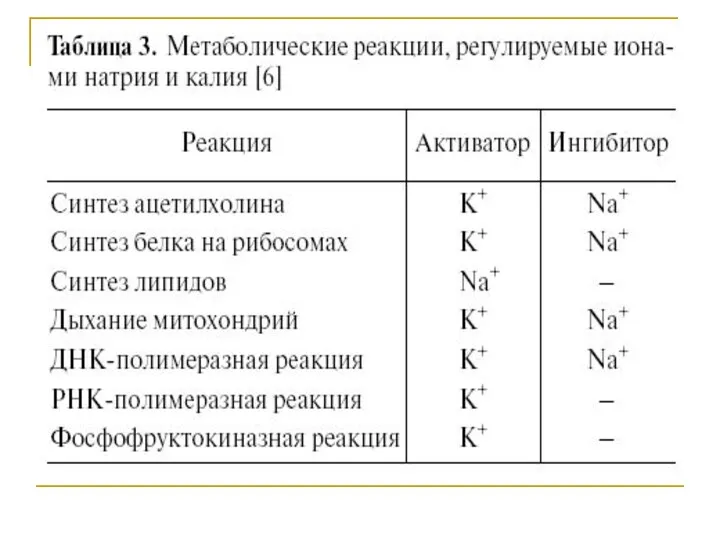

Стехиометрия
Для встречного перемещения (антипорта) натрия и калия Na-K-АТФазой характерна стехиометрия, для
Стехиометрия
Для встречного перемещения (антипорта) натрия и калия Na-K-АТФазой характерна стехиометрия, для

30% метаболизма
Калий-натриевая помпа локализована в мембранах практически всех клеток организма, но
30% метаболизма
Калий-натриевая помпа локализована в мембранах практически всех клеток организма, но

Са-АТФазы
Кальциевый насос поддерживает содержание ионов Са++ в цитозоле на низком уровне.
Са-АТФазы
Кальциевый насос поддерживает содержание ионов Са++ в цитозоле на низком уровне.

Са-АТФазы
В саркоплазматической сети на долю Са-АТФазы приходится 60% общего мембранного белка.
Са-АТФазы
В саркоплазматической сети на долю Са-АТФазы приходится 60% общего мембранного белка.

Ионные каналы
Некоторые из них являются высокоизбирательными для определенных ионов, их относят
Ионные каналы
Некоторые из них являются высокоизбирательными для определенных ионов, их относят

потенциал-активируемые ионные каналы (переход из закрытого в открытое состояние и обратно
потенциал-активируемые ионные каналы (переход из закрытого в открытое состояние и обратно

Структура ионного канала
Структура ионного канала

Потенциал - управляемые каналы – воротный механизм
Сенсор напряжения
Шар на веревочке
Потенциал - управляемые каналы – воротный механизм
Сенсор напряжения
Шар на веревочке

Холинорецептор –
лиганд-управляемый канал
Холинорецептор –
лиганд-управляемый канал

Токи в одном канале
Токи в одном канале

ПП
При потенциале покоя внутренняя сторона клеточной мембраны имеет заряд, знак которого
ПП
При потенциале покоя внутренняя сторона клеточной мембраны имеет заряд, знак которого

ПП
Из-за этого в клетке наблюдается избыток отрицательных ионов, и, следовательно, отрицательных
ПП
Из-за этого в клетке наблюдается избыток отрицательных ионов, и, следовательно, отрицательных

Механизм электрогенеза
Уравнение Гольдмана
Здесь R – газовая постоянная, T –температура, z –
Механизм электрогенеза
Уравнение Гольдмана
Здесь R – газовая постоянная, T –температура, z –

РП
Равновесный потенциал – такой потенциал плазмолеммы клетки, при котором суммарный ток
РП
Равновесный потенциал – такой потенциал плазмолеммы клетки, при котором суммарный ток

Равновесный потенциал
Ек = –75 мВ;
ЕNa = +55 мВ;
ЕCa = +150 мВ;
ЕCl
Равновесный потенциал
Ек = –75 мВ;
ЕNa = +55 мВ;
ЕCa = +150 мВ;
ЕCl

Закон Ома I=U/R
iNa = gNa (Vm – ENa),
где gNa = 1/R
Закон Ома I=U/R
iNa = gNa (Vm – ENa),
где gNa = 1/R

Направление ионного тока
iК = gК(Vm–EК)
При Vm = EК ток равен 0
При
Направление ионного тока
iК = gК(Vm–EК)
При Vm = EК ток равен 0
При

12.1 MANY COMMON FEATURES UNDERLIE THE
DIVERSITY OF BIOLOGICAL MEMBRANES
Membranes are as
12.1 MANY COMMON FEATURES UNDERLIE THE
DIVERSITY OF BIOLOGICAL MEMBRANES
Membranes are as


 Сходства и различия клеток растений, животных и грибов
Сходства и различия клеток растений, животных и грибов Использование дрожжевых грибов для производства разных групп продуктов
Использование дрожжевых грибов для производства разных групп продуктов Открытое мероприятие по профилактике вредных привычек
Открытое мероприятие по профилактике вредных привычек Бактерії - прокаріотичні одноклітинні мікроорганізми
Бактерії - прокаріотичні одноклітинні мікроорганізми Влияние ГМО на организм человека
Влияние ГМО на организм человека Общая физиология сенсорных систем
Общая физиология сенсорных систем Половое поведение животных
Половое поведение животных Дыхание растений
Дыхание растений Круговорот кислорода в природе
Круговорот кислорода в природе Звёздный час. Загадки о животных и растениях
Звёздный час. Загадки о животных и растениях Популяция. Популяционная генетика
Популяция. Популяционная генетика Сахароза. Дисахариды (олигосахариды)
Сахароза. Дисахариды (олигосахариды) Клетки и их разнообразие в многоклеточном организме
Клетки и их разнообразие в многоклеточном организме Основы гистологии. Виды тканей
Основы гистологии. Виды тканей An introduction to metabolism
An introduction to metabolism Глюкоза. Молекулярна формула. Фізичні та хімічні властивості глюкози. Поширення в природі
Глюкоза. Молекулярна формула. Фізичні та хімічні властивості глюкози. Поширення в природі 20231101_vestniki_vesny_viktorina
20231101_vestniki_vesny_viktorina Глаз
Глаз Cattle breeds
Cattle breeds Как зимой помочь птицам (урок окружающего мира, 1 класс)
Как зимой помочь птицам (урок окружающего мира, 1 класс) Эффективный способ посадки картофеля
Эффективный способ посадки картофеля Мы говорим Нет алкоголю!
Мы говорим Нет алкоголю! Физиология растений. Механизмы поступления воды в растительную клетку. Дальний транспорт воды в растении
Физиология растений. Механизмы поступления воды в растительную клетку. Дальний транспорт воды в растении Презентация по биологии для 9 класса по теме:Метаболизм. Энергетический обмен.
Презентация по биологии для 9 класса по теме:Метаболизм. Энергетический обмен. Животные Антарктиды
Животные Антарктиды Красная книга Удмуртии. Животные
Красная книга Удмуртии. Животные Кератин в волосах и восстановление волос
Кератин в волосах и восстановление волос Катархей. Архей
Катархей. Архей