- Ферменты строение и свойства. Регуляция активности ферментов. (Лекция 1.3)

Содержание
- 2. Строение и свойства ферментов. По своей химической природе ферменты (энзимы) являются простыми или сложными белками. В
- 3. Особенности структурной организации ферментов. Ферментам присущи все особенности структурной организации белков. Они имеют четыре уровня организации
- 4. Кроме активного центра у ферментов (Е) имеется регуляторный центр, через который обеспечивается регуляция активного центра посредством
- 5. Четвертичная структура фермента Большинство ферментов, имея четыре уровня структурной организации, состоят из нескольких субъединиц (протомеров). Вследствие
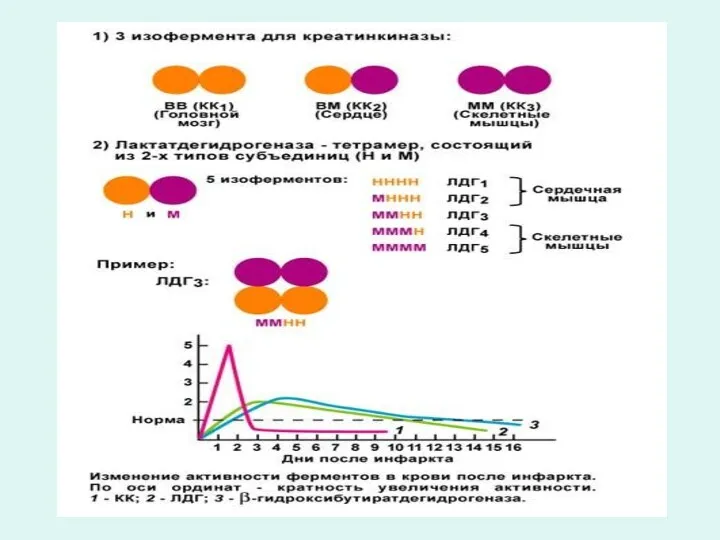
- 6. Каждый орган имеет характерный для него набор ферментов, которые при развитии в этом органе патологического процесса
- 8. Кофакторы ферментов Кофакторы у ферментов бывают двух типов: - ионы металлов - коферменты Металлы, как кофакторы
- 9. Типы коферментов В отличие от ионов металлов, коферменты являются сложными органическими соединениями и по своей химической
- 10. Коферменты алифатического ряда К этой группе коферментов относятся глютатион и липоевая кислота. Глутатион - это кофермент
- 11. Коэнзим Q (убихинон)
- 12. КОЭНЗИМ Q10 (УБИХИНОН) от лат. ubiquitous - повсеместный). Коэнзим Q10 (убихинон) по строению и окислительно-восстановительным свойствам
- 13. Биологические эффекты убихинона ( коэнзима Q10) Коэнзим Q10 участвует в трансмембранном переносе электронов от водорода окисляемого
- 14. Коэнзим Q10 усиливает фагоцитарную активность макрофагов, повышает число гранулоцитов в костном мозге и плазме крови, увеличивает
- 15. . Коэнзим Q10 полезен для лечения последствий радиоактивного поражения. Коэнзим Q10 уменьшать токсичность лекарств и поэтому
- 16. Коферменты гетероциклического ряда К коферментам гетроциклического ряда относятся производные водорастворимых витаминов: витамина В1 - тиаминпирофосфат (ТПФ),
- 17. Тиаминпирофосфат
- 18. Тиаминпирофосфат Тиаминпирофосфат (ТПФ) является коферментом окислительных декарбоксилаз альфа-кетокислот (пирувата, альфа-кетоглутарата) и транскетолаз. ТПФ непосредственно или косвенно
- 19. Фосфопиридоксаль
- 20. Фосфопиридоксаль Фосфопиридоксаль (ФП-СН=О) является коферментом трансаминаз, декарбоксилаз и изомераз аминокислот. Входит в состав моноаминоксидаз и диаминоксидаз
- 21. Карбоксибиотин
- 22. Карбоксибиотин Коферментной формой биотина (витамина Н) является N-карбоксибиотин, который входит в состав ферментов обеспечивающих реакции карбоксилирования:
- 23. тетрагидрофолиевая кислота (ТГФК)
- 24. тетрагидрофолиевая кислота (ТГФК) 5,6,7,8- тетрагидрофолиевая кислота (ТГФК) является коферментом ферментов обеспечивающих перенос одноуглеродистых радикалов необходимых для
- 25. Коферменты нуклеотиды Коферменты нуклеотиды можно рассматривать как производные водорастворимых витаминов: витамина РР (В5) - никотинамидадениндинуклеотид (НАД)
- 26. НАД
- 27. Никотинамид адениндинуклеотид (НАД) и никотинамид адениндинуклеотид фосфат (НАДФ) являются производными витамина РР (В5, никотинамид) и служат
- 28. НАДФ
- 29. ФЛАВИН-АДЕНИН-ДИНУКЛЕОТИД (ФАД) ФЛАВИН-МОНОНУКЛЕОТИД (ФМН) Коферменты ФАД и ФМН являются производными витамина В2 (рибофлавин), являются коферментами аэробных
- 30. ФАД
- 31. ФМН
- 32. Коэнзим А Коэнзим А является производным витамина В3 – пантотеновой кислоты CH3 | O=C---CH--C--CH2-OH | |
- 33. Коэнзим А
- 34. Не содержащие витамины нуклеотидные коферменты. Коферменты этого типа можно поделить на пять групп в зависимости от
- 35. Металлокоферменты К этой группе коферментов относятся железопорфириновые и колбамидные коферменты. Железопорфириновые гемсодержащие коферменты. Коферменты цитохромов, цитохромоксидазы,
- 36. Гем
- 37. Классификация ферментов Согласно международной классификации ферменты делятся на шесть классов по типу катализируемой реакции: 1. Оксидоредуктазы
- 38. 1 класс - Оксидоредуктазы. Ферменты этого класса катализируют окислительно-восстановительные реакции, лежащие в основе процессов биологического окисления,
- 39. 2 класс - Трансферазы. Ферменты этого класса катализируют перенос функциональных групп и делятся по этому принципу
- 40. 3 класс - Гидролазы. Этот класс ферментов катализирует реакции гидролиза, и в зависимости от типа гидролизируемой
- 41. 4 класс - Лиазы. Ферменты этого класса катализируют не гидролитическое разрушение соединений с отщеплением групп (СО2,
- 42. 5 класс - Изомеразы. Эта группа ферментов катализирует реакции изомеризации, и делятся на 6 подклассов: 5.1.
- 43. 6 класс - Лигазы (синтетазы). Этот класс ферментов катализирует реакции синтеза соединений с использованием энергии АТФ
- 44. В каждом классе для структурной организации молекулы фермента используются свои группы коферментов (коэнзимов): - 1 класс
- 45. Особенности ферментативного катализа. Ферментативный катализ отличается целым рядов отличительных признаков: 1.Ферменты намного активнее чем неорганические катализаторы.
- 46. Специфичность бывает абсолютная и относительная. Абсолютная специфичность - фермент катализирует превращение только одного субстрата. Например, уреаза
- 47. Некоторые ферменты проявляют абсолютную групповую специфичность, катализируя превращение сходной группы субстратов. Например, алкогольдегидрогеназа катализирует превращение не
- 48. Скорость ферментативного катализа зависит от температуры Влияние температуры: Влияние температуры на скорость ферментативной реакции может быть
- 49. Влияние рН-среды: При постоянной температуре любой фермент проявляет наибольшую активность в определённом диапазоне рН среды. Оптимальным
- 50. Влияние активаторов и ингибиторов Вещества, повышающие активность ферментов и, следовательно, скорость ферментативной реакции называются активаторами. Ионы
- 52. Различают обратимое и необратимое ингибирование ферментов. Обратимое ингибирование бывает двух видов: конкурентное и неконкурентное обратимое ингибирование.
- 53. Конкурентное обратимое ингибирование. Примером конкурентного обратимого ингибирования может служить ингибирование сукцинатдегидрогеназы малоновой или щавелевоуксусной кислотой, соединениями
- 54. Неконкурентное обратимое ингибирование Некоторые ингибиторы образуют комплекс не со свободным ферментом, а с фермент субстратным комплексом:
- 55. Необратимое ингибирование Необратимое ингибирование вызывается малыми концентрациями ионов тяжелых металлов, например ионов ртути, серебра и мышьяка
- 56. Влияние концентрация субстрата
- 57. Влияние концентрации фермента
- 58. Регуляция активности ферментов. Метаболические процессы, протекающие в клетке находятся в постоянном равновесии, одновременны, подчинены и определяют
- 59. Единицы измерения активности ферментов За международную единицу активности фермента принимается такое его количество, которое катализирует превращение
- 60. Номенклатура ферментов. В 1961 году специальной комиссией Международного биохимического союза была предложена систематическая номенклатура ферментов. Каждый
- 61. Современные подходы к диагностике заболеваний При любых заболеваниях всегда имеет место нарушения различных звеньев обмена веществ,
- 62. 1.Ферменты углеводного обмена Дихотомическимй путь катаболизма глюкозы: гексокиназа (фосфоглюкокиназа) фосфорилаза фосфоглюкоизомераза фосфофруктокиназа альдолаза фосфотриозоизомераза глицеральдегид-дегидрогеназа фосфоглицераткиназа
- 63. Ферменты общих путей катаболизма окислительная декарбоксилаза пирувата цитратсинтетаза аконитаза изоцитратдегидрогеназа α-кетоглутарат дегидрогеназа сукцинаттиокиназа сукцинатдегирогеназа фумараза малатдегидрогеназа
- 64. Метаболизм гликогена Синтез гликогене и глюконеогенез: УДФ-глюкозилпирофосфорилаза гликогенсинтетаза (гликозилтрансфераза) амило-гликозилтрансфераза пируваткарбоксилаза Фосфоенолпируваткарбоксикиназа Мобилизация гликогена : аденилатциклаза
- 65. Превращения олигосахаридов и гликозамингликанов мальтаза лактаза сахараза амилаза глюкуронидаза альдозоредуктаза сорбитдегирогеназа УДФ-глюкозилтрансфераза УДФ-глюкуронилтрансфераза
- 66. Ферменты дыхательной цепи митохондрий Биоэнергетика клетки окислительная декарбоксилаза пирувата изоцитратдегидрогеназа α-кетоглутарат дегидрогеназа сукцинаттиокиназа сукцинатдегирогеназа малатдегидрогеназа НАДН-дегидрогеназа
- 67. Катаболизм триациглицеридов и жирных кислот триацилглицеридлипаза липопротеидлипаза диглицеридлипаза моноглицеридлипаза липопротеинлипаза глицераткиназа α-глицерофосфатдегидрогеназа ацил КоА-синтетаза трансацетилаза карнитин-ацилтрансфераза
- 68. Анаболизм липидов и биосинтез кетоновых тел и холестерина Анаболизм липидов ацетил КоА-карбоксилаза малонитрансацетилаза цитратлиаза β-кетоацил-синтетаза β-кетоацил-редуктаза
- 69. Катаболизм белков и аминокислот пептидазы пищеварительных соков (пепсин, трипсин, химотрипсин) тканевые катепсины трансаминазы глутаматдегидрогеназа АМФ-аминогидролаза гистидаза
- 70. Обезвреживание аммиака и синтез мочевины глутаминсинтетаза глутаминаза карбомоилфосфат-синтетаза карбомоилтрансфераз аргининосукцинасинтетаза аргининосукциназа аргиназа
- 71. Ферменты обмен нуклеотидов и нуклеиновых кислот нуклеазы (ДНК-аза, РНК-аза) 5`-нуклеотидаза нуклеозидазы АМФ дезаминаза аденозиндезаминаза гуаниндезаминаза цитозиндезаминаза
- 73. Скачать презентацию

Строение и свойства ферментов.
По своей химической природе ферменты (энзимы) являются
Строение и свойства ферментов.
По своей химической природе ферменты (энзимы) являются

Особенности структурной организации ферментов.
Ферментам присущи все особенности структурной организации белков. Они
Особенности структурной организации ферментов.
Ферментам присущи все особенности структурной организации белков. Они

Кроме активного центра у ферментов (Е) имеется регуляторный центр, через который
Кроме активного центра у ферментов (Е) имеется регуляторный центр, через который

Четвертичная структура фермента
Большинство ферментов, имея четыре уровня структурной организации, состоят из
Четвертичная структура фермента
Большинство ферментов, имея четыре уровня структурной организации, состоят из

Каждый орган имеет характерный для него набор ферментов, которые при развитии
Каждый орган имеет характерный для него набор ферментов, которые при развитии

Кофакторы ферментов
Кофакторы у ферментов бывают двух типов:
- ионы металлов
-
Кофакторы ферментов
Кофакторы у ферментов бывают двух типов:
- ионы металлов
-

Типы коферментов
В отличие от ионов металлов, коферменты являются сложными органическими соединениями
Типы коферментов
В отличие от ионов металлов, коферменты являются сложными органическими соединениями

Коферменты алифатического ряда
К этой группе коферментов относятся глютатион и липоевая кислота.
Коферменты алифатического ряда
К этой группе коферментов относятся глютатион и липоевая кислота.
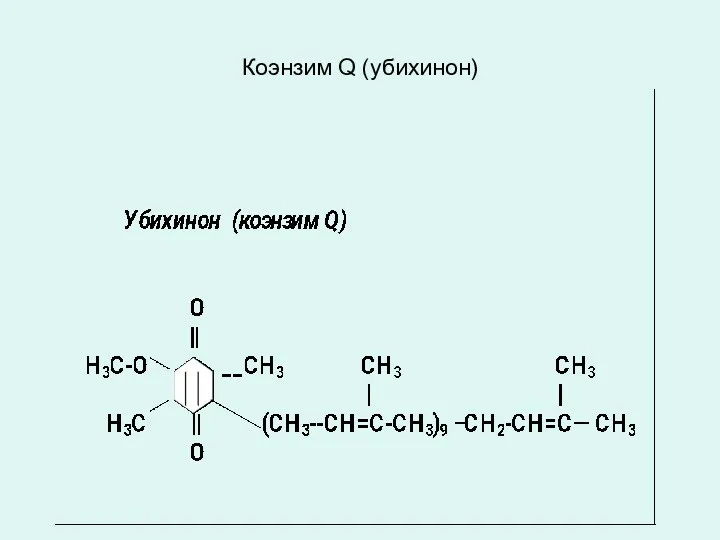
Коэнзим Q (убихинон)
Коэнзим Q (убихинон)

КОЭНЗИМ Q10 (УБИХИНОН) от лат. ubiquitous - повсеместный).
Коэнзим Q10
КОЭНЗИМ Q10 (УБИХИНОН) от лат. ubiquitous - повсеместный).
Коэнзим Q10

Биологические эффекты убихинона ( коэнзима Q10)
Коэнзим Q10 участвует в трансмембранном
Биологические эффекты убихинона ( коэнзима Q10)
Коэнзим Q10 участвует в трансмембранном

Коэнзим Q10 усиливает фагоцитарную активность макрофагов, повышает число гранулоцитов в костном
Коэнзим Q10 усиливает фагоцитарную активность макрофагов, повышает число гранулоцитов в костном

.
Коэнзим Q10 полезен для лечения последствий радиоактивного поражения.
Коэнзим Q10 уменьшать
.
Коэнзим Q10 полезен для лечения последствий радиоактивного поражения.
Коэнзим Q10 уменьшать

Коферменты гетероциклического ряда
К коферментам гетроциклического ряда относятся производные водорастворимых витаминов:
Коферменты гетероциклического ряда
К коферментам гетроциклического ряда относятся производные водорастворимых витаминов:
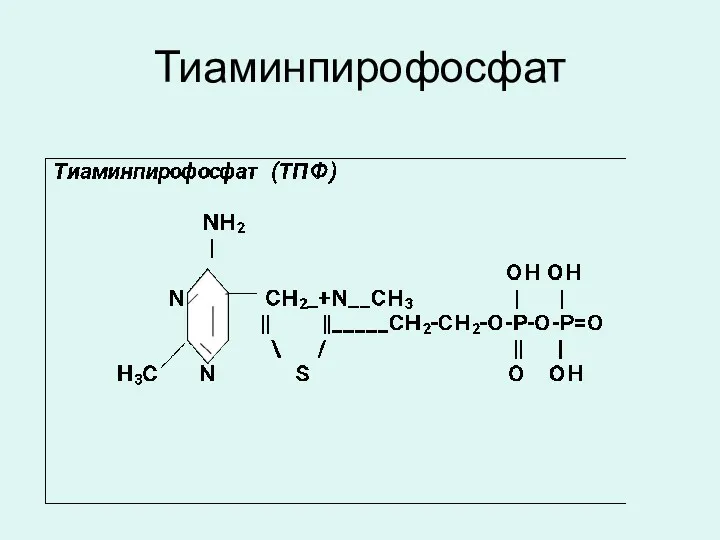
Тиаминпирофосфат
Тиаминпирофосфат

Тиаминпирофосфат
Тиаминпирофосфат (ТПФ) является коферментом окислительных декарбоксилаз альфа-кетокислот (пирувата, альфа-кетоглутарата) и транскетолаз.
Тиаминпирофосфат
Тиаминпирофосфат (ТПФ) является коферментом окислительных декарбоксилаз альфа-кетокислот (пирувата, альфа-кетоглутарата) и транскетолаз.
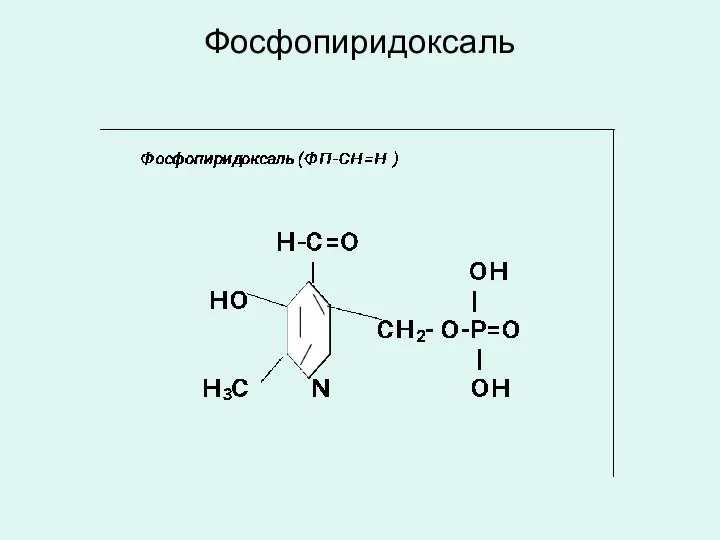
Фосфопиридоксаль
Фосфопиридоксаль

Фосфопиридоксаль
Фосфопиридоксаль (ФП-СН=О) является коферментом трансаминаз, декарбоксилаз и изомераз аминокислот.
Входит
Фосфопиридоксаль
Фосфопиридоксаль (ФП-СН=О) является коферментом трансаминаз, декарбоксилаз и изомераз аминокислот.
Входит
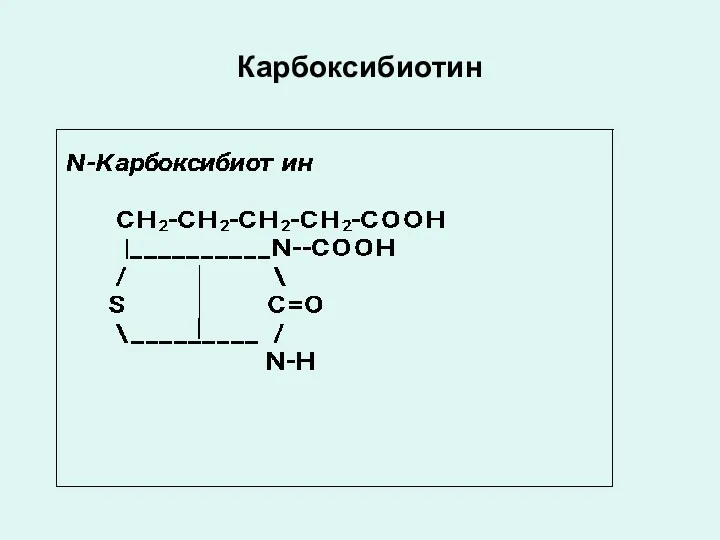
Карбоксибиотин
Карбоксибиотин

Карбоксибиотин
Коферментной формой биотина (витамина Н) является N-карбоксибиотин, который входит в состав
Карбоксибиотин
Коферментной формой биотина (витамина Н) является N-карбоксибиотин, который входит в состав
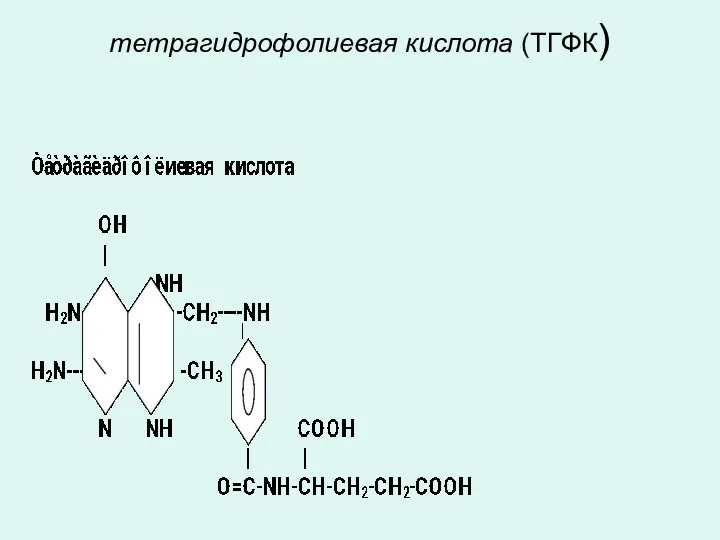
тетрагидрофолиевая кислота (ТГФК)
тетрагидрофолиевая кислота (ТГФК)

тетрагидрофолиевая кислота (ТГФК)
5,6,7,8- тетрагидрофолиевая кислота (ТГФК) является коферментом ферментов обеспечивающих перенос
тетрагидрофолиевая кислота (ТГФК)
5,6,7,8- тетрагидрофолиевая кислота (ТГФК) является коферментом ферментов обеспечивающих перенос

Коферменты нуклеотиды
Коферменты нуклеотиды можно рассматривать как производные водорастворимых витаминов:
витамина РР
Коферменты нуклеотиды
Коферменты нуклеотиды можно рассматривать как производные водорастворимых витаминов:
витамина РР
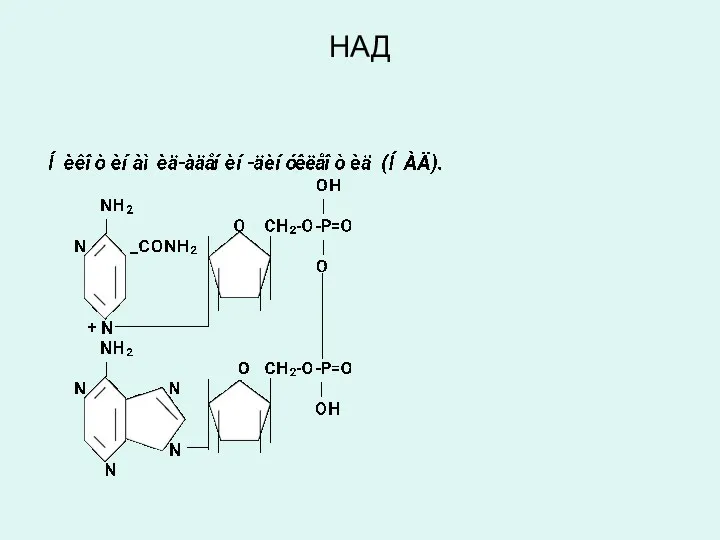
НАД
НАД

Никотинамид адениндинуклеотид (НАД) и никотинамид адениндинуклеотид фосфат (НАДФ)
являются производными витамина
Никотинамид адениндинуклеотид (НАД) и никотинамид адениндинуклеотид фосфат (НАДФ)
являются производными витамина
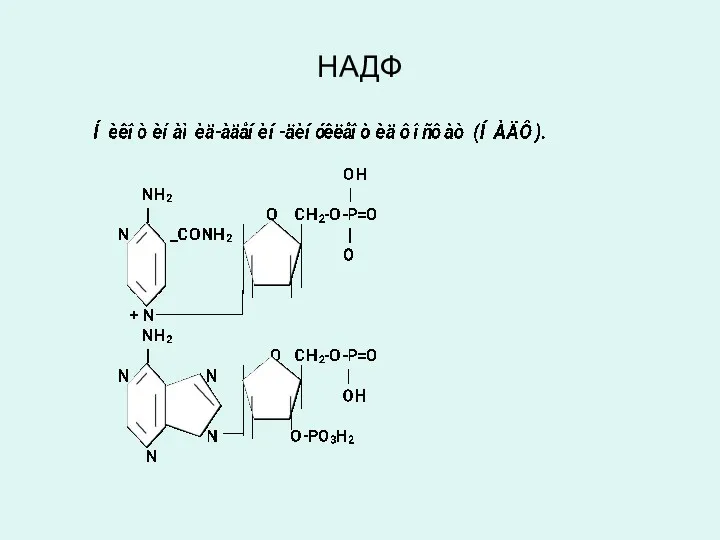
НАДФ
НАДФ

ФЛАВИН-АДЕНИН-ДИНУКЛЕОТИД (ФАД)
ФЛАВИН-МОНОНУКЛЕОТИД (ФМН)
Коферменты ФАД и ФМН являются производными витамина В2
ФЛАВИН-АДЕНИН-ДИНУКЛЕОТИД (ФАД)
ФЛАВИН-МОНОНУКЛЕОТИД (ФМН)
Коферменты ФАД и ФМН являются производными витамина В2
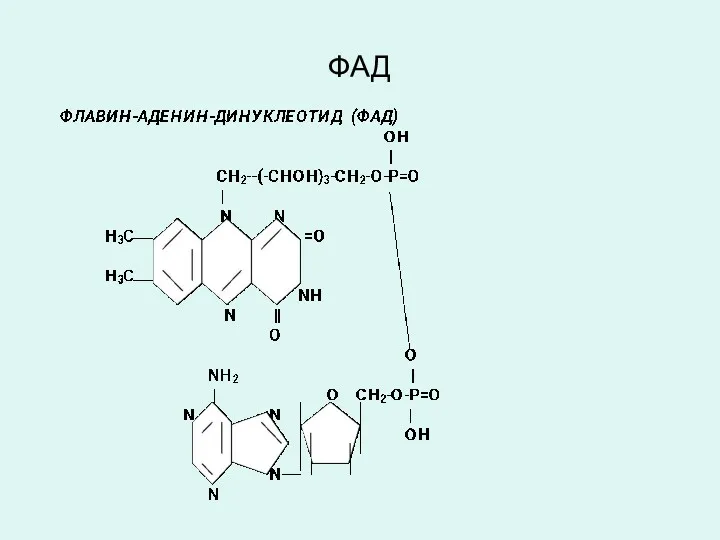
ФАД
ФАД
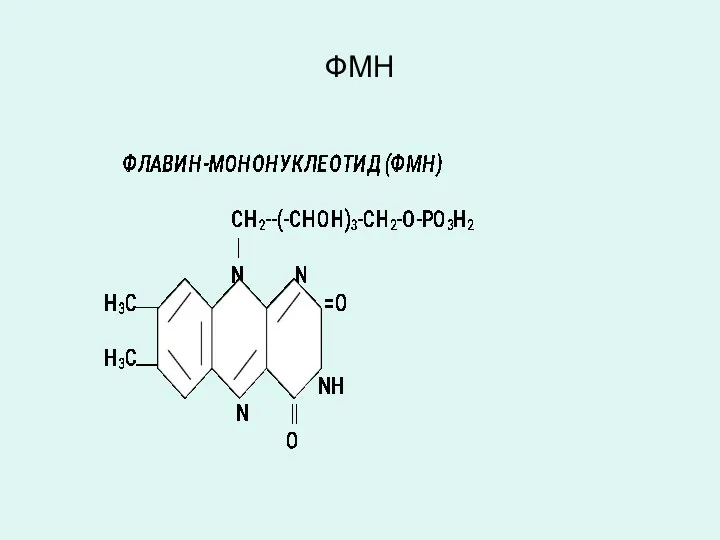
ФМН
ФМН
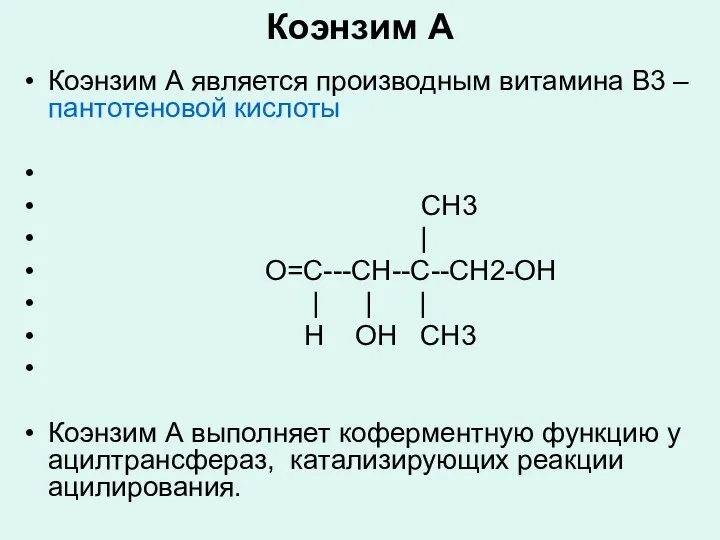
Коэнзим А
Коэнзим А является производным витамина В3 – пантотеновой кислоты
CH3
Коэнзим А
Коэнзим А является производным витамина В3 – пантотеновой кислоты
CH3
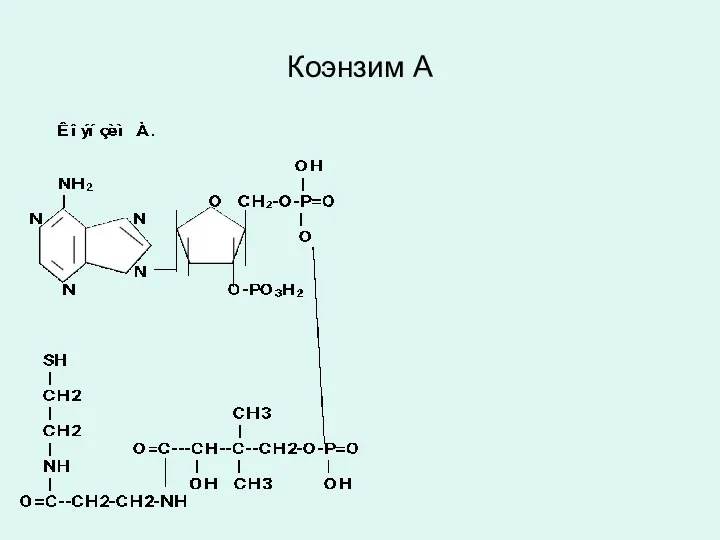
Коэнзим А
Коэнзим А

Не содержащие витамины нуклеотидные коферменты.
Коферменты этого типа можно поделить на
Не содержащие витамины нуклеотидные коферменты.
Коферменты этого типа можно поделить на

Металлокоферменты
К этой группе коферментов относятся железопорфириновые и колбамидные коферменты.
Железопорфириновые гемсодержащие
Металлокоферменты
К этой группе коферментов относятся железопорфириновые и колбамидные коферменты.
Железопорфириновые гемсодержащие
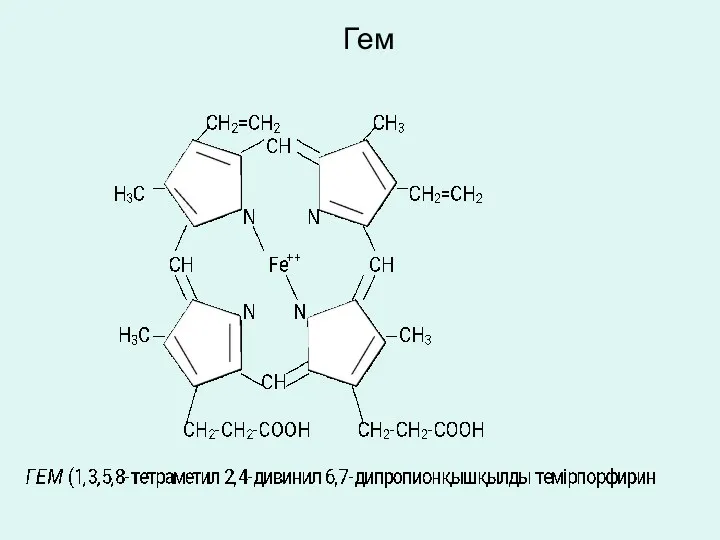
Гем
Гем

Классификация ферментов
Согласно международной классификации ферменты делятся на шесть классов по
Классификация ферментов
Согласно международной классификации ферменты делятся на шесть классов по

1 класс - Оксидоредуктазы.
Ферменты этого класса катализируют окислительно-восстановительные реакции, лежащие в
1 класс - Оксидоредуктазы.
Ферменты этого класса катализируют окислительно-восстановительные реакции, лежащие в

2 класс - Трансферазы.
Ферменты этого класса катализируют перенос функциональных групп и
2 класс - Трансферазы.
Ферменты этого класса катализируют перенос функциональных групп и

3 класс - Гидролазы.
Этот класс ферментов катализирует реакции гидролиза, и в
3 класс - Гидролазы.
Этот класс ферментов катализирует реакции гидролиза, и в

4 класс - Лиазы.
Ферменты этого класса катализируют не гидролитическое разрушение
4 класс - Лиазы.
Ферменты этого класса катализируют не гидролитическое разрушение

5 класс - Изомеразы.
Эта группа ферментов катализирует реакции изомеризации, и делятся
5 класс - Изомеразы.
Эта группа ферментов катализирует реакции изомеризации, и делятся

6 класс - Лигазы (синтетазы).
Этот класс ферментов катализирует реакции синтеза соединений
6 класс - Лигазы (синтетазы).
Этот класс ферментов катализирует реакции синтеза соединений

В каждом классе для структурной организации молекулы фермента используются свои группы
В каждом классе для структурной организации молекулы фермента используются свои группы

Особенности ферментативного катализа.
Ферментативный катализ отличается целым рядов отличительных признаков:
1.Ферменты намного
Особенности ферментативного катализа.
Ферментативный катализ отличается целым рядов отличительных признаков:
1.Ферменты намного

Специфичность бывает абсолютная и относительная.
Абсолютная специфичность - фермент катализирует превращение только
Специфичность бывает абсолютная и относительная.
Абсолютная специфичность - фермент катализирует превращение только

Некоторые ферменты проявляют абсолютную групповую специфичность, катализируя превращение сходной группы субстратов.
Некоторые ферменты проявляют абсолютную групповую специфичность, катализируя превращение сходной группы субстратов.

Скорость ферментативного катализа зависит от температуры
Влияние температуры:
Влияние температуры на скорость
Скорость ферментативного катализа зависит от температуры
Влияние температуры:
Влияние температуры на скорость

Влияние рН-среды:
При постоянной температуре любой фермент проявляет наибольшую активность в определённом
Влияние рН-среды:
При постоянной температуре любой фермент проявляет наибольшую активность в определённом

Влияние активаторов и ингибиторов
Вещества, повышающие активность ферментов и, следовательно, скорость ферментативной
Влияние активаторов и ингибиторов
Вещества, повышающие активность ферментов и, следовательно, скорость ферментативной
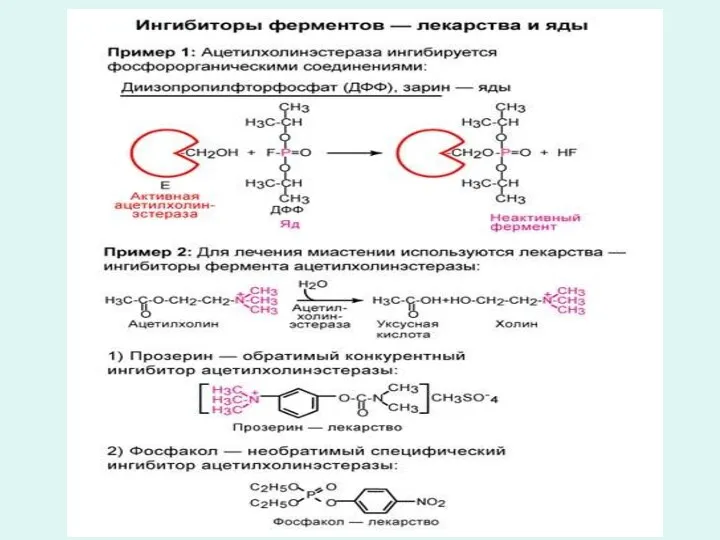

Различают обратимое и необратимое ингибирование ферментов.
Обратимое ингибирование бывает двух видов: конкурентное
Различают обратимое и необратимое ингибирование ферментов.
Обратимое ингибирование бывает двух видов: конкурентное

Конкурентное обратимое ингибирование.
Примером конкурентного обратимого ингибирования может служить ингибирование сукцинатдегидрогеназы малоновой
Конкурентное обратимое ингибирование.
Примером конкурентного обратимого ингибирования может служить ингибирование сукцинатдегидрогеназы малоновой

Неконкурентное обратимое ингибирование
Некоторые ингибиторы образуют комплекс не со свободным ферментом, а
Неконкурентное обратимое ингибирование
Некоторые ингибиторы образуют комплекс не со свободным ферментом, а

Необратимое ингибирование
Необратимое ингибирование вызывается малыми концентрациями ионов тяжелых металлов, например ионов
Необратимое ингибирование
Необратимое ингибирование вызывается малыми концентрациями ионов тяжелых металлов, например ионов
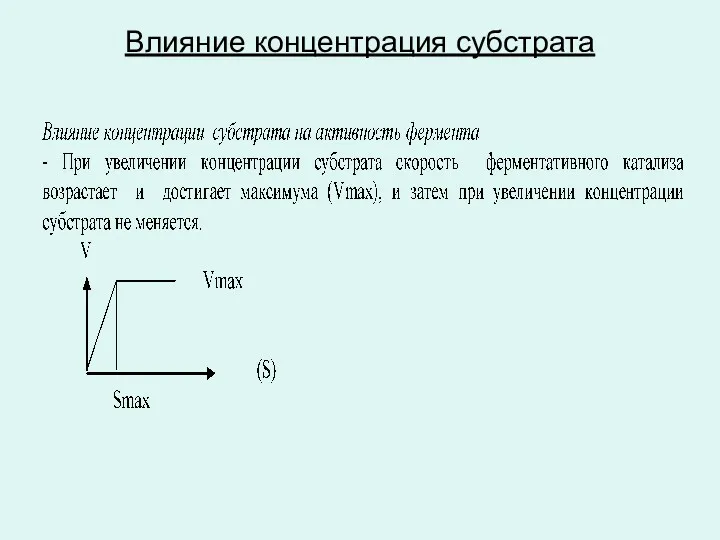
Влияние концентрация субстрата
Влияние концентрация субстрата
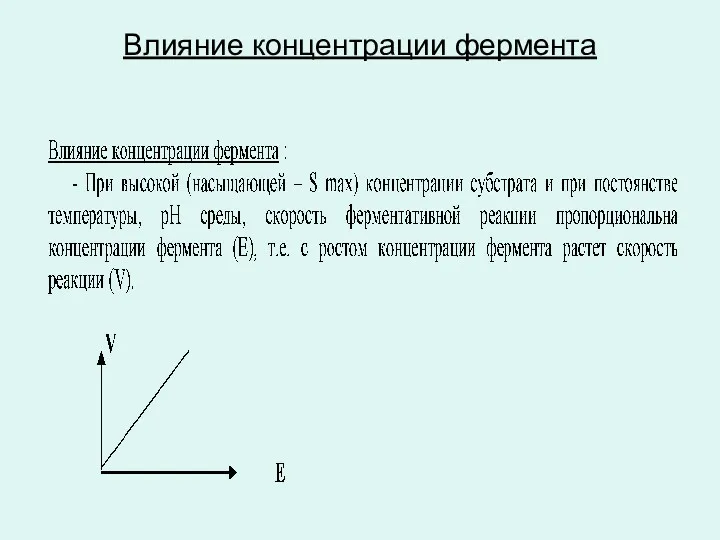
Влияние концентрации фермента
Влияние концентрации фермента

Регуляция активности ферментов.
Метаболические процессы, протекающие в клетке находятся в постоянном
Регуляция активности ферментов.
Метаболические процессы, протекающие в клетке находятся в постоянном

Единицы измерения активности ферментов
За международную единицу активности фермента принимается такое его
Единицы измерения активности ферментов
За международную единицу активности фермента принимается такое его

Номенклатура ферментов.
В 1961 году специальной комиссией Международного биохимического союза была
Номенклатура ферментов.
В 1961 году специальной комиссией Международного биохимического союза была

Современные подходы к диагностике заболеваний
При любых заболеваниях всегда имеет место нарушения
Современные подходы к диагностике заболеваний
При любых заболеваниях всегда имеет место нарушения

1.Ферменты углеводного обмена
Дихотомическимй путь катаболизма глюкозы:
гексокиназа (фосфоглюкокиназа)
фосфорилаза
фосфоглюкоизомераза
фосфофруктокиназа
альдолаза
фосфотриозоизомераза
глицеральдегид-дегидрогеназа
фосфоглицераткиназа
фосфоглицеромутаза
енолаза
пируваткиназа
лактатдегидрогеназа
Апотомический путь катаболизма глюкозы:
глюкоза-6-фосфатдегидрогеназа
лактоназа
6-фосфоглюконат дегидрогеназа
пентозофосфатизомераза
транскетолаза
трансальдолаза
1.Ферменты углеводного обмена
Дихотомическимй путь катаболизма глюкозы:
гексокиназа (фосфоглюкокиназа)
фосфорилаза
фосфоглюкоизомераза
фосфофруктокиназа
альдолаза
фосфотриозоизомераза
глицеральдегид-дегидрогеназа
фосфоглицераткиназа
фосфоглицеромутаза
енолаза
пируваткиназа
лактатдегидрогеназа
Апотомический путь катаболизма глюкозы:
глюкоза-6-фосфатдегидрогеназа
лактоназа
6-фосфоглюконат дегидрогеназа
пентозофосфатизомераза
транскетолаза
трансальдолаза

Ферменты общих путей катаболизма
окислительная декарбоксилаза пирувата
цитратсинтетаза
аконитаза
изоцитратдегидрогеназа
α-кетоглутарат дегидрогеназа
сукцинаттиокиназа
сукцинатдегирогеназа
фумараза
малатдегидрогеназа
Ферменты общих путей катаболизма
окислительная декарбоксилаза пирувата
цитратсинтетаза
аконитаза
изоцитратдегидрогеназа
α-кетоглутарат дегидрогеназа
сукцинаттиокиназа
сукцинатдегирогеназа
фумараза
малатдегидрогеназа

Метаболизм гликогена
Синтез гликогене и глюконеогенез:
УДФ-глюкозилпирофосфорилаза
гликогенсинтетаза (гликозилтрансфераза)
амило-гликозилтрансфераза
пируваткарбоксилаза
Фосфоенолпируваткарбоксикиназа
Мобилизация гликогена :
аденилатциклаза
фосфорилазкиназа
фосфорилаза
протеинкиназа
Метаболизм гликогена
Синтез гликогене и глюконеогенез:
УДФ-глюкозилпирофосфорилаза
гликогенсинтетаза (гликозилтрансфераза)
амило-гликозилтрансфераза
пируваткарбоксилаза
Фосфоенолпируваткарбоксикиназа
Мобилизация гликогена :
аденилатциклаза
фосфорилазкиназа
фосфорилаза
протеинкиназа

Превращения олигосахаридов и гликозамингликанов
мальтаза
лактаза
сахараза
амилаза
глюкуронидаза
альдозоредуктаза
сорбитдегирогеназа
УДФ-глюкозилтрансфераза
УДФ-глюкуронилтрансфераза
Превращения олигосахаридов и гликозамингликанов
мальтаза
лактаза
сахараза
амилаза
глюкуронидаза
альдозоредуктаза
сорбитдегирогеназа
УДФ-глюкозилтрансфераза
УДФ-глюкуронилтрансфераза

Ферменты дыхательной цепи митохондрий Биоэнергетика клетки
окислительная декарбоксилаза пирувата
изоцитратдегидрогеназа
α-кетоглутарат дегидрогеназа
сукцинаттиокиназа
сукцинатдегирогеназа
малатдегидрогеназа
НАДН-дегидрогеназа
ФМНН-дегидрогеназа
КоQ -редуктаза
цитохром-С-оксидаза
Ферменты дыхательной цепи митохондрий Биоэнергетика клетки
окислительная декарбоксилаза пирувата
изоцитратдегидрогеназа
α-кетоглутарат дегидрогеназа
сукцинаттиокиназа
сукцинатдегирогеназа
малатдегидрогеназа
НАДН-дегидрогеназа
ФМНН-дегидрогеназа
КоQ -редуктаза
цитохром-С-оксидаза

Катаболизм триациглицеридов и жирных кислот
триацилглицеридлипаза
липопротеидлипаза
диглицеридлипаза
моноглицеридлипаза
липопротеинлипаза
глицераткиназа
α-глицерофосфатдегидрогеназа
ацил КоА-синтетаза
трансацетилаза
карнитин-ацилтрансфераза
ацил-КоА-дегидрогеназа
еноил-КоА-гидратаза
гидроксиацил-КоА-дегидрогеназа
ацил-КоА-ацилтрансфераза
циклооксигеназа
липооксигеназа
Катаболизм триациглицеридов и жирных кислот
триацилглицеридлипаза
липопротеидлипаза
диглицеридлипаза
моноглицеридлипаза
липопротеинлипаза
глицераткиназа
α-глицерофосфатдегидрогеназа
ацил КоА-синтетаза
трансацетилаза
карнитин-ацилтрансфераза
ацил-КоА-дегидрогеназа
еноил-КоА-гидратаза
гидроксиацил-КоА-дегидрогеназа
ацил-КоА-ацилтрансфераза
циклооксигеназа
липооксигеназа

Анаболизм липидов и биосинтез кетоновых тел и холестерина
Анаболизм липидов
ацетил КоА-карбоксилаза
малонитрансацетилаза
цитратлиаза
β-кетоацил-синтетаза
β-кетоацил-редуктаза
гидроксиацил-гидратаза
еноилредуктаза
ацилтрансфераза
фосфатаза
Биосинтез кетоновых
Анаболизм липидов и биосинтез кетоновых тел и холестерина
Анаболизм липидов
ацетил КоА-карбоксилаза
малонитрансацетилаза
цитратлиаза
β-кетоацил-синтетаза
β-кетоацил-редуктаза
гидроксиацил-гидратаза
еноилредуктаза
ацилтрансфераза
фосфатаза
Биосинтез кетоновых

Катаболизм белков и аминокислот
пептидазы пищеварительных соков (пепсин, трипсин, химотрипсин)
тканевые катепсины
трансаминазы
глутаматдегидрогеназа
АМФ-аминогидролаза
гистидаза
серин (треонин)
Катаболизм белков и аминокислот
пептидазы пищеварительных соков (пепсин, трипсин, химотрипсин)
тканевые катепсины
трансаминазы
глутаматдегидрогеназа
АМФ-аминогидролаза
гистидаза
серин (треонин)

Обезвреживание аммиака и синтез мочевины
глутаминсинтетаза
глутаминаза
карбомоилфосфат-синтетаза
карбомоилтрансфераз
аргининосукцинасинтетаза
аргининосукциназа
аргиназа
Обезвреживание аммиака и синтез мочевины
глутаминсинтетаза
глутаминаза
карбомоилфосфат-синтетаза
карбомоилтрансфераз
аргининосукцинасинтетаза
аргининосукциназа
аргиназа

Ферменты обмен нуклеотидов и нуклеиновых кислот
нуклеазы (ДНК-аза, РНК-аза)
5`-нуклеотидаза
нуклеозидазы
АМФ дезаминаза
аденозиндезаминаза
гуаниндезаминаза
цитозиндезаминаза
ксантиноксидаза
фосфорибозилтрансфераза
дигидрооротатдегидрогеназа
рибонуклеозидредуктаза
тиоредоксинредуктаза
Ферменты обмен нуклеотидов и нуклеиновых кислот
нуклеазы (ДНК-аза, РНК-аза)
5`-нуклеотидаза
нуклеозидазы
АМФ дезаминаза
аденозиндезаминаза
гуаниндезаминаза
цитозиндезаминаза
ксантиноксидаза
фосфорибозилтрансфераза
дигидрооротатдегидрогеназа
рибонуклеозидредуктаза
тиоредоксинредуктаза
 Применение нейронных сетей в биотехнических системах
Применение нейронных сетей в биотехнических системах Теория антропосоциогенеза
Теория антропосоциогенеза Презентации к урокам
Презентации к урокам Залози тіла людини
Залози тіла людини Fats and oils
Fats and oils Клинико-физиологические особенности женского организма в репродуктивный период
Клинико-физиологические особенности женского организма в репродуктивный период Природа человека. Теории происхождения человека
Природа человека. Теории происхождения человека Насекомые. Отличительные признаки насекомых (1 класса)
Насекомые. Отличительные признаки насекомых (1 класса) Nervous system
Nervous system Центральная нервна система
Центральная нервна система Селекция животных
Селекция животных Отдел Покрытосеменные
Отдел Покрытосеменные Общие пути катаболизма
Общие пути катаболизма Класс млекопитающие
Класс млекопитающие Удивительный мир веществ. Урок-игра
Удивительный мир веществ. Урок-игра Класс Однодольные
Класс Однодольные Борьба за существование и ее формы. 11 класс
Борьба за существование и ее формы. 11 класс Качкодзьоб. Спеціальні пристосування качкодзьоба
Качкодзьоб. Спеціальні пристосування качкодзьоба Ультраструктура бактериальной клетки
Ультраструктура бактериальной клетки Общие закономерности ВНД у человека и животных
Общие закономерности ВНД у человека и животных Парнокопытные и непарнокопытные животные
Парнокопытные и непарнокопытные животные Водоросли. Среда обитания
Водоросли. Среда обитания Анатомия глаза
Анатомия глаза Кровеносная система
Кровеносная система Отдел Покрытосеменные: Ч.1
Отдел Покрытосеменные: Ч.1 Стволовые клетки
Стволовые клетки Уровни организации живой материи
Уровни организации живой материи Тварини в екосистемі
Тварини в екосистемі