- Филогенез клеток и функции клеточных мембран

Содержание
- 2. Филогенез эукариотической клетки
- 3. Клетка Прокариот Эукариот Царство бактерии Царство археи Bacilla Cocci Только одноклеточные и бактериальные сообщества (пленки и
- 4. Last Eukaryote Common Ancestor Последний эукариотический общий предок
- 5. В 1905 K.С. Мережковский предположил, что эукариотическая клетка образовалась в результате симбиогенеза – слияния бактериальных клеток
- 6. Теория симбиогенеза anaerobes aerobes 1905 «Uber Natur und Ursprung der Chromatophoren im Pflanzen reiche», K.С. Мережковский
- 7. V. A. Kozlov, full Bilogical Science doctor, Ph.D. Medical Science, professor Р. Гупта предположил, что эукариотическая
- 8. Последний эукариотический общий предок имел значительно более сложный геном, чем у современных многоклеточных. Многоклеточность осуществлялась через
- 9. Типы соединения клеток
- 10. нексус Клетка – Клетка контакты простого типа а) адгезионные б) интердигитация (пальцевые соединения) контакты сцепляющего типа
- 13. Межклеточные контакты необходимы для соединения клеток информационного обмена а) межклеточного б) с внешней средой Для этого
- 14. Ионные каналы
- 16. Общее представление о структуре и функциях ионных каналов Каналообразующие белковые молекулы всех ионных каналов имеют некоторые
- 17. Трехмерная пространственная структура каналообразующей молекулы белка располагается в цитоплазматической мембране устья канала, обращенные к наружной и
- 18. Описано > 100 разновидностей ионных каналов, для их классификации используют различные подходы. Один из них основан
- 19. Пассивные каналы могут быть открытыми (активными) в покоящихся клетках, т.е. в отсутствие каких-либо воздействий не являются
- 20. Каналы щелевых (плотных) контактов клеток Каналы щелевых контактов формируются в области контакта двух соседних клеток, очень
- 21. Мембраны двух соседних клеток Канал Щелевой контакт 3,5 нм Коннексин – белок Коннексон – функциональный комплекс
- 22. Ионные каналы открываются и закрываются бывают в ответ на механозависимые потенциалзависимые лигандзависимые взаимодействие низкомолекулярного медиатора с
- 23. Потенциалзависимые каналы Состояние этих каналов управляется силами электрического поля, создаваемого величиной разности потенциалов (Δϕ) на мембране.
- 24. Потенциал-зависимый канал может находиться в трех последовательных состояниях: закрытый; открытый; инактивированный Состояние канала зависит от величины
- 25. Скорость открытия активационных ворот может быть очень высокой и низкой. По этому показателю потенциалзависимые ионные каналы
- 26. Активационные ворота – последовательность аминокислот в виде плотного шарика (клубка) на нити, располагающегося у выхода устья
- 27. Существуют вещества, блокирующие работу потенциалзависимых ионных каналов. Таким образом, возможно еще одно состояние ионного канала –
- 28. Все блокаторы ионных каналов действуют одинаково – встраиваются как пробка в просвете канала. Различия между блокаторами
- 29. Порообразующая часть ионного канала может представлять собой 1) единый полипептид, организованный в виде нескольких идентичных трансмембранных
- 30. Строение Na+-ионного канала клеточной мембраны: а – двухмерное строение α-субъединицы ионного канала клеточной мембраны; б –
- 31. Нарушение работы ионных каналов часто приводят к заболеваниям — каналопатиям. Основная причина таких нарушений — наследственные
- 32. Работа ионного канала (разным цветом выделены разные белки, образующие домены) пропускная способность открытого канала поразительна: ток
- 33. Лигандзависимые ионные каналы Формируются белковыми макромолекулами, одновременно выполняющими функцию ионных каналов и селективных рецепторов лигандов. Поэтому
- 34. Лиганд-зависимый канал Центр связывания лиганда (трансмиттера) Клеточная мембрана 2α, β, γ, δ белковые субъединицы, образующие систему
- 35. 14 нанометров 12 нанометров Антагонист рецептора Трансмембранный домен Лигандсвязывающий домен Аминоконцевой домен Молекулярная (белковая) структура лигандзависимого
- 36. Лигандзависимые ионные каналы обычно локализованы в постсинаптических мембранах нервных клеток и их отростков, а также мышечных
- 37. Лигандзависимые каналы могут изменять проницаемость для катионов Na+ и К+ или для К+ и анионов СI–.
- 38. Существуют медленные лигандзависимые ионные каналы. Их открытие реализуется через цепь последовательных событий: первичный медиатор (лиганд, нейромедиатор)
- 39. Ацетилхолин К+-канал Гиперполяризация Синоатриальный узел миокарда Связь ионного канала с рецептором Ацетилхолин возбуждает m2-рецептор, это через
- 40. Особым типом лигандзависимых каналов являются каналы, локализованные в мембранах эндоплазматического ретикулума гладкомышечной клетки. Их лигандом является
- 41. Рецепторы РЕЦЕПЦИЯ – процесс восприятия и трансформации (преобразования) энергии внешнего раздражителя в нервный импульс или сложную
- 42. Сенсорные Мембранные Внутриклеточные Специализированные клетки: 1) механорецепторы, реагируют на механическое сжатие или рас-тяжение рецептора или смежных
- 43. Пропускает ион в ответ на механическую деформацию клеточной мембраны – механозависимые каналы тактильных, гравитационных, болевых нейронных
- 44. Сенсорные рецепторы
- 45. механорецепторы кожи, мышц, сердечно-сосудистой системы, внутренних органов механорецепторы органов слуха и вестибулярного аппарата Механорецепторы Вторичночувствующие: восприятие
- 46. Кортиев орган Орган равновесия
- 47. Периферические неинкапсулированные нервные окончания: кожа, подкожные ткани, кожные и подкожные сосуды медиальная преоптическая область гипоталамуса (центральные
- 48. Ноцицепторы Свободные нервные окончания безмиелиновые (медленные нервные волокна) и миелиновые (быстрые нервные волокна) Реакция на превышение
- 49. Электромагнитные рецепторы Под действием фотона родопсин разрушается на люмиродопсин и метародопсин, который гидролизуется с водой с
- 50. 3D организация сетчатки
- 51. Центральные (медуллярные) хеморецепторы расположены непосредственно в ростральных отделах вентральной дыхательной группы, в структурах голубого пятна (locus
- 52. Осморецепторы Расположены в гипоталамусе и печени. Реагируют на изменение осмотического давления плазмы крови. Между осморецепторами печени
- 53. Молекулярные рецепторы
- 55. Структура рецепторов а) ионотропного и б) связанного с G-белком
- 56. Мембранные рецепторы сопряженные с ионными каналами Механизм работы: Активация рецептора Открытие ионного канала Изменение электрического потенциала
- 57. NMDA-рецептор представляет из себя рецепторно-ионофорный комплекс, включающий в себя: 1) сайт специфического связывания медиатора (L-глутаминовой кислоты);
- 58. Мембранные рецепторы сопряженные с G-белками Механизм работы: Активация рецептора Активация G–белка Активация фермента, катализирующего образование второго
- 59. Лиганд связывается с участком узнавания политопного рецептора, это меняет его конформацию и запускает активность гуанозинфосфатазы, образующей
- 60. Молекулярная организация рецепторов, связанных с G-белком ФОСФОИНОЗИТИДНАЯ СИСТЕМА
- 61. G-белки двух типов: GS – стимулирующие клеточный ответ GI – ингибирующие клеточный ответ Лиганды (первичные посредники)
- 63. Молекулярная организация монотопных рецепторов с собственной ферментативной активностью Лиганды: факторы роста нервов, тромбоцитов, инсулин и др.,
- 64. Внутриклеточные (ядерные) рецепторы – факторы транскрипции Только гидрофобные молекулы!
- 66. Скачать презентацию

Филогенез эукариотической клетки
Филогенез эукариотической клетки
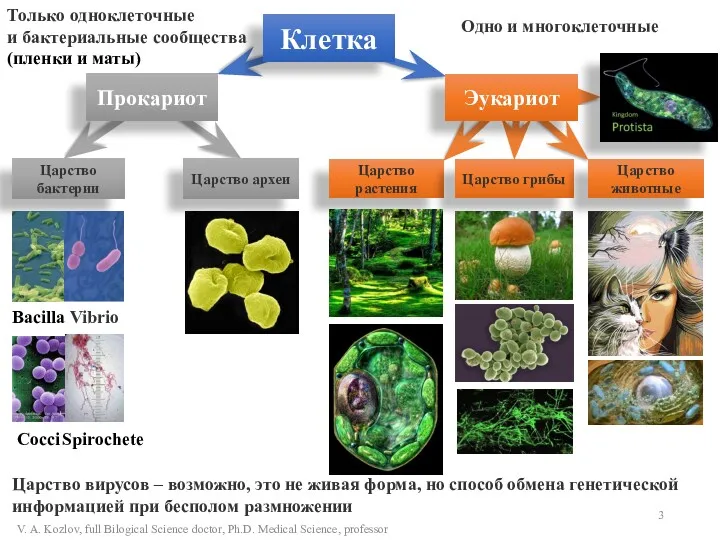
Клетка
Прокариот
Эукариот
Царство бактерии
Царство археи
Bacilla
Cocci
Только одноклеточные
и бактериальные сообщества
(пленки и маты)
Одно и многоклеточные
Царство
Клетка
Прокариот
Эукариот
Царство бактерии
Царство археи
Bacilla
Cocci
Только одноклеточные
и бактериальные сообщества
(пленки и маты)
Одно и многоклеточные
Царство
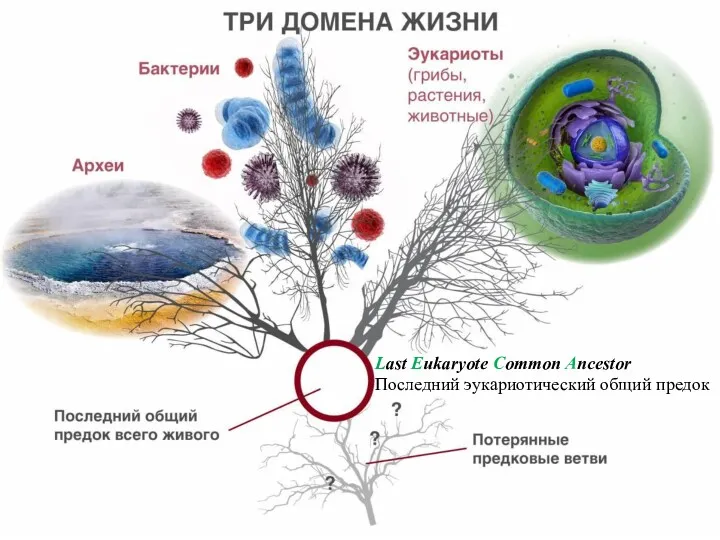
Last Eukaryote Common Ancestor
Последний эукариотический общий предок
Last Eukaryote Common Ancestor
Последний эукариотический общий предок

В 1905 K.С. Мережковский предположил, что эукариотическая клетка образовалась в результате
В 1905 K.С. Мережковский предположил, что эукариотическая клетка образовалась в результате
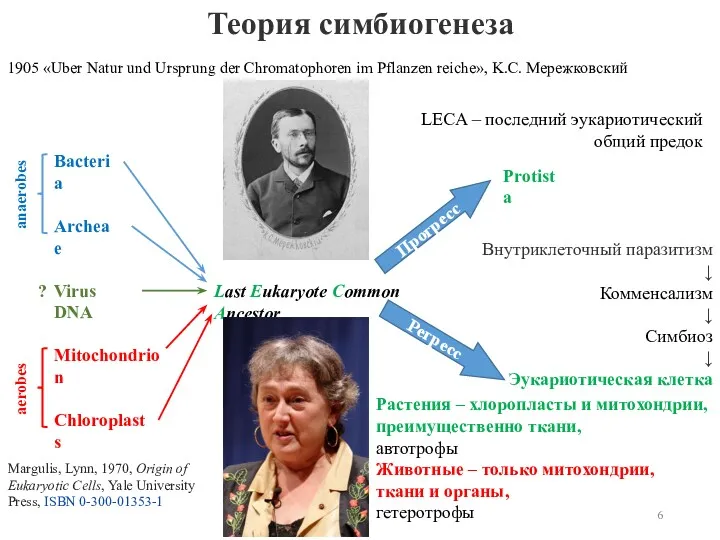
Теория симбиогенеза
anaerobes
aerobes
1905 «Uber Natur und Ursprung der Chromatophoren im Pflanzen reiche»,
Теория симбиогенеза
anaerobes
aerobes
1905 «Uber Natur und Ursprung der Chromatophoren im Pflanzen reiche»,

V. A. Kozlov, full Bilogical Science doctor, Ph.D. Medical Science, professor
Р.
V. A. Kozlov, full Bilogical Science doctor, Ph.D. Medical Science, professor
Р.

Последний эукариотический общий предок имел значительно более сложный геном, чем у
Последний эукариотический общий предок имел значительно более сложный геном, чем у

Типы соединения клеток
Типы соединения клеток
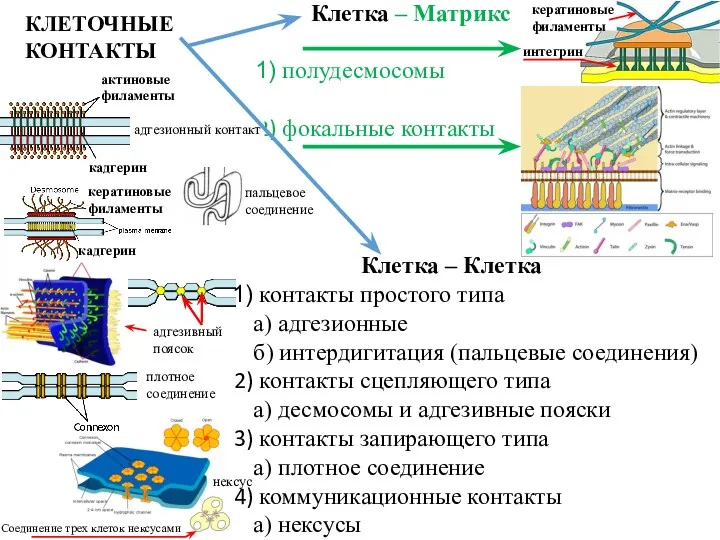
нексус
Клетка – Клетка
контакты простого типа
а) адгезионные
б) интердигитация (пальцевые соединения)
нексус
Клетка – Клетка
контакты простого типа
а) адгезионные
б) интердигитация (пальцевые соединения)
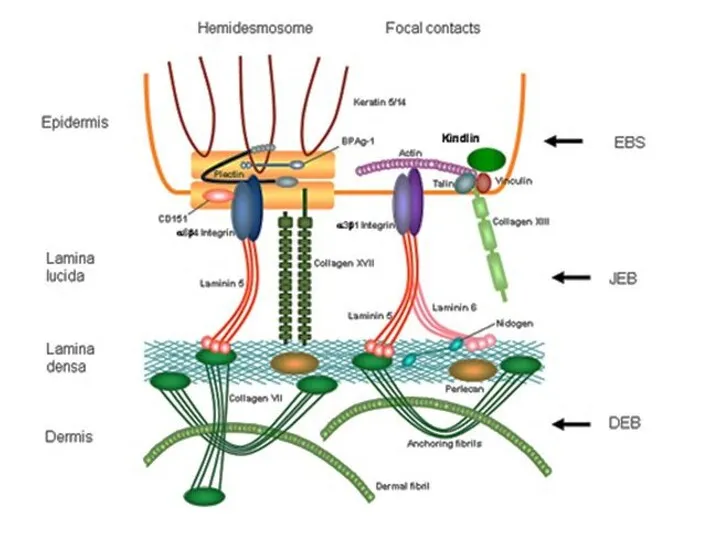
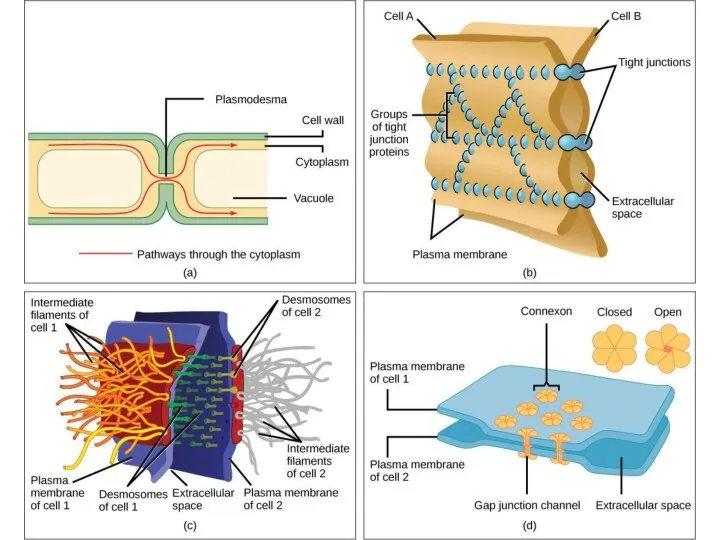

Межклеточные контакты необходимы для
соединения клеток
информационного обмена
а) межклеточного
б) с внешней средой
Для
Межклеточные контакты необходимы для
соединения клеток
информационного обмена
а) межклеточного
б) с внешней средой
Для

Ионные каналы
Ионные каналы
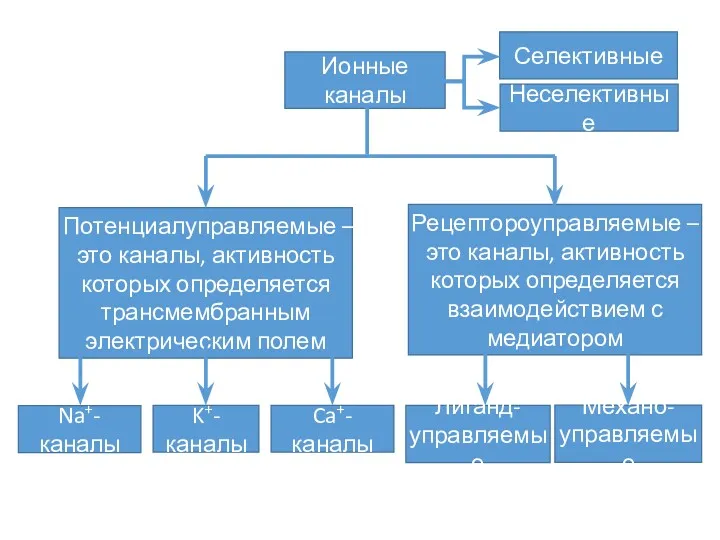

Общее представление о структуре и функциях ионных каналов
Каналообразующие белковые молекулы всех
Общее представление о структуре и функциях ионных каналов
Каналообразующие белковые молекулы всех
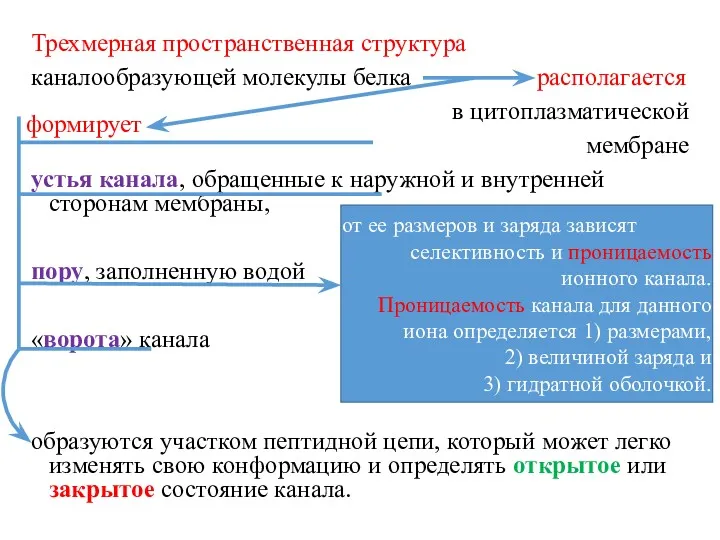
Трехмерная пространственная структура
каналообразующей молекулы белка располагается
в цитоплазматической
мембране
устья канала, обращенные
Трехмерная пространственная структура
каналообразующей молекулы белка располагается
в цитоплазматической
мембране
устья канала, обращенные
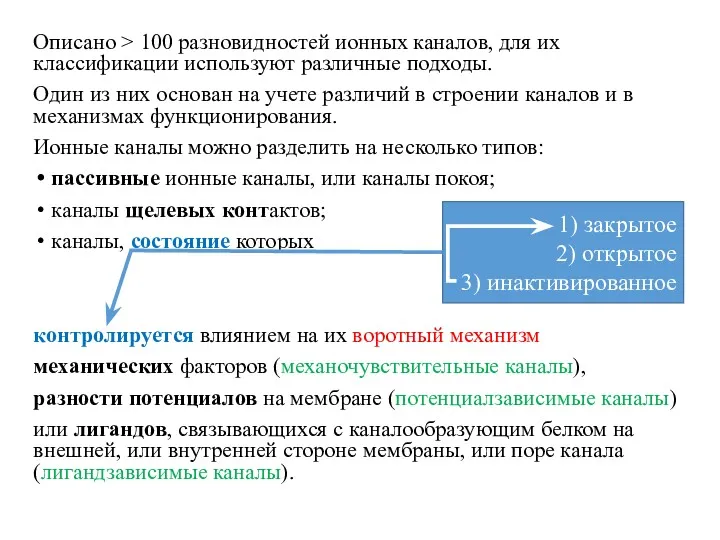
Описано > 100 разновидностей ионных каналов, для их классификации используют различные
Описано > 100 разновидностей ионных каналов, для их классификации используют различные

Пассивные каналы
могут быть открытыми (активными) в покоящихся клетках, т.е. в отсутствие
Пассивные каналы
могут быть открытыми (активными) в покоящихся клетках, т.е. в отсутствие

Каналы щелевых (плотных) контактов клеток
Каналы щелевых контактов формируются в области контакта
Каналы щелевых (плотных) контактов клеток
Каналы щелевых контактов формируются в области контакта
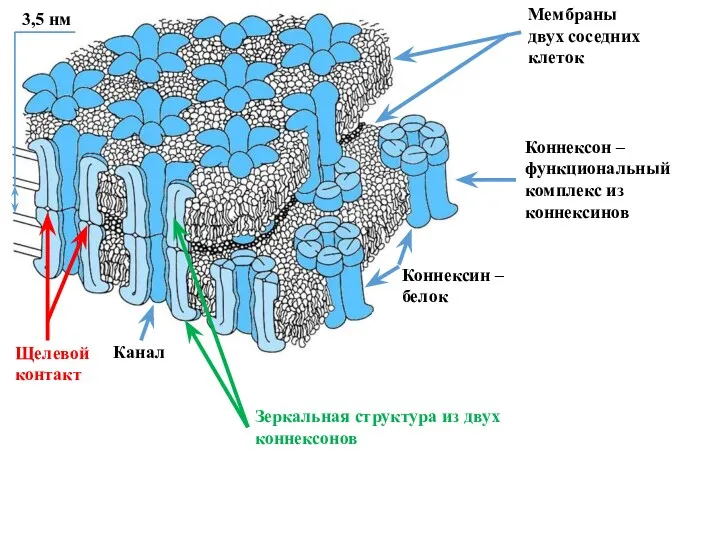
Мембраны
двух соседних
клеток
Канал
Щелевой
контакт
3,5 нм
Коннексин – белок
Коннексон – функциональный комплекс
Мембраны
двух соседних
клеток
Канал
Щелевой
контакт
3,5 нм
Коннексин – белок
Коннексон – функциональный комплекс
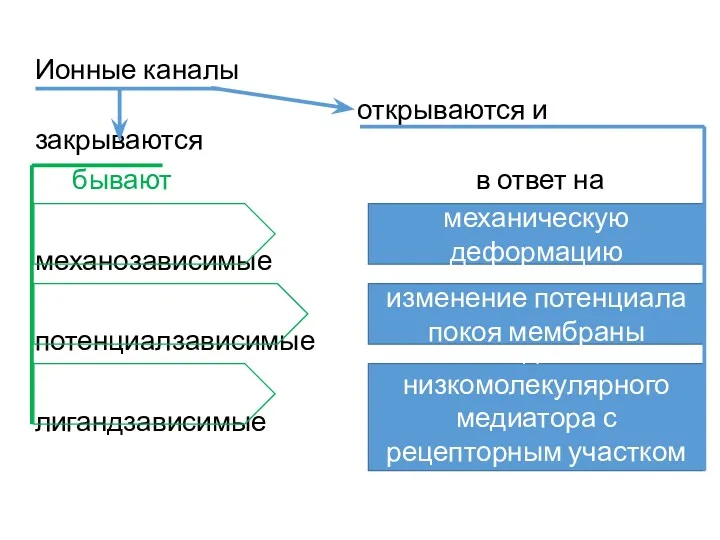
Ионные каналы
открываются и закрываются
бывают в ответ на
механозависимые
потенциалзависимые
лигандзависимые
взаимодействие низкомолекулярного
Ионные каналы
открываются и закрываются
бывают в ответ на
механозависимые
потенциалзависимые
лигандзависимые
взаимодействие низкомолекулярного

Потенциалзависимые каналы
Состояние этих каналов управляется силами электрического поля, создаваемого величиной разности
Потенциалзависимые каналы
Состояние этих каналов управляется силами электрического поля, создаваемого величиной разности
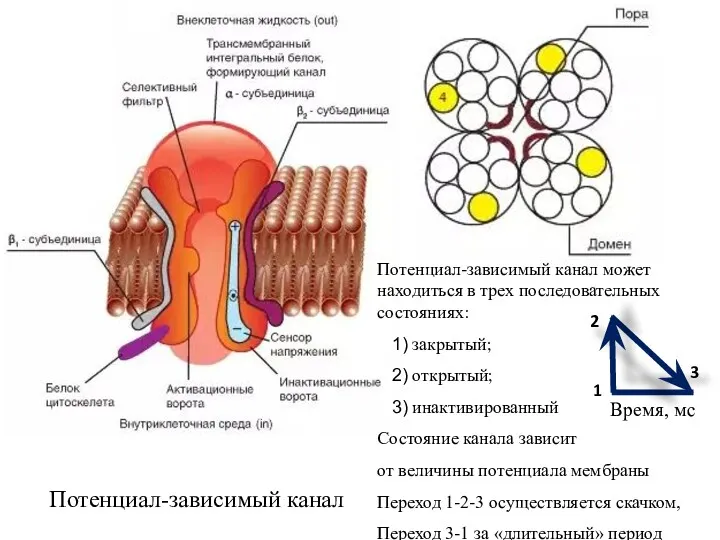
Потенциал-зависимый канал может находиться в трех последовательных состояниях:
закрытый;
открытый;
инактивированный
Состояние
Потенциал-зависимый канал может находиться в трех последовательных состояниях:
закрытый;
открытый;
инактивированный
Состояние

Скорость открытия активационных ворот может быть
очень высокой
и
Скорость открытия активационных ворот может быть
очень высокой
и
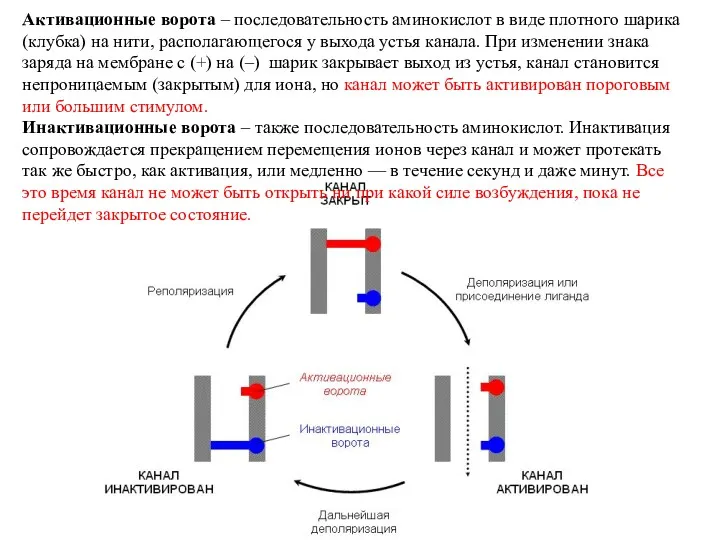
Активационные ворота – последовательность аминокислот в виде плотного шарика (клубка) на
Активационные ворота – последовательность аминокислот в виде плотного шарика (клубка) на

Существуют вещества, блокирующие работу потенциалзависимых ионных каналов. Таким образом, возможно еще
Существуют вещества, блокирующие работу потенциалзависимых ионных каналов. Таким образом, возможно еще
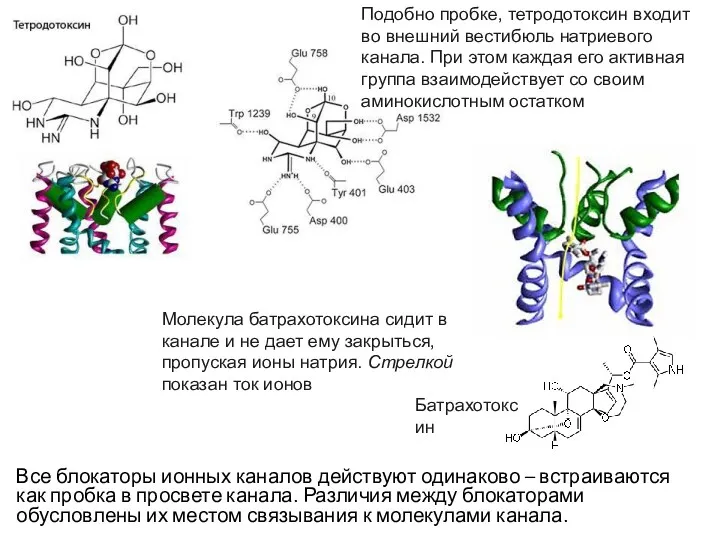
Все блокаторы ионных каналов действуют одинаково – встраиваются как пробка в
Все блокаторы ионных каналов действуют одинаково – встраиваются как пробка в

Порообразующая часть ионного канала может представлять собой
1) единый полипептид, организованный в
Порообразующая часть ионного канала может представлять собой
1) единый полипептид, организованный в
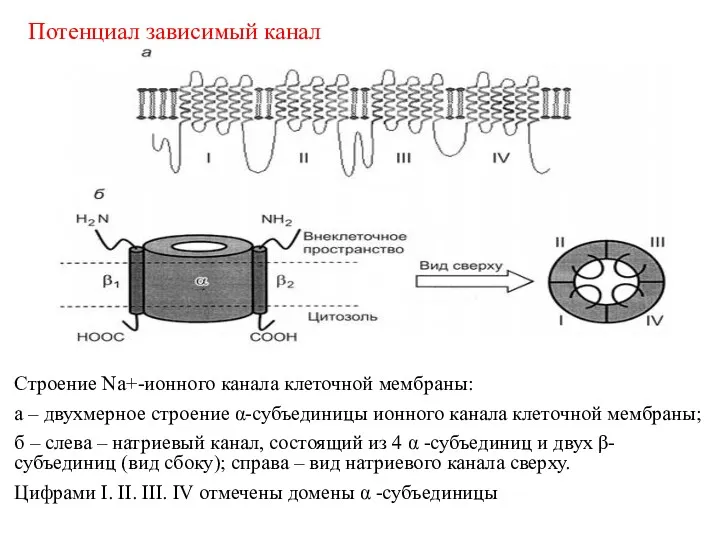
Строение Na+-ионного канала клеточной мембраны:
а – двухмерное строение α-субъединицы ионного
Строение Na+-ионного канала клеточной мембраны:
а – двухмерное строение α-субъединицы ионного

Нарушение работы ионных каналов часто приводят к заболеваниям — каналопатиям. Основная
Нарушение работы ионных каналов часто приводят к заболеваниям — каналопатиям. Основная
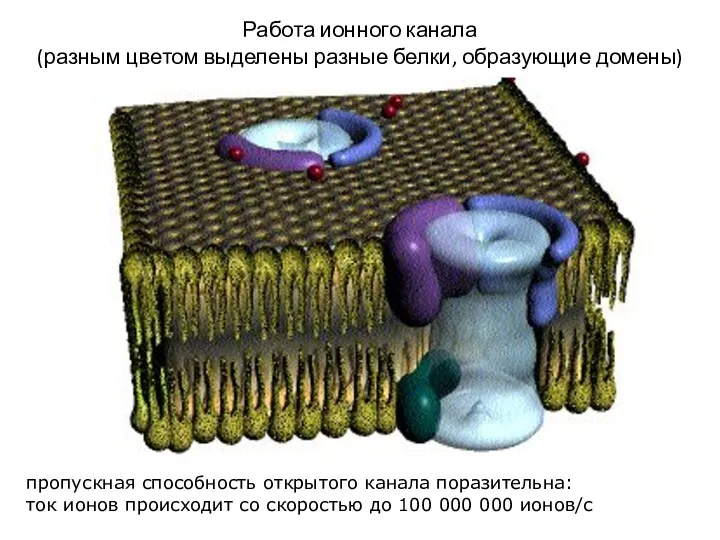
Работа ионного канала
(разным цветом выделены разные белки, образующие домены)
пропускная способность
Работа ионного канала
(разным цветом выделены разные белки, образующие домены)
пропускная способность

Лигандзависимые ионные каналы
Формируются белковыми макромолекулами, одновременно выполняющими функцию
ионных каналов
Лигандзависимые ионные каналы
Формируются белковыми макромолекулами, одновременно выполняющими функцию
ионных каналов
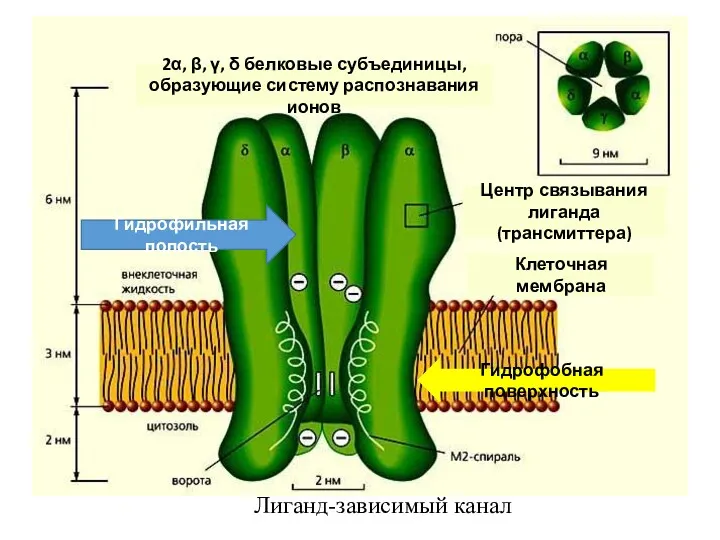
Лиганд-зависимый канал
Центр связывания лиганда (трансмиттера)
Клеточная мембрана
2α, β, γ, δ белковые субъединицы,
Лиганд-зависимый канал
Центр связывания лиганда (трансмиттера)
Клеточная мембрана
2α, β, γ, δ белковые субъединицы,
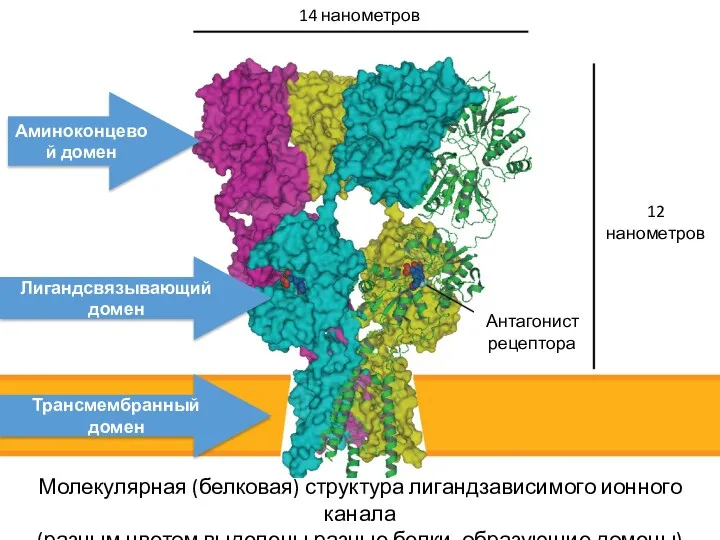
14 нанометров
12 нанометров
Антагонист рецептора
Трансмембранный домен
Лигандсвязывающий домен
Аминоконцевой домен
Молекулярная (белковая) структура лигандзависимого ионного
14 нанометров
12 нанометров
Антагонист рецептора
Трансмембранный домен
Лигандсвязывающий домен
Аминоконцевой домен
Молекулярная (белковая) структура лигандзависимого ионного

Лигандзависимые ионные каналы обычно локализованы в постсинаптических мембранах нервных клеток и
Лигандзависимые ионные каналы обычно локализованы в постсинаптических мембранах нервных клеток и

Лигандзависимые каналы могут изменять проницаемость для катионов Na+ и К+ или
Лигандзависимые каналы могут изменять проницаемость для катионов Na+ и К+ или

Существуют медленные лигандзависимые ионные каналы.
Их открытие реализуется через цепь последовательных
Существуют медленные лигандзависимые ионные каналы.
Их открытие реализуется через цепь последовательных
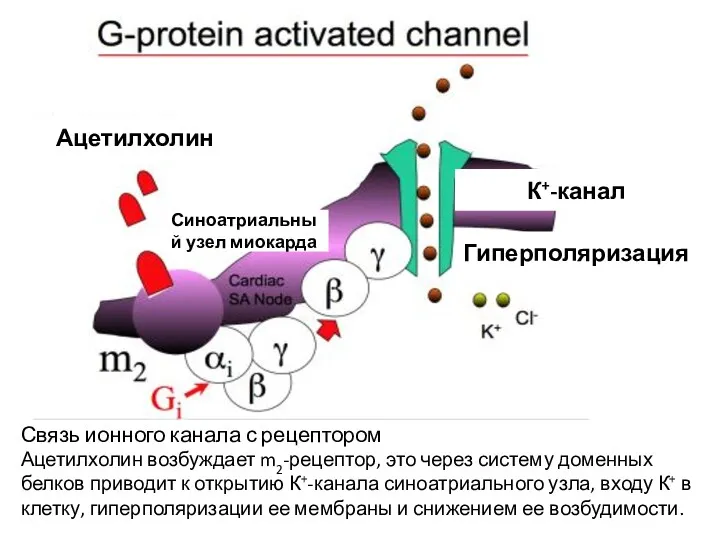
Ацетилхолин
К+-канал
Гиперполяризация
Синоатриальный узел миокарда
Связь ионного канала с рецептором
Ацетилхолин возбуждает m2-рецептор, это через
Ацетилхолин
К+-канал
Гиперполяризация
Синоатриальный узел миокарда
Связь ионного канала с рецептором
Ацетилхолин возбуждает m2-рецептор, это через

Особым типом лигандзависимых каналов являются каналы, локализованные в мембранах эндоплазматического ретикулума
Особым типом лигандзависимых каналов являются каналы, локализованные в мембранах эндоплазматического ретикулума

Рецепторы
РЕЦЕПЦИЯ – процесс восприятия и трансформации (преобразования) энергии внешнего раздражителя в
Рецепторы
РЕЦЕПЦИЯ – процесс восприятия и трансформации (преобразования) энергии внешнего раздражителя в
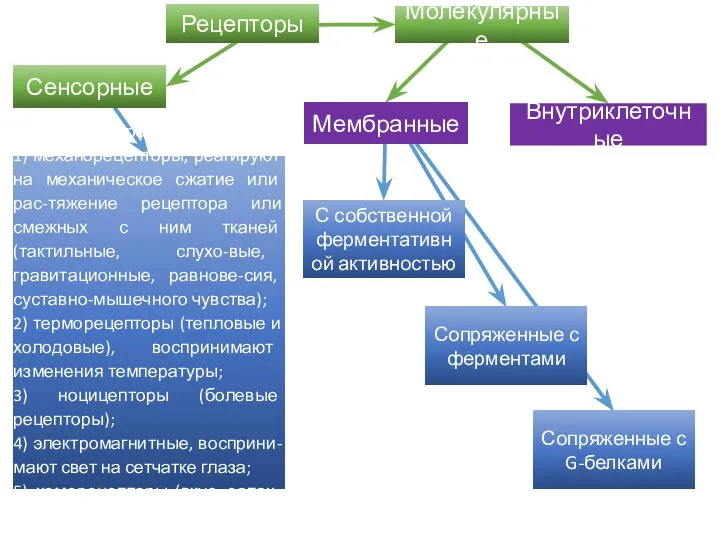
Сенсорные
Мембранные
Внутриклеточные
Специализированные клетки:
1) механорецепторы, реагируют на механическое сжатие или рас-тяжение рецептора или
Сенсорные
Мембранные
Внутриклеточные
Специализированные клетки:
1) механорецепторы, реагируют на механическое сжатие или рас-тяжение рецептора или
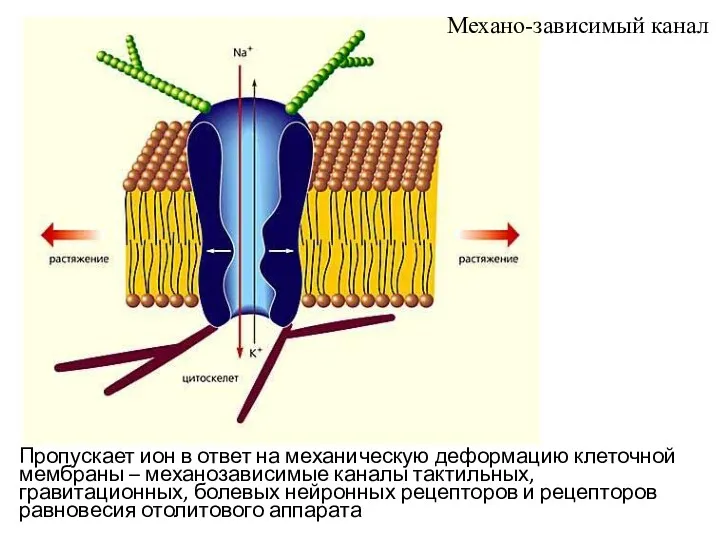
Пропускает ион в ответ на механическую деформацию клеточной мембраны – механозависимые
Пропускает ион в ответ на механическую деформацию клеточной мембраны – механозависимые

Сенсорные рецепторы
Сенсорные рецепторы
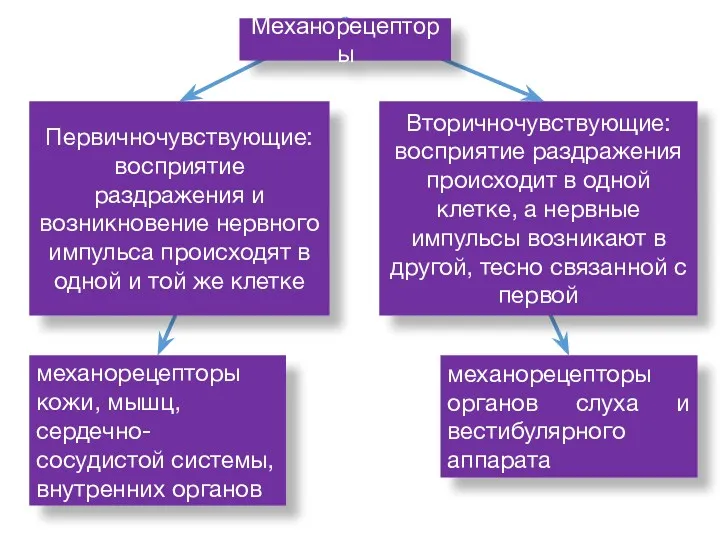
механорецепторы кожи, мышц,
сердечно-сосудистой системы,
внутренних органов
механорецепторы органов слуха и вестибулярного
механорецепторы кожи, мышц,
сердечно-сосудистой системы,
внутренних органов
механорецепторы органов слуха и вестибулярного
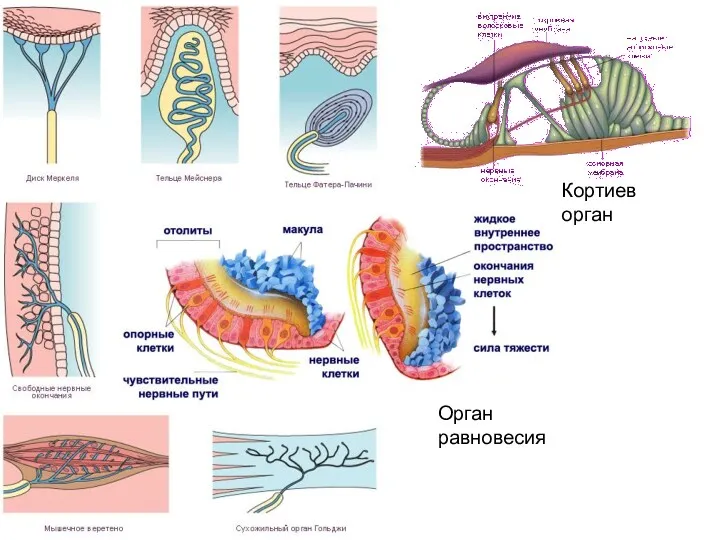
Кортиев орган
Орган равновесия
Кортиев орган
Орган равновесия
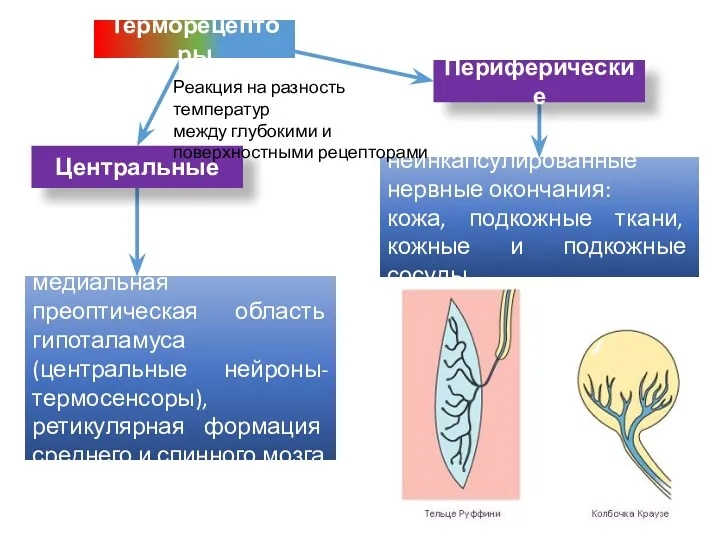
Периферические
неинкапсулированные нервные окончания:
кожа, подкожные ткани, кожные и подкожные сосуды
медиальная преоптическая область
Периферические
неинкапсулированные нервные окончания:
кожа, подкожные ткани, кожные и подкожные сосуды
медиальная преоптическая область
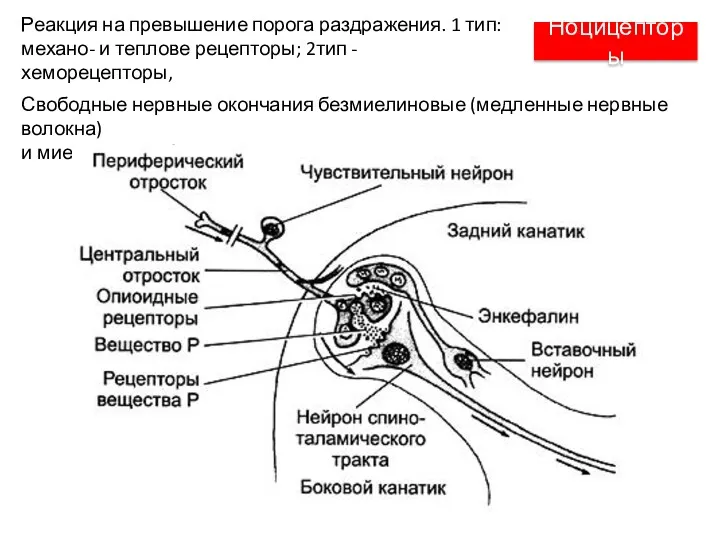
Ноцицепторы
Свободные нервные окончания безмиелиновые (медленные нервные волокна)
и миелиновые (быстрые нервные волокна)
Реакция
Ноцицепторы
Свободные нервные окончания безмиелиновые (медленные нервные волокна)
и миелиновые (быстрые нервные волокна)
Реакция
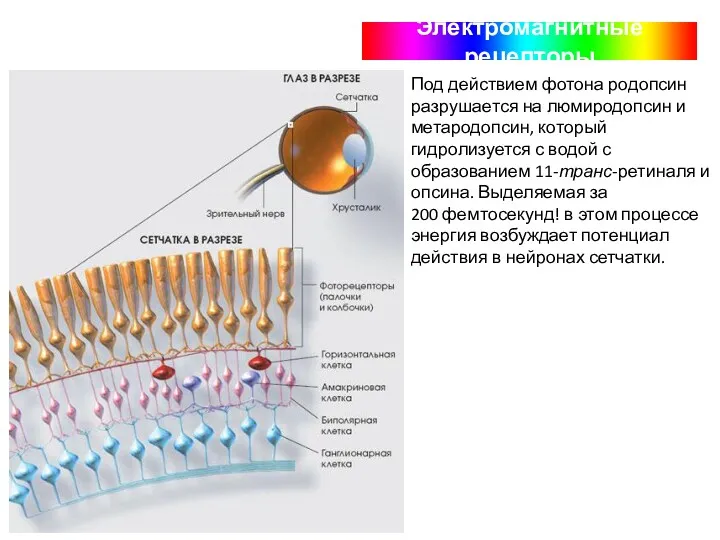
Электромагнитные рецепторы
Под действием фотона родопсин разрушается на люмиродопсин и метародопсин, который
Электромагнитные рецепторы
Под действием фотона родопсин разрушается на люмиродопсин и метародопсин, который
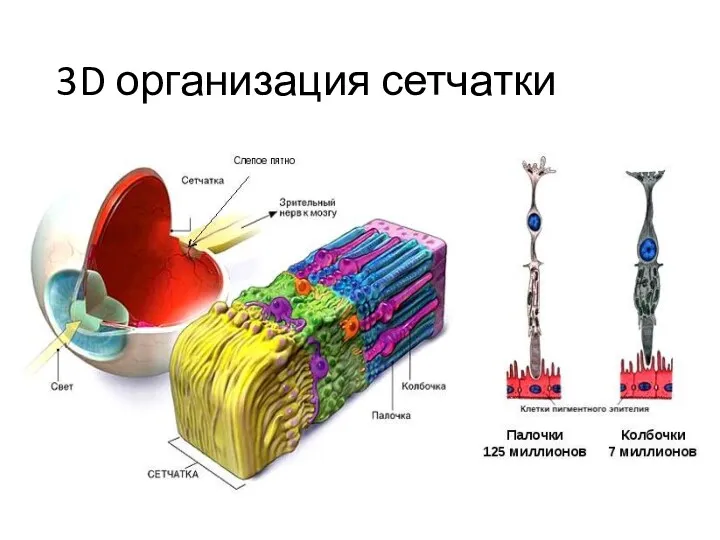
3D организация сетчатки
3D организация сетчатки
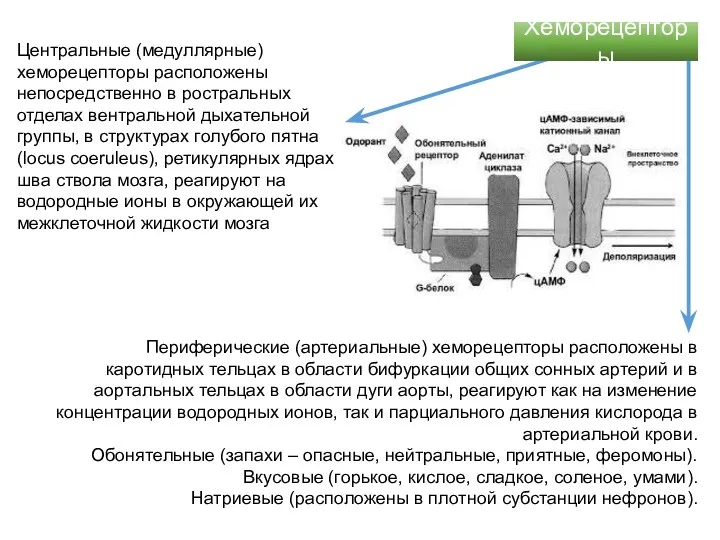
Центральные (медуллярные) хеморецепторы расположены непосредственно в ростральных отделах вентральной дыхательной группы,
Центральные (медуллярные) хеморецепторы расположены непосредственно в ростральных отделах вентральной дыхательной группы,
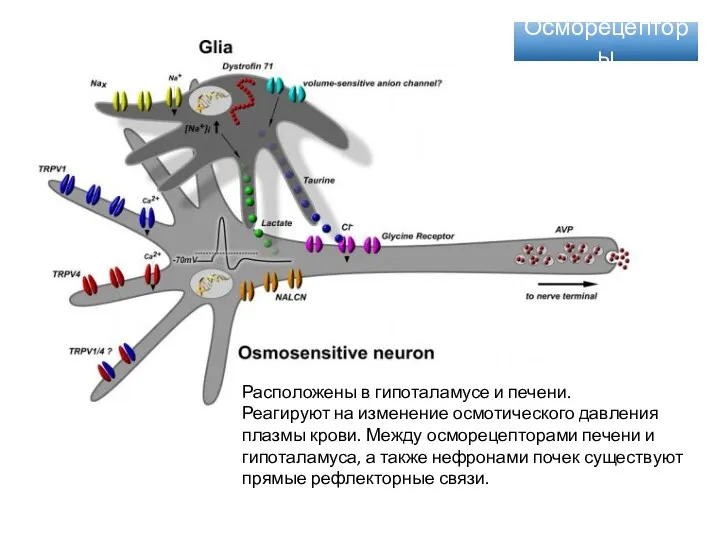
Осморецепторы
Расположены в гипоталамусе и печени.
Реагируют на изменение осмотического давления плазмы крови.
Осморецепторы
Расположены в гипоталамусе и печени.
Реагируют на изменение осмотического давления плазмы крови.

Молекулярные рецепторы
Молекулярные рецепторы
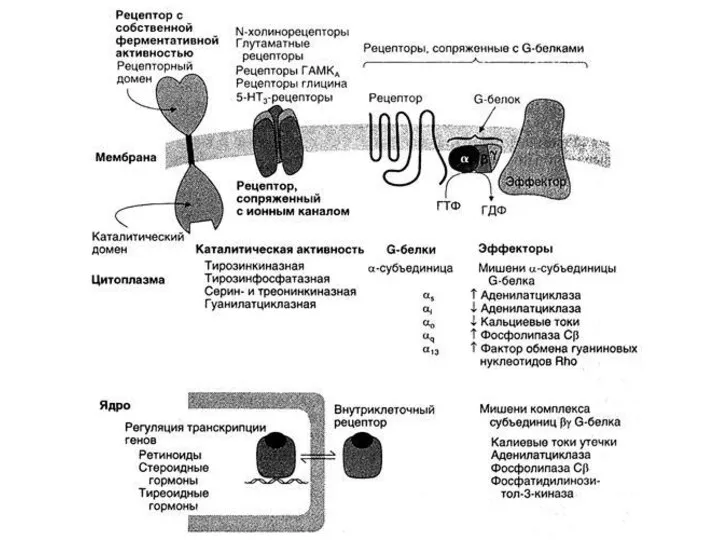
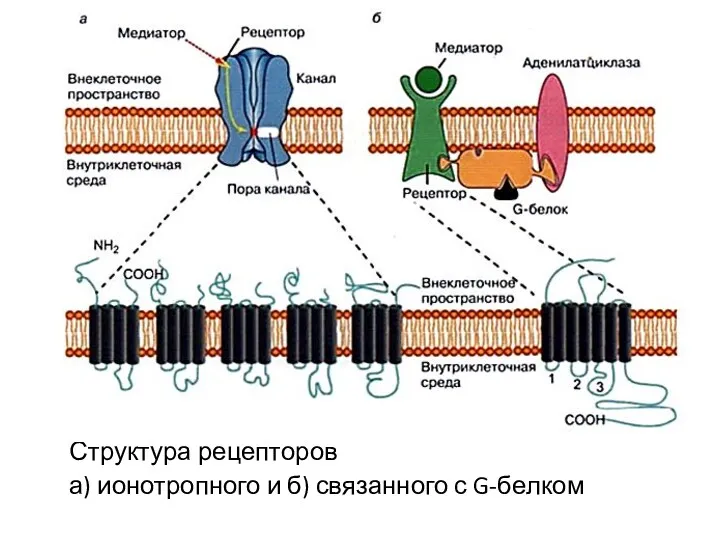
Структура рецепторов
а) ионотропного и б) связанного с G-белком
Структура рецепторов
а) ионотропного и б) связанного с G-белком

Мембранные рецепторы сопряженные с ионными каналами
Механизм работы:
Активация рецептора
Мембранные рецепторы сопряженные с ионными каналами
Механизм работы:
Активация рецептора
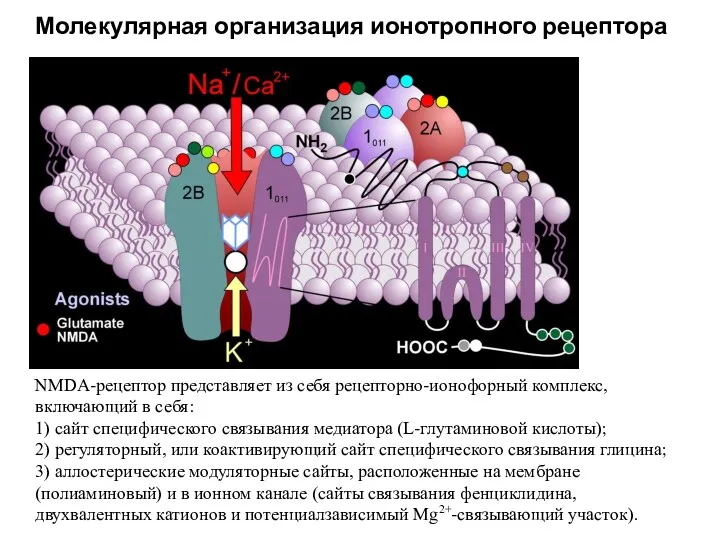
NMDA-рецептор представляет из себя рецепторно-ионофорный комплекс, включающий в себя:
1) сайт специфического
NMDA-рецептор представляет из себя рецепторно-ионофорный комплекс, включающий в себя:
1) сайт специфического

Мембранные рецепторы сопряженные с G-белками
Механизм работы:
Активация рецептора
Активация
Мембранные рецепторы сопряженные с G-белками
Механизм работы:
Активация рецептора
Активация
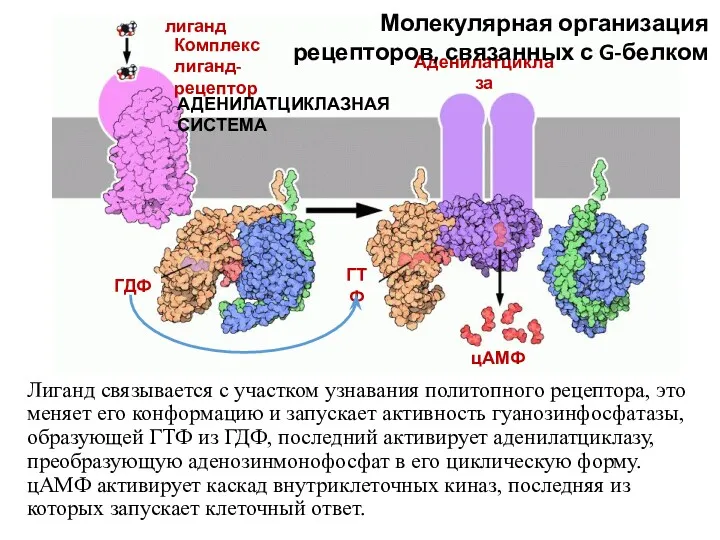
Лиганд связывается с участком узнавания политопного рецептора, это меняет его конформацию
Лиганд связывается с участком узнавания политопного рецептора, это меняет его конформацию
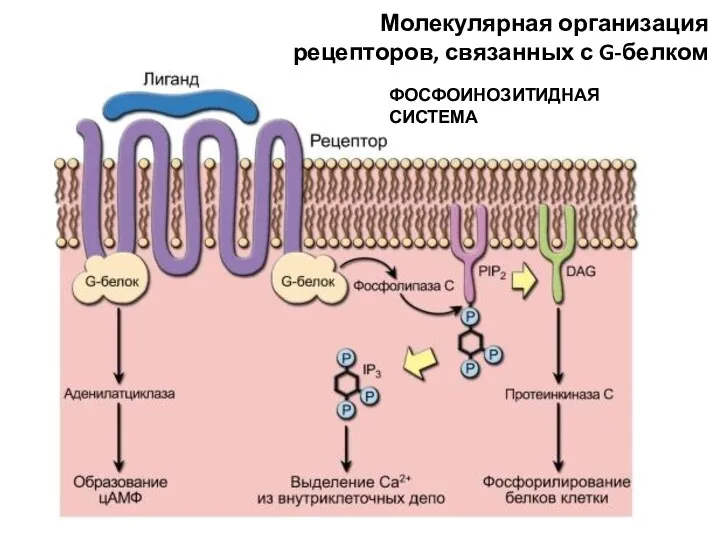
Молекулярная организация рецепторов, связанных с G-белком
ФОСФОИНОЗИТИДНАЯ СИСТЕМА
Молекулярная организация рецепторов, связанных с G-белком
ФОСФОИНОЗИТИДНАЯ СИСТЕМА

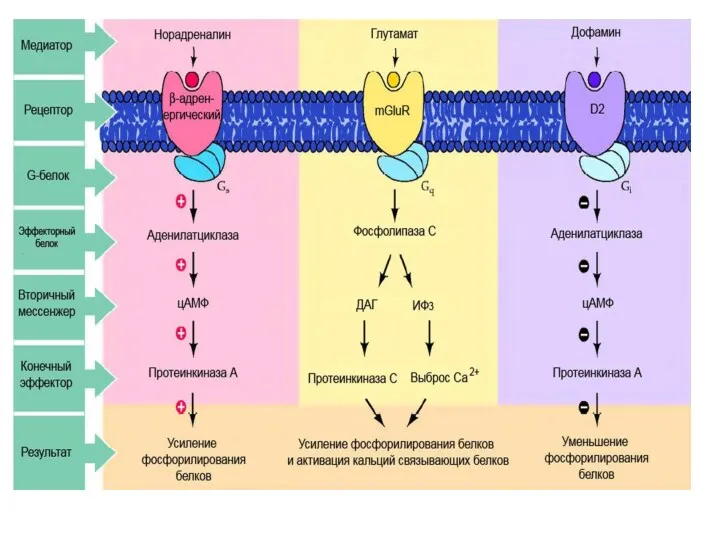
G-белки двух типов:
GS – стимулирующие клеточный ответ
GI – ингибирующие клеточный ответ
Лиганды
G-белки двух типов:
GS – стимулирующие клеточный ответ
GI – ингибирующие клеточный ответ
Лиганды
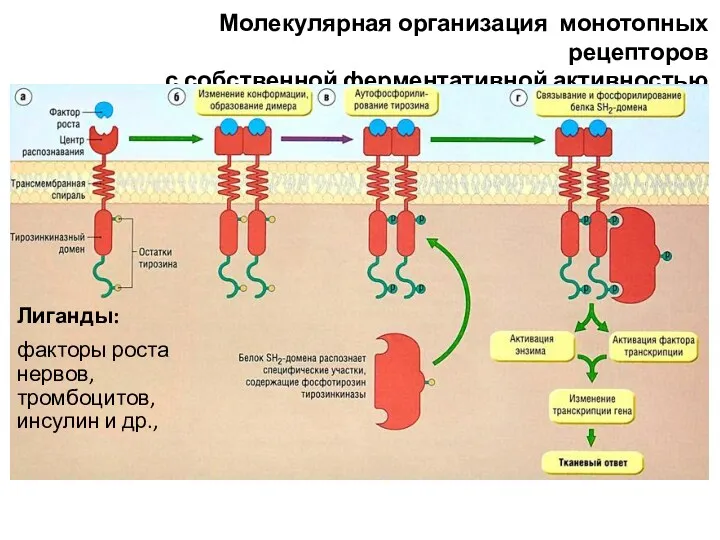
Молекулярная организация монотопных рецепторов
с собственной ферментативной активностью
Лиганды:
факторы роста нервов, тромбоцитов,
Молекулярная организация монотопных рецепторов
с собственной ферментативной активностью
Лиганды:
факторы роста нервов, тромбоцитов,
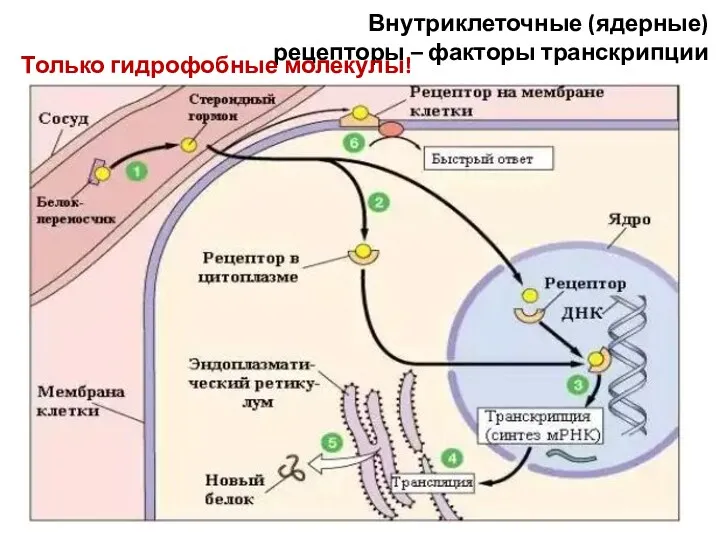
Внутриклеточные (ядерные) рецепторы – факторы транскрипции
Только гидрофобные молекулы!
Внутриклеточные (ядерные) рецепторы – факторы транскрипции
Только гидрофобные молекулы!
 Молекулярно-генетические механизмы развития корня
Молекулярно-генетические механизмы развития корня Интерсекс. Терминология и идентичность. Гендерная идентичность
Интерсекс. Терминология и идентичность. Гендерная идентичность Хромосомная теория наследственности. Сцепленное наследование
Хромосомная теория наследственности. Сцепленное наследование Теория И.П. Павлова о типах высшей нервной деятельности
Теория И.П. Павлова о типах высшей нервной деятельности Строение семян двудольных и однодольных растений
Строение семян двудольных и однодольных растений Тонкий кишечник
Тонкий кишечник The muscular system
The muscular system Мы - друзья пернатых
Мы - друзья пернатых Вода - основной источник жизни и здоровья
Вода - основной источник жизни и здоровья Асқазан. Дамуы
Асқазан. Дамуы Різноманітність ссавців
Різноманітність ссавців Биологические ритмы
Биологические ритмы 7 Б класс. Принял эстафету
7 Б класс. Принял эстафету 1 апреля – Международный день птиц
1 апреля – Международный день птиц Проверка знаний
Проверка знаний Внутрішня будова ссавців
Внутрішня будова ссавців Физиология сенсорных систем
Физиология сенсорных систем Квіткові рослини
Квіткові рослини Урок биологии Многообразие ракообразных с презентацией в 7 классе по учебнику Н.И.Сонина
Урок биологии Многообразие ракообразных с презентацией в 7 классе по учебнику Н.И.Сонина Водоросли. Особенности строения, питания, размножения
Водоросли. Особенности строения, питания, размножения Внеклассное мероприятие по биологии в 7-8 классах на тему: Мы- за здоровье!
Внеклассное мероприятие по биологии в 7-8 классах на тему: Мы- за здоровье! Репликация ДНК. (Лекция 3)
Репликация ДНК. (Лекция 3) Витамины
Витамины Фиалка домашняя
Фиалка домашняя Мегаспорогенез және микроспорогенез
Мегаспорогенез және микроспорогенез Бактериофаги. Основная роль
Бактериофаги. Основная роль Урок-игра по теме Системы органов животных
Урок-игра по теме Системы органов животных Полевые цветы. Фото
Полевые цветы. Фото