- Физиология скелетных мышц

Содержание
- 2. Схема строения мышечного волокна Саркомер - с двух сторон ограничен Z – мембранами. Толстые – миозиновые,
- 3. Саркоплазма В саркоплазме находится весь набор типичных для любой клетки органоидов. Особо следует подчеркнуть наличие: -
- 4. Актиновые миофиламенты Актиновые филаменты, скомпанованы из двух актиновых нитей, представляющих собой как бы бусинки глобулярных молекул
- 5. Схема строения актиновых и миозиновых филаментов Миозиновые филаменты образуются более чем двумястами молекулами миозина. Каждая из
- 6. Двигательные единицы К каждому мышечному волокну подходит отросток мотонейрона. Как правило, 1 мотонейрон иннервирует несколько мышечных
- 7. Нервно-мышечный синапс 1 - пресинаптическая мембрана, 2 - пузырьки с ацетилхолином, 3 - митохондрии, 4 -
- 8. Передача ПД через синапс 1 – везикула, 2 – медиатор (ацетилхолин, АХ), 3 – холинорецептор, 4
- 9. Взаимодействие медиатора с постсинаптической мебраной Медиатор (АХ) диффундирует по синаптической жидкости и большая часть молекул его
- 10. Лигандзависимый канал
- 11. Ресинтез АХ Для передачи через синапс 1 ПД требуется около 300 пузырьков с АХ. ПОЭТОМУ НЕОБХОДИМО
- 12. Нарушение синаптической проводимости Некоторые яды могут частично нарушать или полностью блокировать нервно-мышечную передачу. Механизм их действия
- 13. МП Мышечное волокно имеет мембранный потенциал -80 - -90 мВ. Для того, чтобы вызвать возникновение возбуждения
- 14. Явление суммации. Обозначения: а, б - деполяризация не достигает критического уровня, в - результат суммации –
- 15. Для чего необходимо поступление ПД к мышце? ВПСП распространяется по сарколемме. Проскакивает по мембране саркоплазматического ретикулума.
- 16. Депо кальция – саркоплазматический ретикулум 1- миофибриллы, 2 – саркоплазматический ретикулум, 3 – цистерны, 4 –
- 17. Роль кальция в мышечном сокращении Последовательные этапы: а – расслабление, б – соединение миозиновых головок с
- 18. «Шаговый» механизм
- 19. ЭМГ Электрические явления в мышце, связанные с механизмами перехода ПД (возбуждения) можно зарегистрировать вводя в нее
- 20. Различные режимы сокращения мышц А - одиночное сокращение, Б – неполный тетанус, В – полный тетанус.
- 21. Анатомический и физиологический поперечники мышц В естественных условиях на проявление силы мышцы оказывает влияние не только
- 22. Роль АТФ в мышце АТФ в мышце необходима для: а) сокращения (образования мостиков); б) расслабления (разрыва
- 23. Максимальная мощность путей ресинтеза АТФ: а) фосфагенный (КФ) - 3,6 моль АТФ/мин, б) гликолитический - 1,2
- 24. Типы ДЕ (двигательные единицы) - процентное соотношение врожденное и у разных людей различное (спринтеры, стайеры) Быстрые
- 25. Гладкие мышцы Гладкие мышцы находятся в стенке внут-ренних органов, сосудов, коже. Структурной единицей их является вытянутой
- 26. Компановка сократимых миофиламентов внутри клетки. Актиновые филаменты сгруппированы в пучки, которые время от времени образуют уплотнения
- 27. Разновидности деполяризации гладко-мышечных клеток Один из них (а) напоминает ПД скелетной мышцы, отличаясь от нее большей
- 29. Скачать презентацию
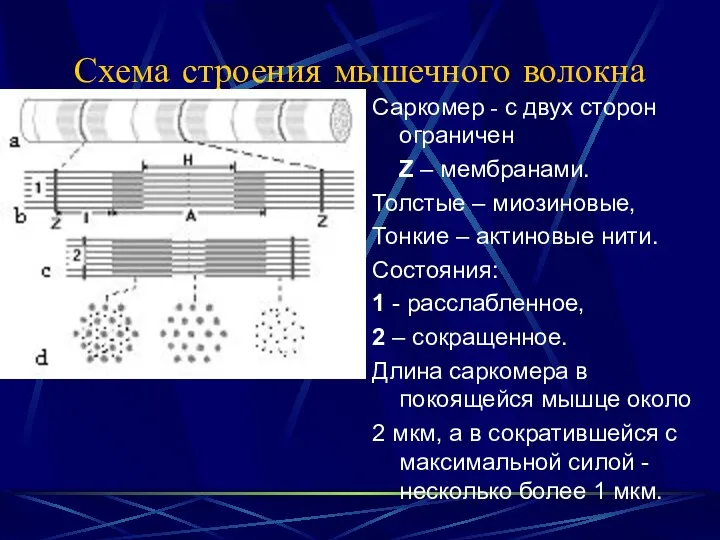
Схема строения мышечного волокна
Саркомер - с двух сторон ограничен
Z
Схема строения мышечного волокна
Саркомер - с двух сторон ограничен
Z

Саркоплазма
В саркоплазме находится весь набор типичных для любой клетки органоидов.
Особо следует
Саркоплазма
В саркоплазме находится весь набор типичных для любой клетки органоидов.
Особо следует

Актиновые миофиламенты
Актиновые филаменты, скомпанованы из двух актиновых нитей, представляющих собой как
Актиновые миофиламенты
Актиновые филаменты, скомпанованы из двух актиновых нитей, представляющих собой как

Схема строения актиновых и миозиновых филаментов
Миозиновые филаменты образуются более чем двумястами
Схема строения актиновых и миозиновых филаментов
Миозиновые филаменты образуются более чем двумястами

Двигательные единицы
К каждому мышечному волокну подходит отросток мотонейрона.
Как правило, 1 мотонейрон
Двигательные единицы
К каждому мышечному волокну подходит отросток мотонейрона.
Как правило, 1 мотонейрон
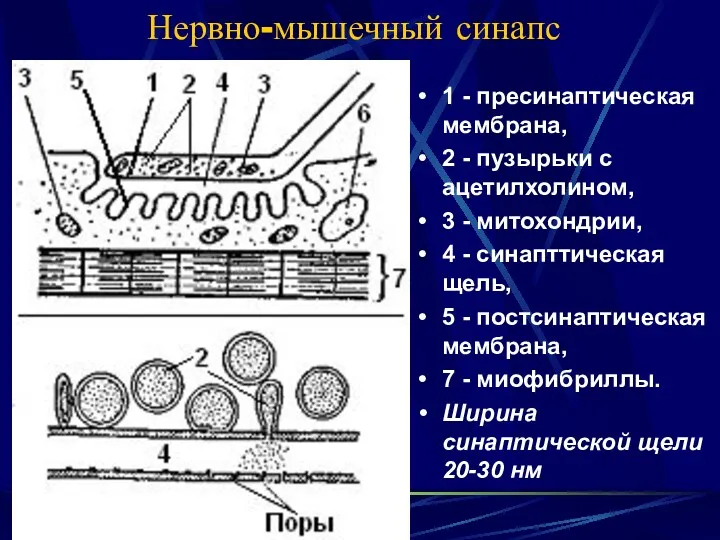
Нервно-мышечный синапс
1 - пресинаптическая мембрана,
2 - пузырьки с ацетилхолином,
3
Нервно-мышечный синапс
1 - пресинаптическая мембрана,
2 - пузырьки с ацетилхолином,
3
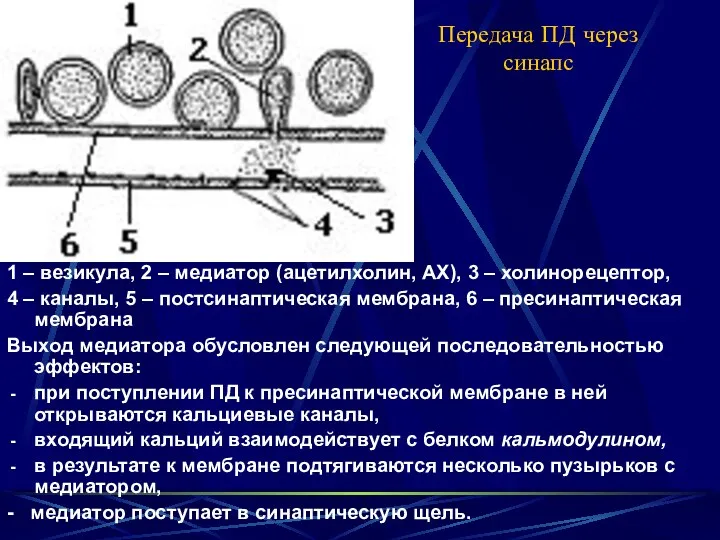
Передача ПД через синапс
1 – везикула, 2 – медиатор (ацетилхолин, АХ),
Передача ПД через синапс
1 – везикула, 2 – медиатор (ацетилхолин, АХ),

Взаимодействие медиатора с постсинаптической мебраной
Медиатор (АХ) диффундирует по синаптической жидкости и
Взаимодействие медиатора с постсинаптической мебраной
Медиатор (АХ) диффундирует по синаптической жидкости и
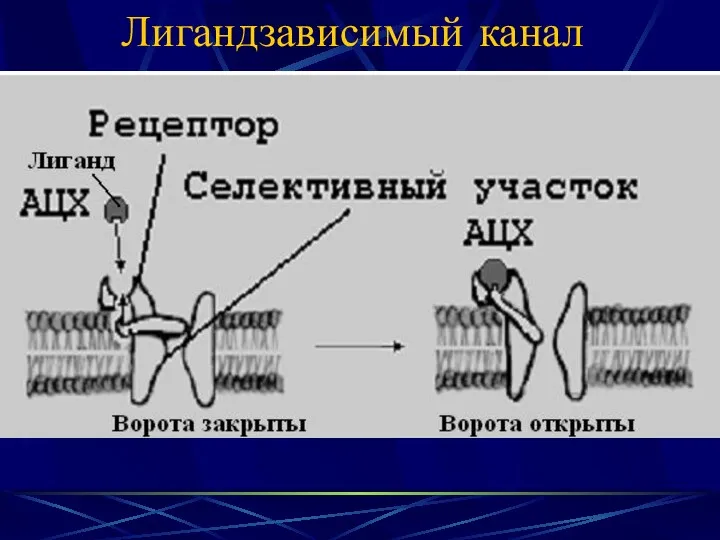
Лигандзависимый канал
Лигандзависимый канал

Ресинтез АХ
Для передачи через синапс 1 ПД требуется около 300 пузырьков
Ресинтез АХ
Для передачи через синапс 1 ПД требуется около 300 пузырьков

Нарушение синаптической проводимости
Некоторые яды могут частично нарушать или полностью блокировать нервно-мышечную
Нарушение синаптической проводимости
Некоторые яды могут частично нарушать или полностью блокировать нервно-мышечную

МП
Мышечное волокно имеет мембранный потенциал -80 - -90 мВ. Для того,
МП
Мышечное волокно имеет мембранный потенциал -80 - -90 мВ. Для того,
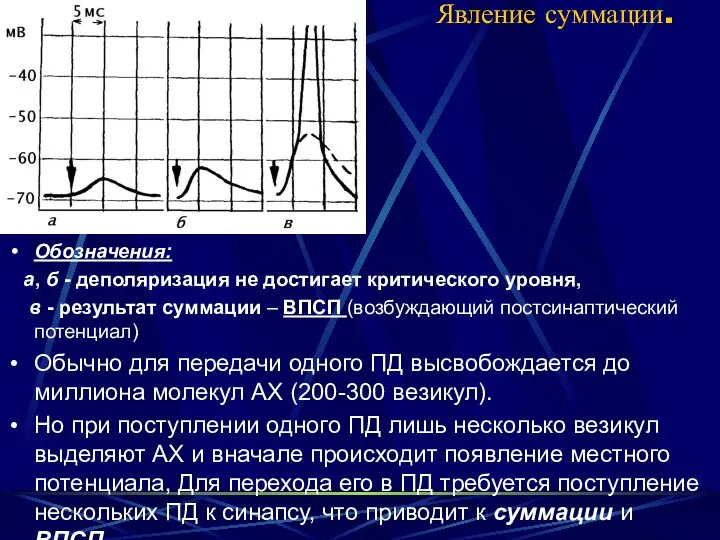
Явление суммации.
Обозначения:
а, б - деполяризация не достигает критического уровня,
Явление суммации.
Обозначения:
а, б - деполяризация не достигает критического уровня,

Для чего необходимо поступление ПД к мышце?
ВПСП распространяется по сарколемме.
Для чего необходимо поступление ПД к мышце?
ВПСП распространяется по сарколемме.
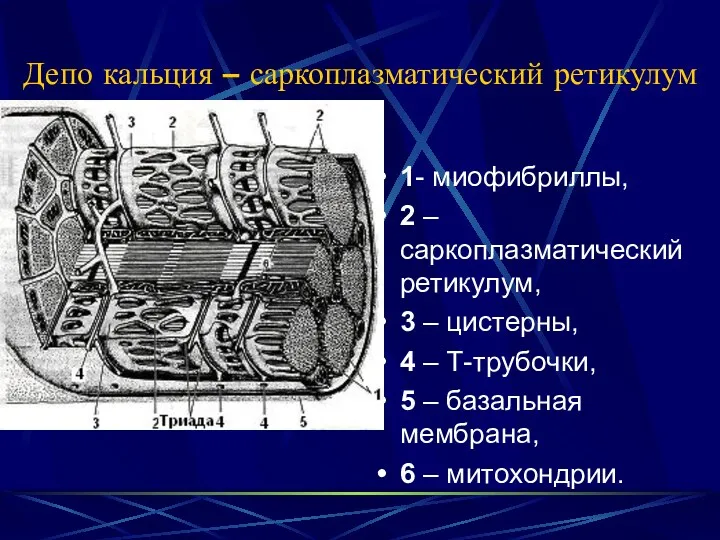
Депо кальция – саркоплазматический ретикулум
1- миофибриллы,
2 – саркоплазматический ретикулум,
Депо кальция – саркоплазматический ретикулум
1- миофибриллы,
2 – саркоплазматический ретикулум,

Роль кальция в мышечном сокращении
Последовательные этапы:
а – расслабление,
б – соединение миозиновых
Роль кальция в мышечном сокращении
Последовательные этапы:
а – расслабление,
б – соединение миозиновых
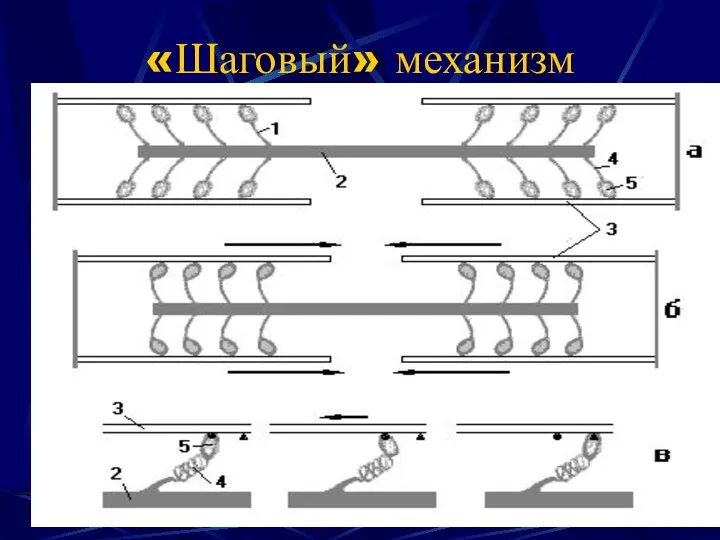
«Шаговый» механизм
«Шаговый» механизм

ЭМГ
Электрические явления в мышце, связанные с механизмами перехода ПД (возбуждения) можно
ЭМГ
Электрические явления в мышце, связанные с механизмами перехода ПД (возбуждения) можно
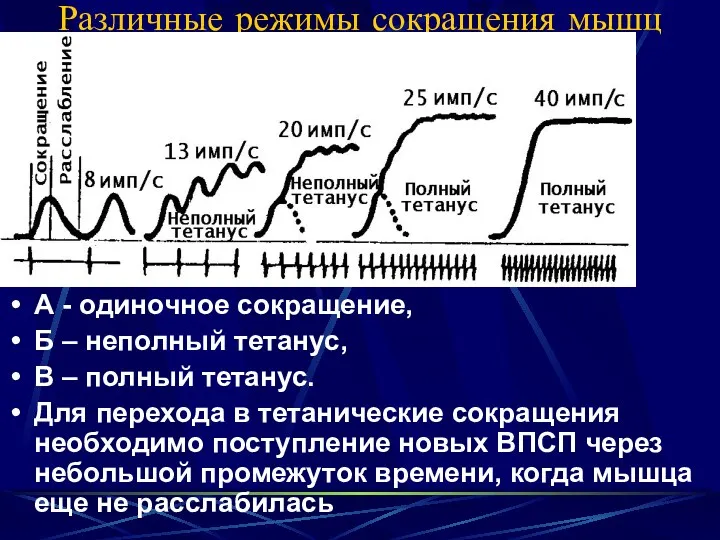
Различные режимы сокращения мышц
А - одиночное сокращение,
Б – неполный тетанус,
Различные режимы сокращения мышц
А - одиночное сокращение,
Б – неполный тетанус,

Анатомический и физиологический поперечники мышц
В естественных условиях на проявление силы мышцы
Анатомический и физиологический поперечники мышц
В естественных условиях на проявление силы мышцы

Роль АТФ в мышце
АТФ в мышце необходима для: а) сокращения (образования
Роль АТФ в мышце
АТФ в мышце необходима для: а) сокращения (образования

Максимальная мощность путей ресинтеза АТФ:
а) фосфагенный (КФ) - 3,6 моль АТФ/мин,
Максимальная мощность путей ресинтеза АТФ:
а) фосфагенный (КФ) - 3,6 моль АТФ/мин,

Типы ДЕ (двигательные единицы)
- процентное соотношение врожденное и у разных людей
Типы ДЕ (двигательные единицы) - процентное соотношение врожденное и у разных людей

Гладкие мышцы
Гладкие мышцы находятся в стенке внут-ренних органов, сосудов, коже. Структурной
Гладкие мышцы
Гладкие мышцы находятся в стенке внут-ренних органов, сосудов, коже. Структурной

Компановка сократимых миофиламентов внутри клетки.
Актиновые филаменты сгруппированы в пучки, которые время
Компановка сократимых миофиламентов внутри клетки.
Актиновые филаменты сгруппированы в пучки, которые время
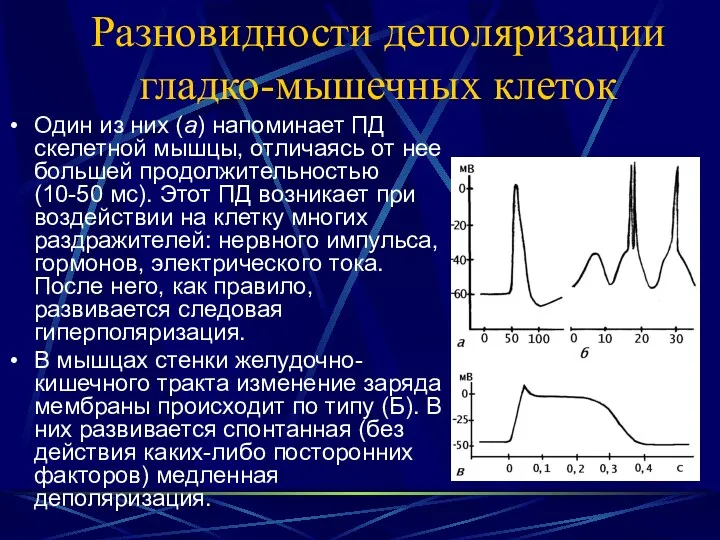
Разновидности деполяризации гладко-мышечных клеток
Один из них (а) напоминает ПД скелетной
Разновидности деполяризации гладко-мышечных клеток
Один из них (а) напоминает ПД скелетной
 Тренировочные занятия по подготовке к ЕГЭ. Задачи по цитологии и генетике. (11 класс)
Тренировочные занятия по подготовке к ЕГЭ. Задачи по цитологии и генетике. (11 класс) Пресноводная гидра
Пресноводная гидра презентация биосинтез белков
презентация биосинтез белков Соя – самый трансгенный вид
Соя – самый трансгенный вид Редокс-статус клетки, окислительный стресс и митохондрии
Редокс-статус клетки, окислительный стресс и митохондрии Органы чувств животных
Органы чувств животных красная книга Марий Эл
красная книга Марий Эл Лекарственные растения
Лекарственные растения Красители. Классификация. Приготовление красителей. Артефакты. Методы микроскопии
Красители. Классификация. Приготовление красителей. Артефакты. Методы микроскопии Особенности молекулярной эволюции
Особенности молекулярной эволюции Биологическая подвижность
Биологическая подвижность Отдел моховидные
Отдел моховидные Царство растений
Царство растений Медицинская гельминтология
Медицинская гельминтология Хоботные
Хоботные Группа ископаемых высших приматов австралопитеки
Группа ископаемых высших приматов австралопитеки Человек родился. Биологическая наследственность
Человек родился. Биологическая наследственность Регуляция обмена веществ
Регуляция обмена веществ Биологическое окисление
Биологическое окисление Медицинская арахноэнтомология
Медицинская арахноэнтомология Мир насекомых
Мир насекомых Сердечно-сосудистая система
Сердечно-сосудистая система Энергетический обмен
Энергетический обмен Сахарозаменители. Натуральные сахарозаменители
Сахарозаменители. Натуральные сахарозаменители Природа родного края
Природа родного края Властивості та характеристики екосистем. Типи зв'язків між популяціями в екосистемі
Властивості та характеристики екосистем. Типи зв'язків між популяціями в екосистемі Мутації людини
Мутації людини Простейшие и кишечнополостные. Игра
Простейшие и кишечнополостные. Игра