- Физиология возбудимых тканей. Часть 1

Содержание
- 2. ЭЛЕКТРОФИЗИОЛОГИЯ При регуляции в организме нервная система использует биопотенциалы. С их помощью осуществляется: А)передача информации (с
- 3. Строение клеточной мембраны Мембрана представляет собой двойной слой фосфолипидов, в который включены другие группы липидов и
- 4. ЛИПИДЫ КЛЕТОЧНОЙ МЕМБРАНЫ 1группа. Фосфолипиды. Составляют около 80 % от массы липидов мембраны. Представлены в основном
- 5. При помещении в водную фазу фосфолипиды образуют: 2 группа.Сфинголипиды (содержат аминоспирт-сфингозин). Образуют миелиновую оболочку нервных клеток
- 6. БЕЛКИ КЛЕТОЧНОЙ МЕМБРАНЫ составляют до 50—60 % сухой массы мембраны Функциональные группы белков: белки-каналы (обеспечивают опосредованный
- 7. Транспорт веществ через мембрану Прямой транспорт предполагает диффузию через мембрану гидрофобных молекул и гидрофильных через разрывы
- 8. Облегченная диффузия с участием каналов. Облегченная диффузия подчиняется законам диффузии. Ее скорость описывается уравнением Фика. Направление
- 9. Транспорт с участием переносчика. Предполагает существование в мембране белка переносчика, который связывается с переносимым веществом, без
- 10. Активный транспорт. Предполагает энергозависимый перенос ионов и молекул. Он может быть без изменения структуры мембраны (участие
- 11. ИТОГО Биопотенциалы чаще создаются в результате простой диффузии ионов через мемебрану Диффузия каждого иона зависит от
- 12. Количественные показатели :равновестный потенциал и электрохимический Равновестный потенциал для конкретного иона-мембранный потенциал, при котором Концентрационный и
- 13. Общая характеристика возбудимых тканей Свойства возбудимых тканей: возбудимость проводимость сократимость секреторной активность лабильность Возбудимость - способность
- 14. Мембранно-ионная теория происхождения потенциала покоя
- 15. Физиологический смысл ПП Электрохимический градиент для К невелик: концентрационный и электрический градиенты направлены взаимнопротивоположно и ПП
- 16. Уравнение Нернста где Е калия - равновесный калиевый потенциал, R -универсальная газовая постоянная, Т - температура
- 17. В процессе поляризации мембраны клетка теряет катионы калия и получает ионы натрия и хлора, однако нарушения
- 18. Значит, потенциал покоя: Создается неравновесным распределением калия, натрия, хлора и кальция внутри клетки по отношению к
- 19. Мембранно-ионная теория происхождения потенциала действия Потенциал действия, или импульсное возбуждение, возникает в ответ на пороговые или
- 20. Работа натриевого канала (3 состояния) Математическая модель, описывающая работу натриевого канала, предполагает существование в его составе
- 21. Работа калиевого канала Математическая модель, описывающая работу калиевого канала, предполагает в структуре одну субъединицу, аналогичную легкой
- 22. Потенциал действия Фазы потенциала действия (ПД): медленной деполяризации (1), быстрой деполяризации (2), реполяризации (3), следового отрицательного
- 23. Значит, потенциал действия: Возникает в ответ на пороговые и сверхпороговые раздражители; Обусловлен увеличением проницаемости мембраны для
- 24. Локальный ответ. Возникает в ответ на раздражители, составляющие не менее 60—80% от порога. Обусловлен увеличением проницаемости
- 26. Скачать презентацию

ЭЛЕКТРОФИЗИОЛОГИЯ
При регуляции в организме нервная система использует биопотенциалы. С их помощью
ЭЛЕКТРОФИЗИОЛОГИЯ
При регуляции в организме нервная система использует биопотенциалы. С их помощью
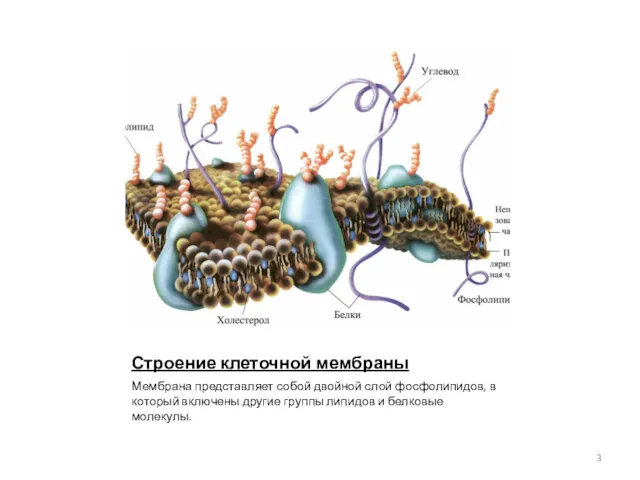
Строение клеточной мембраны
Мембрана представляет собой двойной слой фосфолипидов, в который включены
Строение клеточной мембраны
Мембрана представляет собой двойной слой фосфолипидов, в который включены
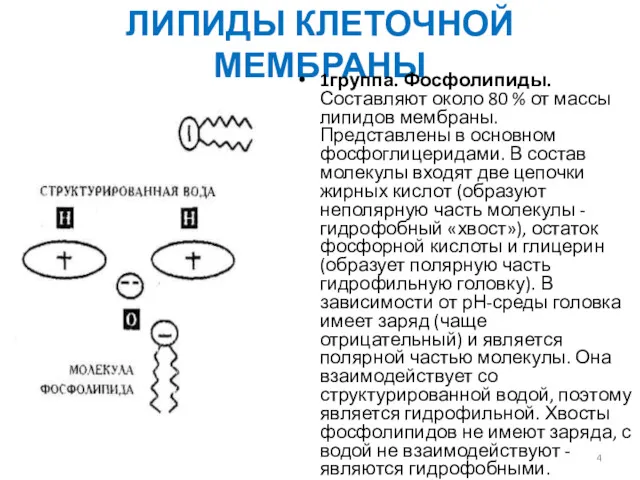
ЛИПИДЫ КЛЕТОЧНОЙ МЕМБРАНЫ
1группа. Фосфолипиды. Составляют около 80 % от массы липидов
ЛИПИДЫ КЛЕТОЧНОЙ МЕМБРАНЫ
1группа. Фосфолипиды. Составляют около 80 % от массы липидов
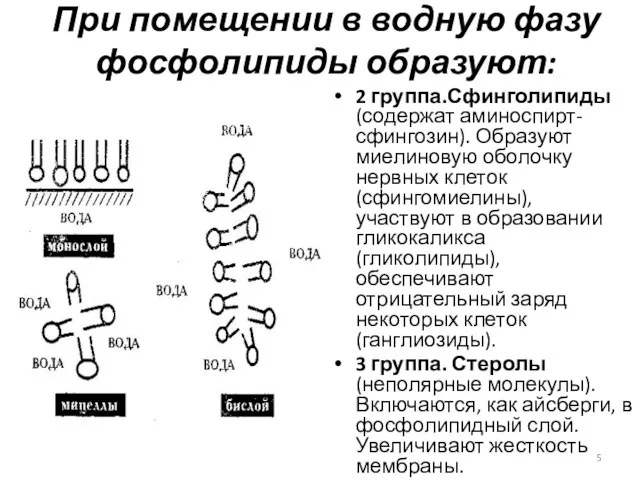
При помещении в водную фазу фосфолипиды образуют:
2 группа.Сфинголипиды (содержат аминоспирт-сфингозин). Образуют
При помещении в водную фазу фосфолипиды образуют:
2 группа.Сфинголипиды (содержат аминоспирт-сфингозин). Образуют

БЕЛКИ КЛЕТОЧНОЙ МЕМБРАНЫ составляют до 50—60 % сухой массы мембраны
Функциональные
БЕЛКИ КЛЕТОЧНОЙ МЕМБРАНЫ составляют до 50—60 % сухой массы мембраны
Функциональные

Транспорт веществ через мембрану
Прямой транспорт предполагает диффузию через мембрану гидрофобных
Транспорт веществ через мембрану
Прямой транспорт предполагает диффузию через мембрану гидрофобных

Облегченная диффузия с участием каналов.
Облегченная диффузия подчиняется законам диффузии. Ее
Облегченная диффузия с участием каналов.
Облегченная диффузия подчиняется законам диффузии. Ее
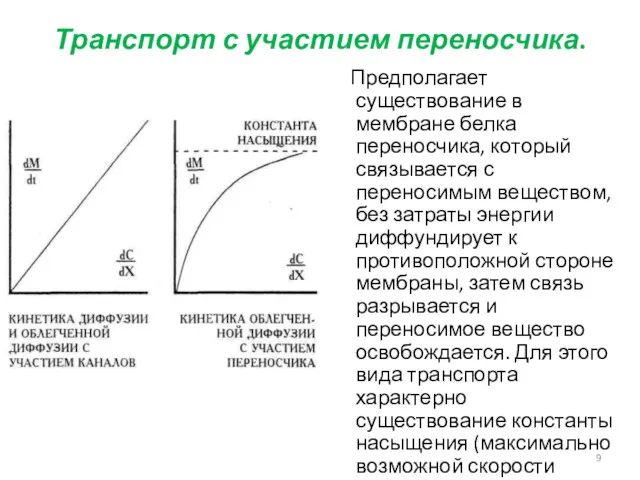
Транспорт с участием переносчика.
Предполагает существование в мембране белка переносчика, который
Транспорт с участием переносчика.
Предполагает существование в мембране белка переносчика, который
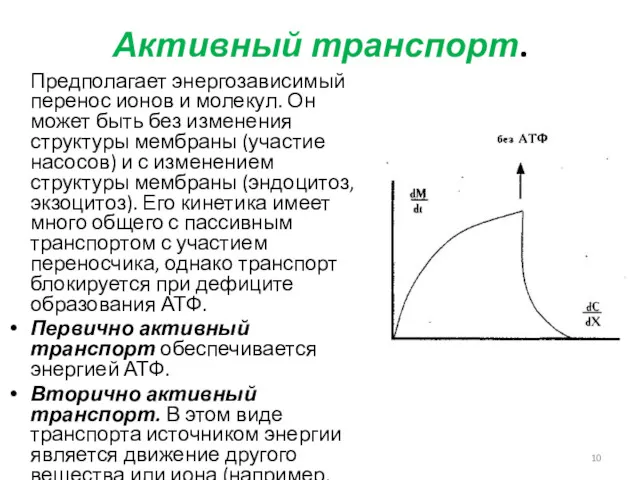
Активный транспорт.
Предполагает энергозависимый перенос ионов и молекул. Он может быть
Активный транспорт.
Предполагает энергозависимый перенос ионов и молекул. Он может быть

ИТОГО
Биопотенциалы чаще создаются в результате простой диффузии ионов через мемебрану
Диффузия каждого
ИТОГО
Биопотенциалы чаще создаются в результате простой диффузии ионов через мемебрану
Диффузия каждого

Количественные показатели :равновестный потенциал и электрохимический
Равновестный потенциал для конкретного иона-мембранный
Количественные показатели :равновестный потенциал и электрохимический
Равновестный потенциал для конкретного иона-мембранный

Общая характеристика возбудимых тканей
Свойства возбудимых тканей:
возбудимость
проводимость
сократимость
секреторной
Общая характеристика возбудимых тканей
Свойства возбудимых тканей:
возбудимость
проводимость
сократимость
секреторной
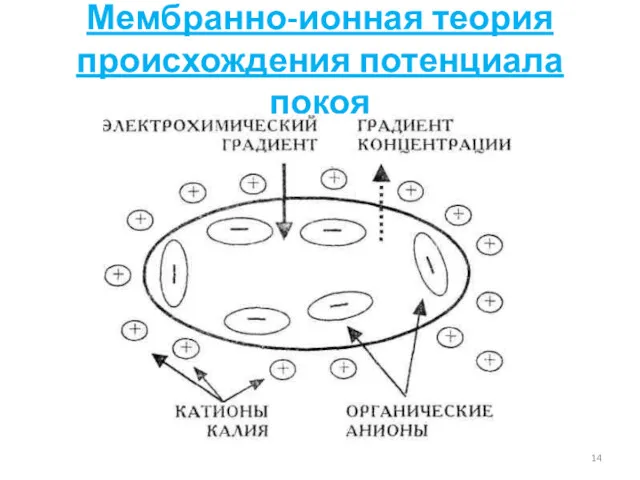
Мембранно-ионная теория происхождения потенциала покоя
Мембранно-ионная теория происхождения потенциала покоя

Физиологический смысл ПП
Электрохимический градиент для К невелик:
концентрационный и электрический
Физиологический смысл ПП
Электрохимический градиент для К невелик:
концентрационный и электрический

Уравнение Нернста
где Е калия - равновесный калиевый потенциал,
R
Уравнение Нернста
где Е калия - равновесный калиевый потенциал,
R

В процессе поляризации мембраны клетка теряет катионы калия и получает ионы
В процессе поляризации мембраны клетка теряет катионы калия и получает ионы

Значит, потенциал покоя:
Создается неравновесным распределением калия, натрия, хлора и кальция внутри
Значит, потенциал покоя:
Создается неравновесным распределением калия, натрия, хлора и кальция внутри

Мембранно-ионная теория происхождения потенциала действия
Потенциал действия, или импульсное возбуждение, возникает в
Мембранно-ионная теория происхождения потенциала действия
Потенциал действия, или импульсное возбуждение, возникает в
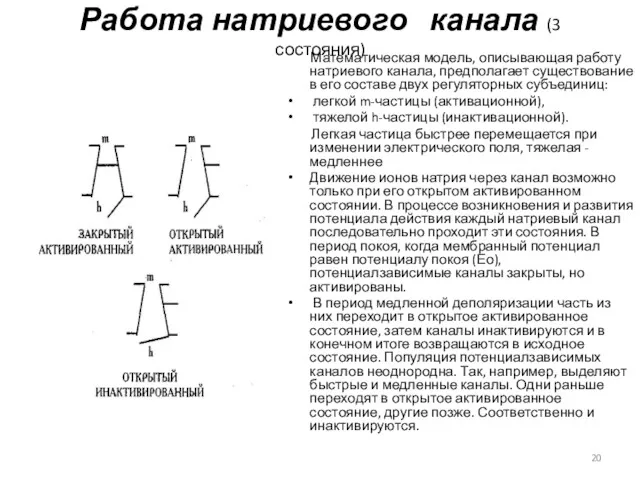
Работа натриевого канала (3 состояния)
Математическая модель, описывающая работу натриевого канала,
Работа натриевого канала (3 состояния)
Математическая модель, описывающая работу натриевого канала,

Работа калиевого канала
Математическая модель, описывающая работу калиевого канала, предполагает в структуре
Работа калиевого канала
Математическая модель, описывающая работу калиевого канала, предполагает в структуре

Потенциал действия
Фазы потенциала действия (ПД):
медленной деполяризации (1),
быстрой деполяризации
Потенциал действия
Фазы потенциала действия (ПД):
медленной деполяризации (1),
быстрой деполяризации

Значит, потенциал действия:
Возникает в ответ на пороговые и сверхпороговые раздражители;
Обусловлен увеличением
Возникает в ответ на пороговые и сверхпороговые раздражители;
Обусловлен увеличением

Локальный ответ.
Возникает в ответ на раздражители, составляющие не менее 60—80%
Локальный ответ.
Возникает в ответ на раздражители, составляющие не менее 60—80%
 Биологическое и социальное в человеке. Обществознание. 6 класс. 1 урок
Биологическое и социальное в человеке. Обществознание. 6 класс. 1 урок Геном человека. Мультифакториальные болезни
Геном человека. Мультифакториальные болезни Различия в строении клеток эукариот и прокариот
Различия в строении клеток эукариот и прокариот Розмноження птахів
Розмноження птахів Разбор заданий ВПР 5 класс
Разбор заданий ВПР 5 класс Урок-практикум Опорно-двигательная система
Урок-практикум Опорно-двигательная система Масти лошадей
Масти лошадей Нәруыздардың құрамы бойынша жіктеу. Денатурация
Нәруыздардың құрамы бойынша жіктеу. Денатурация Химический состав клетки. Опыты
Химический состав клетки. Опыты Белый медведь
Белый медведь Введение в гистологию
Введение в гистологию Селекция растений
Селекция растений Рослинна клітина
Рослинна клітина кожа и ее производные
кожа и ее производные Выравнивание генетических последовательностей представителей семейства dipsacaceae juss
Выравнивание генетических последовательностей представителей семейства dipsacaceae juss Дикие и домашние животные
Дикие и домашние животные Учебно-исследовательская работа Как влияет тканевый сок суккулентов на развитие семян?
Учебно-исследовательская работа Как влияет тканевый сок суккулентов на развитие семян? Презентация к уроку биологии по теме: Рыбы- водные животные
Презентация к уроку биологии по теме: Рыбы- водные животные Динозавры. (Окружающий мир. 1 класс)
Динозавры. (Окружающий мир. 1 класс) Як вчаться пташенята
Як вчаться пташенята Структура ДНК. Доказательства роли ДНК в передаче наследственной информации
Структура ДНК. Доказательства роли ДНК в передаче наследственной информации Функции желез внутренней секреции
Функции желез внутренней секреции Протопласты растительных клеток как объекты биологического конструирования
Протопласты растительных клеток как объекты биологического конструирования Государство Австралия. (2 класс)
Государство Австралия. (2 класс) Репликация ДНК. (Лекция 3)
Репликация ДНК. (Лекция 3) Дикие и домашние животные
Дикие и домашние животные Клеточные и неклеточные формы жизни
Клеточные и неклеточные формы жизни Отдел Охрофита. Лекция 5-6
Отдел Охрофита. Лекция 5-6