- Структура ДНК. Доказательства роли ДНК в передаче наследственной информации

Содержание
- 2. История открытия и исследования нуклеиновых кислот. 2
- 3. 1869г.: Фридерих Мишер выделенное из ядер лейкоцитов вещество назвал нуклеином 1891г.: Альбрехт Коссель в составе нуклеина
- 4. 1) количество пуриновых оснований = количеству пиримидиновых азотистых оснований; 2) содержание в клетке А = Т;
- 5. 1950г.: Морис Уилкинс и Розалинда Франклин на поперечном срезе ДНК с помощью рентгеноструктурного анализа получают интересную
- 6. ДНК состоит из 2 цепей; цепь спирально закручена, ее диаметр = 2 нм; цепь состоит из
- 7. 1953г.: Джеймс Уотсон и Френсис Крик пришли к выводу, что нити ДНК антипараллельны друг другу История
- 8. Доказательство роли ДНК в передаче наследственной информации. ВОПРОС № 1 Опыты Гриффитса, Эвери, Мак-Леода и Мак-Карти.
- 9. Опыты Фредерика Гриффитса В 1928г. Ф. Гриффитс обнаружил у пневмококков (Streptococcus pneumonia) явление трансформации. 9
- 10. S – тип: пневмококки, окруженные капсулой, образуют крупные гладкие колонии (от англ. smooth — гладкий) ПАТОГЕННЫ,
- 11. Схема опытов Ф. Гриффитса Непатогенный штамм Патогенный штамм Патогенный штамм после нагревания Микст - вариант 11
- 12. Вывод опыта Ф. Гриффитса: Гриффитс выявил существование некоего “трансформирующего начала”, превращающего клетки пневмококков типа IIR в
- 13. Опыты О. Эвери, К. Мак-Леода и М. Мак-Карти В 1944г. К. Мак-Леод и О. Эвери К.
- 14. Схема опытов О. Эвери, К. Мак-Леода и М. Мак-Карти Препараты ДНК из пневмококков типа IIIS делили
- 15. Схема опытов О. Эвери, К. Мак-Леода и М. Мак-Карти 15
- 16. Вывод опытов О. Эвери, К. Мак-Леода и М. Мак-Карти: Только обработка ДНК-азой полностью снимала трансформирующую активность
- 17. Доказательство роли ДНК в передаче наследственной информации. ВОПРОС № 2 Опыты Херши и Чейз. 17
- 18. Опыты А. Херши и М. Чейз В 1952г. А. Херши и М. Чейз в качестве объекта
- 19. Схема опытов А. Херши и М. Чейз Белок капсида фага Т2 помечали радиоактивным изотопом серы S35,
- 20. Вывод опытов А. Херши и М. Чейз: Во время инфекции в клетку проникает преимущественно фаговая ДНК
- 21. Структура нуклеиновых кислот. Нуклеотиды, их разновидности. ВОПРОС № 3 21
- 22. Нуклеиновые кислоты представляют собой макромолекулы, образованные повторяющимися структурами- нуклеотидами. 22
- 23. 23 Состав нуклеотида: циклическое азотсодержащее соединение, называемое основанием; сахар пентоза, включающий пять атомов углерода; остаток фосфорной
- 24. Известны пять главных азотистых основания : 24
- 25. Сахара, входящие в состав НК : В составе РНК В составе ДНК 25
- 26. Молекула РНК Молекула РНК состоит из одной цепи, в которой последовательно чередуются четыре возможных нуклеотида. 26
- 27. Первичная структура ДНК - нуклеотиды соединяются друг с другом 3’,5’-фосфодиэфирной связью. Фосфат связывает 3’-ОН группу одного
- 28. Вторичная структура ДНК – это Двойная спираль, состоящая из двух полинуклеотидных цепей Цепи комплементарны, антипараллельны и
- 29. Связи, стабилизирующие вторичную структуру ДНК : Водородные связи - между комплементарными азотистыми основаниями Стэкинг-взаимодействия - между
- 30. Пространственная конфигурация молекулы ДНК. Модель Уотсона и Крика. B и Z формы ДНК. ВОПРОС № 4
- 31. Пространственные конфигурации молекулы ДНК 31
- 32. Способы репликации ДНК: консервативный, полуконсервативный, дисперсионный. ВОПРОС № 5 Опыты М. Мезельсона и Ф. Сталя. 32
- 33. Репликация ДНК Репликация (от лат. replicatio — возобновление) — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на
- 34. Консервативный способ репликации ДНК Исходная ДНК остается неизменной во время всего процесса репликации и дочерние ДНК
- 35. Дисперсионный способ репликации ДНК Дробление молекул ДНК, в результате которого каждая отдельная цепь новых дочерних молекул
- 36. Полуконсервативный способ репликации ДНК Способ репликации двухцепочечной молекулы ДНК, при котором исходная молекула разделяется на две
- 37. Эксперименты М. Мезельсона и Ф. Сталя В 1958г. М. Мезельсон и Ф. Сталь с помощью метода
- 38. Схема опытов М. Мезельсона и Ф. Сталя 38
- 39. Вывод из опытов М. Мезельсона и Ф. Сталя: Вся ДНК, выделенная из клеток, выращенных в течение
- 40. Направление репликации ДНК. Образование репликативной вилки. Точка ori. ВОПРОС № 6 40
- 41. Направление репликации ДНК В 1963г. Дж. Кэрнс, используя метод авторадиографии, визуализировал процесс репликации ДНК у бактерий.
- 42. Точка начала репликации - ori (от англ. origin- начало). Точка ori Репликон – расстояние между двумя
- 43. Образование репликативной вилки связано с раскручиванием дуплекса ДНК и локальным разделением ее цепей. Образование репликативной вилки
- 44. Ферменты репликации. Инициация репликации. Факторы инициации. ВОПРОС № 7 44
- 45. Ферменты, обеспечивающие репликацию ДНК 45
- 46. Топоизомераза (ДНК-гираза) находит точку начала репликации (ori), гидролизует одну фосфодиэфирную связь и дает возможность компонентам репликативной
- 47. ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным связям между комплементарными нуклеотидами; ДНК-полимераза α
- 48. Элонгация репликации. ДНК-топоизомераза, ДНК-затравка, ДНК-полимераза. ВОПРОС № 8 48
- 49. ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей нити, а ДНК-полимеразы α и ε -
- 51. Скачать презентацию

История открытия и исследования нуклеиновых кислот.
2
История открытия и исследования нуклеиновых кислот.
2

1869г.: Фридерих Мишер выделенное из ядер лейкоцитов вещество назвал нуклеином
1891г.:
1869г.: Фридерих Мишер выделенное из ядер лейкоцитов вещество назвал нуклеином
1891г.:

1) количество пуриновых оснований = количеству пиримидиновых азотистых оснований;
2) содержание в
2) содержание в

1950г.: Морис Уилкинс и Розалинда Франклин на поперечном срезе ДНК с
1950г.: Морис Уилкинс и Розалинда Франклин на поперечном срезе ДНК с

ДНК состоит из 2 цепей;
цепь спирально закручена, ее диаметр =
ДНК состоит из 2 цепей;
цепь спирально закручена, ее диаметр =

1953г.: Джеймс Уотсон и Френсис Крик пришли к выводу, что нити
1953г.: Джеймс Уотсон и Френсис Крик пришли к выводу, что нити

Доказательство роли ДНК в передаче наследственной информации.
ВОПРОС № 1
Опыты
Доказательство роли ДНК в передаче наследственной информации.
ВОПРОС № 1
Опыты

Опыты Фредерика Гриффитса
В 1928г. Ф. Гриффитс обнаружил у пневмококков (Streptococcus pneumonia)
Опыты Фредерика Гриффитса
В 1928г. Ф. Гриффитс обнаружил у пневмококков (Streptococcus pneumonia)

S – тип:
пневмококки, окруженные капсулой, образуют крупные гладкие колонии (от
S – тип:
пневмококки, окруженные капсулой, образуют крупные гладкие колонии (от
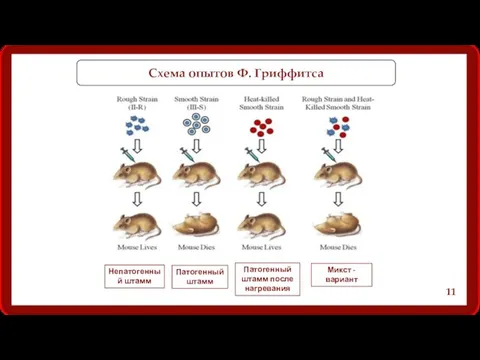
Схема опытов Ф. Гриффитса
Непатогенный штамм
Патогенный штамм
Патогенный штамм после нагревания
Микст - вариант
11
Схема опытов Ф. Гриффитса
Непатогенный штамм
Патогенный штамм
Патогенный штамм после нагревания
Микст - вариант
11

Вывод опыта Ф. Гриффитса:
Гриффитс выявил существование некоего “трансформирующего начала”,
Вывод опыта Ф. Гриффитса:
Гриффитс выявил существование некоего “трансформирующего начала”,

Опыты О. Эвери, К. Мак-Леода и М. Мак-Карти
В 1944г. К.
Опыты О. Эвери, К. Мак-Леода и М. Мак-Карти
В 1944г. К.

Схема опытов О. Эвери, К. Мак-Леода и М. Мак-Карти
Препараты ДНК
Схема опытов О. Эвери, К. Мак-Леода и М. Мак-Карти
Препараты ДНК
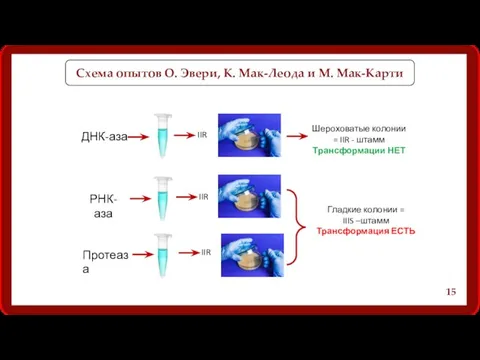
Схема опытов О. Эвери, К. Мак-Леода и М. Мак-Карти
15
Схема опытов О. Эвери, К. Мак-Леода и М. Мак-Карти
15

Вывод опытов О. Эвери, К. Мак-Леода и М. Мак-Карти:
Только
Вывод опытов О. Эвери, К. Мак-Леода и М. Мак-Карти:
Только

Доказательство роли ДНК в передаче наследственной информации.
ВОПРОС № 2
Опыты
Доказательство роли ДНК в передаче наследственной информации.
ВОПРОС № 2
Опыты

Опыты А. Херши и М. Чейз
В 1952г. А. Херши и М.
Опыты А. Херши и М. Чейз
В 1952г. А. Херши и М.

Схема опытов А. Херши и М. Чейз
Белок капсида фага Т2
Схема опытов А. Херши и М. Чейз
Белок капсида фага Т2

Вывод опытов А. Херши и М. Чейз:
Во время инфекции в
Вывод опытов А. Херши и М. Чейз:
Во время инфекции в

Структура нуклеиновых кислот. Нуклеотиды, их разновидности.
ВОПРОС № 3
21
Структура нуклеиновых кислот. Нуклеотиды, их разновидности.
ВОПРОС № 3
21
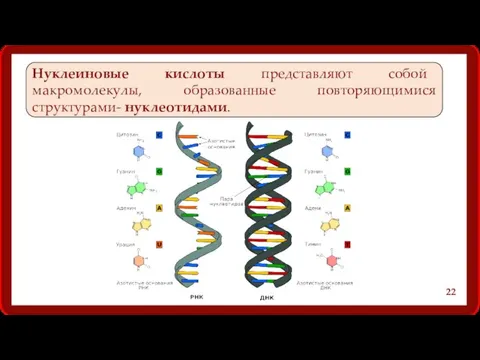
Нуклеиновые кислоты представляют собой макромолекулы, образованные повторяющимися структурами- нуклеотидами.
22
Нуклеиновые кислоты представляют собой макромолекулы, образованные повторяющимися структурами- нуклеотидами.
22
23
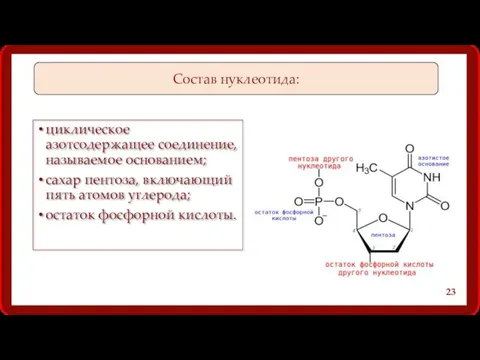
Состав нуклеотида:
циклическое азотсодержащее соединение, называемое основанием;
сахар пентоза, включающий пять атомов
23
Состав нуклеотида:
циклическое азотсодержащее соединение, называемое основанием;
сахар пентоза, включающий пять атомов
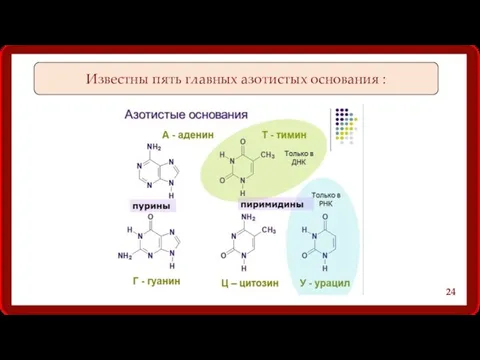
Известны пять главных азотистых основания :
24
Известны пять главных азотистых основания :
24
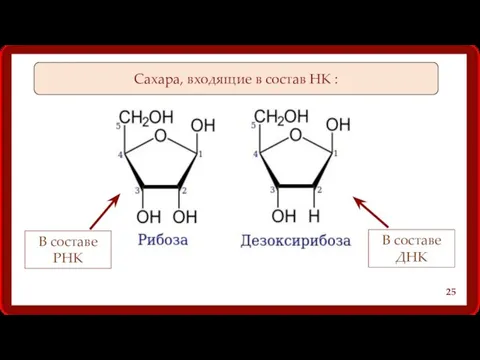
Сахара, входящие в состав НК :
В составе РНК
В составе
Сахара, входящие в состав НК :
В составе РНК
В составе

Молекула РНК
Молекула РНК состоит из одной цепи, в которой последовательно
Молекула РНК
Молекула РНК состоит из одной цепи, в которой последовательно

Первичная структура ДНК - нуклеотиды соединяются друг с другом 3’,5’-фосфодиэфирной связью.
Первичная структура ДНК - нуклеотиды соединяются друг с другом 3’,5’-фосфодиэфирной связью.

Вторичная структура ДНК – это
Двойная спираль, состоящая из двух полинуклеотидных
Вторичная структура ДНК – это
Двойная спираль, состоящая из двух полинуклеотидных
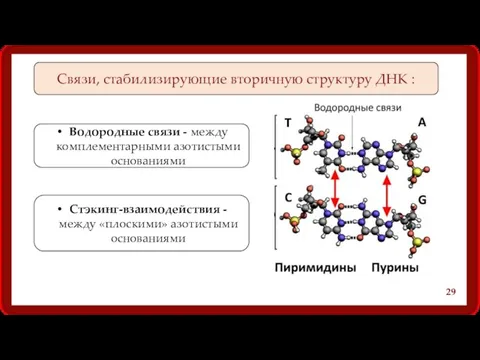
Связи, стабилизирующие вторичную структуру ДНК :
Водородные связи - между комплементарными
Связи, стабилизирующие вторичную структуру ДНК :
Водородные связи - между комплементарными

Пространственная конфигурация молекулы ДНК. Модель Уотсона и Крика. B и Z
Пространственная конфигурация молекулы ДНК. Модель Уотсона и Крика. B и Z

Пространственные конфигурации молекулы ДНК
31
Пространственные конфигурации молекулы ДНК
31

Способы репликации ДНК: консервативный, полуконсервативный, дисперсионный.
ВОПРОС № 5
Опыты М.
Способы репликации ДНК: консервативный, полуконсервативный, дисперсионный.
ВОПРОС № 5
Опыты М.
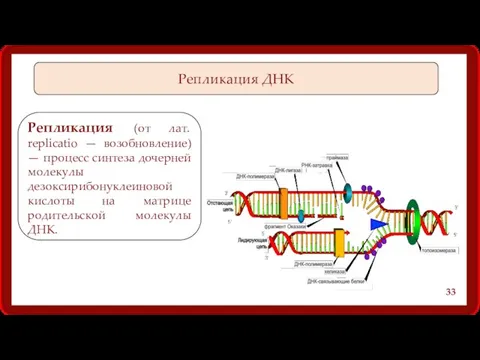
Репликация ДНК
Репликация (от лат. replicatio — возобновление) — процесс синтеза
Репликация ДНК
Репликация (от лат. replicatio — возобновление) — процесс синтеза
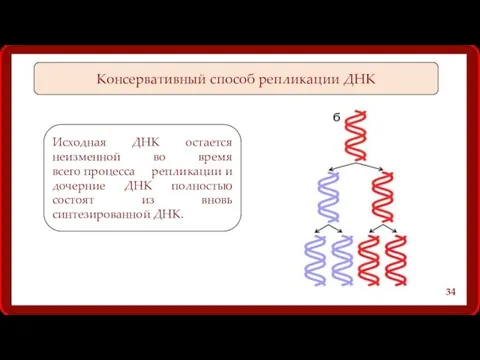
Консервативный способ репликации ДНК
Исходная ДНК остается неизменной во время всего процесса
Консервативный способ репликации ДНК
Исходная ДНК остается неизменной во время всего процесса
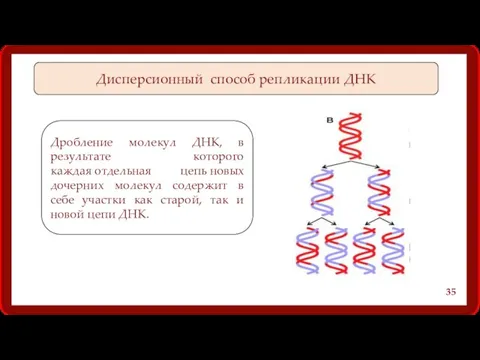
Дисперсионный способ репликации ДНК
Дробление молекул ДНК, в результате которого каждая отдельная
Дисперсионный способ репликации ДНК
Дробление молекул ДНК, в результате которого каждая отдельная

Полуконсервативный способ репликации ДНК
Способ репликации двухцепочечной молекулы ДНК, при котором
Полуконсервативный способ репликации ДНК
Способ репликации двухцепочечной молекулы ДНК, при котором

Эксперименты М. Мезельсона и Ф. Сталя
В 1958г. М. Мезельсон и
Эксперименты М. Мезельсона и Ф. Сталя
В 1958г. М. Мезельсон и

Схема опытов М. Мезельсона и Ф. Сталя
38
Схема опытов М. Мезельсона и Ф. Сталя
38

Вывод из опытов М. Мезельсона и Ф. Сталя:
Вся ДНК, выделенная
Вывод из опытов М. Мезельсона и Ф. Сталя:
Вся ДНК, выделенная

Направление репликации ДНК. Образование репликативной вилки. Точка ori.
ВОПРОС № 6
40
Направление репликации ДНК. Образование репликативной вилки. Точка ori.
ВОПРОС № 6
40

Направление репликации ДНК
В 1963г. Дж. Кэрнс, используя метод авторадиографии, визуализировал
Направление репликации ДНК
В 1963г. Дж. Кэрнс, используя метод авторадиографии, визуализировал
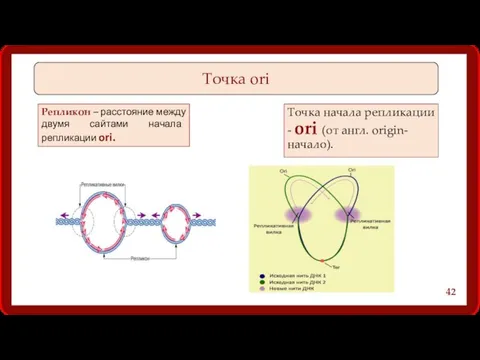
Точка начала репликации - ori (от англ. origin- начало).
Точка ori
Репликон –
Точка начала репликации - ori (от англ. origin- начало).
Точка ori
Репликон –
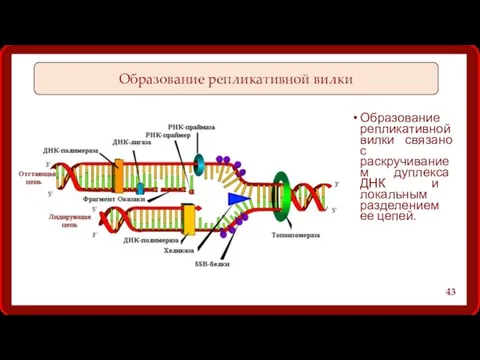
Образование репликативной вилки связано с раскручиванием дуплекса ДНК и локальным разделением
Образование репликативной вилки связано с раскручиванием дуплекса ДНК и локальным разделением

Ферменты репликации. Инициация репликации. Факторы инициации.
ВОПРОС № 7
44
Ферменты репликации. Инициация репликации. Факторы инициации.
ВОПРОС № 7
44

Ферменты, обеспечивающие репликацию ДНК
45
Ферменты, обеспечивающие репликацию ДНК
45

Топоизомераза (ДНК-гираза) находит точку начала репликации (ori), гидролизует одну фосфодиэфирную связь
Топоизомераза (ДНК-гираза) находит точку начала репликации (ori), гидролизует одну фосфодиэфирную связь

ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным связям
ДНК-связывающие белки (SSB-белки) стабилизируют репликативную вилку, не давая восстанавливаться водородным связям

Элонгация репликации.
ДНК-топоизомераза, ДНК-затравка, ДНК-полимераза.
ВОПРОС № 8
48
Элонгация репликации.
ДНК-топоизомераза, ДНК-затравка, ДНК-полимераза.
ВОПРОС № 8
48

ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей нити, а
ДНК-полимераза δ продолжает удлинять нить из дезоксирибонуклеотидов на лидирующей нити, а
 Как зимой помочь птицам. (Окружающий мир, 1 класс)
Как зимой помочь птицам. (Окружающий мир, 1 класс) Итоговая контрольная работа. 6 класс
Итоговая контрольная работа. 6 класс Ферменти: класифікація, структурно-функціональна організація
Ферменти: класифікація, структурно-функціональна організація Neisseria. General Characteristics of Neisseria spp
Neisseria. General Characteristics of Neisseria spp Спинной мозг
Спинной мозг Дыхание растений
Дыхание растений Биоинженерия. Полимеразная цепная реакция
Биоинженерия. Полимеразная цепная реакция Грегор Мендель (1822 - 1884 г.г.). Законы, задачи
Грегор Мендель (1822 - 1884 г.г.). Законы, задачи Пищеварение в кишечнике.Всасывание питательных веществ.
Пищеварение в кишечнике.Всасывание питательных веществ. Обмен простых белков. Образование биологически активных аминов
Обмен простых белков. Образование биологически активных аминов Домашняя бабочка
Домашняя бабочка Биология 8 класс. Иммунитет
Биология 8 класс. Иммунитет Самый большой отряд насекомых
Самый большой отряд насекомых Внутреннее строение рыбы
Внутреннее строение рыбы Лазовский(Судзухинский) заповедник
Лазовский(Судзухинский) заповедник Вегетативные органы растений. Побег
Вегетативные органы растений. Побег Движение. 6 класс
Движение. 6 класс Свойства живого
Свойства живого Опорно-двигательный аппарат
Опорно-двигательный аппарат Размножение и развитие растений
Размножение и развитие растений Животные (фотографии)
Животные (фотографии) Репетитор ЕГЭ. Человек
Репетитор ЕГЭ. Человек Знакомство с внешним строением цветкового растения
Знакомство с внешним строением цветкового растения Немного о дельфинах
Немного о дельфинах Скелет позвоночных
Скелет позвоночных Воспроизводительная система рыб
Воспроизводительная система рыб Чим бактерії корисні людині. Міні-проект
Чим бактерії корисні людині. Міні-проект Тема 12
Тема 12