- Фотосинтез

Содержание
- 2. Михаил Семёнович Цвет Другие каротиноиды Пигменты зелёного листа
- 3. Хлорофилл: двуликий Янус Red-Ox реакций Хлорофиллов >10: Хл. а, b, c1, с2, d, e; Б-хл. a,
- 4. Многообразие хлорофиллов Хлорофиллы (Chl) Бактериохлорофиллы (BChl) Водоросли
- 5. Основные структурные особенности молекулы хлорофилла Конъюгированная система двойных связей: основная 18-членная π-система + дополнителные в I,
- 6. δ-амннолевулиновая кислота – предшественник гема и хлорофиллов.
- 7. Биосинтез порфиринов у растений происходит в пластидах цитохромы, каталаза, пероксидазы, etc. фитохромы (фоторецепторы) хлорофиллы
- 8. Спектры поглощения хлорофиллов Почему видимый свет ? 1. Оптимум энергии: 1 – 3 эв. 2. Максимальная
- 9. Энергетические уровни хлорофилла
- 10. Белковое окружение изменяет спектр поглощения хлорофилла
- 11. Первичные процессы фотосинтеза – временные интервалы различаются на 6 -7 порядков Поглощение фотона и перенос энергии
- 12. Простейшая схема фотосинтеза — пурпурные бактерии. За счет работы b6c1-комплекса протоны «перекачиваются» через мембрану, и за
- 13. Вторая простейшая схема фотосинтеза - серные зеленые бактерии. Восстановленные соединения образуются! FeS кластеры (840 нм) (MQ)
- 14. Основные требования к донору электронов: Ео’ меньше Ео’ хл+ (т.е. может отдавать е- на хл+) Продукты
- 15. Как решить проблемы водички в качестве донора электрона? Энергия квантов видимого света. 400нм (синий свет) —
- 16. Гениальное решение: соединить две фотосистемы! Фотосистема II Фотосистема I
- 17. Z-схема: оптимальное сочетание фотосистем позволяет «втиснуть» между ними еще и b6f-комплекс Фотосистема II Фотосистема I цитохромный
- 18. Кофакторы ЭТЦ фотосинтеза: знакомые все лица... 2Fe-2S и 4Fe-4S-белки, хиноны (пластохиноны и филлохинон), цитохромы Убихинон (UQ)
- 19. Z-схема фотосинтеза Расположение кофакторов в координатах ОВ потенциала И «механический» аналог - для наглядности...
- 20. Z-схема фотосинтеза: RedOx потенциалы компонентов ЭТЦ хлоропластов
- 21. Два типа реакционных центров: феофитин-хиноновый и железо-серный Ну очень положительный потенциал…
- 22. Организация фотосинтетического аппарата Тилакоиды образовались по-видимому, за счет инвагинации внутренней мембраны с последующим «отшнуровы-ванием». Таким образом,
- 23. Организация фотосинтетического аппарата весьма похожа на ЭТЦ дыхания
- 24. Организация фотосинтетического аппарата, «реальная картинка». Строма Люмен
- 25. Фотосистема II: схема реакционного центра (РЦ) 25 белков: D1 + D2 – Гетеродимер, на нем: Р680
- 26. Кинетика работы водоокисляющего комплекса номер вспышки выделение О2 (отн.ед.) выделение O2 при освещении вспышками света
- 27. Марганцевый кластер системы фотоокисления воды Тирозин-161 Гистидин-190 Глут.к-та-189 Только один из ионов марганца, а именно Mn4,
- 28. Не все так просто и не все так ясно в системе водоокисления… В организации системы, по-видимому,
- 29. Фотосистема II в «реальном виде»
- 30. В6f-комплекс: схема. Два такта работы Q-цикла. Как в митохондриях… PQH2/PQ – восстановленный/окисленный пластохинон RFeS – белок
- 31. В6f-комплекс (димер) в «реальном виде» Цитохромы-b6 – фиолетовые ленточки, Субъединицы IV - серые, Белок Риске -
- 32. Фотосистема I: схема реакционного центра (РЦ) ФСI – 13 белков: А + В – гетеродимер, на
- 33. Фотосистема I в «реальном виде» В отличие от бактериального РЦ, в ФСI скорее всего работают обе
- 34. ЭТЦ хлоропластов: образование NADPH и ATP Количественное соотношение компонентов в ЭТЦ
- 35. Другие варианты использования электронов с ферредоксина Восстановление азота и серы (минеральное питание) Регуляция активности ферментов, в
- 36. Антенны: дополнительные пигменты фикоэритрин фикоцианин аллофикоцианин Chl a Chl a Chl b Спектр солнечного света лютеин
- 37. Антенны. Фикобилисомы – светособирающий комплекс цианобактерий и красных водорослей пигмент λmax поглощения PE – фикоэритрины 570
- 38. Фикобилипротеиды. По набору пигментов в антенных комплексах можно заключить, что симбиоз фототрофов происходил неоднократно.. Цианобактерии Chla
- 39. Антенны. Светособирающие комплексы различных организмов А – хлоросома зеленых серных бактерий (Chlorobium) ССК расположен на цитоплазматической
- 40. Каротиноиды: каротины и ксантофиллы – тетратерпены (С40) Различия по концевым группам, содержанию кислорода, изомерии, числу двойных
- 41. Развлечения господ инженеров. Генных… Колонии E.coli, экспрессирующие гены биосинтеза соответствующих каротиноидов. 5 7 9 11 11
- 42. Схема мономера фотосистемы II с антенными комплексами СР43, СР47 – внутренние антенны ФСII CP24, CP26, CP29
- 43. Светособирающий комплекс LHC II (Lhcb 1+2+3) : мономер и тример Каждый белок содержит 7 молекул хлорофилла
- 44. Схема мономера и димера фотосистемы II с антенными комплексами мономер ФСII: РЦ, внутренние и внешние антенны
- 45. Светособирающие комплексы фотосистем I и II ССКII (тример)
- 46. Состав хлорофилл-белковых комплексов ФСI и ФСII Одна «фотосинтетическая единица» – т.е. одна ЭТЦ (от ФСII до
- 47. Гетерогенность тилакоидых мембран
- 48. Hosta ‘Piedmont Gold’ Hosta sieboldiana ‘Elegans’ Hosta ‘Golden Haze’ Фотовыцветание
- 49. Механизмы регулирования и защиты ФСII от фотодеструкции нециклический поток, регулирование мобильными антеннами; циклические потоки вокруг каждой
- 50. Регулирование поглощения света мобильными антеннами Р Р Р Р Р Состояние 1 Состояние 2 Состояние 1
- 51. Варианты электронного транспорта в ЭТЦ хлоропластов Варианты транспорта ē: Продукты: 0 – Нециклический транспорт ē (основной
- 52. Варианты электронного транспорта в ЭТЦ хлоропластов, чуть подробнее… 1 2 4 Варианты транспорта ē: 1 –
- 53. Циклический поток электронов вокруг фотосистемы II cytb559 – цитохром b559 – периферические субъединицы ФСII ChlZ/D –
- 54. Варианты работы ЭТЦ фотосинтеза: Z-схема Варианты: Циклические потоки ē у каждой из фотосистем Псевдоциклический транспорт электронов
- 55. Функции каротиноидов 1. Антенны (400 – 500 нм) 2. Структурная (организация ССК) 3. Фотопротекторная (виолаксантиновый цикл)
- 56. Виолоксантиновый цикл – основа «нефотохимического тушения» Энергия S1 ниже S1 хлорофилла Энергия S1 выше S1 хлорофилла
- 57. Фотозащита Виолоксантин-эпоксидаза, 43 kDa, тример рН 7 – неактивна, рН 5 – активна. Т.к. находится со
- 58. D1 белок – «камикадзе» растительной клетки Разборка ФС II: уходят белки OEC, снимаются атомы Mn, отсоединяются
- 59. Темновая фаза фотосинтеза – образование «основных фондов» из НАДФН и АТФ
- 61. Скачать презентацию
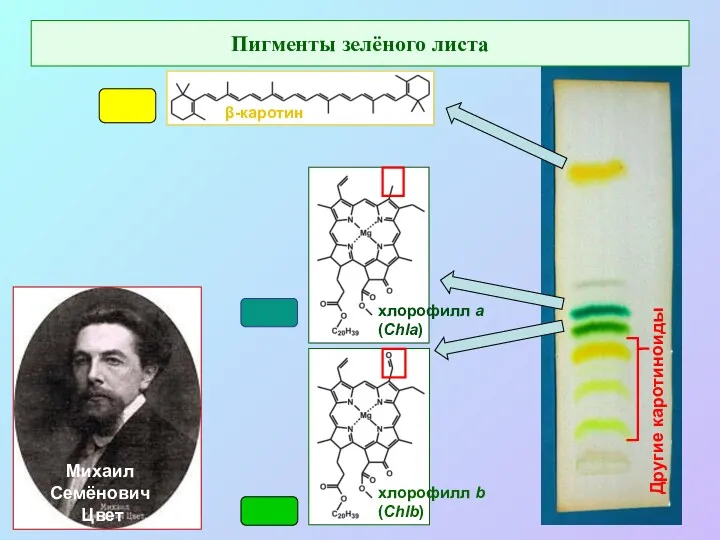
Михаил
Семёнович
Цвет
Другие каротиноиды
Пигменты зелёного листа
Михаил
Семёнович
Цвет
Другие каротиноиды
Пигменты зелёного листа
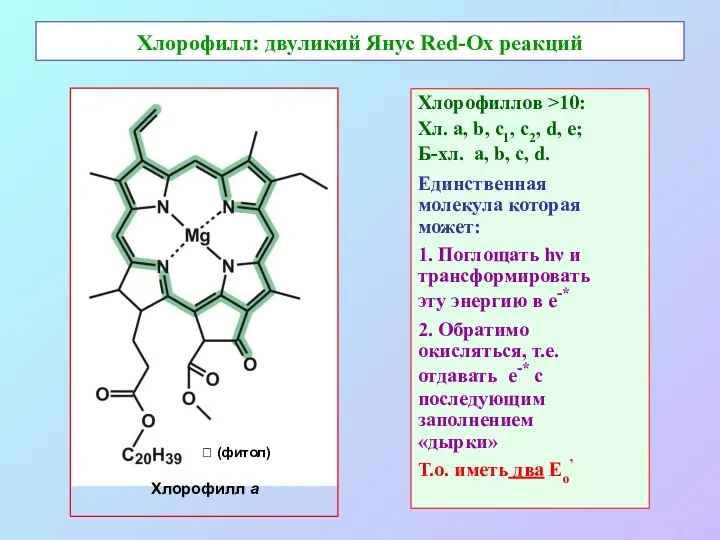
Хлорофилл: двуликий Янус Red-Ox реакций
Хлорофиллов >10:
Хл. а, b, c1, с2,
Хлорофилл: двуликий Янус Red-Ox реакций
Хлорофиллов >10:
Хл. а, b, c1, с2,
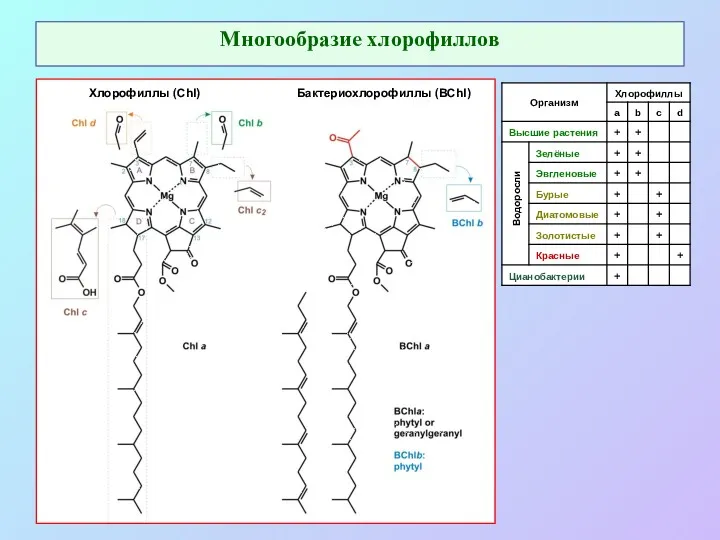
Многообразие хлорофиллов
Хлорофиллы (Chl) Бактериохлорофиллы (BChl)
Водоросли
Многообразие хлорофиллов
Хлорофиллы (Chl) Бактериохлорофиллы (BChl)
Водоросли

Основные структурные особенности молекулы хлорофилла
Конъюгированная система двойных связей: основная 18-членная π-система
Основные структурные особенности молекулы хлорофилла
Конъюгированная система двойных связей: основная 18-членная π-система
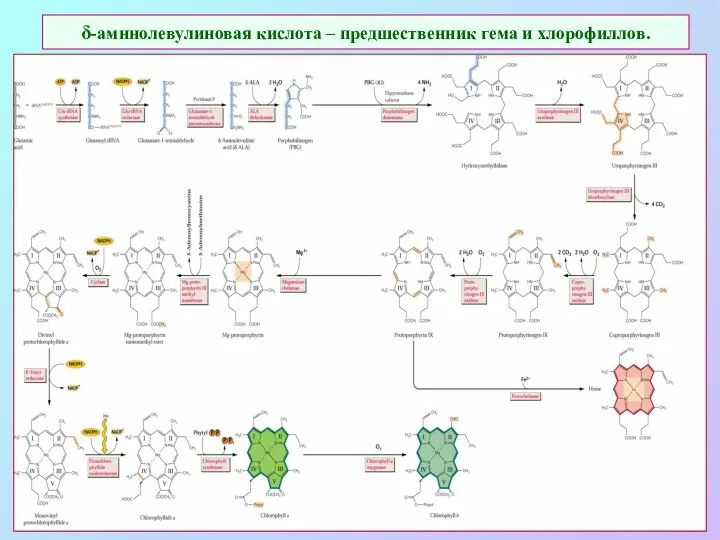
δ-амннолевулиновая кислота – предшественник гема и хлорофиллов.
δ-амннолевулиновая кислота – предшественник гема и хлорофиллов.
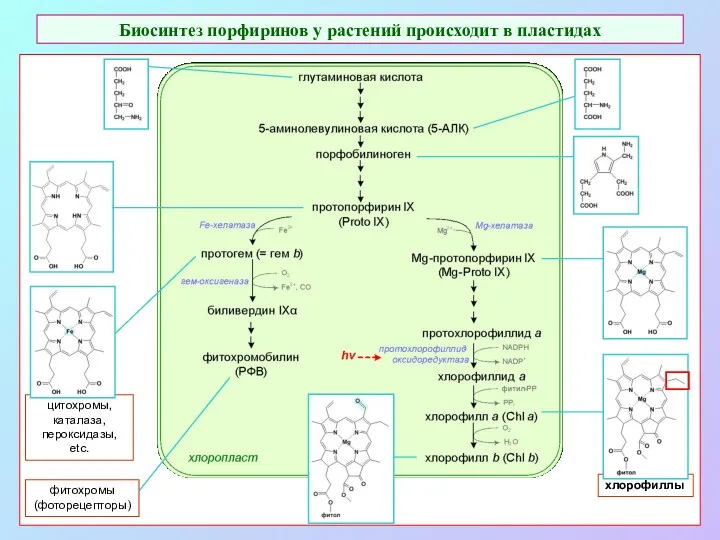
Биосинтез порфиринов у растений происходит в пластидах
цитохромы,
каталаза,
пероксидазы,
etc.
фитохромы
(фоторецепторы)
хлорофиллы
Биосинтез порфиринов у растений происходит в пластидах
цитохромы,
каталаза,
пероксидазы,
etc.
фитохромы
(фоторецепторы)
хлорофиллы
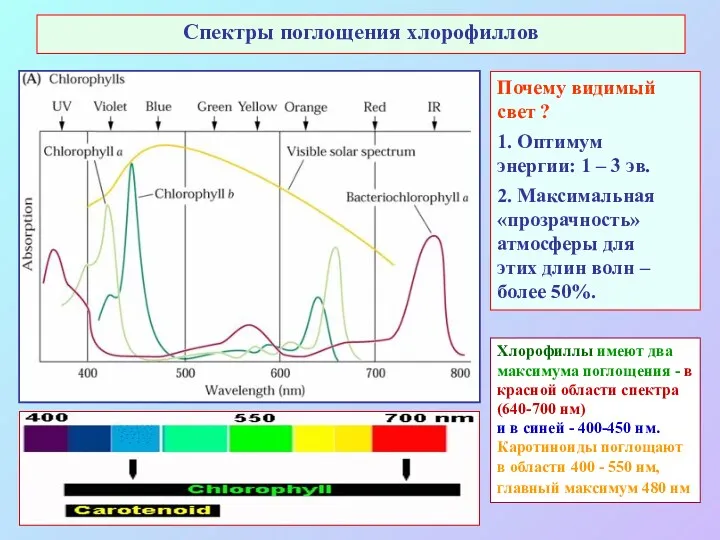
Спектры поглощения хлорофиллов
Почему видимый
свет ?
1. Оптимум
энергии: 1 – 3
Спектры поглощения хлорофиллов
Почему видимый
свет ?
1. Оптимум
энергии: 1 – 3
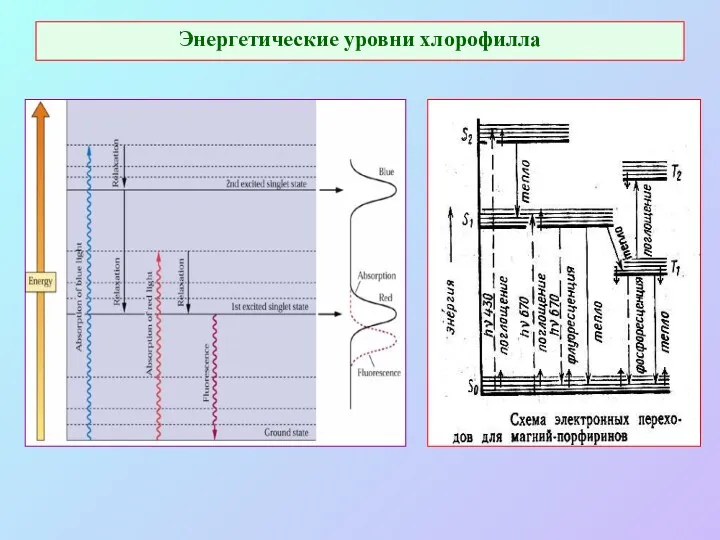
Энергетические уровни хлорофилла
Энергетические уровни хлорофилла
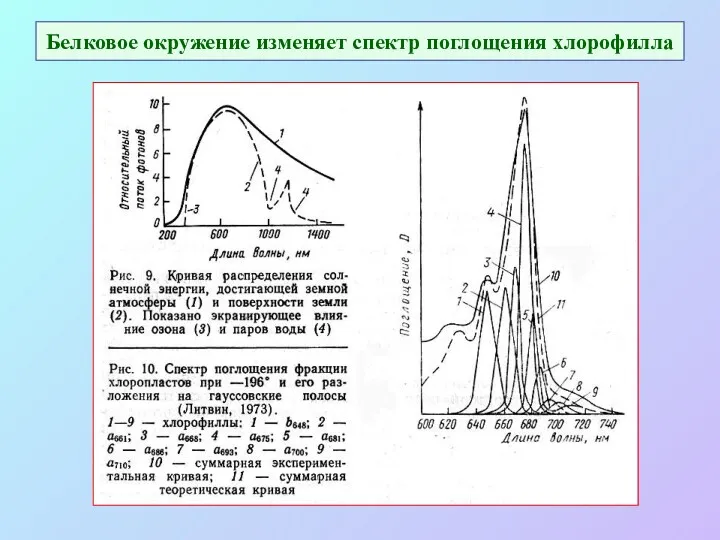
Белковое окружение изменяет спектр поглощения хлорофилла
Белковое окружение изменяет спектр поглощения хлорофилла
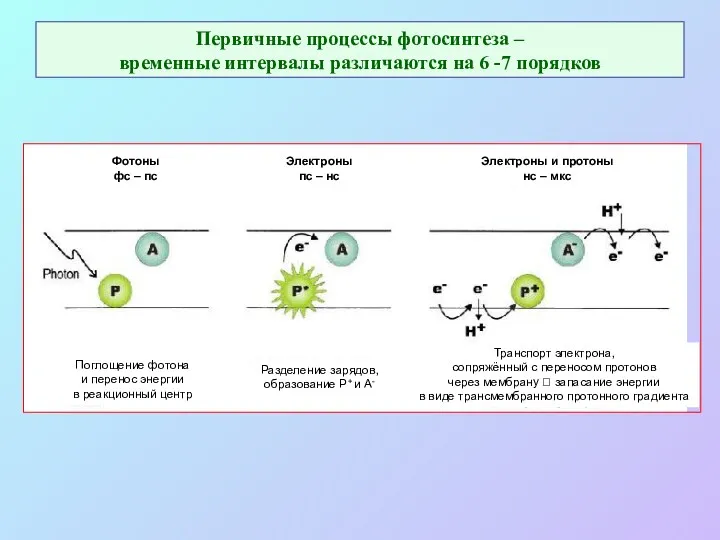
Первичные процессы фотосинтеза –
временные интервалы различаются на 6 -7 порядков
Поглощение
Первичные процессы фотосинтеза –
временные интервалы различаются на 6 -7 порядков
Поглощение
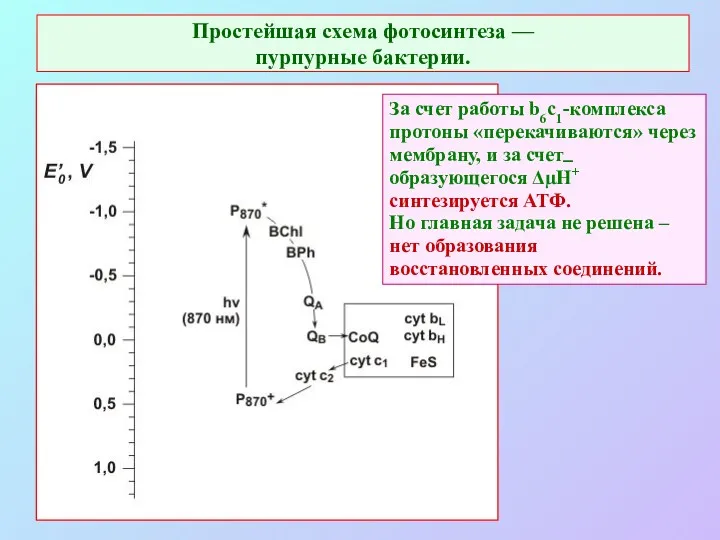
Простейшая схема фотосинтеза —
пурпурные бактерии.
За счет работы b6c1-комплекса протоны
Простейшая схема фотосинтеза —
пурпурные бактерии.
За счет работы b6c1-комплекса протоны
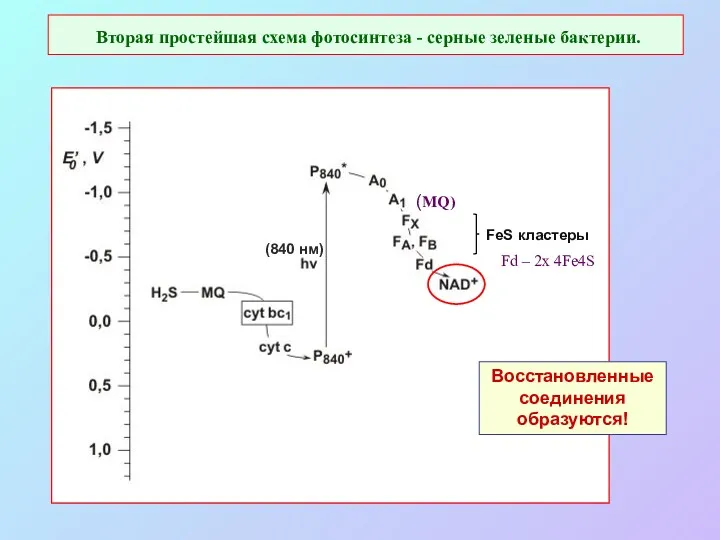
Вторая простейшая схема фотосинтеза - серные зеленые бактерии.
Восстановленные соединения
Вторая простейшая схема фотосинтеза - серные зеленые бактерии.
Восстановленные соединения

Основные требования к донору электронов:
Ео’ меньше Ео’ хл+ (т.е. может отдавать
Основные требования к донору электронов:
Ео’ меньше Ео’ хл+ (т.е. может отдавать
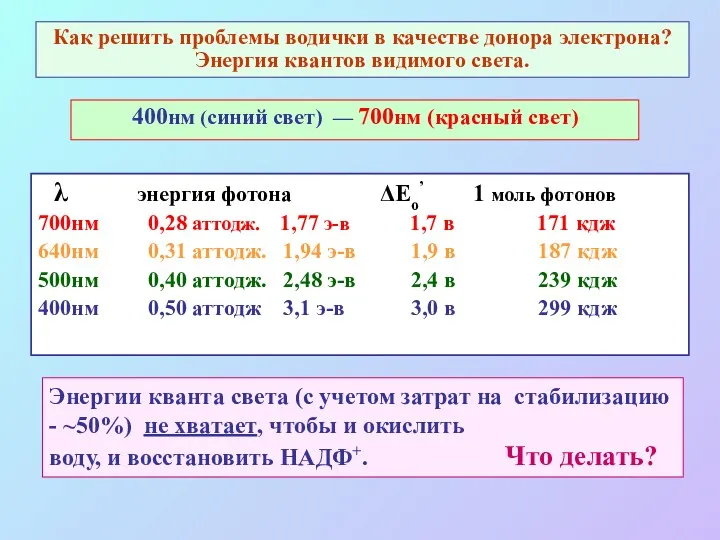
Как решить проблемы водички в качестве донора электрона? Энергия квантов видимого
Как решить проблемы водички в качестве донора электрона? Энергия квантов видимого
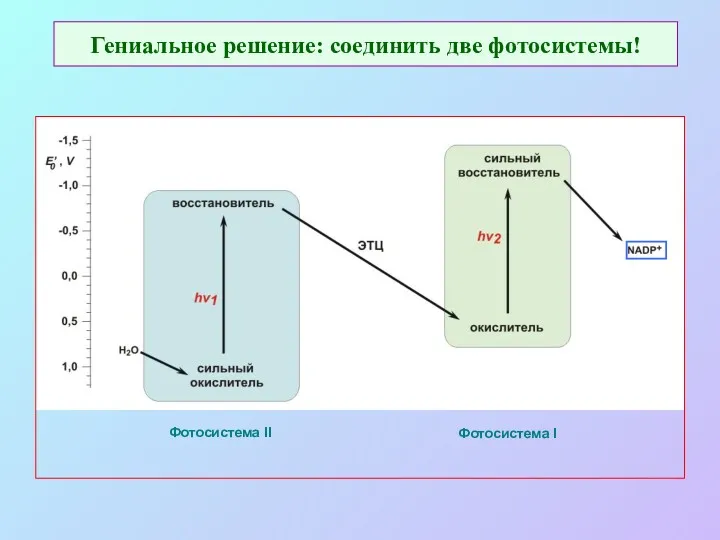
Гениальное решение: соединить две фотосистемы!
Фотосистема II
Фотосистема I
Гениальное решение: соединить две фотосистемы!
Фотосистема II
Фотосистема I
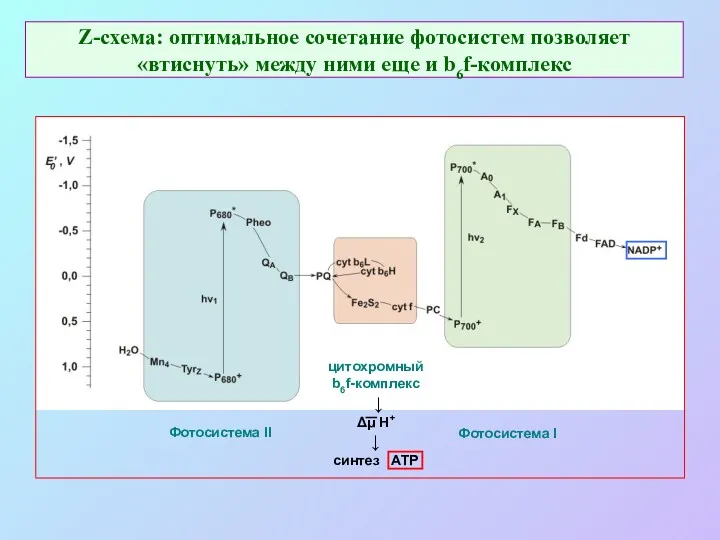
Z-схема: оптимальное сочетание фотосистем позволяет «втиснуть» между ними еще и b6f-комплекс
Z-схема: оптимальное сочетание фотосистем позволяет «втиснуть» между ними еще и b6f-комплекс
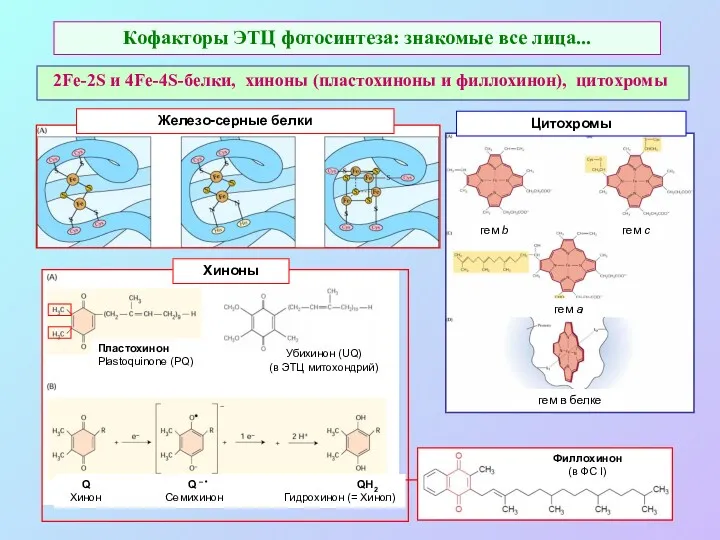
Кофакторы ЭТЦ фотосинтеза: знакомые все лица...
2Fe-2S и 4Fe-4S-белки, хиноны (пластохиноны
Кофакторы ЭТЦ фотосинтеза: знакомые все лица...
2Fe-2S и 4Fe-4S-белки, хиноны (пластохиноны
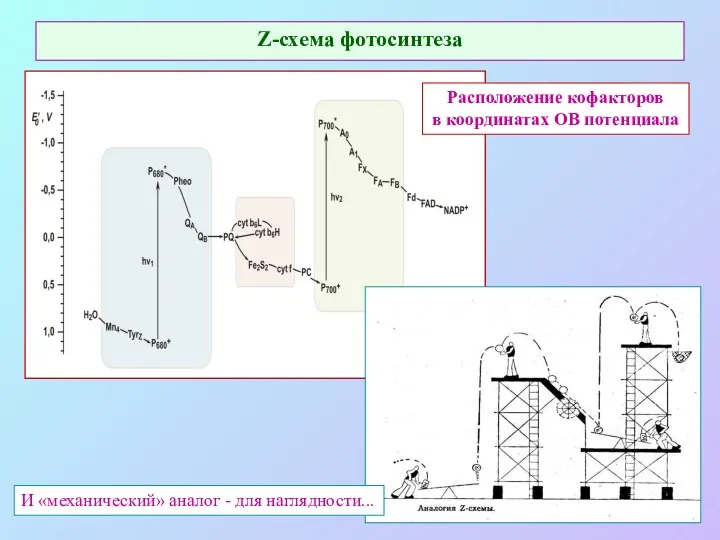
Z-схема фотосинтеза
Расположение кофакторов
в координатах ОВ потенциала
И «механический» аналог - для наглядности...
Z-схема фотосинтеза
Расположение кофакторов
в координатах ОВ потенциала
И «механический» аналог - для наглядности...
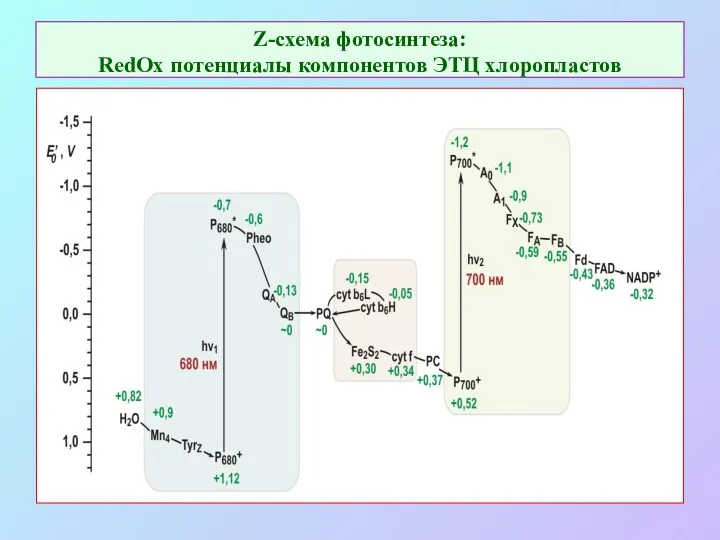
Z-схема фотосинтеза:
RedOx потенциалы компонентов ЭТЦ хлоропластов
Z-схема фотосинтеза:
RedOx потенциалы компонентов ЭТЦ хлоропластов
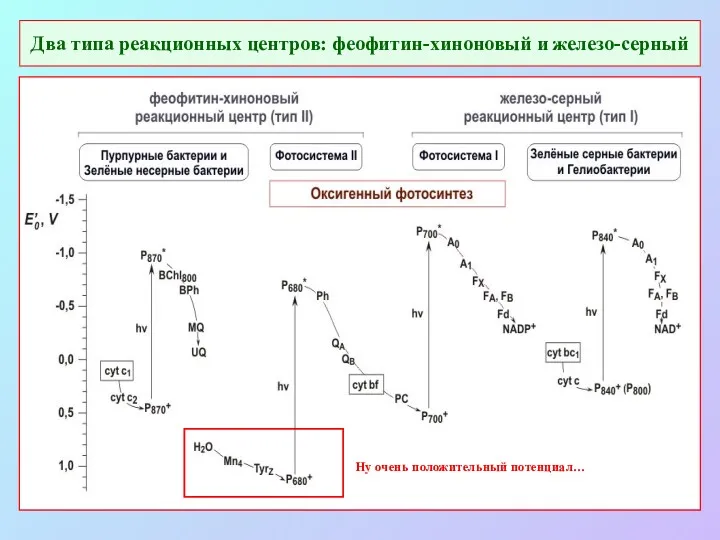
Два типа реакционных центров: феофитин-хиноновый и железо-серный
Ну очень положительный потенциал…
Два типа реакционных центров: феофитин-хиноновый и железо-серный
Ну очень положительный потенциал…
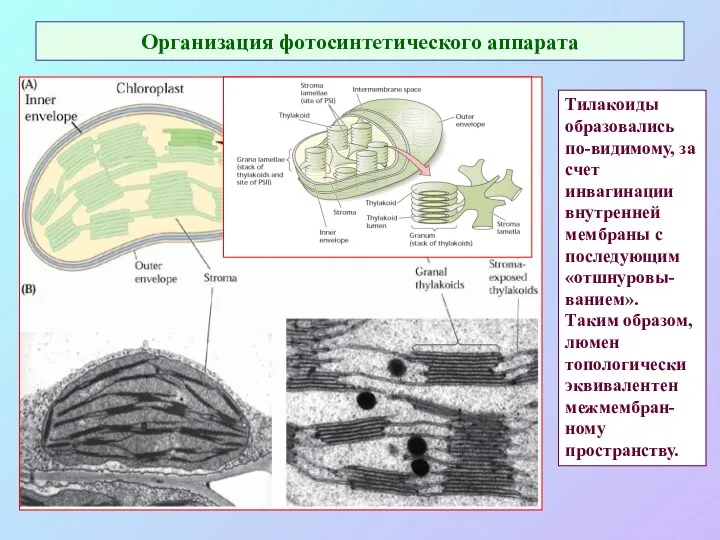
Организация фотосинтетического аппарата
Тилакоиды образовались по-видимому, за счет инвагинации внутренней мембраны с
Организация фотосинтетического аппарата
Тилакоиды образовались по-видимому, за счет инвагинации внутренней мембраны с
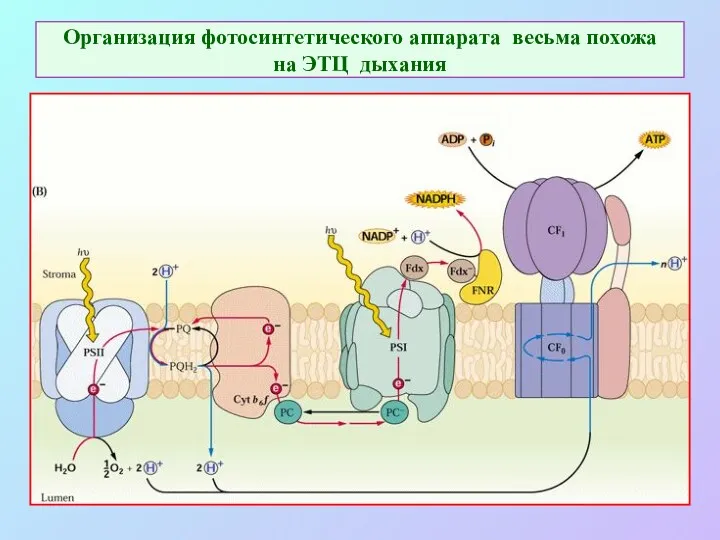
Организация фотосинтетического аппарата весьма похожа
на ЭТЦ дыхания
Организация фотосинтетического аппарата весьма похожа
на ЭТЦ дыхания

Организация фотосинтетического аппарата, «реальная картинка».
Строма
Люмен
Организация фотосинтетического аппарата, «реальная картинка».
Строма
Люмен
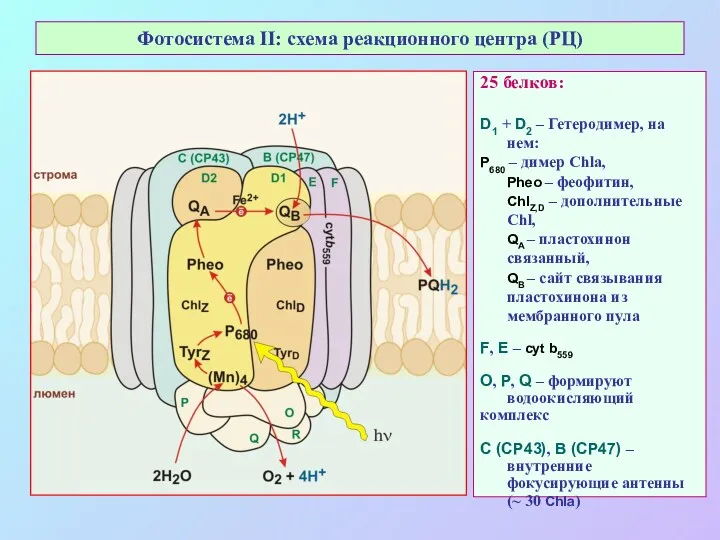
Фотосистема II: схема реакционного центра (РЦ)
25 белков:
D1 + D2 – Гетеродимер,
Фотосистема II: схема реакционного центра (РЦ)
25 белков:
D1 + D2 – Гетеродимер,
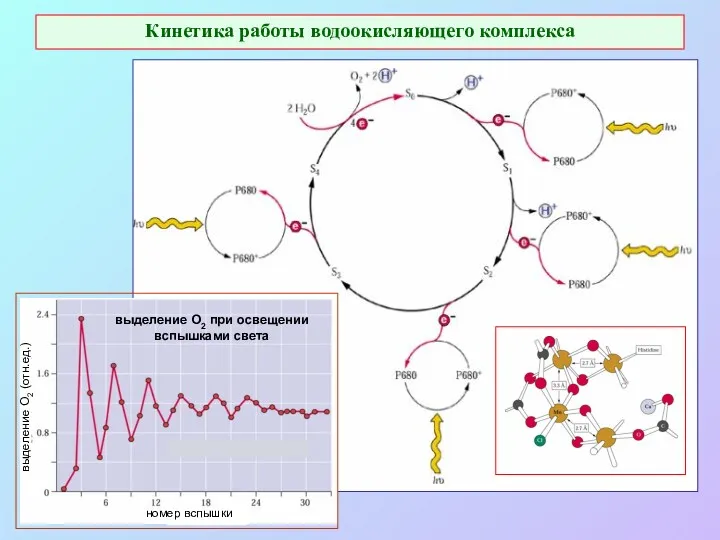
Кинетика работы водоокисляющего комплекса
номер вспышки
выделение О2 (отн.ед.)
выделение O2 при освещении
вспышками
Кинетика работы водоокисляющего комплекса
номер вспышки
выделение О2 (отн.ед.)
выделение O2 при освещении вспышками
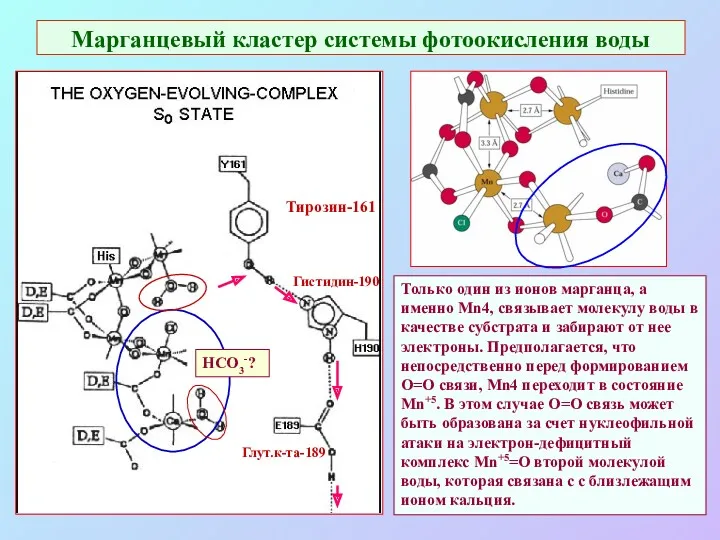
Марганцевый кластер системы фотоокисления воды
Тирозин-161
Гистидин-190
Глут.к-та-189
Только один из ионов марганца, а именно
Марганцевый кластер системы фотоокисления воды
Тирозин-161
Гистидин-190
Глут.к-та-189
Только один из ионов марганца, а именно
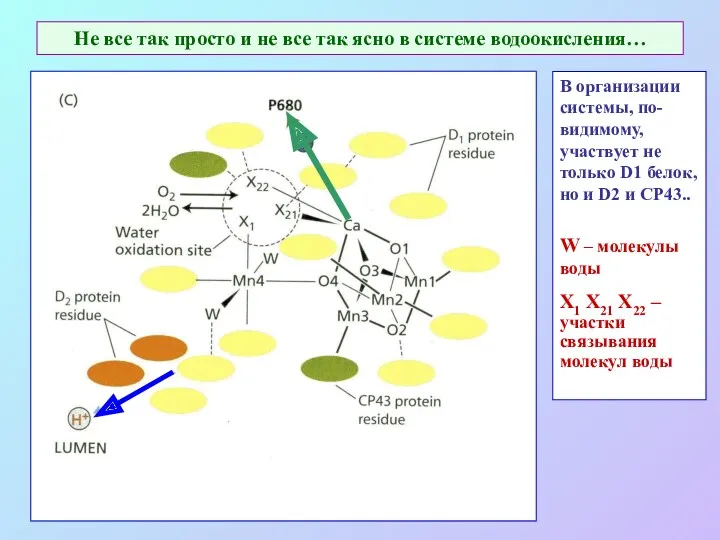
Не все так просто и не все так ясно в системе
Не все так просто и не все так ясно в системе

Фотосистема II в «реальном виде»
Фотосистема II в «реальном виде»
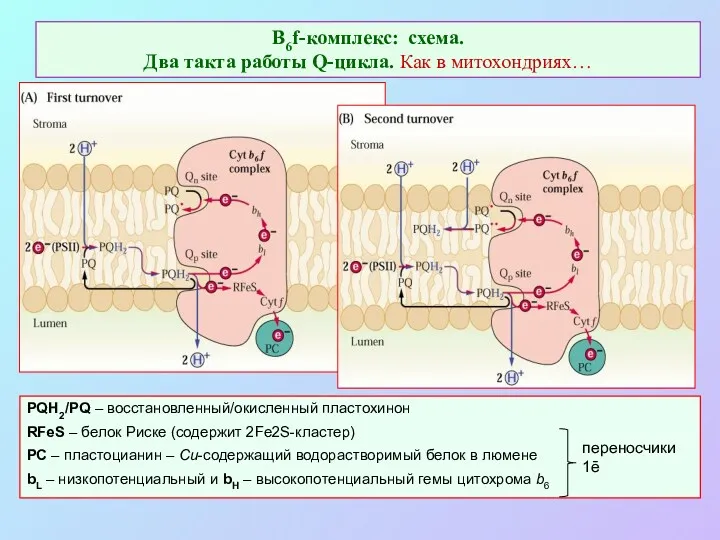
В6f-комплекс: схема.
Два такта работы Q-цикла. Как в митохондриях…
PQH2/PQ – восстановленный/окисленный пластохинон
RFeS
В6f-комплекс: схема.
Два такта работы Q-цикла. Как в митохондриях…
PQH2/PQ – восстановленный/окисленный пластохинон
RFeS
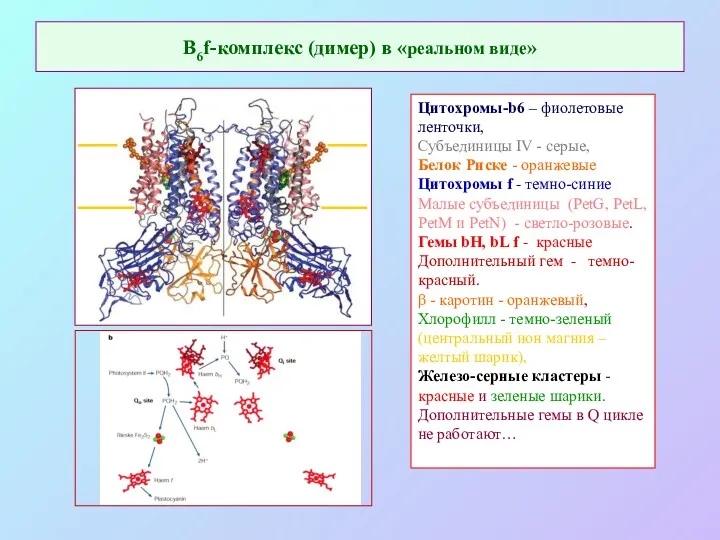
В6f-комплекс (димер) в «реальном виде»
Цитохромы-b6 – фиолетовые ленточки,
Субъединицы IV -
В6f-комплекс (димер) в «реальном виде»
Цитохромы-b6 – фиолетовые ленточки,
Субъединицы IV -

Фотосистема I: схема реакционного центра (РЦ)
ФСI – 13 белков:
А + В
Фотосистема I: схема реакционного центра (РЦ)
ФСI – 13 белков:
А + В
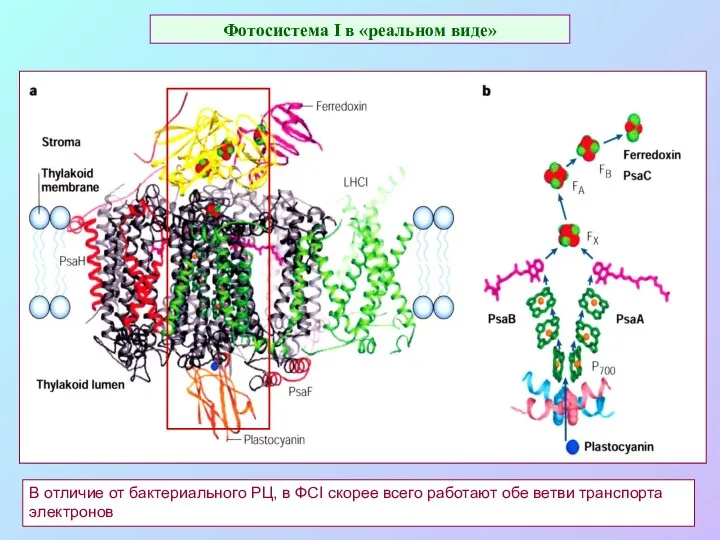
Фотосистема I в «реальном виде»
В отличие от бактериального РЦ, в ФСI
Фотосистема I в «реальном виде»
В отличие от бактериального РЦ, в ФСI
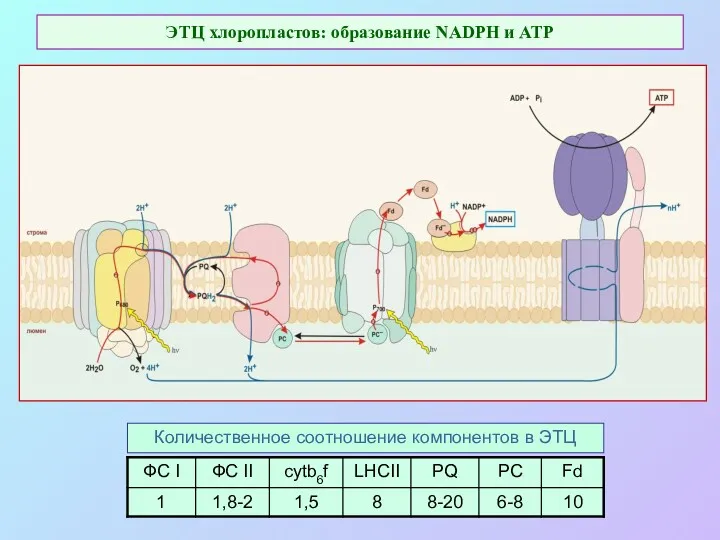
ЭТЦ хлоропластов: образование NADPH и ATP
Количественное соотношение компонентов в ЭТЦ
ЭТЦ хлоропластов: образование NADPH и ATP
Количественное соотношение компонентов в ЭТЦ
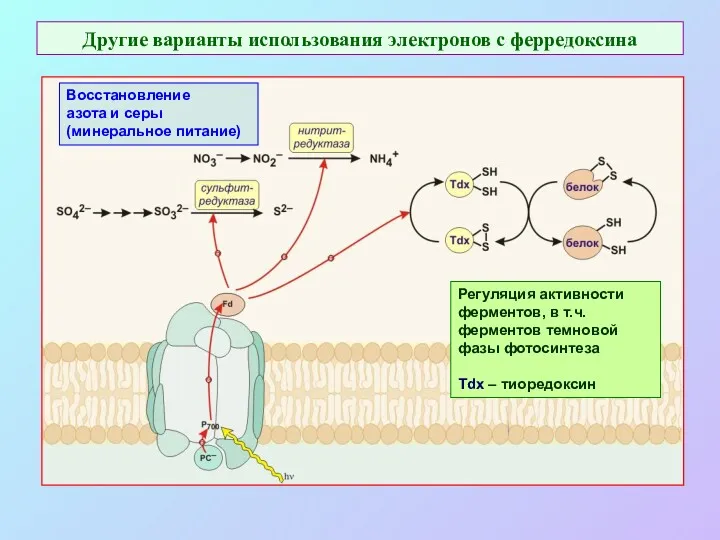
Другие варианты использования электронов с ферредоксина
Восстановление
азота и серы
(минеральное
Другие варианты использования электронов с ферредоксина
Восстановление азота и серы (минеральное
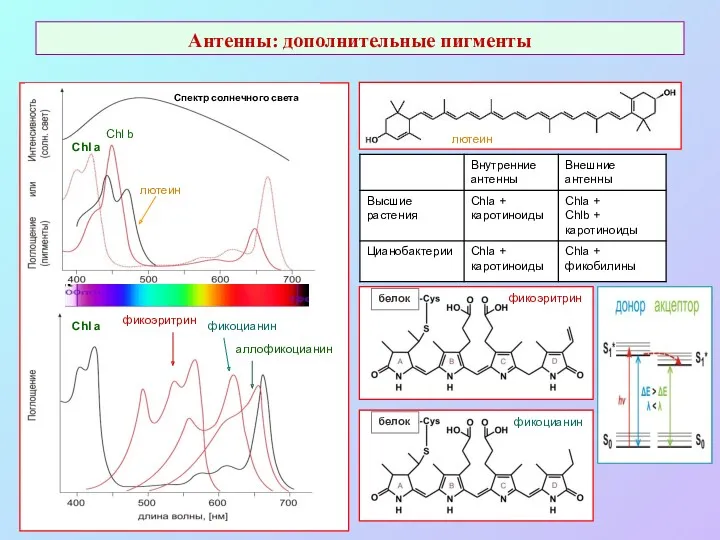
Антенны: дополнительные пигменты
фикоэритрин
фикоцианин
аллофикоцианин
Chl a
Chl a
Chl b
Спектр солнечного света
лютеин
фикоэритрин
фикоцианин
лютеин
белок
белок
Антенны: дополнительные пигменты
фикоэритрин
фикоцианин
аллофикоцианин
Chl a
Chl a
Chl b
Спектр солнечного света
лютеин
фикоэритрин
фикоцианин
лютеин
белок
белок
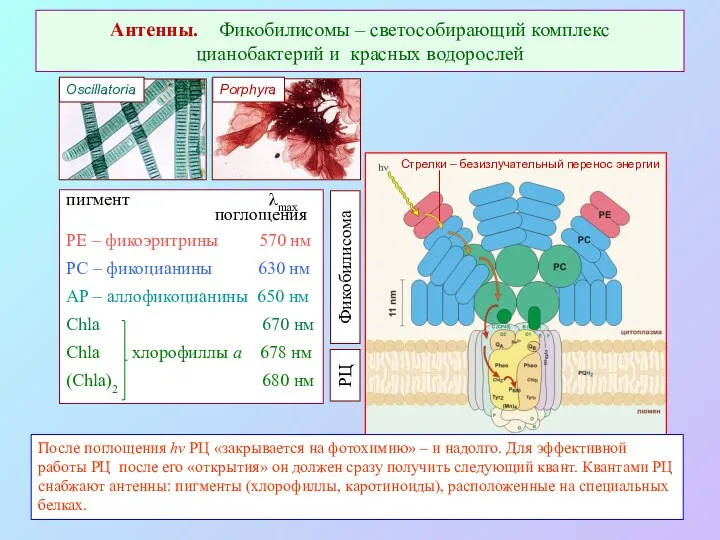
Антенны. Фикобилисомы – светособирающий комплекс цианобактерий и красных водорослей
пигмент λmax поглощения
Антенны. Фикобилисомы – светособирающий комплекс цианобактерий и красных водорослей
пигмент λmax поглощения
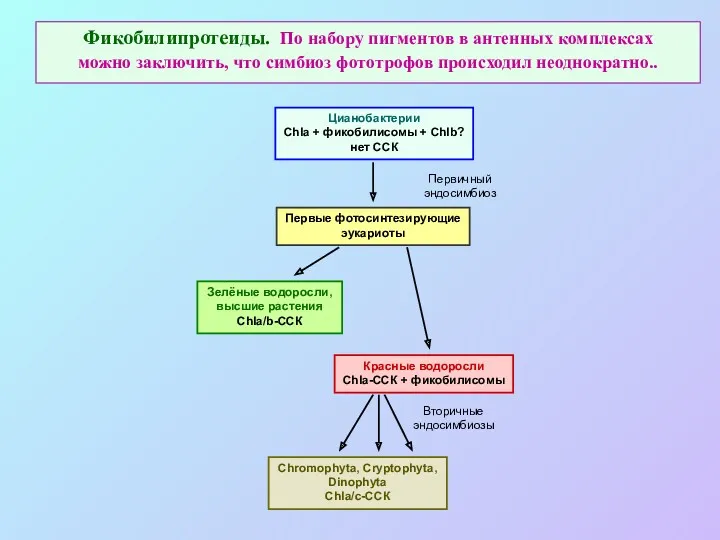
Фикобилипротеиды. По набору пигментов в антенных комплексах
можно заключить, что симбиоз
Фикобилипротеиды. По набору пигментов в антенных комплексах можно заключить, что симбиоз
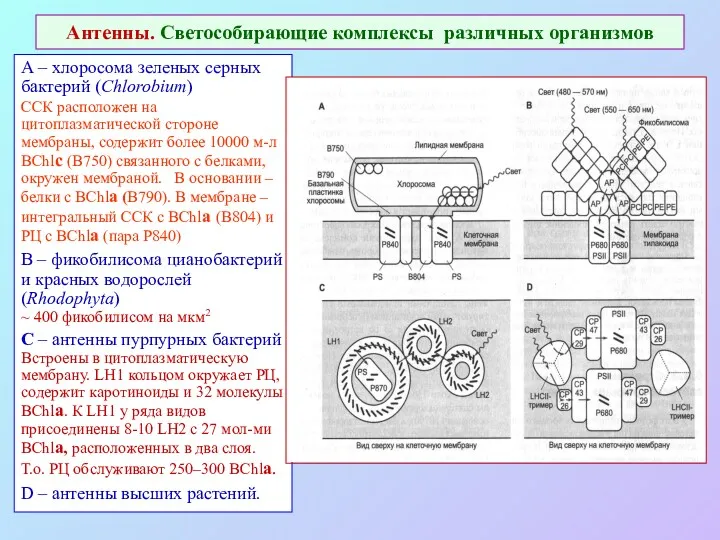
Антенны. Светособирающие комплексы различных организмов
А – хлоросома зеленых серных бактерий (Chlorobium)
ССК
Антенны. Светособирающие комплексы различных организмов
А – хлоросома зеленых серных бактерий (Chlorobium)
ССК
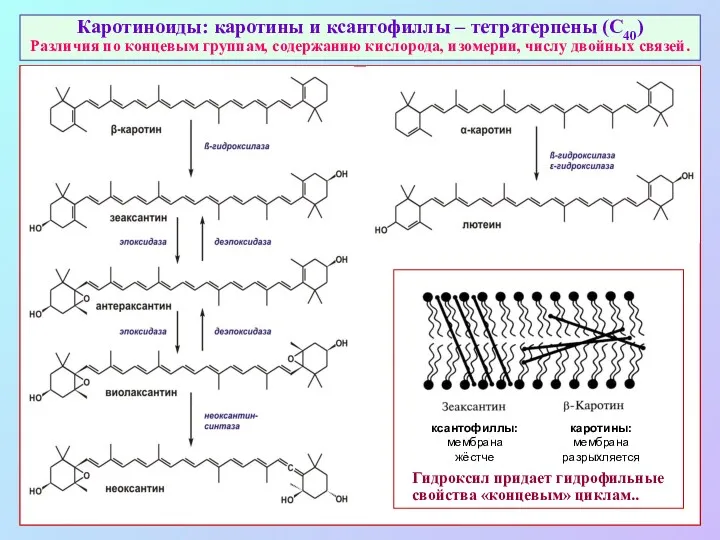
Каротиноиды: каротины и ксантофиллы – тетратерпены (С40)
Различия по концевым группам, содержанию
Каротиноиды: каротины и ксантофиллы – тетратерпены (С40) Различия по концевым группам, содержанию
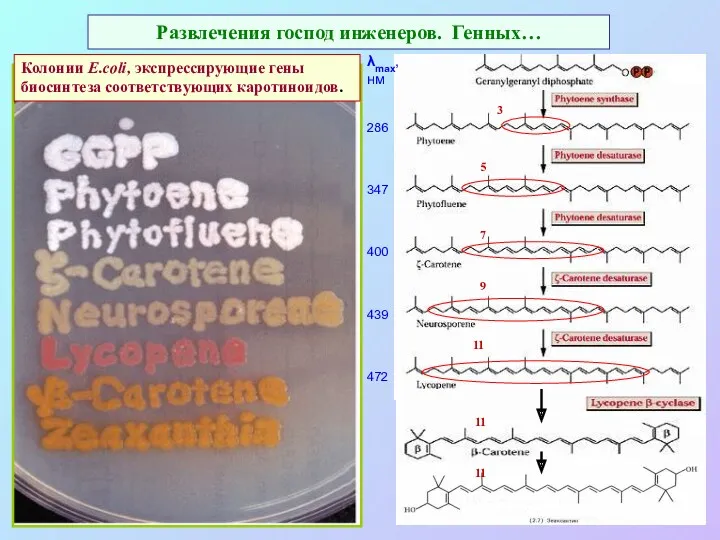
Развлечения господ инженеров. Генных…
Колонии E.coli, экспрессирующие гены биосинтеза соответствующих каротиноидов.
5
7
9
11
11
11
3
λmax,
нм
286
347
400
439
472
Развлечения господ инженеров. Генных…
Колонии E.coli, экспрессирующие гены биосинтеза соответствующих каротиноидов.
5
7
9
11
11
11
3
λmax,
нм
286
347
400
439
472
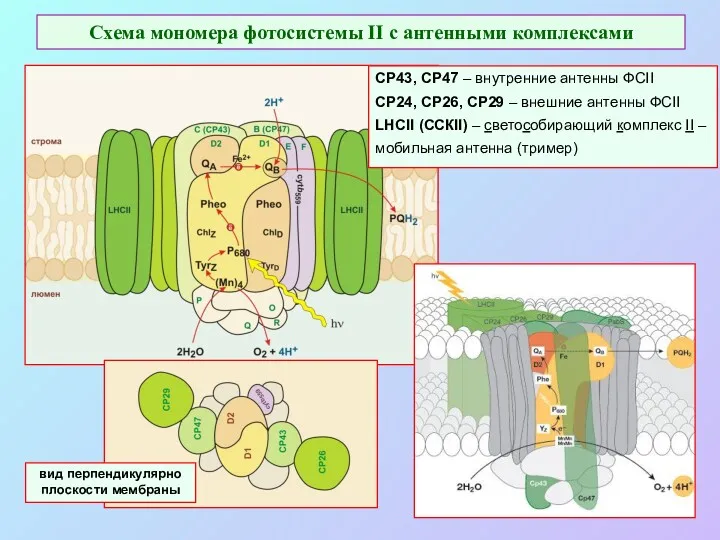
Схема мономера фотосистемы II с антенными комплексами
СР43, СР47 – внутренние антенны
Схема мономера фотосистемы II с антенными комплексами
СР43, СР47 – внутренние антенны
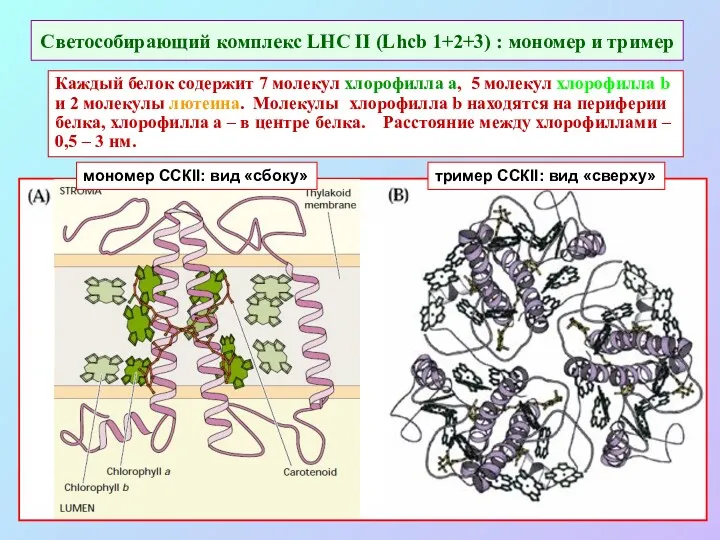
Светособирающий комплекс LHC II (Lhcb 1+2+3) : мономер и тример
Каждый белок
Светособирающий комплекс LHC II (Lhcb 1+2+3) : мономер и тример
Каждый белок
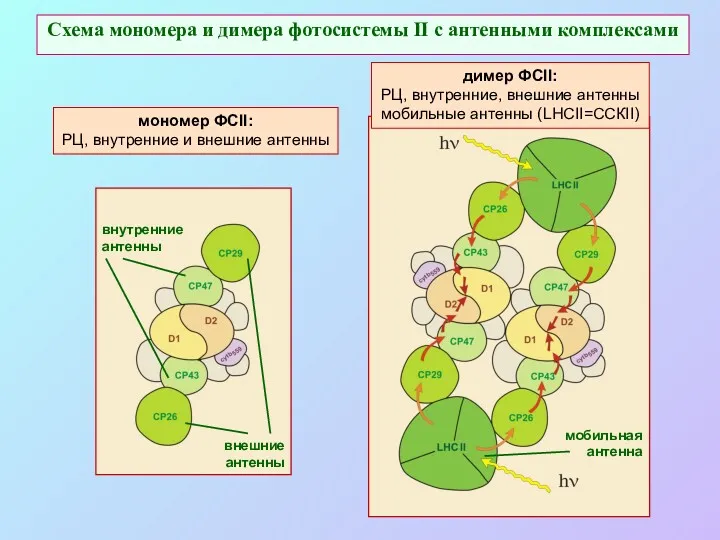
Схема мономера и димера фотосистемы II с антенными комплексами
мономер ФСII:
РЦ, внутренние
Схема мономера и димера фотосистемы II с антенными комплексами
мономер ФСII:
РЦ, внутренние
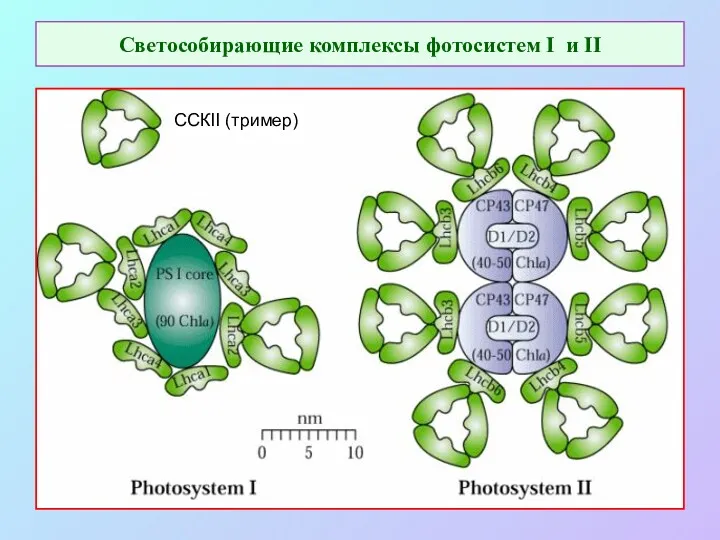
Светособирающие комплексы фотосистем I и II
ССКII (тример)
Светособирающие комплексы фотосистем I и II
ССКII (тример)
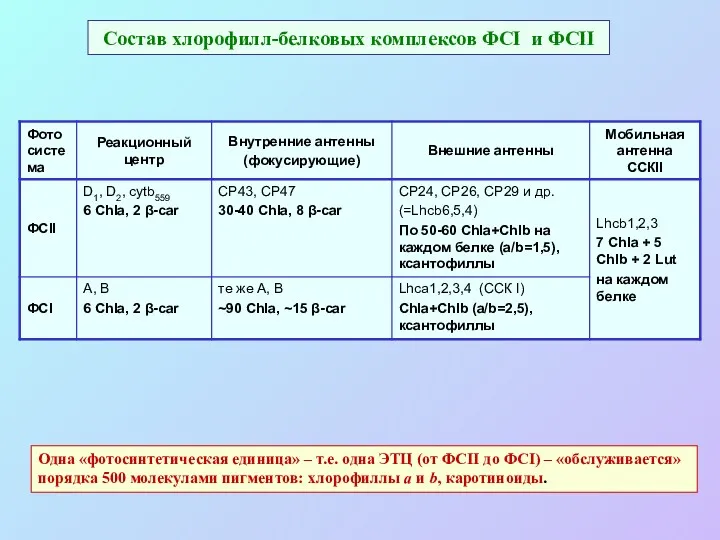
Состав хлорофилл-белковых комплексов ФСI и ФСII
Одна «фотосинтетическая единица» – т.е. одна
Состав хлорофилл-белковых комплексов ФСI и ФСII
Одна «фотосинтетическая единица» – т.е. одна
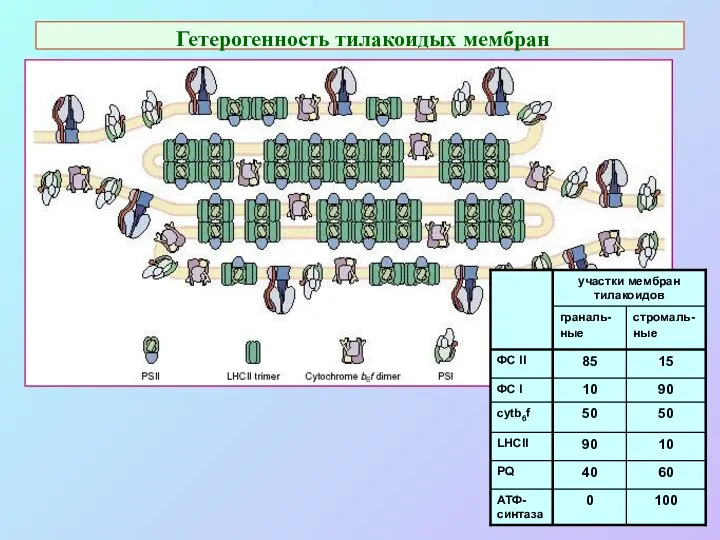
Гетерогенность тилакоидых мембран
Гетерогенность тилакоидых мембран

Hosta
‘Piedmont Gold’
Hosta sieboldiana
‘Elegans’
Hosta
‘Golden Haze’
Фотовыцветание
Hosta
‘Piedmont Gold’
Hosta sieboldiana
‘Elegans’
Hosta
‘Golden Haze’
Фотовыцветание

Механизмы регулирования и защиты ФСII от фотодеструкции
нециклический поток, регулирование мобильными антеннами;
циклические
Механизмы регулирования и защиты ФСII от фотодеструкции
нециклический поток, регулирование мобильными антеннами;
циклические
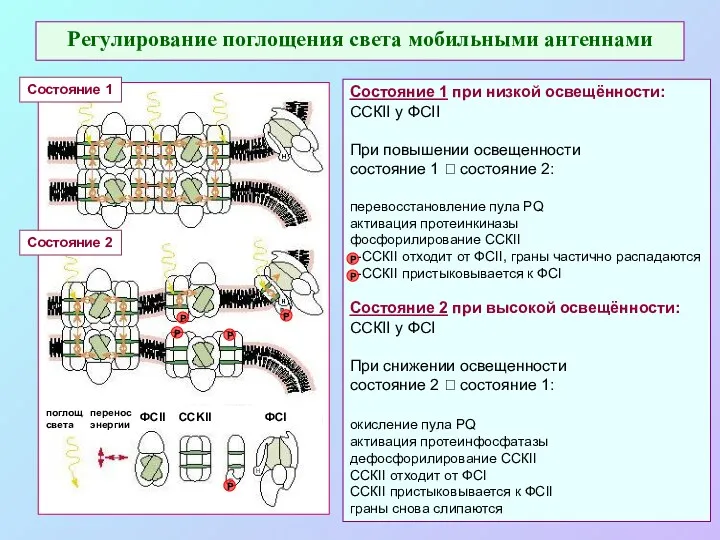
Регулирование поглощения света мобильными антеннами
Р
Р
Р
Р
Р
Состояние 1
Состояние 2
Состояние 1 при низкой освещённости:
ССКII
Регулирование поглощения света мобильными антеннами
Р
Р
Р
Р
Р
Состояние 1
Состояние 2
Состояние 1 при низкой освещённости:
ССКII

Варианты электронного транспорта в ЭТЦ хлоропластов
Варианты транспорта ē: Продукты:
0 – Нециклический
Варианты электронного транспорта в ЭТЦ хлоропластов
Варианты транспорта ē: Продукты:
0 – Нециклический
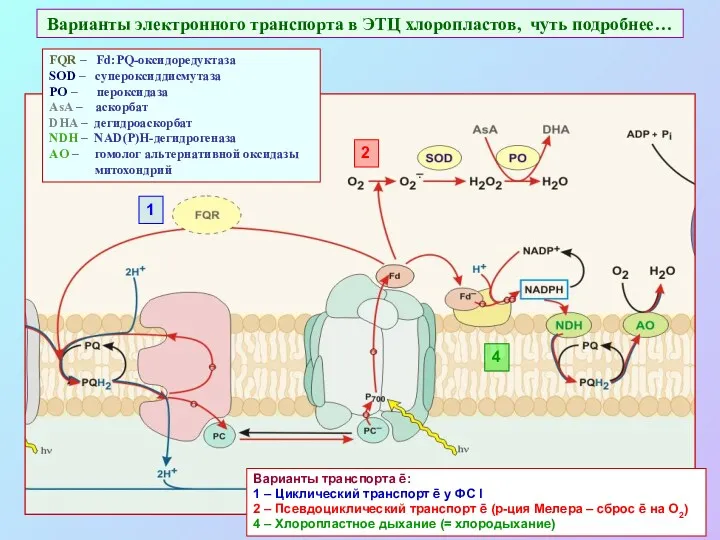
Варианты электронного транспорта в ЭТЦ хлоропластов, чуть подробнее…
1
2
4
Варианты транспорта ē:
1
Варианты электронного транспорта в ЭТЦ хлоропластов, чуть подробнее…
1
2
4
Варианты транспорта ē:
1
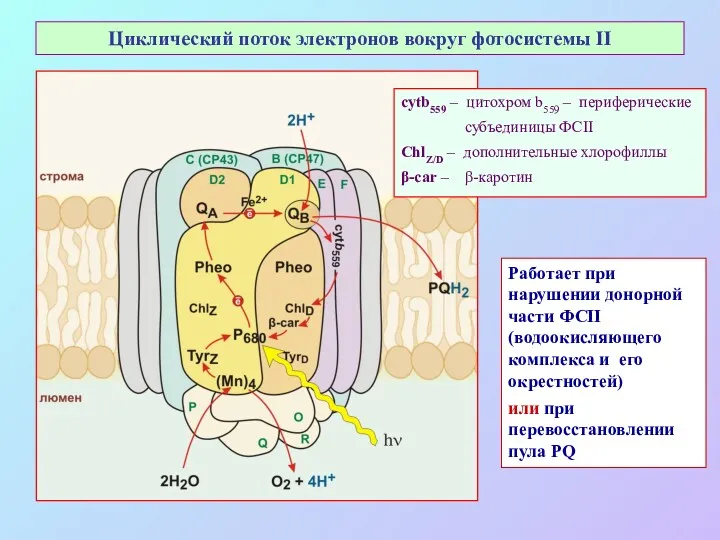
Циклический поток электронов вокруг фотосистемы II
cytb559 – цитохром b559 – периферические
Циклический поток электронов вокруг фотосистемы II
cytb559 – цитохром b559 – периферические
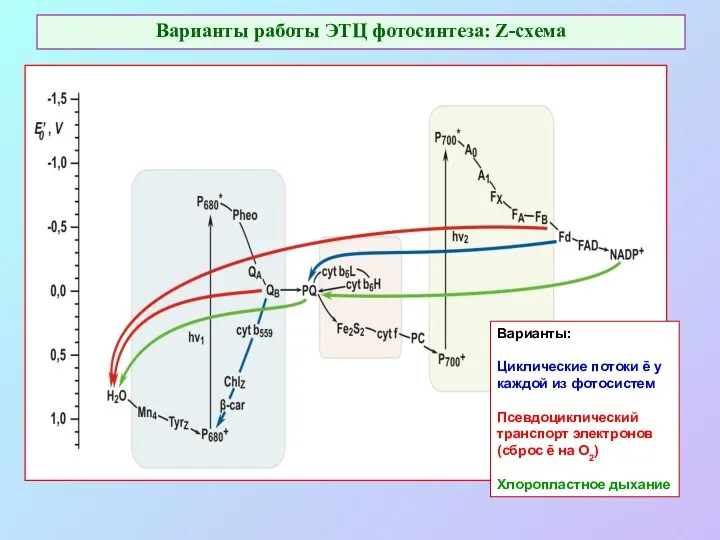
Варианты работы ЭТЦ фотосинтеза: Z-схема
Варианты:
Циклические потоки ē у каждой из фотосистем
Псевдоциклический
Варианты работы ЭТЦ фотосинтеза: Z-схема
Варианты:
Циклические потоки ē у каждой из фотосистем
Псевдоциклический
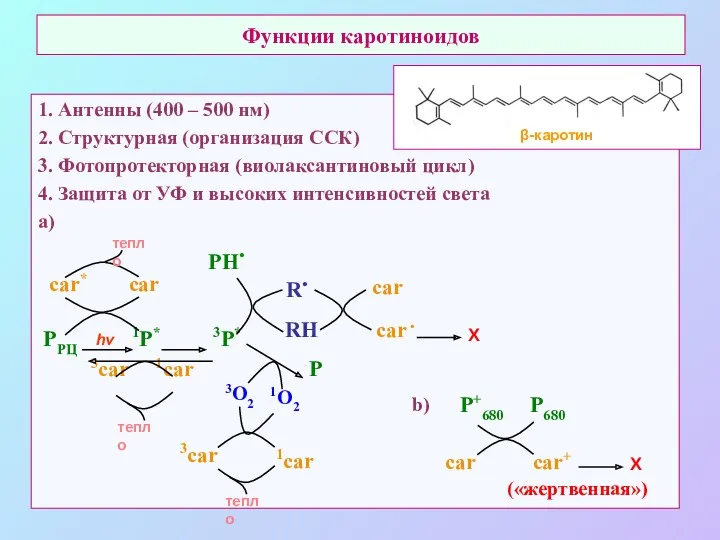
Функции каротиноидов
1. Антенны (400 – 500 нм)
2. Структурная (организация ССК)
3. Фотопротекторная
Функции каротиноидов
1. Антенны (400 – 500 нм)
2. Структурная (организация ССК)
3. Фотопротекторная
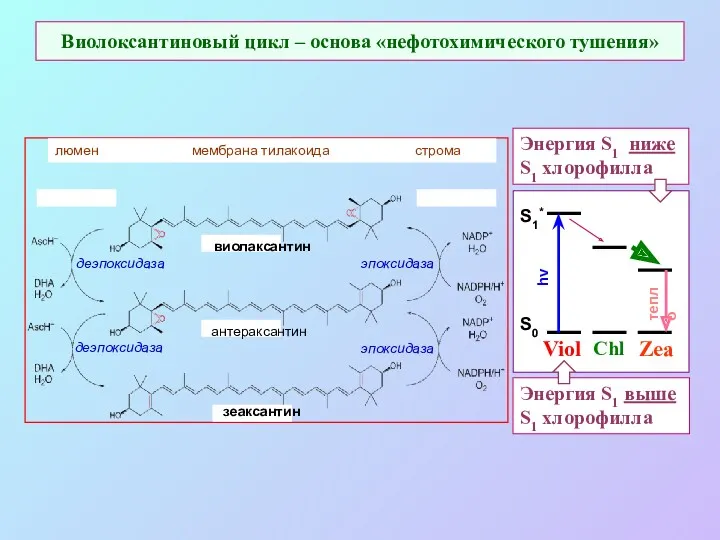
Виолоксантиновый цикл – основа «нефотохимического тушения»
Энергия S1 ниже
S1 хлорофилла
Энергия S1
Виолоксантиновый цикл – основа «нефотохимического тушения»
Энергия S1 ниже
S1 хлорофилла
Энергия S1
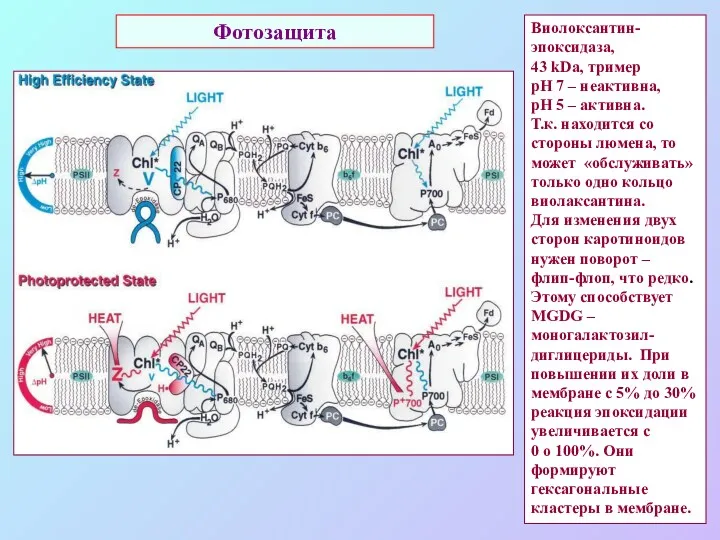
Фотозащита
Виолоксантин-эпоксидаза,
43 kDa, тример
рН 7 – неактивна,
рН 5 – активна.
Т.к. находится со
Фотозащита
Виолоксантин-эпоксидаза,
43 kDa, тример
рН 7 – неактивна,
рН 5 – активна.
Т.к. находится со
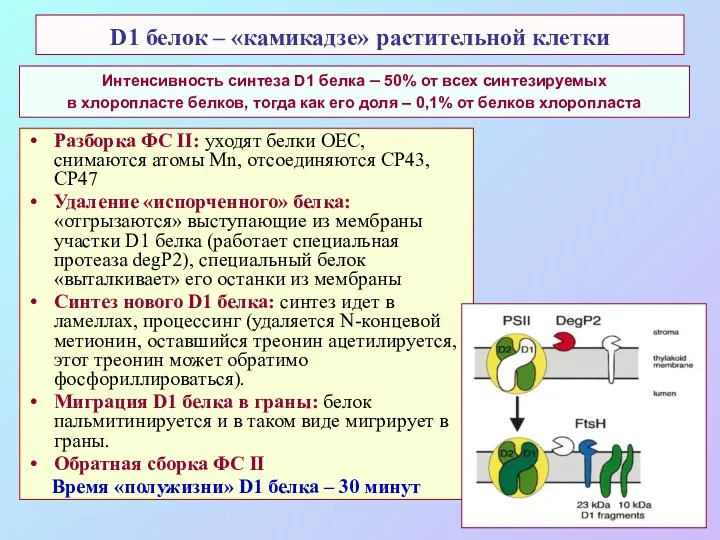
D1 белок – «камикадзе» растительной клетки
Разборка ФС II: уходят белки OEC,
D1 белок – «камикадзе» растительной клетки
Разборка ФС II: уходят белки OEC,
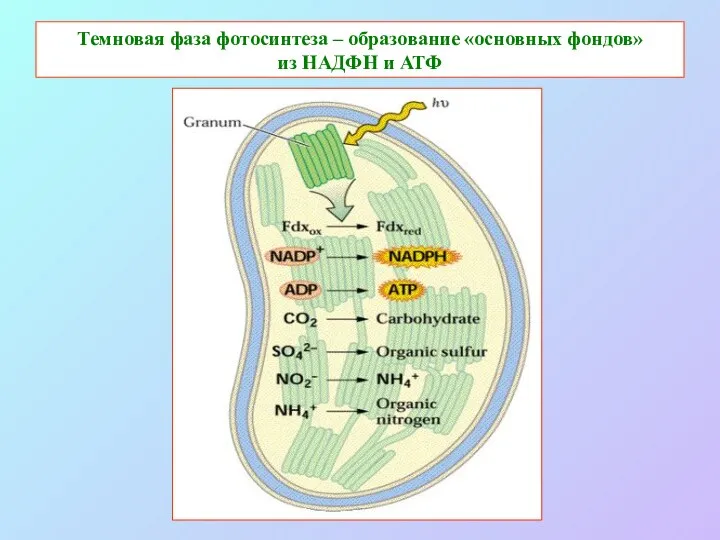
Темновая фаза фотосинтеза – образование «основных фондов»
из НАДФН и АТФ
Темновая фаза фотосинтеза – образование «основных фондов»
из НАДФН и АТФ
 Тест Земноводные, Пресмыкающиеся
Тест Земноводные, Пресмыкающиеся Многообразие Млекопитающих
Многообразие Млекопитающих Класс млекопитающие. Отряд хищные
Класс млекопитающие. Отряд хищные Растения. Систематика
Растения. Систематика Разновидности мышечного волокна. Сократительные белки. Механизм сокращения
Разновидности мышечного волокна. Сократительные белки. Механизм сокращения Нормальная микрофлора тела человека
Нормальная микрофлора тела человека Отряд Стрекозы
Отряд Стрекозы Развитие кровеносной системы
Развитие кровеносной системы Fauna
Fauna Анатомическое строение корня
Анатомическое строение корня Variability is the ability of living organisms to acquire new marks
Variability is the ability of living organisms to acquire new marks История изучения клетки. Клеточная теория
История изучения клетки. Клеточная теория Семейства растений
Семейства растений Микробиология мяса, как продукта питания
Микробиология мяса, как продукта питания Клонирование и этические проблемы
Клонирование и этические проблемы Animals high. Body parts
Animals high. Body parts Биологическое оружие
Биологическое оружие Кровообращение и лимфообращение
Кровообращение и лимфообращение Классы рыб
Классы рыб Своя игра
Своя игра Семейство крестоцветные
Семейство крестоцветные Отдаленные последствия облучения
Отдаленные последствия облучения Мичурин Иван Владимирович
Мичурин Иван Владимирович тема самообразования
тема самообразования игра Что Где Когда по биологии
игра Что Где Когда по биологии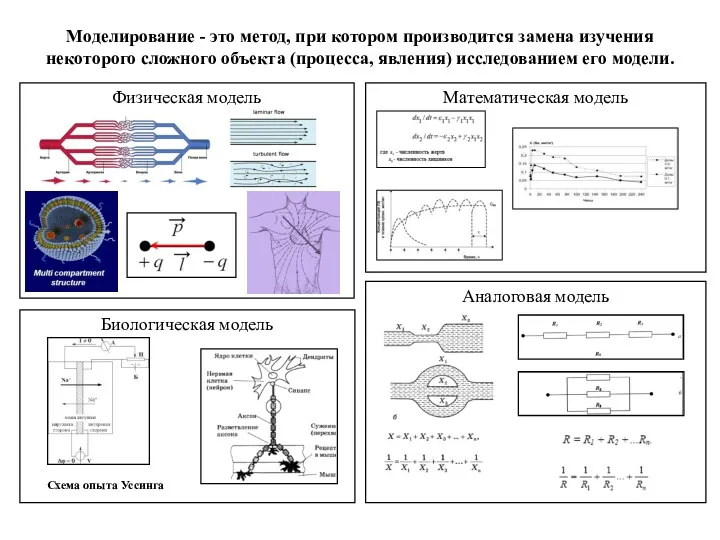 Физическая, математическая, аналоговая, биологическая модели процессов. Моделирование. (Лекция 1)
Физическая, математическая, аналоговая, биологическая модели процессов. Моделирование. (Лекция 1) Симбиотические организмы лишайники
Симбиотические организмы лишайники Основы микробиологии и биотехнологии
Основы микробиологии и биотехнологии