- Разновидности мышечного волокна. Сократительные белки. Механизм сокращения

Содержание
- 2. Типы и свойства мышечной ткани Возбудимость Сократимость Проводимость Эластичность
- 3. Макроскопическое строение мышц Брюшко Головка мышцы, Хвост мышцы, Мышцы-антагонисты, Мышцы-синергисты, Апоневроз?
- 4. МЫШЦЫ СТРОЕНИЕ КЛАССИФИКАЦИЯ БРЮШКО ФАСЦИЯ ПОВЕРХНОСТНАЯ ГЛУБОКАЯ МЫШЕЧНЫЕ ПУЧКИ ВСПОМОГА-ТЕЛЬНЫЙ АПАРАТ СЫНОВИАЛЬНАЯ СУМКА СЕСАМОВИДНЫЕ КОСТИ ЗА
- 5. Микроскопическое строение мышц. Мышечный пучок Под какой буквой обозначены гладкая и поперечнополосатая мускулатура? А-; Б-. Что
- 6. Схема строения мышечного волокна Саркомер - с двух сторон ограничен Z – мембранами. Толстые – миозиновые,
- 8. Саркомер в расслабленном состоянии Саркомер в состоянии сокращения
- 9. Саркоплазма В саркоплазме находится весь набор типичных для любой клетки органоидов. Особо следует подчеркнуть наличие: -
- 10. Актиновые миофиламенты Актиновые филаменты, скомпанованы из двух актиновых нитей, представляющих собой как бы бусинки глобулярных молекул
- 11. Схема строения актиновых и миозиновых филаментов Миозиновые филаменты образуются более чем двумястами молекулами миозина. Каждая из
- 12. актин тропонин тропомиозин Са+2
- 13. миозин головка шейка
- 14. Актино-миозиновые мостики
- 15. Цикл работы головки миозина
- 16. Нервно-мышечный синапс 1 - пресинаптическая мембрана, 2 - пузырьки с ацетилхолином, 3 - митохондрии, 4 -
- 17. Ах Аксонная терминаль Аксонная бляшка Ацетилхолин Пресинаптическая мембрана Постсинаптическая мембрана Синаптическая щель Рецептор Хемозависимый канал Электроза-
- 18. Ах ПД Са++ Са++ Ах ПД Na+ Na+ ПКП Ацетилхолин- эстераза Na+ Na+
- 19. Строение нервно-мышечного синапса в разрезе
- 20. Электрохимическое преобразование ПД по мембране + дегидропиридина в триадах + рианодина в СПР Открытие каналов для
- 21. Без ПД кальций в цитоплазму не выйдет!!
- 22. Механизм расслабления мышцы [Са+2] 107 М + кальциевый насос в СПР СПР Са+2 Са+2 Са+2 АТФ
- 23. Са+2
- 24. Затраты энергии АТФ во время мышечного сокращения На поддержание ионной асимметрии (натрий-калиевй насос) Образование мостиков («приклеивание»
- 25. Двигательная или моторная единица Каждое мышечное волокно имеет только один синапс
- 26. Двигательная или моторная единица Мотонейрон и группа мышечных волокон, иннервируемых разветвлениями аксона этого мотонейрона Число мышечных
- 27. Строение двигательной единицы
- 28. Распределение синапсов аксона мотонейрона в скелетной мышце
- 29. Классификация мышечных волокон
- 30. Медленные фазические волокна окислительного типа большое содержание миоглобина и митохондрий красного цвета медленно утомляются В одной
- 31. Быстрые фазические волокна окислительного типа Содержат много митохондрий Способны синтезировать АТФ путем окислительного фосфорилирования Выполняют быстрые
- 32. Быстрые фазические с гликолитическим типом окисления Мало митохондрий АТФ образуется за счет гликолиза Миоглобина нет (белый
- 33. Тонические волокна Двигательный аксон образует множество синапсов Медленно сокращаются и медленно расслеабдяются Низкая АТФ-азная активность миозина
- 34. Закон «все или ничего» Одиночное мышечное волокно подчиняется этому закону: Подпороговое раздражение не вызывает сокращение, а
- 35. Закон «все или ничего» Целая мышца данному закону не подчиняется потому, что состоит из множества моторных
- 36. Режимы сокращения Изометрический – увеличение напряжения без изменения длины мышцы Изотонические – уменьшение длины мышцы без
- 37. Виды мышечных сокращений Одиночное Тетанус Тонус
- 38. Виды раздражения мышцы в эксперименте Непрямое раздражение – импульс воздействует на нервное окончание мотонейрона Прямое раздражение
- 39. Одиночное мышечное сокращение
- 40. Одиночное мышечное сокращение 1 2 3 1 – латентный период 2 – период напряжения 3 –
- 41. Тетанус Это сильное и длительное сокращение мышцы в ответ на серию раздражений. Происходит за счет суммации
- 42. Суммация одиночных сокращений
- 43. Зубчатый тетанус Возникает в условиях когда каждый последующий импульс попадает в период расслабления или интервал между
- 46. Гладкий тетанус Возникает в условиях когда каждый последующий импульс попадает в период укорочения Или интервал между
- 49. оптимум Частота раздражения, при которой наблюдается суммарное сокращение (тетанус) наибольшей амплитуды При этом каждый последующий импульс
- 50. пессимум Частота раздражения, при которй не наблюдается суммации сокращения При этом каждый последующий импульс попадает в
- 52. 1 2 3 4 5 Е0 Екр 0 +30
- 53. 1 2 3 4 5 Е0 Екр 0 +30 В Г А Б
- 54. Сила мышц Зависит от толщины мышцы и её поперечного физиологического сечения
- 55. Работа мышцы Это энергия, затрачиваемая на перемещение тела с определенной силой на определенное расстояние: A =
- 56. утомление Процесс временного снижения работоспособности мышцы. Возникает в связи с уменьшением энергетических запасов (АТФ) в мышечном
- 58. Утомление в организме Утомление развивается вначале в нервных центрах – это защитный механизм При этом в
- 59. Гладкие мышцы
- 60. Висцеральные гладкие мышцы Все внутренние органы Большое количество нексусов (красные) Мало нервных окончаний (зеленые)
- 61. Мультиунитарные гладкие мышцы Представлены ресничной и мышцей радужки глаза Отмечается большая плотность нервных синаптических (варикозных) контактов
- 62. Иннервация ГМК
- 63. Особенности ГМК Веретенообразные клетки, тесно прилежат друг к другу Между клетками – плотные контакты – нексусы
- 64. Иннервация ГМК Иннервируют симпатические и парасимпатические нервные волокна Нет синапсов, нервные окончания образуют варикозные расширения Медиатор
- 65. Адекватные раздражители Нервный импульс Механическое растяжение Химические вещества
- 66. Особенности возбудимости Порог возбудимости ниже, чем у поперечно-полосатых мышц Мембрана более проницаема для натрия, поэтому МП
- 67. Автоматия Способность клетки самостоятельно без внешнего раздражителя генерировать ПД ГМК в состоянии покоя пропускает натрий, который
- 68. Электрохимическое сопряжение в ГМК ПД открывает кальциевые каналы и в клетку входит кальций. Лиганд - активирует
- 69. Механизм регуляции содержания кальция в ГМК
- 70. Особенности механизма сокращения в ГМК Кальций в цитоплазме связывается с кальмодулином и активирует протеинкиназу . Протеинкиназа
- 72. Скачать презентацию
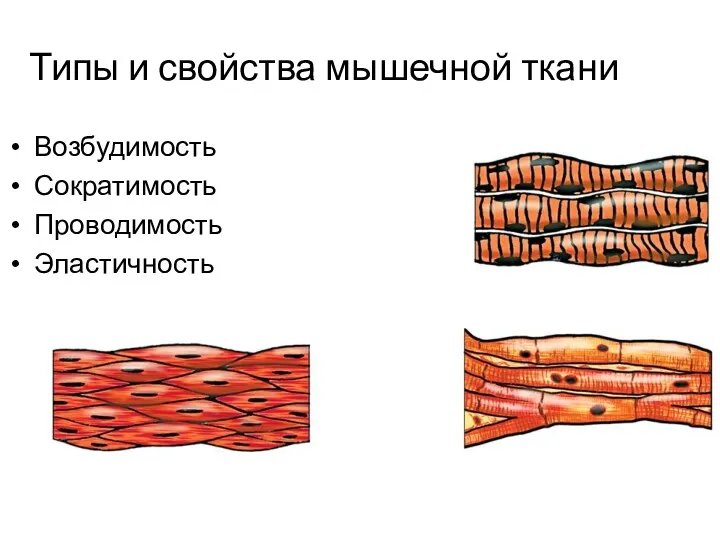
Типы и свойства мышечной ткани
Возбудимость
Сократимость
Проводимость
Эластичность
Типы и свойства мышечной ткани
Возбудимость
Сократимость
Проводимость
Эластичность
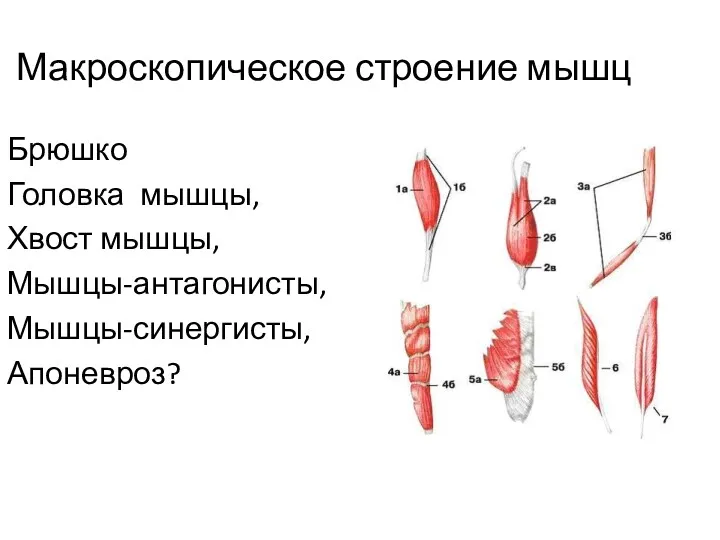
Макроскопическое строение мышц
Брюшко
Головка мышцы,
Хвост мышцы,
Мышцы-антагонисты,
Мышцы-синергисты,
Апоневроз?
Макроскопическое строение мышц
Брюшко
Головка мышцы,
Хвост мышцы,
Мышцы-антагонисты,
Мышцы-синергисты,
Апоневроз?
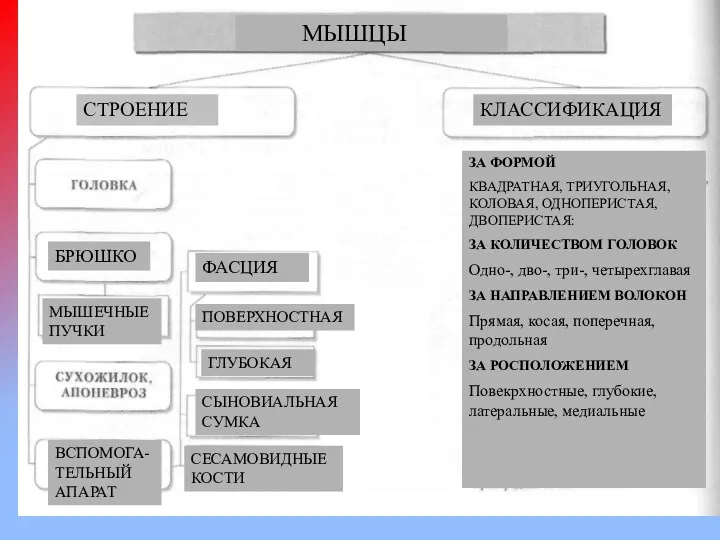
МЫШЦЫ
СТРОЕНИЕ
КЛАССИФИКАЦИЯ
БРЮШКО
ФАСЦИЯ
ПОВЕРХНОСТНАЯ
ГЛУБОКАЯ
МЫШЕЧНЫЕ ПУЧКИ
ВСПОМОГА-ТЕЛЬНЫЙ АПАРАТ
СЫНОВИАЛЬНАЯ СУМКА
СЕСАМОВИДНЫЕ КОСТИ
ЗА ФОРМОЙ
КВАДРАТНАЯ, ТРИУГОЛЬНАЯ, КОЛОВАЯ, ОДНОПЕРИСТАЯ,
МЫШЦЫ
СТРОЕНИЕ
КЛАССИФИКАЦИЯ
БРЮШКО
ФАСЦИЯ
ПОВЕРХНОСТНАЯ
ГЛУБОКАЯ
МЫШЕЧНЫЕ ПУЧКИ
ВСПОМОГА-ТЕЛЬНЫЙ АПАРАТ
СЫНОВИАЛЬНАЯ СУМКА
СЕСАМОВИДНЫЕ КОСТИ
ЗА ФОРМОЙ
КВАДРАТНАЯ, ТРИУГОЛЬНАЯ, КОЛОВАЯ, ОДНОПЕРИСТАЯ,
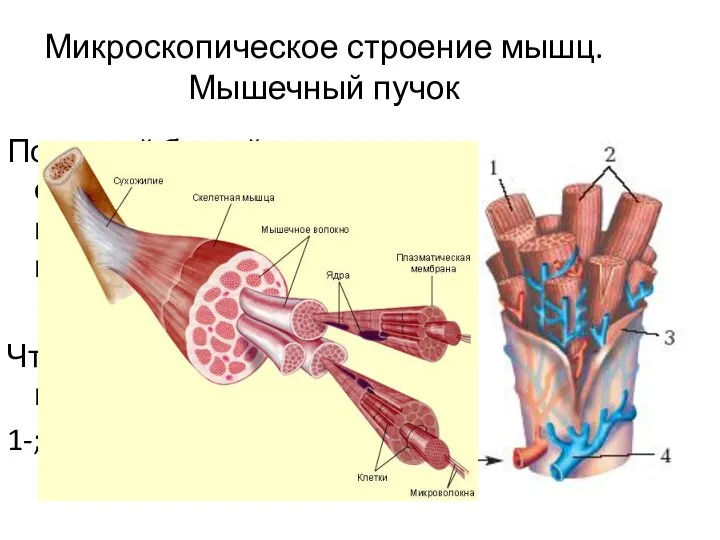
Микроскопическое строение мышц. Мышечный пучок
Под какой буквой обозначены гладкая и поперечнополосатая
Микроскопическое строение мышц. Мышечный пучок
Под какой буквой обозначены гладкая и поперечнополосатая
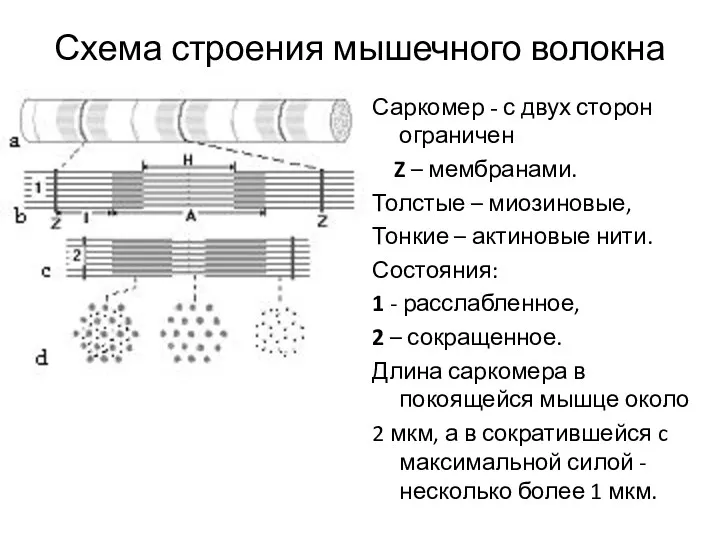
Схема строения мышечного волокна
Саркомер - с двух сторон ограничен
Z
Схема строения мышечного волокна
Саркомер - с двух сторон ограничен
Z
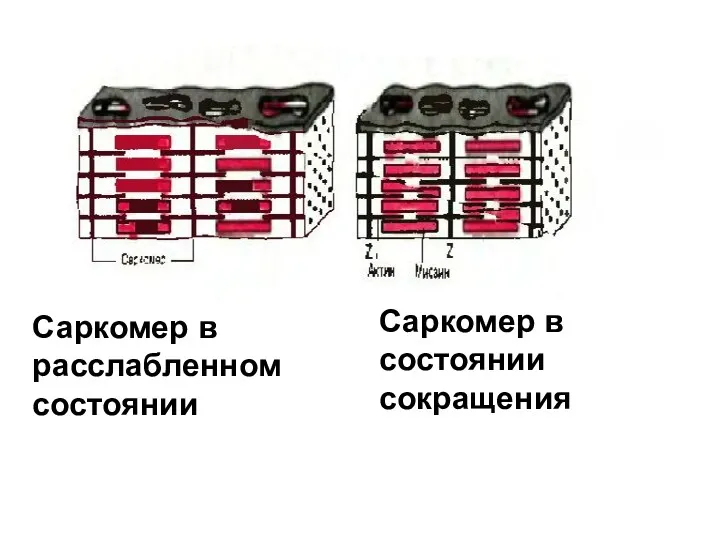
Саркомер в расслабленном
состоянии
Саркомер в состоянии
сокращения
Саркомер в расслабленном
состоянии
Саркомер в состоянии
сокращения

Саркоплазма
В саркоплазме находится весь набор типичных для любой клетки органоидов.
Особо следует
Саркоплазма
В саркоплазме находится весь набор типичных для любой клетки органоидов.
Особо следует

Актиновые миофиламенты
Актиновые филаменты, скомпанованы из двух актиновых нитей, представляющих собой как
Актиновые миофиламенты
Актиновые филаменты, скомпанованы из двух актиновых нитей, представляющих собой как

Схема строения актиновых и миозиновых филаментов
Миозиновые филаменты образуются более чем двумястами
Схема строения актиновых и миозиновых филаментов
Миозиновые филаменты образуются более чем двумястами
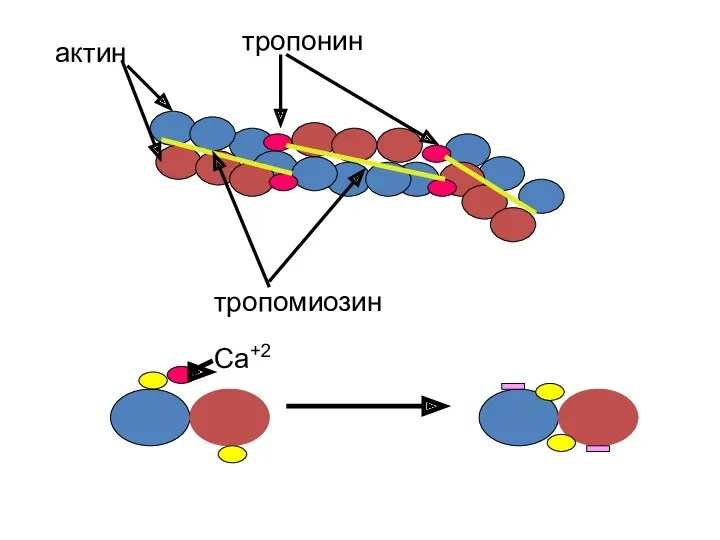
актин
тропонин
тропомиозин
Са+2
актин
тропонин
тропомиозин
Са+2
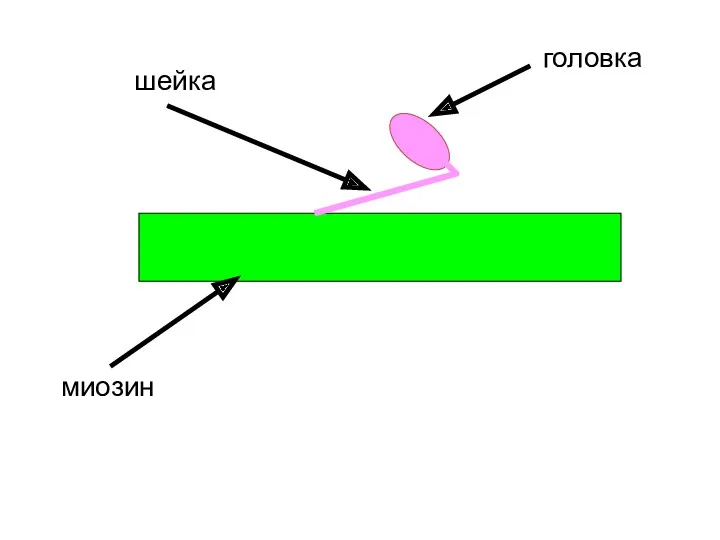
миозин
головка
шейка
миозин
головка
шейка
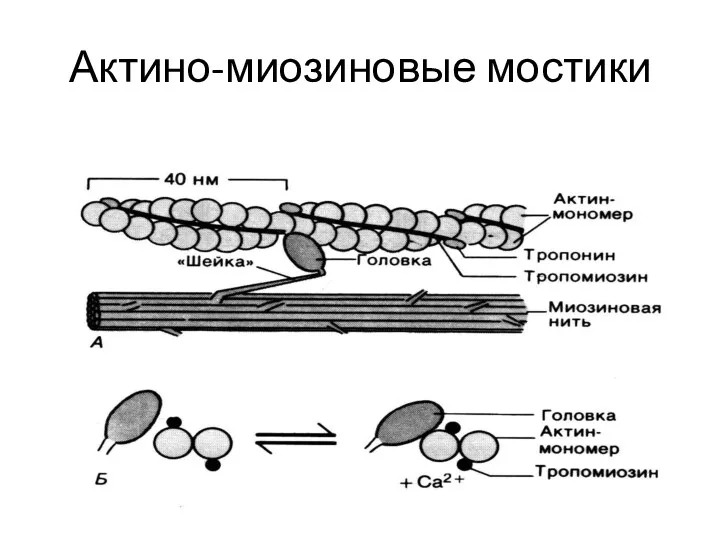
Актино-миозиновые мостики
Актино-миозиновые мостики
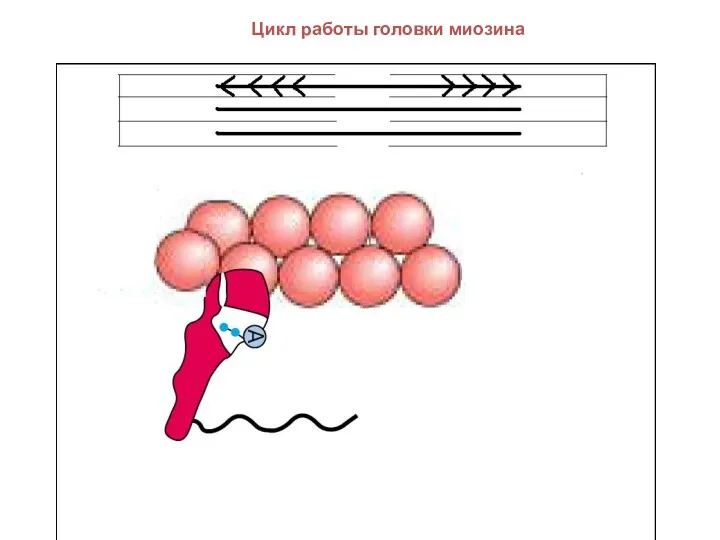
Цикл работы головки миозина
Цикл работы головки миозина
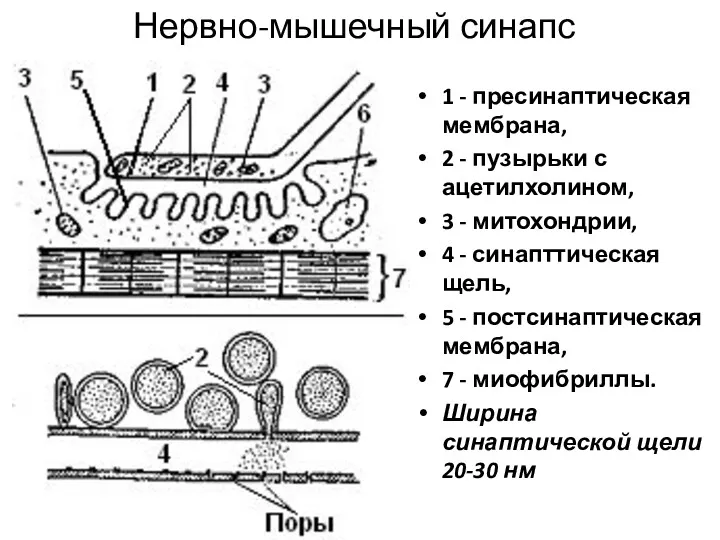
Нервно-мышечный синапс
1 - пресинаптическая мембрана,
2 - пузырьки с ацетилхолином,
3
Нервно-мышечный синапс
1 - пресинаптическая мембрана,
2 - пузырьки с ацетилхолином,
3
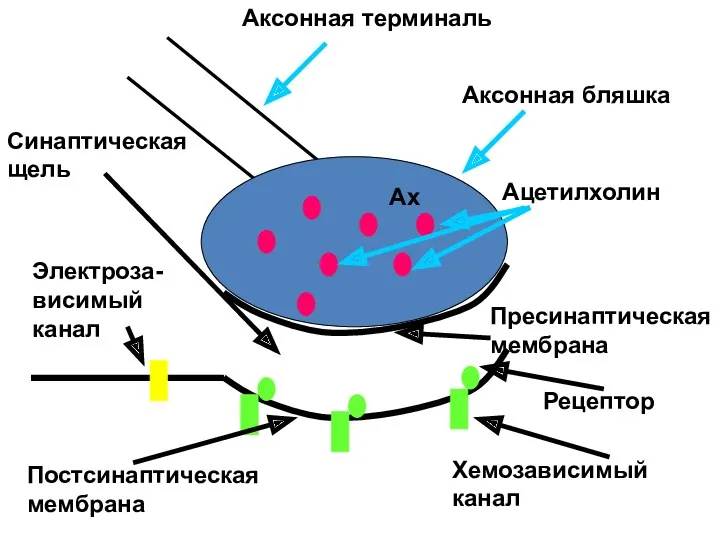
Ах
Аксонная терминаль
Аксонная бляшка
Ацетилхолин
Пресинаптическая
мембрана
Постсинаптическая
мембрана
Синаптическая
щель
Рецептор
Хемозависимый
канал
Электроза-
висимый
канал
Ах
Аксонная терминаль
Аксонная бляшка
Ацетилхолин
Пресинаптическая
мембрана
Постсинаптическая
мембрана
Синаптическая
щель
Рецептор
Хемозависимый
канал
Электроза-
висимый
канал
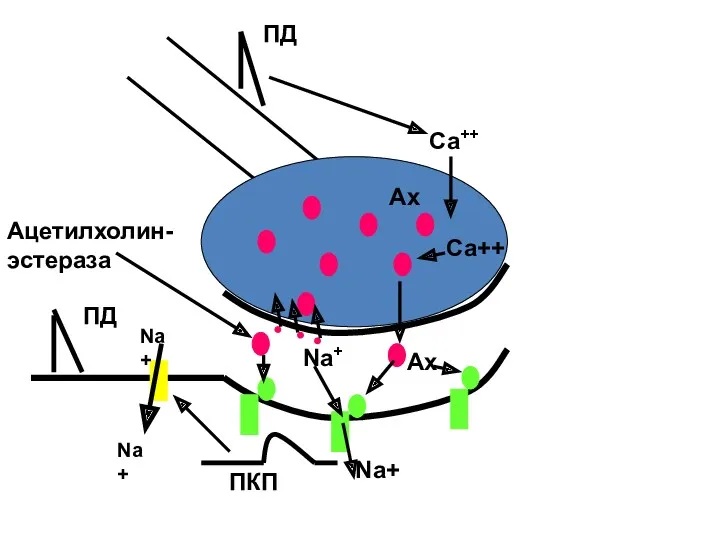
Ах
ПД
Са++
Са++
Ах
ПД
Na+
Na+
ПКП
Ацетилхолин-
эстераза
Na+
Na+
Ах
ПД
Са++
Са++
Ах
ПД
Na+
Na+
ПКП
Ацетилхолин-
эстераза
Na+
Na+
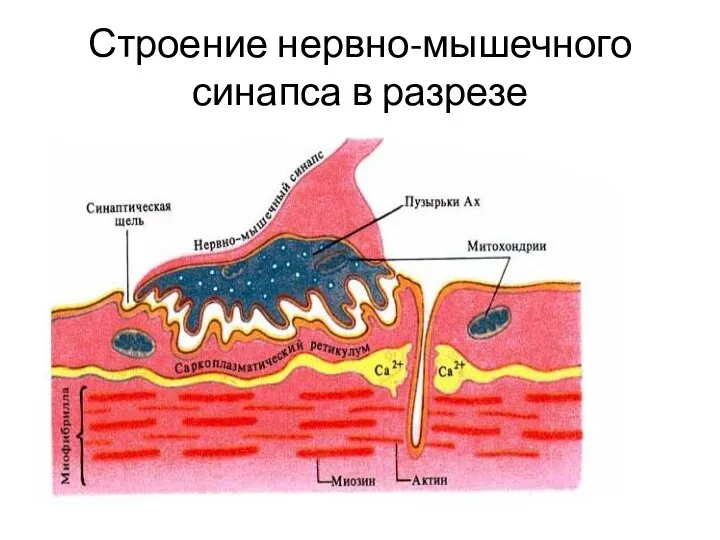
Строение нервно-мышечного синапса в разрезе
Строение нервно-мышечного синапса в разрезе
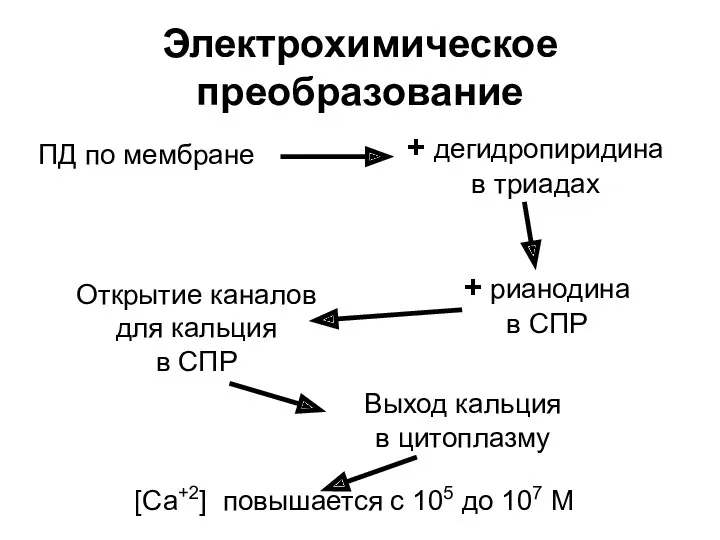
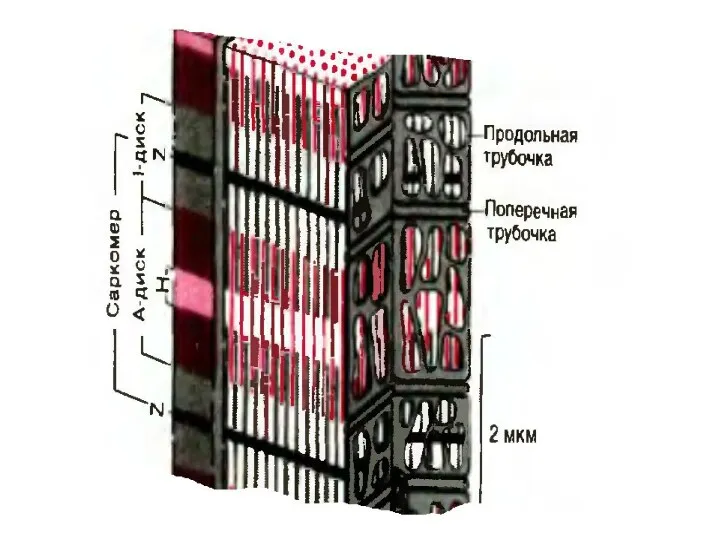
Электрохимическое преобразование
ПД по мембране
+ дегидропиридина
в триадах
+ рианодина
в СПР
Открытие каналов
для кальция
в
Электрохимическое преобразование
ПД по мембране
+ дегидропиридина
в триадах
+ рианодина
в СПР
Открытие каналов
для кальция
в

Без ПД кальций в цитоплазму не выйдет!!
Без ПД кальций в цитоплазму не выйдет!!
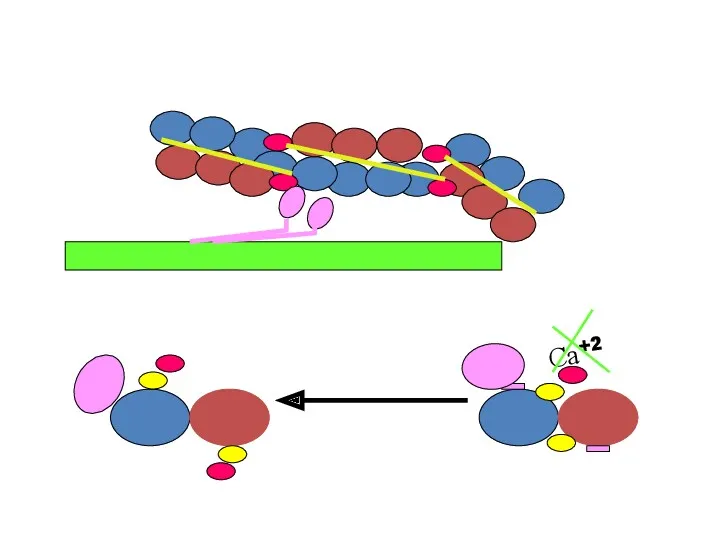
![Механизм расслабления мышцы [Са+2] 107 М + кальциевый насос в](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/261398/slide-21.jpg)
Механизм расслабления мышцы
[Са+2]
107 М
+ кальциевый насос в СПР
СПР
Са+2
Са+2
Са+2
АТФ
[Са+2]
>
<
105 М
Механизм расслабления мышцы
[Са+2]
107 М
+ кальциевый насос в СПР
СПР
Са+2
Са+2
Са+2
АТФ
[Са+2]
>
<
105 М
Са+2
Са+2

Затраты энергии АТФ во время мышечного сокращения
На поддержание ионной асимметрии (натрий-калиевй
Затраты энергии АТФ во время мышечного сокращения
На поддержание ионной асимметрии (натрий-калиевй

Двигательная или моторная единица
Каждое мышечное волокно имеет только один синапс
Двигательная или моторная единица
Каждое мышечное волокно имеет только один синапс

Двигательная или моторная единица
Мотонейрон и группа мышечных волокон, иннервируемых разветвлениями аксона
Двигательная или моторная единица
Мотонейрон и группа мышечных волокон, иннервируемых разветвлениями аксона
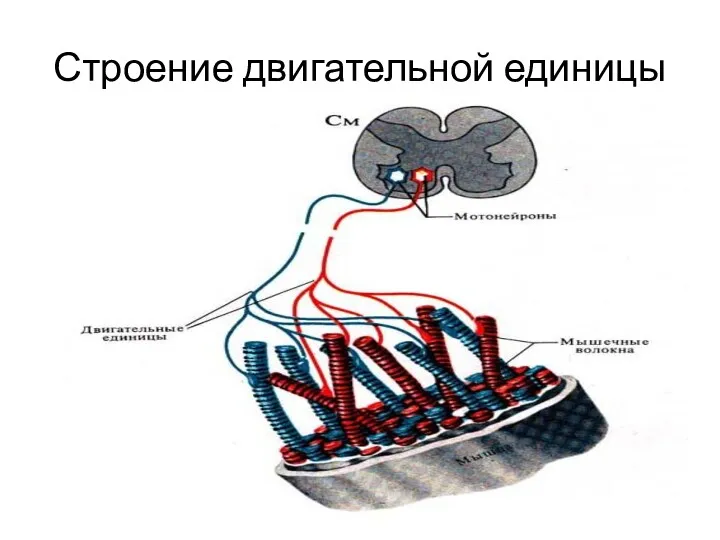
Строение двигательной единицы
Строение двигательной единицы
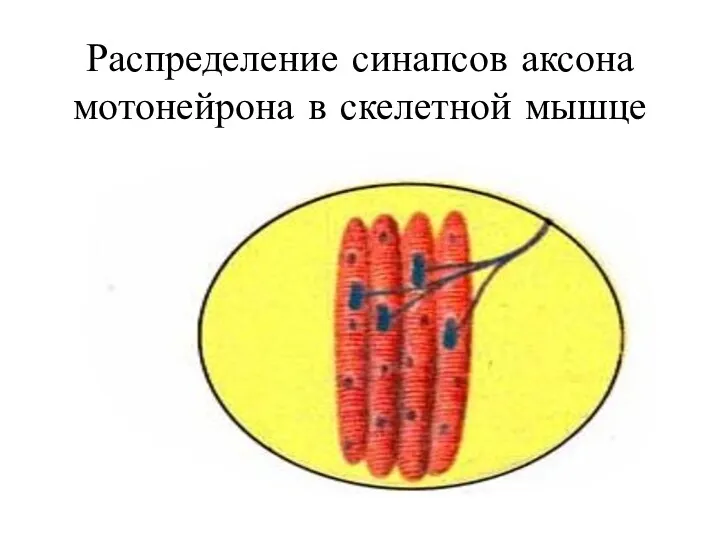
Распределение синапсов аксона мотонейрона в скелетной мышце
Распределение синапсов аксона мотонейрона в скелетной мышце

Классификация мышечных волокон
Классификация мышечных волокон

Медленные фазические волокна окислительного типа
большое содержание миоглобина и митохондрий
красного
Медленные фазические волокна окислительного типа
большое содержание миоглобина и митохондрий
красного

Быстрые фазические волокна окислительного типа
Содержат много митохондрий
Способны синтезировать АТФ путем окислительного
Быстрые фазические волокна окислительного типа
Содержат много митохондрий
Способны синтезировать АТФ путем окислительного

Быстрые фазические с гликолитическим типом окисления
Мало митохондрий
АТФ образуется за счет гликолиза
Миоглобина
Быстрые фазические с гликолитическим типом окисления
Мало митохондрий
АТФ образуется за счет гликолиза
Миоглобина

Тонические волокна
Двигательный аксон образует множество синапсов
Медленно сокращаются и медленно расслеабдяются
Низкая АТФ-азная
Тонические волокна
Двигательный аксон образует множество синапсов
Медленно сокращаются и медленно расслеабдяются
Низкая АТФ-азная

Закон «все или ничего»
Одиночное мышечное волокно подчиняется этому закону:
Подпороговое раздражение не
Закон «все или ничего»
Одиночное мышечное волокно подчиняется этому закону:
Подпороговое раздражение не

Закон «все или ничего»
Целая мышца данному закону не подчиняется потому, что
Закон «все или ничего»
Целая мышца данному закону не подчиняется потому, что

Режимы сокращения
Изометрический – увеличение напряжения без изменения длины мышцы
Изотонические – уменьшение
Режимы сокращения
Изометрический – увеличение напряжения без изменения длины мышцы
Изотонические – уменьшение

Виды мышечных сокращений
Одиночное
Тетанус
Тонус
Виды мышечных сокращений
Одиночное
Тетанус
Тонус

Виды раздражения мышцы в эксперименте
Непрямое раздражение – импульс воздействует на нервное
Виды раздражения мышцы в эксперименте
Непрямое раздражение – импульс воздействует на нервное
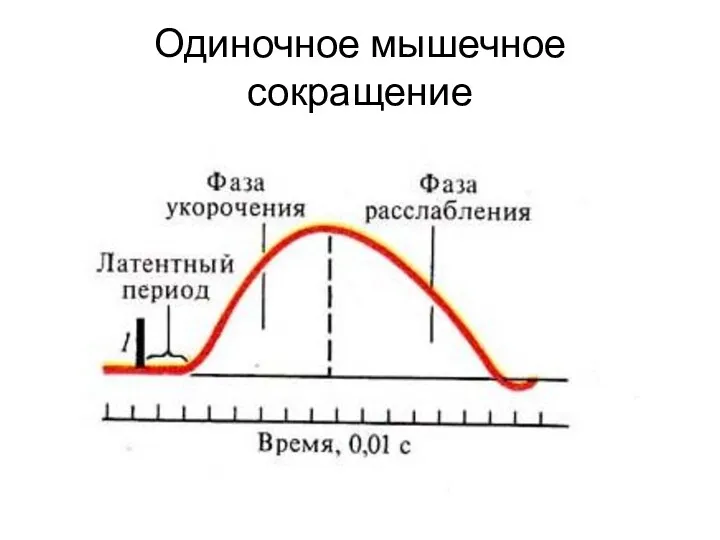
Одиночное мышечное сокращение
Одиночное мышечное сокращение
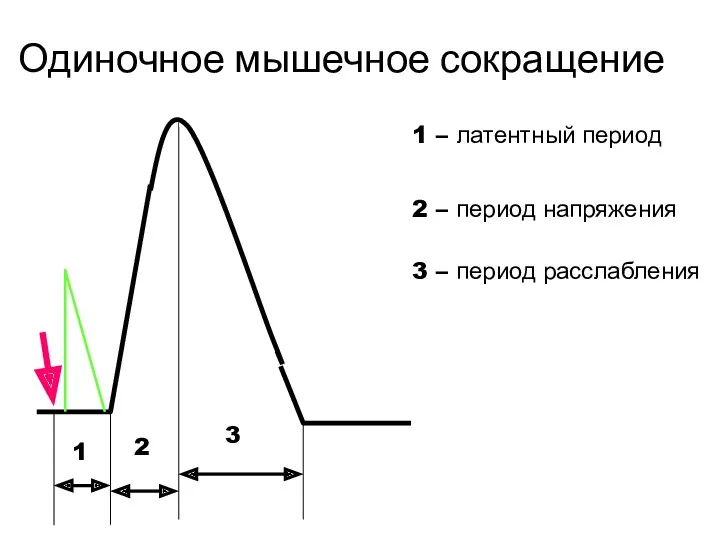
Одиночное мышечное сокращение
1
2
3
1 – латентный период
2 – период напряжения
3 – период
Одиночное мышечное сокращение
1
2
3
1 – латентный период
2 – период напряжения
3 – период

Тетанус
Это сильное и длительное сокращение мышцы в ответ на серию
Тетанус
Это сильное и длительное сокращение мышцы в ответ на серию
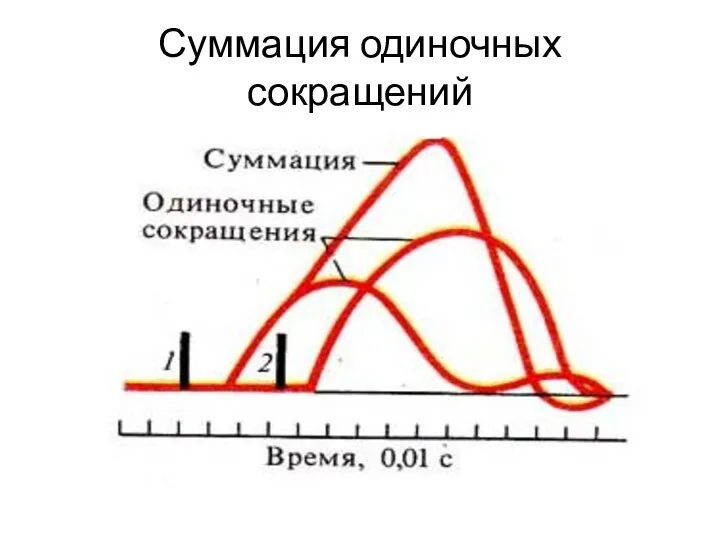
Суммация одиночных сокращений
Суммация одиночных сокращений

Зубчатый тетанус
Возникает в условиях когда каждый последующий импульс попадает в период
Зубчатый тетанус
Возникает в условиях когда каждый последующий импульс попадает в период



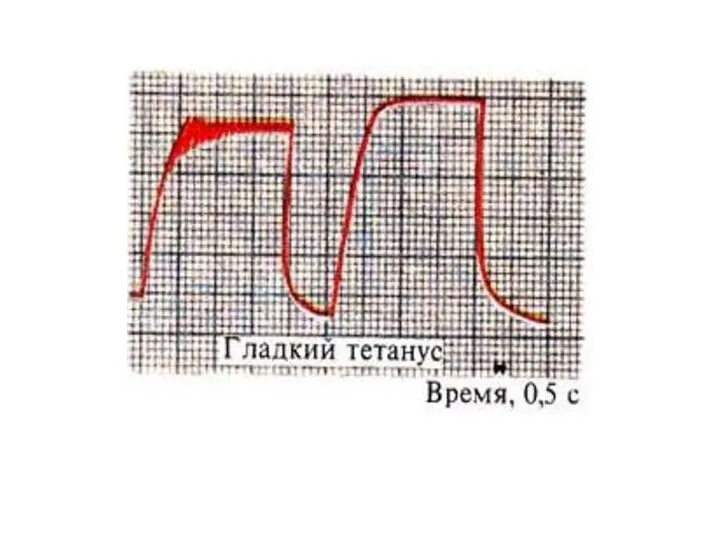
Гладкий тетанус
Возникает в условиях когда каждый последующий импульс попадает в период
Гладкий тетанус
Возникает в условиях когда каждый последующий импульс попадает в период
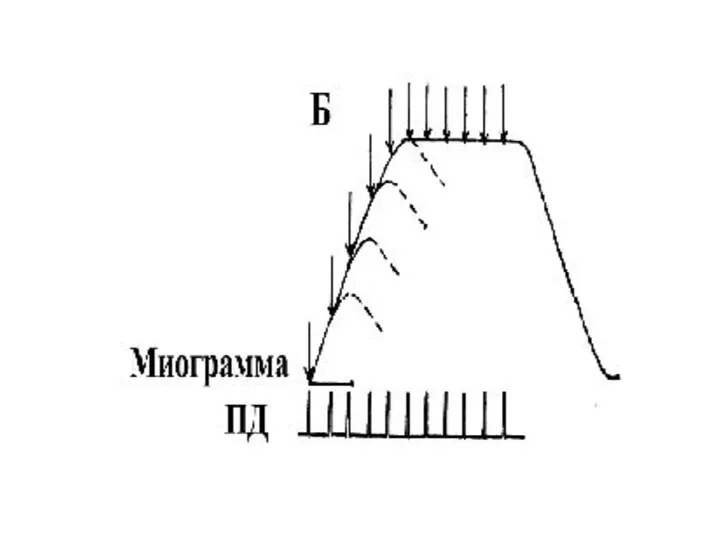

оптимум
Частота раздражения, при которой наблюдается суммарное сокращение (тетанус) наибольшей амплитуды
При этом
оптимум
Частота раздражения, при которой наблюдается суммарное сокращение (тетанус) наибольшей амплитуды
При этом

пессимум
Частота раздражения, при которй не наблюдается суммации сокращения
При этом каждый
пессимум
Частота раздражения, при которй не наблюдается суммации сокращения
При этом каждый

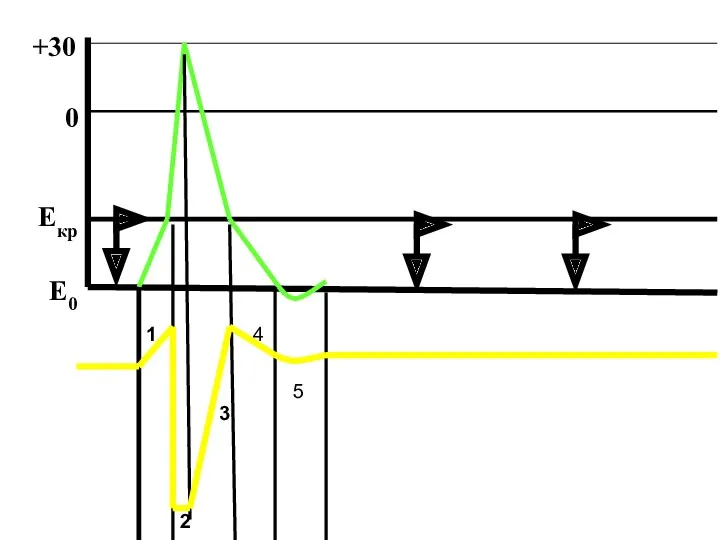
1
2
3
4
5
Е0
Екр
0
+30
1
2
3
4
5
Е0
Екр
0
+30
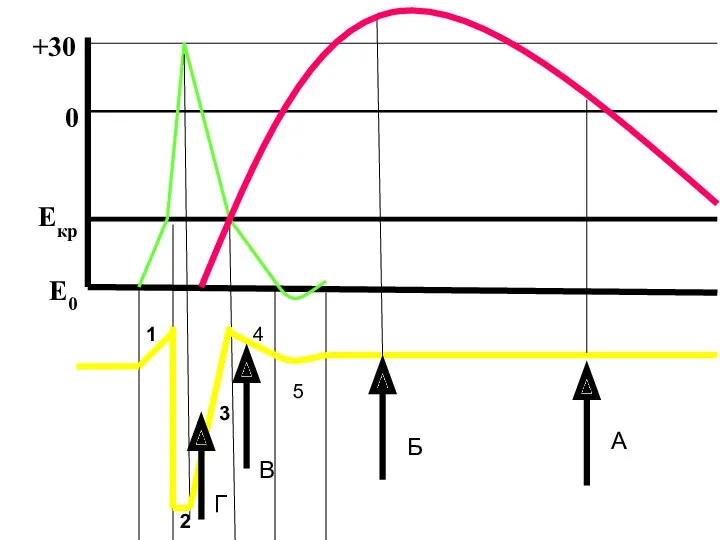
1
2
3
4
5
Е0
Екр
0
+30
В
Г
А
Б
1
2
3
4
5
Е0
Екр
0
+30
В
Г
А
Б
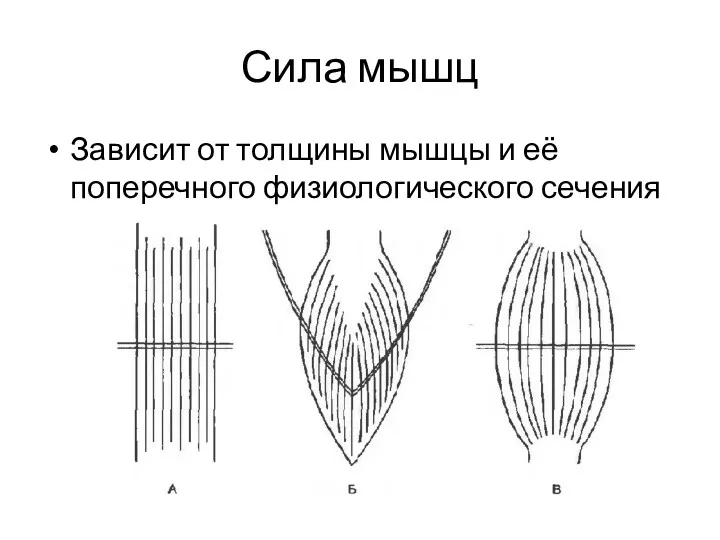
Сила мышц
Зависит от толщины мышцы и её поперечного физиологического сечения
Сила мышц
Зависит от толщины мышцы и её поперечного физиологического сечения

Работа мышцы
Это энергия, затрачиваемая на перемещение тела с определенной силой на
Работа мышцы
Это энергия, затрачиваемая на перемещение тела с определенной силой на

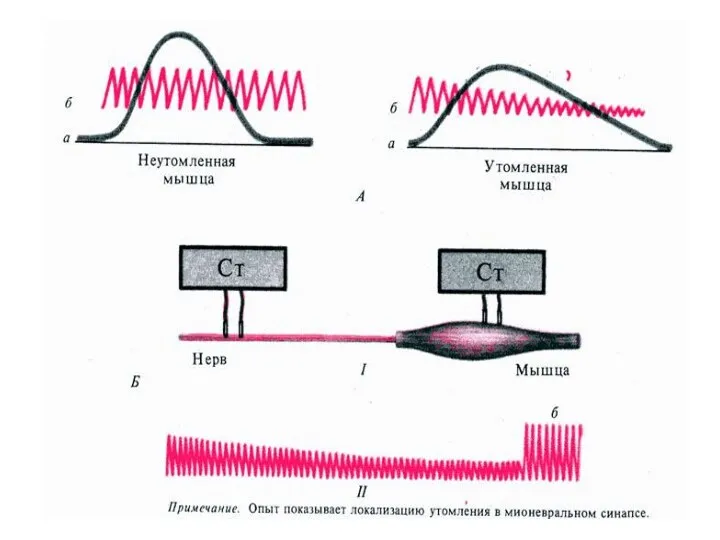
утомление
Процесс временного снижения работоспособности мышцы.
Возникает в связи с уменьшением энергетических запасов
утомление
Процесс временного снижения работоспособности мышцы.
Возникает в связи с уменьшением энергетических запасов

Утомление в организме
Утомление развивается вначале в нервных центрах – это защитный
Утомление в организме
Утомление развивается вначале в нервных центрах – это защитный

Гладкие мышцы
Гладкие мышцы
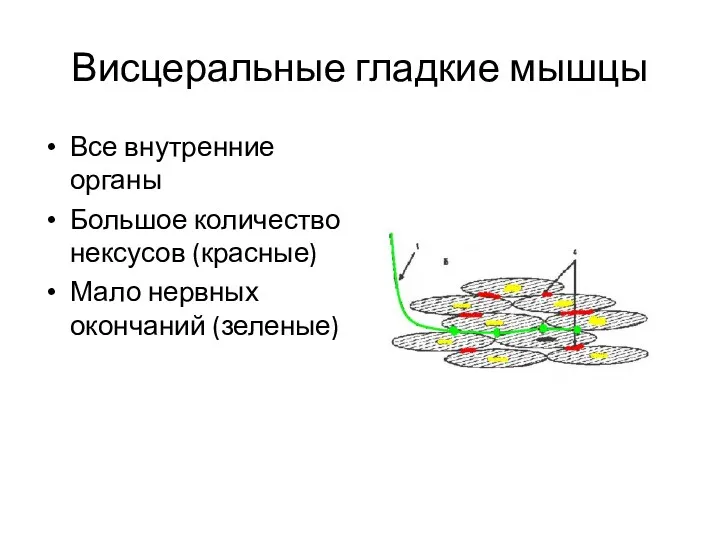
Висцеральные гладкие мышцы
Все внутренние органы
Большое количество нексусов (красные)
Мало нервных окончаний (зеленые)
Висцеральные гладкие мышцы
Все внутренние органы
Большое количество нексусов (красные)
Мало нервных окончаний (зеленые)
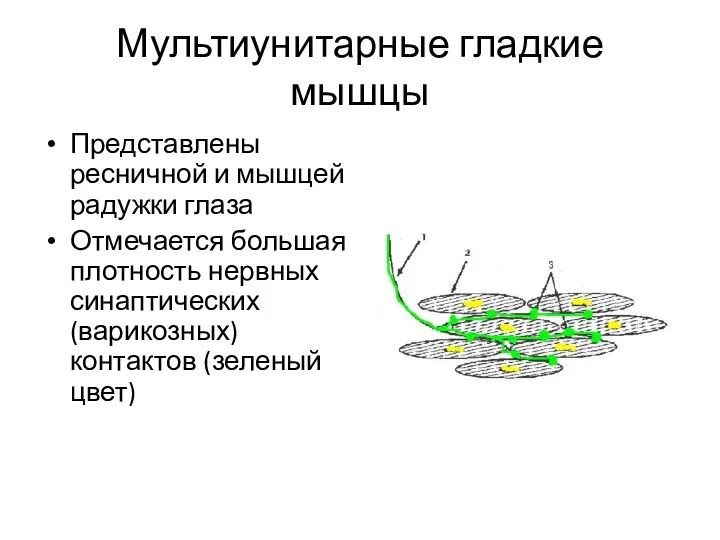
Мультиунитарные гладкие мышцы
Представлены ресничной и мышцей радужки глаза
Отмечается большая плотность нервных
Мультиунитарные гладкие мышцы
Представлены ресничной и мышцей радужки глаза
Отмечается большая плотность нервных
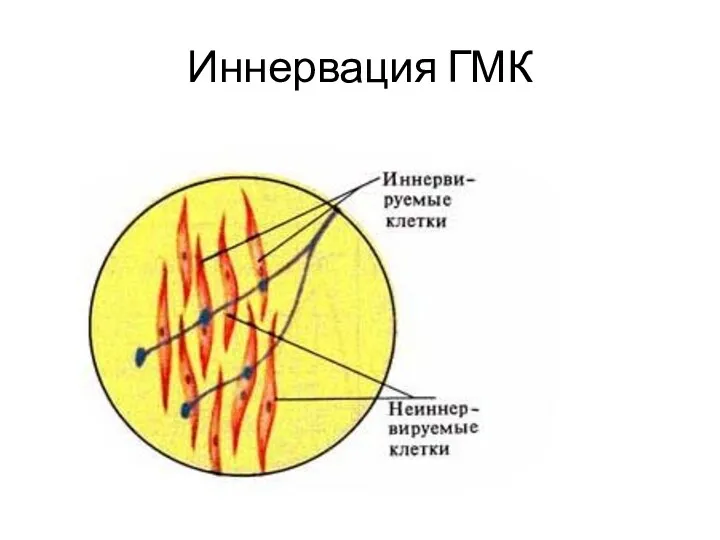
Иннервация ГМК
Иннервация ГМК

Особенности ГМК
Веретенообразные клетки, тесно прилежат друг к другу
Между клетками – плотные
Особенности ГМК
Веретенообразные клетки, тесно прилежат друг к другу
Между клетками – плотные

Иннервация ГМК
Иннервируют симпатические и парасимпатические нервные волокна
Нет синапсов, нервные окончания образуют
Иннервация ГМК
Иннервируют симпатические и парасимпатические нервные волокна
Нет синапсов, нервные окончания образуют

Адекватные раздражители
Нервный импульс
Механическое растяжение
Химические вещества
Адекватные раздражители
Нервный импульс
Механическое растяжение
Химические вещества

Особенности возбудимости
Порог возбудимости ниже, чем у поперечно-полосатых мышц
Мембрана более проницаема для
Особенности возбудимости
Порог возбудимости ниже, чем у поперечно-полосатых мышц
Мембрана более проницаема для

Автоматия
Способность клетки самостоятельно без внешнего раздражителя генерировать ПД
ГМК в состоянии покоя
Автоматия
Способность клетки самостоятельно без внешнего раздражителя генерировать ПД
ГМК в состоянии покоя

Электрохимическое сопряжение в ГМК
ПД открывает кальциевые каналы и в клетку входит
Электрохимическое сопряжение в ГМК
ПД открывает кальциевые каналы и в клетку входит
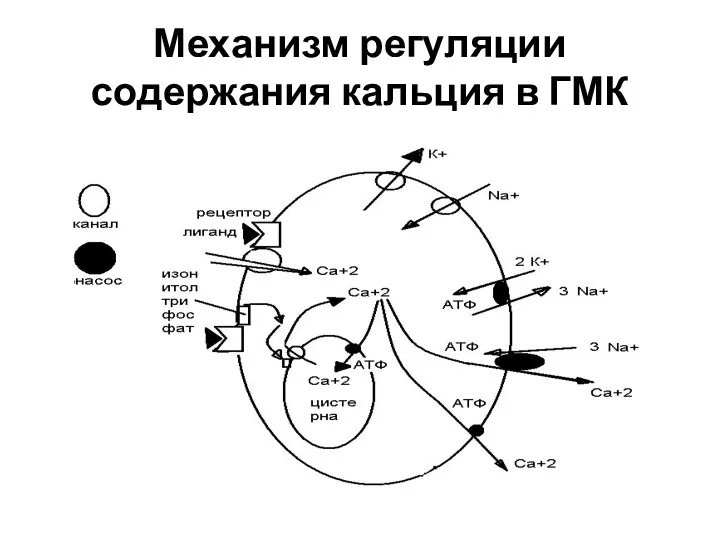
Механизм регуляции содержания кальция в ГМК
Механизм регуляции содержания кальция в ГМК

Особенности механизма сокращения в ГМК
Кальций в цитоплазме связывается с кальмодулином и
Особенности механизма сокращения в ГМК
Кальций в цитоплазме связывается с кальмодулином и
 Законы действия экологических факторов. Популяции
Законы действия экологических факторов. Популяции Мотивация. Биологическая мотивация. Общие свойства различных видов мотивации. Мотивация как доминанта
Мотивация. Биологическая мотивация. Общие свойства различных видов мотивации. Мотивация как доминанта Royal Botanic Gardens — Kew World Heritage in London
Royal Botanic Gardens — Kew World Heritage in London Матричный принцип, как основа современной эволюционной теории
Матричный принцип, как основа современной эволюционной теории Витамины
Витамины Строение глаза
Строение глаза Лекарственные растения луга
Лекарственные растения луга Как животные заботятся о своем потомстве. Как им помогает в этом Бог
Как животные заботятся о своем потомстве. Как им помогает в этом Бог Класс Пресмыкающиеся
Класс Пресмыкающиеся Витамины.
Витамины. Жабы и лягушки. 2 класс
Жабы и лягушки. 2 класс В царстве грибов
В царстве грибов Екологія мікроорганізмів. Мікрофлора організму людини. Мікрофлора ротової порожнини
Екологія мікроорганізмів. Мікрофлора організму людини. Мікрофлора ротової порожнини Интерактивный кроссворд Общее знакомство с цветковыми растениями
Интерактивный кроссворд Общее знакомство с цветковыми растениями Физиология микроорганизмов
Физиология микроорганизмов Матричный синтез информационных макромолекул. Экспрессия генетического материала
Матричный синтез информационных макромолекул. Экспрессия генетического материала Время посева и глубина заделки семян
Время посева и глубина заделки семян Аквариум и его обитатели
Аквариум и его обитатели Разнообразие природы Республики Татарстан
Разнообразие природы Республики Татарстан Система желудочков мозга
Система желудочков мозга Растительный мир Дальнего Востока
Растительный мир Дальнего Востока Дидактическая игра Эволюционное учение
Дидактическая игра Эволюционное учение Обмен углеводов. Функции углеводов
Обмен углеводов. Функции углеводов Урок биологии в 6 классе Внешнее и клеточное строение листа.
Урок биологии в 6 классе Внешнее и клеточное строение листа. Соматическая и вегетативная нервная система
Соматическая и вегетативная нервная система Хищные животные Хабаровского края
Хищные животные Хабаровского края Микробы (Окружающий мир, 3 класс)
Микробы (Окружающий мир, 3 класс) Питание микроорганизмов
Питание микроорганизмов