- Genetic load of human population

Содержание
- 2. Topic: Genetic load of human population Some Important Points:- Genetic load is the reduction in mean
- 3. genetic load Mutation that leads to lethal traits are often eliminated from the gene pool, however
- 4. Abstract Genetic load is the reduction in the mean fitness of a population relative to a
- 5. Genetic load is the difference between the fitness of an average genotype in a population and
- 6. Genetic load : Sources Mainly form three sources: 1.Mutational Load 2. Substitution load 3.Segregation load
- 7. Direct Evidence of an Increasing Mutational Load in Humans The extent to which selection has shaped
- 8. Mutational load is the total genetic burden in a population resulting from accumulated deleterious mutations. It
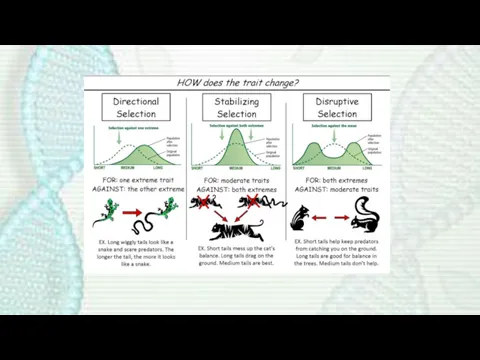
- 11. Directional selection occurs when individuals homozygous for one allele have a fitness greater than that of
- 12. When a dominant favored allele is rare most individuals carrying it are heterozygous, and the large
- 13. The Effects of Sexual Selection on the Heritability of Trait Strong directional selection usually exhausts additive
- 15. Substitutional load Substitutional load In genetics, the cost in genetic deaths to the population of replacing
- 16. Segregational or recombination load Segregation load is the presence of under dominant heterozygote's (i.e. heterozygote's that
- 17. CAUSES Some causes are :- Deleterious mutation Beneficial mutation Inbreeding
- 18. Deleterious mutation It is the main contributing factor to genetic load overall. Most mutations are neutral
- 19. Beneficial mutation New beneficial mutations create fitter genotypes than those previously present in the population. When
- 20. Inbreeding Inbreeding increases homozygosity In the short run, an increase in inbreeding increases the probability with
- 22. Скачать презентацию

Topic: Genetic load of human population
Some Important Points:-
Genetic load is the
Topic: Genetic load of human population
Some Important Points:-
Genetic load is the

genetic load
Mutation that leads to lethal traits are often
genetic load
Mutation that leads to lethal traits are often

Abstract
Genetic load is the reduction in the mean fitness of a
Abstract
Genetic load is the reduction in the mean fitness of a

Genetic load is the difference between the fitness of an average genotype in a population and the fitness
Genetic load is the difference between the fitness of an average genotype in a population and the fitness
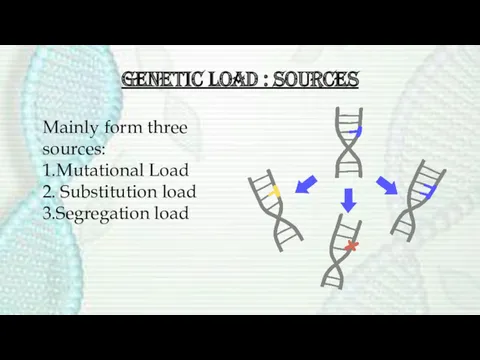
Genetic load : Sources
Mainly form three sources:
1.Mutational Load
2. Substitution load
3.Segregation load
Genetic load : Sources
Mainly form three sources:
1.Mutational Load
2. Substitution load
3.Segregation load

Direct Evidence of an Increasing Mutational Load in Humans
The extent to
Direct Evidence of an Increasing Mutational Load in Humans
The extent to

Mutational load is the total genetic burden in a population resulting from
Mutational load is the total genetic burden in a population resulting from



Directional selection occurs when individuals homozygous for one allele have a fitness
Directional selection occurs when individuals homozygous for one allele have a fitness
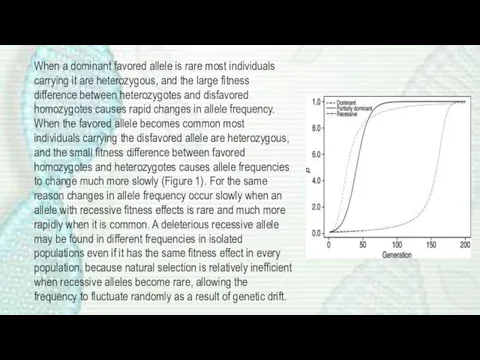
When a dominant favored allele is rare most individuals carrying it
When a dominant favored allele is rare most individuals carrying it

The Effects of Sexual Selection on the Heritability of Trait
Strong directional
The Effects of Sexual Selection on the Heritability of Trait
Strong directional

Substitutional load
Substitutional load In genetics, the cost in genetic deaths to the
Substitutional load
Substitutional load In genetics, the cost in genetic deaths to the

Segregational or recombination load
Segregation load is the presence of under dominant heterozygote's
Segregational or recombination load
Segregation load is the presence of under dominant heterozygote's

CAUSES
Some causes are :-
Deleterious mutation
Beneficial mutation
Inbreeding
CAUSES
Some causes are :-
Deleterious mutation
Beneficial mutation
Inbreeding

Deleterious mutation
It is the main contributing factor to genetic load overall. Most
Deleterious mutation
It is the main contributing factor to genetic load overall. Most

Beneficial mutation
New beneficial mutations create fitter genotypes than those previously
Beneficial mutation
New beneficial mutations create fitter genotypes than those previously

Inbreeding
Inbreeding increases homozygosity In the short run, an increase in inbreeding
Inbreeding
Inbreeding increases homozygosity In the short run, an increase in inbreeding
 Комнатные растения в интерьере
Комнатные растения в интерьере Cellular neurophysiology
Cellular neurophysiology Зеленые водоросли
Зеленые водоросли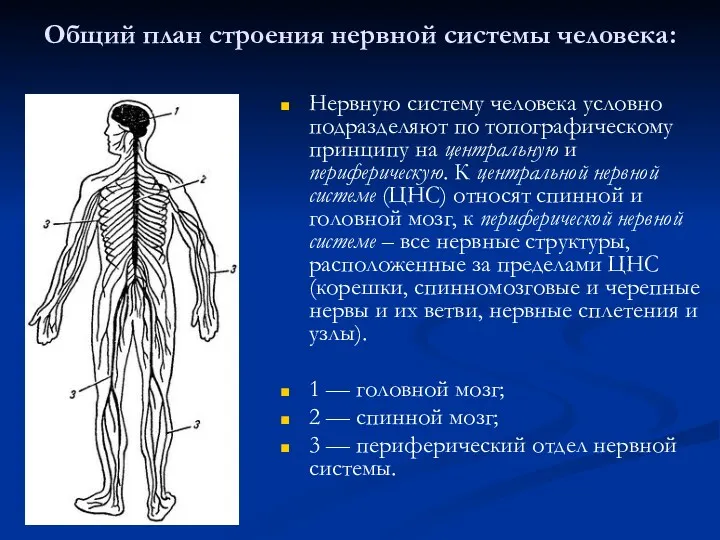 Спинной мозг человека
Спинной мозг человека Апоптоз. Исследования феномена программируемой клеточной смерти
Апоптоз. Исследования феномена программируемой клеточной смерти Ознакомление с фитонцидными растениями и выявление возможности их использования в интерьере. Практическая работа
Ознакомление с фитонцидными растениями и выявление возможности их использования в интерьере. Практическая работа Ритмы здоровья. Витаминно-минеральный комплекс
Ритмы здоровья. Витаминно-минеральный комплекс Чарлз Дарвин об эволюции органического мира
Чарлз Дарвин об эволюции органического мира урок биологии 6 класс по теме Побег
урок биологии 6 класс по теме Побег Эволюция человека
Эволюция человека Корова – домашнее и сельскохозяйственное животное
Корова – домашнее и сельскохозяйственное животное Онтогенез. Размножение организмов
Онтогенез. Размножение организмов Выращивание томата в теплице и парнике. 6 класс
Выращивание томата в теплице и парнике. 6 класс Подготовка к ЕГЭ. Решение генетических задач по схемам родословных
Подготовка к ЕГЭ. Решение генетических задач по схемам родословных Плоды и семена
Плоды и семена Приобретенные формы поведения. (Лекция 3)
Приобретенные формы поведения. (Лекция 3) Презентация по теме Основные типы экологических взаимодействий
Презентация по теме Основные типы экологических взаимодействий Новосибирский зоопарк
Новосибирский зоопарк Отдел Злаковые
Отдел Злаковые Клеточная оболочка. Оболочка животных и растительных клеток
Клеточная оболочка. Оболочка животных и растительных клеток Тваринний світ України
Тваринний світ України Органические вещества клетки – нуклеиновые кислоты ДНК и РНК
Органические вещества клетки – нуклеиновые кислоты ДНК и РНК Начальные этапы развития жизни на Земле
Начальные этапы развития жизни на Земле Synecology or - Community Ecology лекция
Synecology or - Community Ecology лекция Опасные дикие животные Якутии (Волк). 6 класс
Опасные дикие животные Якутии (Волк). 6 класс Методы исследования генетики человека
Методы исследования генетики человека Нервная система, ее структурно-функциональная характеристика. Центральная нервная система
Нервная система, ее структурно-функциональная характеристика. Центральная нервная система Клеточная теория. Цитология
Клеточная теория. Цитология