- Генетика мікроорганізмів

Содержание
- 2. Использование множественно ауксотрофных штаммов для, демонстрации скрещивания у Е. coli.
- 3. Життєві цикли. Процеси, що ведуть до рекомбінації в бактерій і бактеріофагів Між процесами, що ведуть до
- 4. Схема будови прокаріотичної клітини, що ділиться: КС - клітинна стінка, КМ - клітинна мембрана, Н -
- 5. Кон'югація Кон'югацією називається безпосередній контакт між клітинами бактерій, супроводжуваний переносом генетичного матеріалу із клітин донора в
- 6. Контроль статевого диференціювання. Епісоми і плазміди. Виявилося, що серед штамів Е. coli існує різноманітність по частоті
- 7. Таким чином, F-фактор може існувати у двох станах: автономному й інтегрованому. Такі генетичні детермінанти одержали найменування
- 8. Электронная микрофотография клеток F- Е. соli(справа) и F+ (слева). F-пили можно отличить от пилей другого типа,
- 9. Кон'югація між клітинами F (Hfr — довгаста) і b (кругла) Е. coli
- 10. Схематическое изображение механизма, с помощью которого ДНК F-фактора передастся от F + -клетки F- -клетке в
- 11. Мобилизация бактериальной хромосомы при включении F-фактора в клетке Hfr. При конъюгации репликация хромосомы начинается с встроенного
- 12. Опыт Ф. Жакоба и Е. Вольмана по прерыванию конъюгации А — схема эксперимента; Б — зависимость
- 13. Схема, иллюстрирующая полярность переноса Hfr-хромосомы в эксперименте с прерванной конъюгацией. К рекомбинации с хромосомой F- -клетки
- 14. Сайты интеграции F-фактора в хромосому некоторых Hfr-штаммов Е. coli
- 15. Генетична карта E. coli
- 16. Реплікація при кон'югації Взаємини F-епісоми і бактеріальної хромосоми можна представити в такий спосіб. У клітинах F–
- 17. F'-фактори й сексдукція Вирізання (ексцизія) F-фактору із хромосоми Hfr -штаму іноді буває неточним, і тоді ділянка
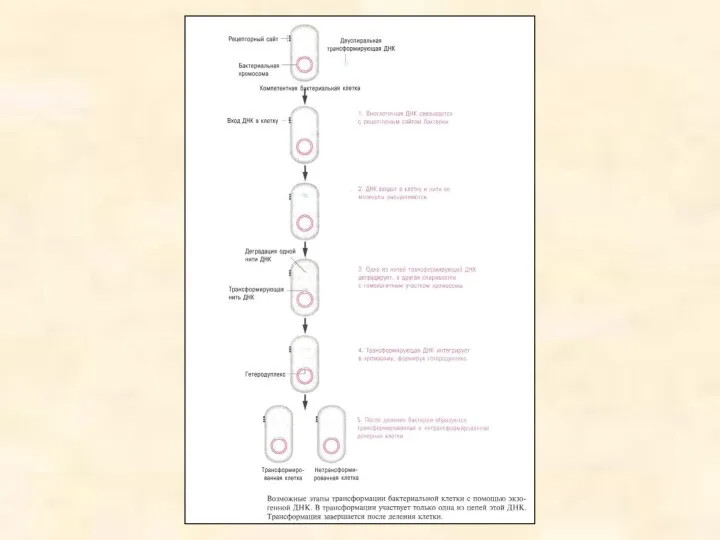
- 18. Трансформація Трансформація бактерій — це перенос ДНК, ізольованої з одних клітин в інші. Дослідження на пневмококах,
- 19. Схема досліду, що демонструє явище трансформації: Штами бактерій Sms – чутливі до стрептоміцину Smr - нечутливі
- 20. Схема дослідів Евірі, Мак-Леода і Мак-Карті по трансформації пневмококів
- 21. Механізм бактеріальної трансформації У результаті трансформації відбувається генетична рекомбінація між бактеріальними клітинами. Невеликі фрагментипозаклітинної ДНК потрапляють
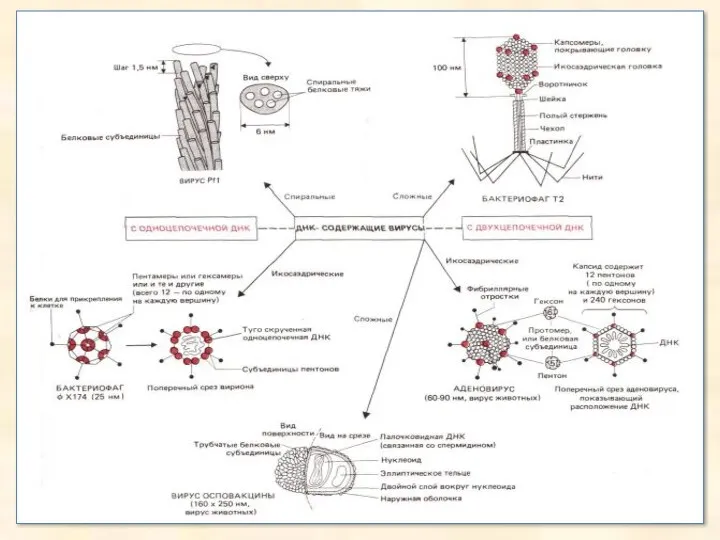
- 23. Віруси Віруси-частки, що містять нуклеїнові кислоти, білки, а іноді й ліпіди й здатні розмножуватися лише в
- 24. Электронные микрофотографии различных вирусов. Верхний ряд: РНК-вирусы полиомиелита (увеличение х 115 000); табачной мозаики ( х
- 26. Інфікування — процес, за допомогою якого вірус впроваджується в клітину-хазяїна й «набудовує» її метаболічний апарат на
- 27. Узагальнена схема розмноження ДНК вірусів
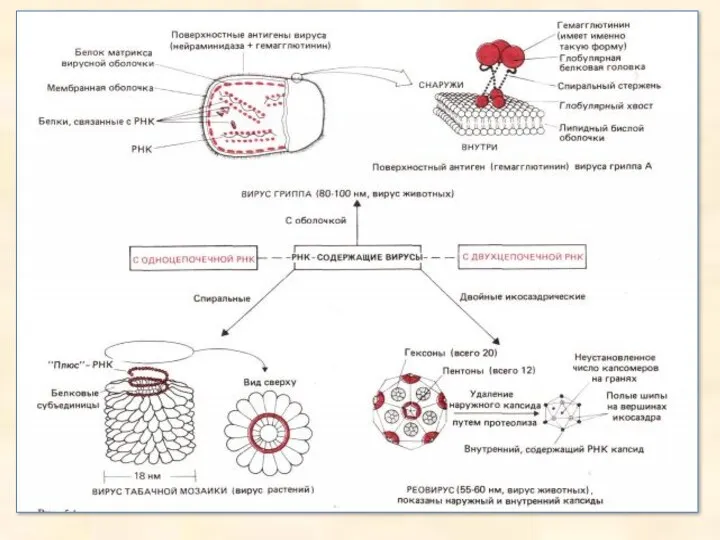
- 28. РНК-містячі віруси не мають ДНК: генетична інформація цих вірусів закодована в РНК. РНК може бути одне-
- 30. Узагальнена схема розмноження РНК вірусів
- 31. Вірус імунодефіциту людини
- 32. Схема будови вірусу імунодефіциту людини
- 37. Трансдукція Трансдукцією називають переніс генів з одних бактеріальних клітин в інші за допомогою бактеріофага. Вірулентні й
- 38. Схема досліду, що демонструє явище трансдукції у сальмонели: Штами бактерій 22А – які нездатні синтезувати триптофан
- 39. Загальна, або неспецифічна, трансдукція Трансдукцію здійснюють помірні бактеріофаги. До їхнього числа ставиться й фаг Р22, за
- 40. Специфічна трансдукція. Сайт-специфічна рекомбінація Специфічна трансдукція відрізняється від неспецифічної тим, що бактеріофаг може переносити тільки певні
- 41. Відносини помірного бактеріофага й бактеріальної клітини А и Б — літичний цикл; В — лізогенізація; Г
- 42. Перенос ДНК из донорной бактериальной клетки в реципиентную в результате трансдукции фагом Р1. В популяции трансдуцирующих
- 43. Схема інтеграції й ексцизії геному бактеріофага λ за рахунок сайт-специфічної рекомбінації із хромосомою Е. coli і
- 44. Генетичний аналіз у бактерій При кон'югації в клітину-реципієнт переноситься звичайно менше половини бактеріального геному, і дуже
- 46. Скачать презентацию
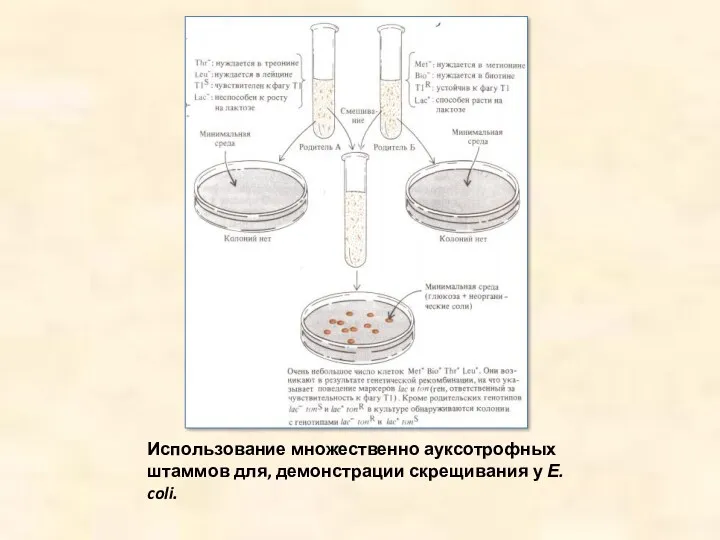
Использование множественно ауксотрофных штаммов для, демонстрации скрещивания у Е. coli.
Использование множественно ауксотрофных штаммов для, демонстрации скрещивания у Е. coli.

Життєві цикли. Процеси, що ведуть до рекомбінації в бактерій і бактеріофагів
Між
Життєві цикли. Процеси, що ведуть до рекомбінації в бактерій і бактеріофагів
Між
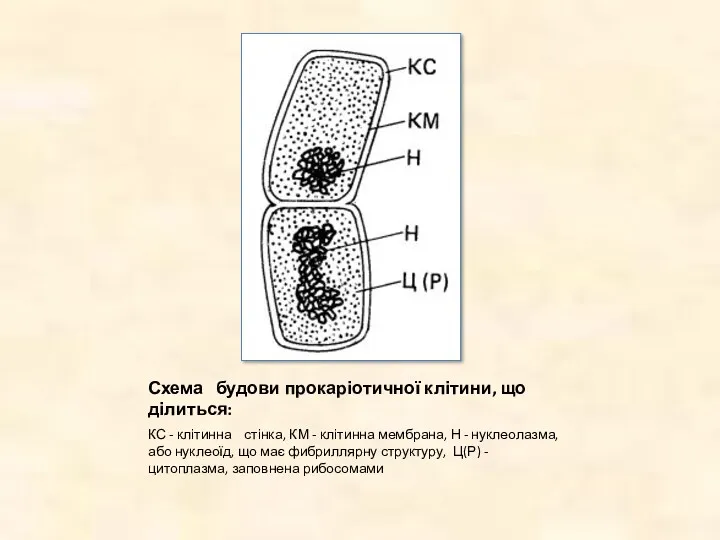
Схема будови прокаріотичної клітини, що ділиться:
КС - клітинна стінка, КМ
Схема будови прокаріотичної клітини, що ділиться:
КС - клітинна стінка, КМ

Кон'югація
Кон'югацією називається безпосередній контакт між клітинами бактерій, супроводжуваний переносом генетичного матеріалу
Кон'югація
Кон'югацією називається безпосередній контакт між клітинами бактерій, супроводжуваний переносом генетичного матеріалу

Контроль статевого диференціювання. Епісоми і плазміди.
Виявилося, що серед штамів Е. coli
Контроль статевого диференціювання. Епісоми і плазміди.
Виявилося, що серед штамів Е. coli

Таким чином, F-фактор може існувати у двох станах: автономному й інтегрованому.
Таким чином, F-фактор може існувати у двох станах: автономному й інтегрованому.
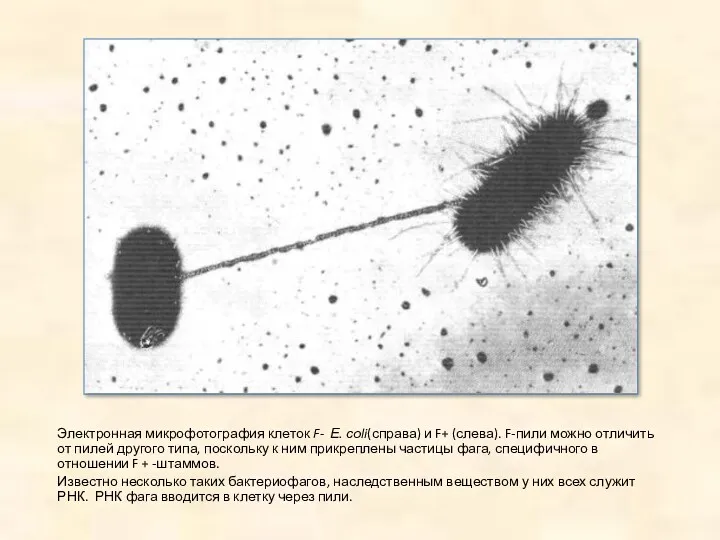
Электронная микрофотография клеток F- Е. соli(справа) и F+ (слева). F-пили можно
Электронная микрофотография клеток F- Е. соli(справа) и F+ (слева). F-пили можно

Кон'югація між клітинами F (Hfr — довгаста) і b (кругла) Е.
Кон'югація між клітинами F (Hfr — довгаста) і b (кругла) Е.
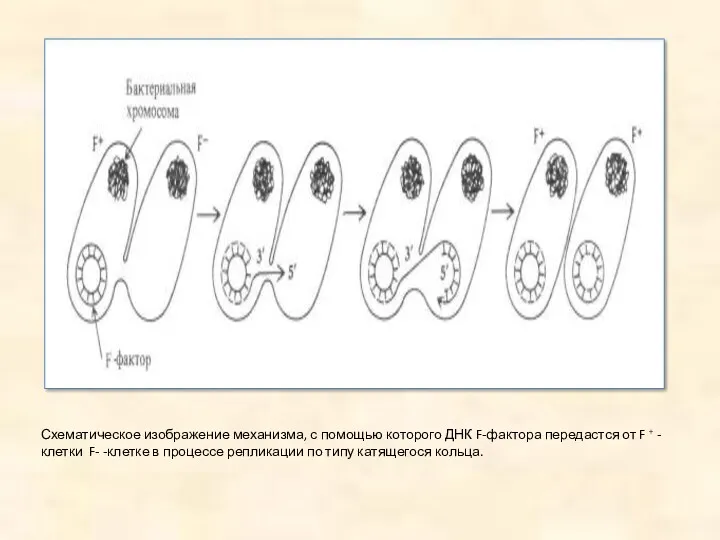
Схематическое изображение механизма, с помощью которого ДНК F-фактора передастся от F
Схематическое изображение механизма, с помощью которого ДНК F-фактора передастся от F
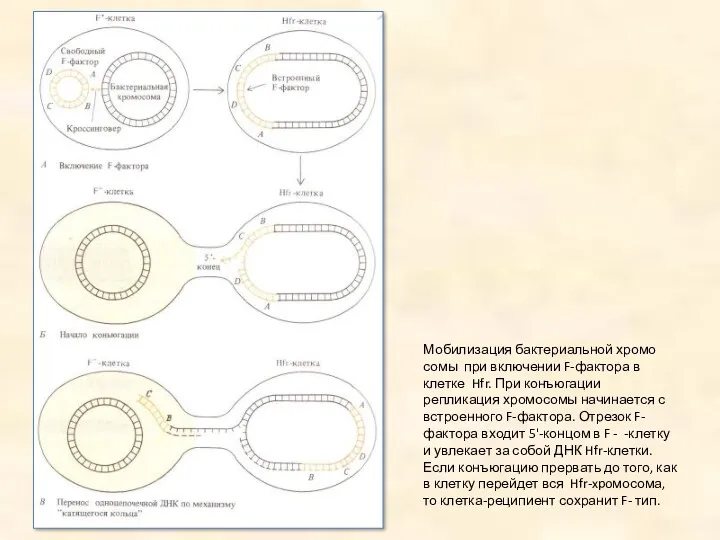
Мобилизация бактериальной хромосомы при включении F-фактора в клетке Hfr. При конъюгации
Мобилизация бактериальной хромосомы при включении F-фактора в клетке Hfr. При конъюгации
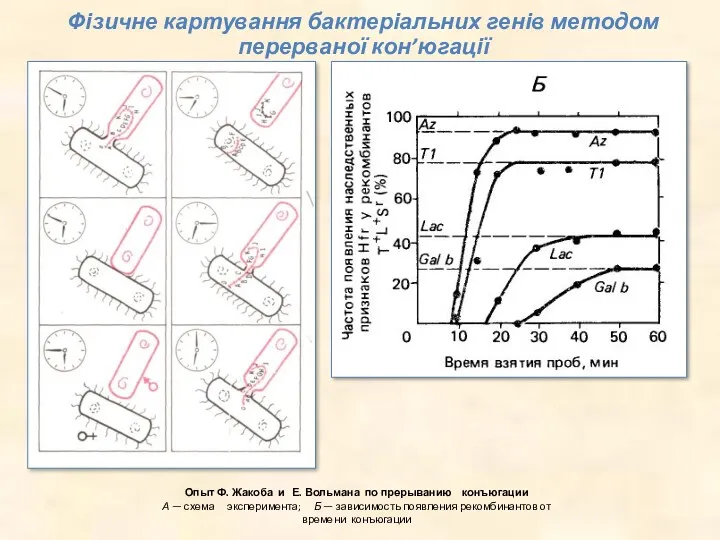
Опыт Ф. Жакоба и Е. Вольмана по прерыванию конъюгации
А — схема
Опыт Ф. Жакоба и Е. Вольмана по прерыванию конъюгации А — схема
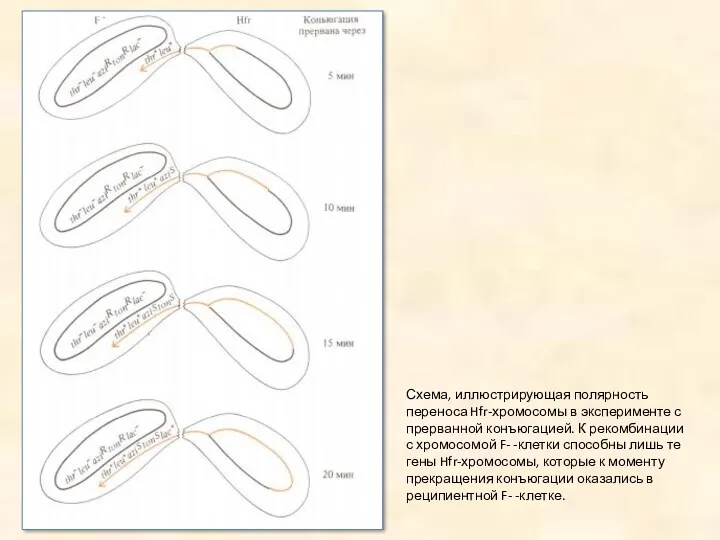
Схема, иллюстрирующая полярность переноса Hfr-хромосомы в эксперименте с прерванной конъюгацией. К
Схема, иллюстрирующая полярность переноса Hfr-хромосомы в эксперименте с прерванной конъюгацией. К
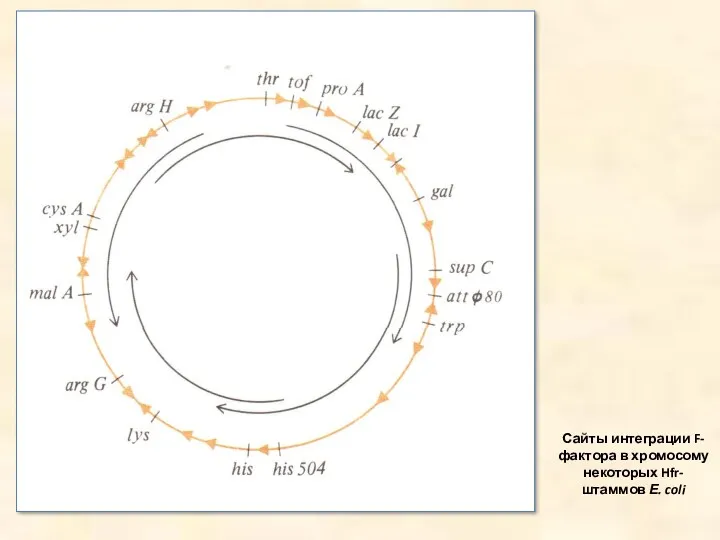
Сайты интеграции F-фактора в хромосому некоторых Hfr-штаммов Е. coli
Сайты интеграции F-фактора в хромосому некоторых Hfr-штаммов Е. coli
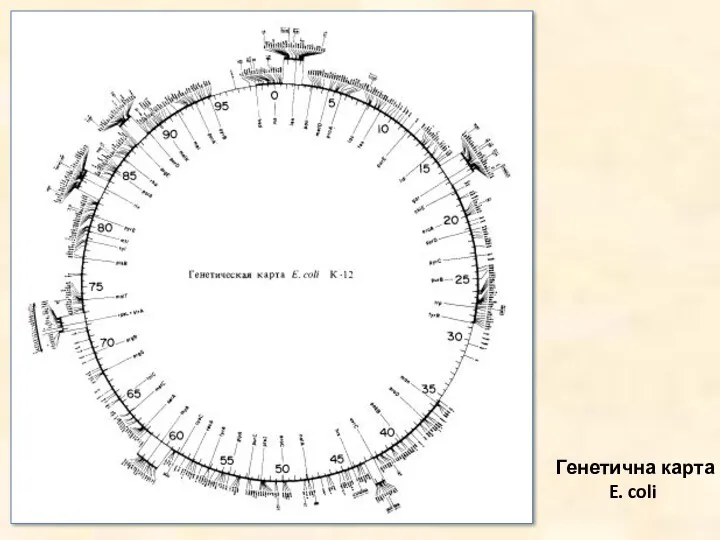
Генетична карта E. coli
Генетична карта E. coli

Реплікація при кон'югації
Взаємини F-епісоми і бактеріальної хромосоми можна представити в такий
Реплікація при кон'югації
Взаємини F-епісоми і бактеріальної хромосоми можна представити в такий

F'-фактори й сексдукція
Вирізання (ексцизія) F-фактору із хромосоми Hfr -штаму іноді буває
F'-фактори й сексдукція
Вирізання (ексцизія) F-фактору із хромосоми Hfr -штаму іноді буває

Трансформація
Трансформація бактерій — це перенос ДНК, ізольованої з одних клітин в
Трансформація
Трансформація бактерій — це перенос ДНК, ізольованої з одних клітин в
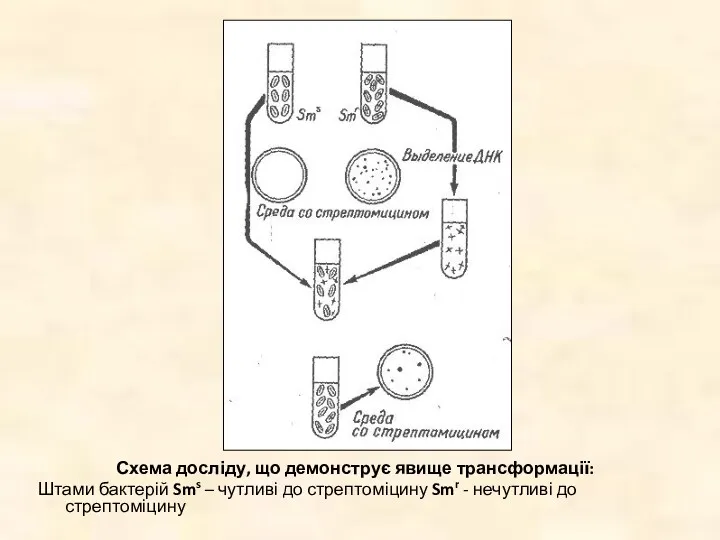
Схема досліду, що демонструє явище трансформації:
Штами бактерій Sms – чутливі до
Схема досліду, що демонструє явище трансформації:
Штами бактерій Sms – чутливі до
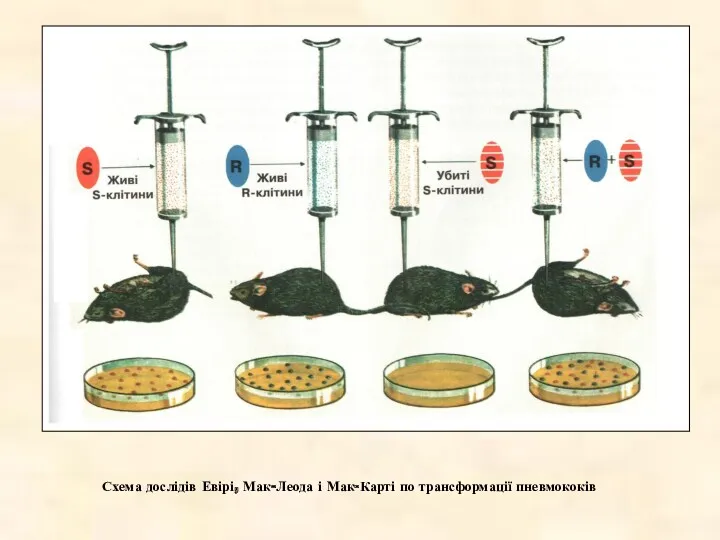
Схема дослідів Евірі, Мак-Леода і Мак-Карті по трансформації пневмококів
Схема дослідів Евірі, Мак-Леода і Мак-Карті по трансформації пневмококів

Механізм бактеріальної трансформації
У результаті трансформації відбувається генетична рекомбінація між бактеріальними клітинами. Невеликі фрагментипозаклітинної ДНК потрапляють у бактерію й забезпечують клітині-реципієнту нові
Механізм бактеріальної трансформації
У результаті трансформації відбувається генетична рекомбінація між бактеріальними клітинами. Невеликі фрагментипозаклітинної ДНК потрапляють у бактерію й забезпечують клітині-реципієнту нові

Віруси
Віруси-частки, що містять нуклеїнові кислоти, білки, а іноді й ліпіди й
Віруси
Віруси-частки, що містять нуклеїнові кислоти, білки, а іноді й ліпіди й

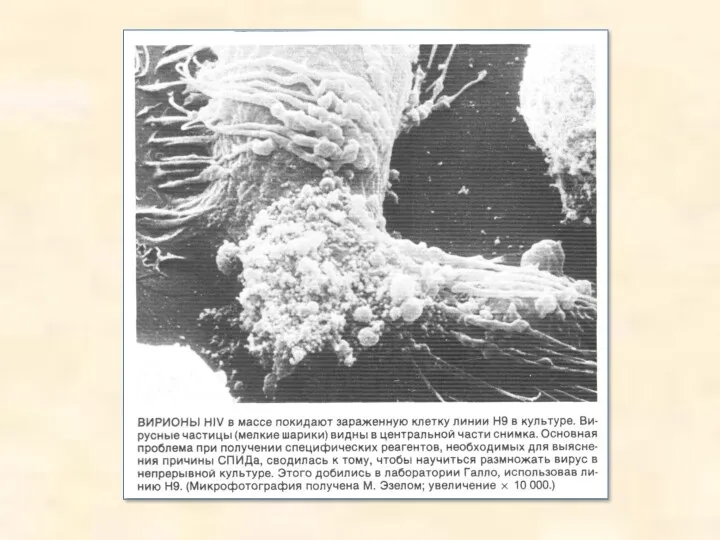
Электронные микрофотографии различных вирусов. Верхний ряд: РНК-вирусы полиомиелита (увеличение х 115
Электронные микрофотографии различных вирусов. Верхний ряд: РНК-вирусы полиомиелита (увеличение х 115

Інфікування — процес, за допомогою якого вірус впроваджується в клітину-хазяїна й
Інфікування — процес, за допомогою якого вірус впроваджується в клітину-хазяїна й
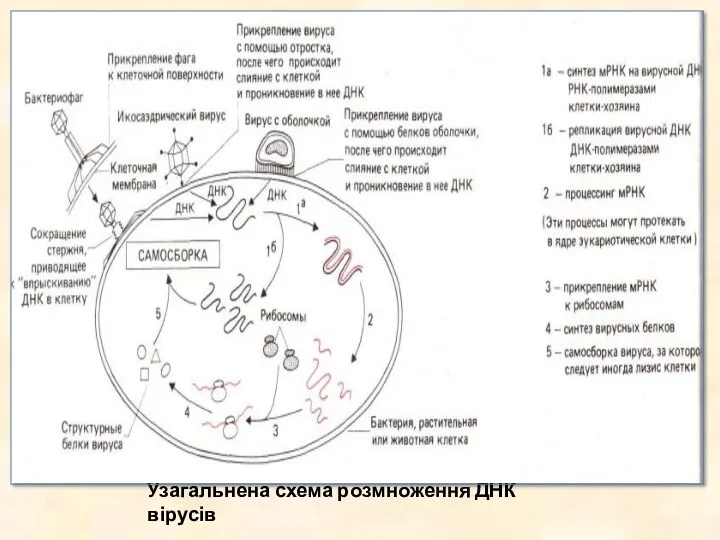
Узагальнена схема розмноження ДНК вірусів
Узагальнена схема розмноження ДНК вірусів

РНК-містячі віруси не мають ДНК: генетична інформація цих вірусів закодована в
РНК-містячі віруси не мають ДНК: генетична інформація цих вірусів закодована в
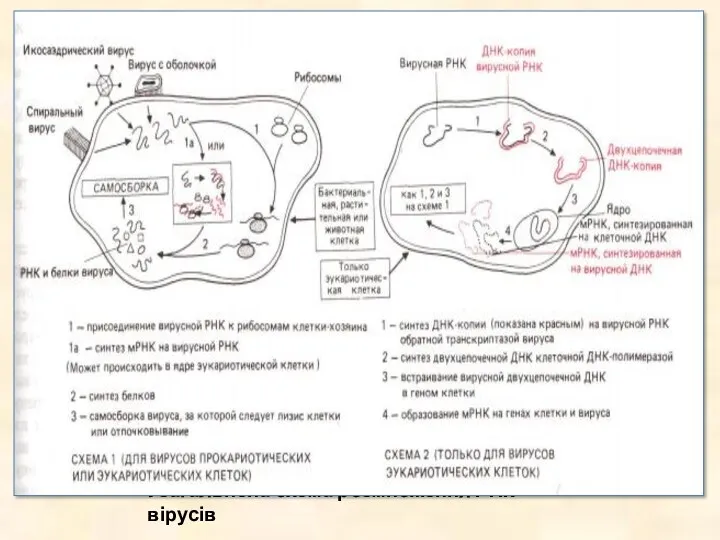
Узагальнена схема розмноження РНК вірусів
Узагальнена схема розмноження РНК вірусів
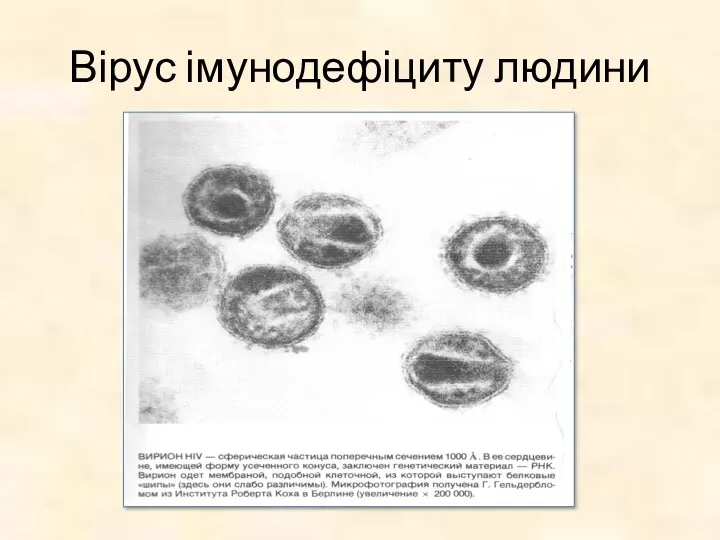
Вірус імунодефіциту людини
Вірус імунодефіциту людини
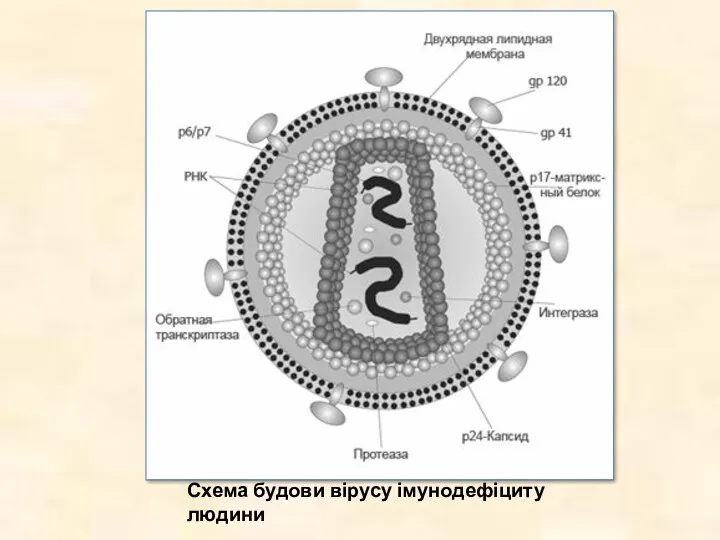
Схема будови вірусу імунодефіциту людини
Схема будови вірусу імунодефіциту людини
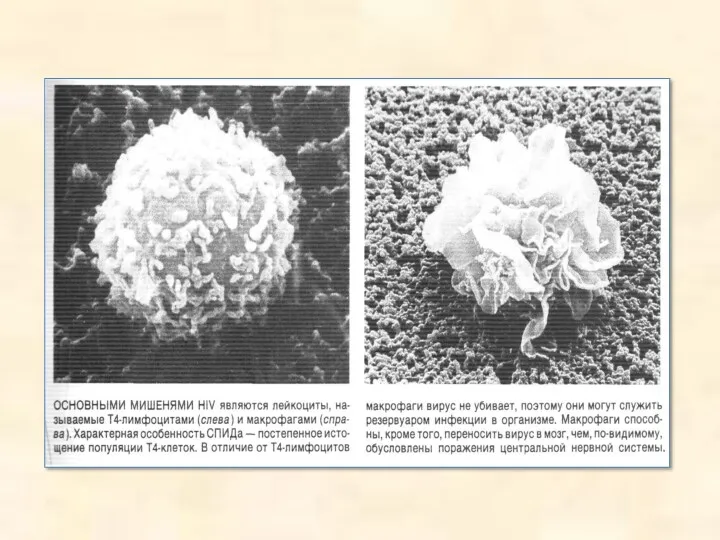
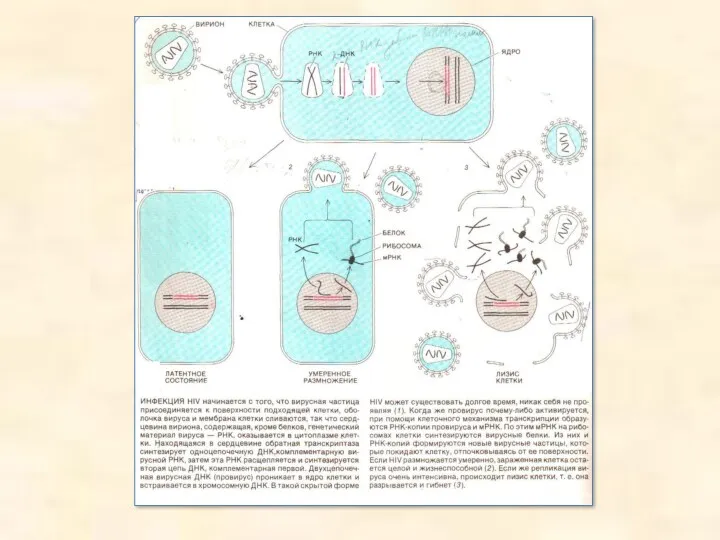
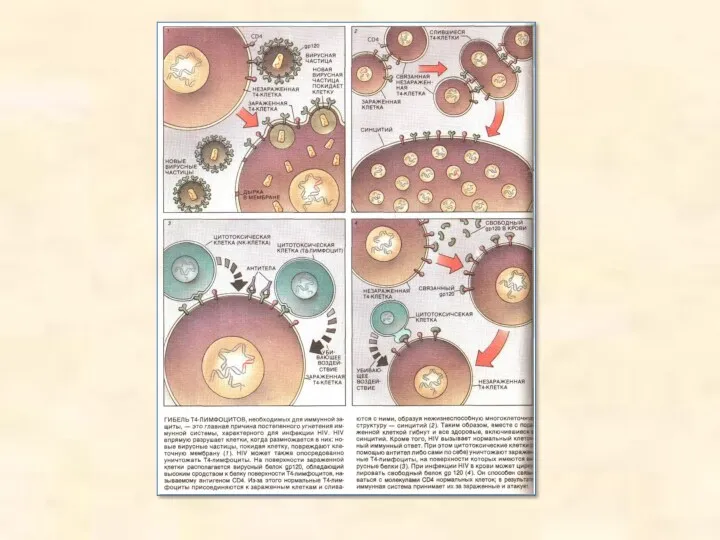

Трансдукція
Трансдукцією називають переніс генів з одних бактеріальних клітин в інші за
Трансдукція
Трансдукцією називають переніс генів з одних бактеріальних клітин в інші за
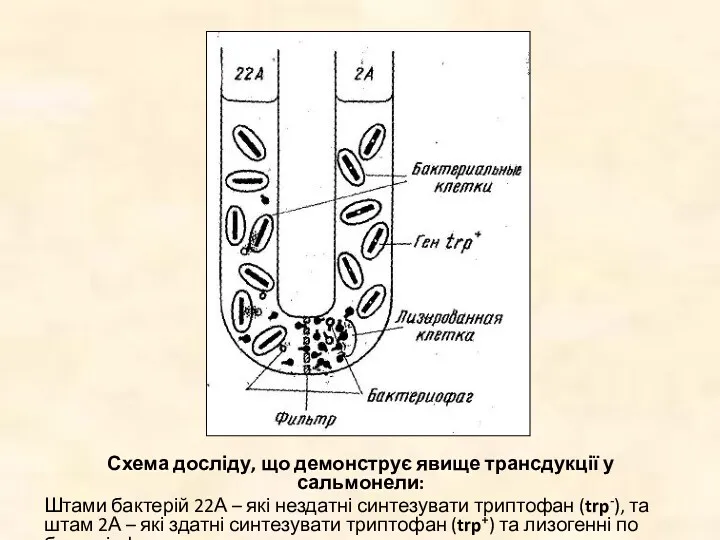
Схема досліду, що демонструє явище трансдукції у сальмонели:
Штами бактерій 22А –
Схема досліду, що демонструє явище трансдукції у сальмонели:
Штами бактерій 22А –

Загальна, або неспецифічна, трансдукція
Трансдукцію здійснюють помірні бактеріофаги. До їхнього числа ставиться
Загальна, або неспецифічна, трансдукція
Трансдукцію здійснюють помірні бактеріофаги. До їхнього числа ставиться

Специфічна трансдукція. Сайт-специфічна рекомбінація
Специфічна трансдукція відрізняється від неспецифічної тим, що бактеріофаг
Специфічна трансдукція. Сайт-специфічна рекомбінація
Специфічна трансдукція відрізняється від неспецифічної тим, що бактеріофаг
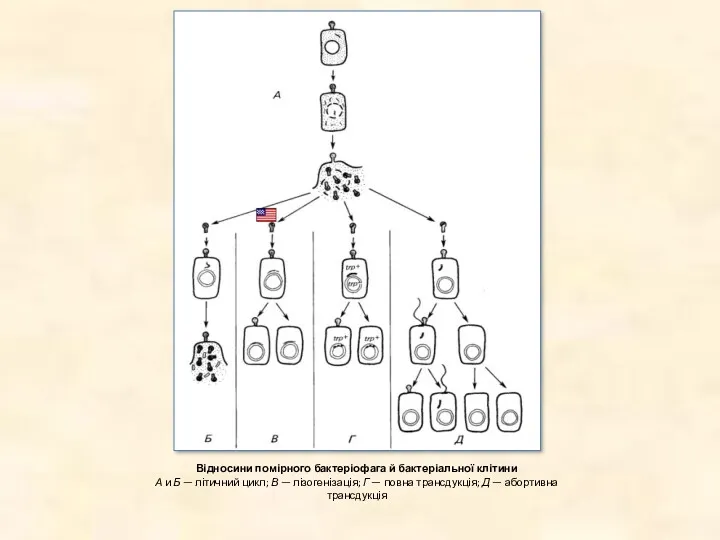
Відносини помірного бактеріофага й бактеріальної клітини
А и Б — літичний цикл;
Відносини помірного бактеріофага й бактеріальної клітини А и Б — літичний цикл;
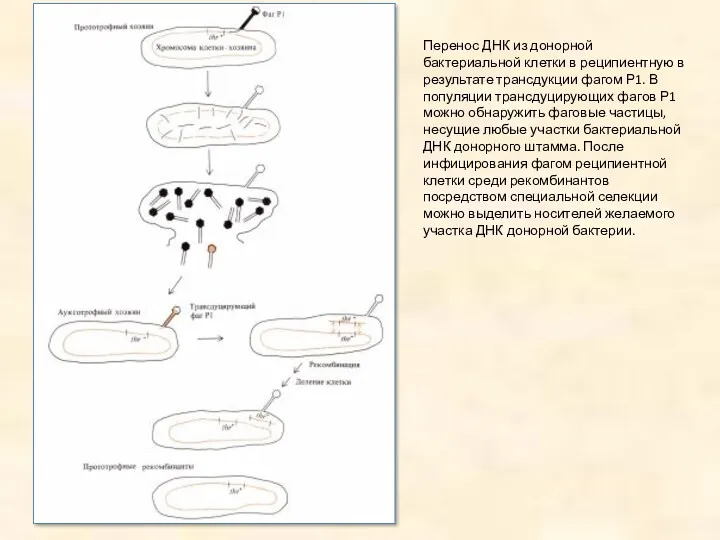
Перенос ДНК из донорной бактериальной клетки в реципиентную в результате трансдукции
Перенос ДНК из донорной бактериальной клетки в реципиентную в результате трансдукции
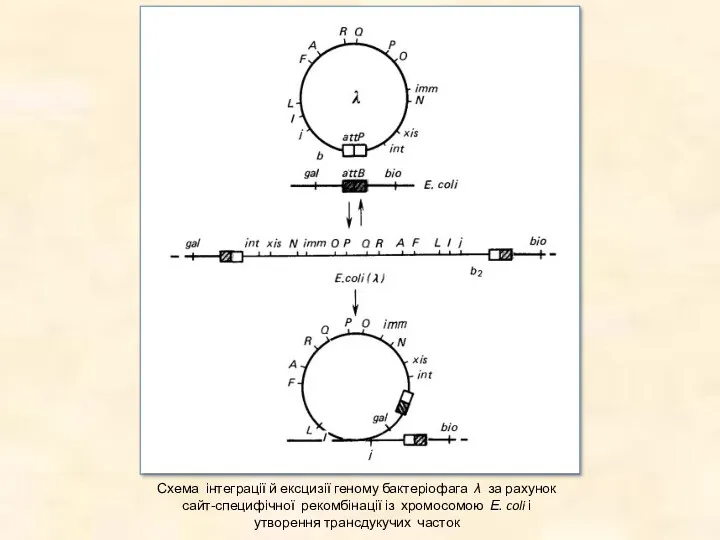
Схема інтеграції й ексцизії геному бактеріофага λ за рахунок сайт-специфічної рекомбінації
Схема інтеграції й ексцизії геному бактеріофага λ за рахунок сайт-специфічної рекомбінації

Генетичний аналіз у бактерій
При кон'югації в клітину-реципієнт переноситься звичайно менше половини
Генетичний аналіз у бактерій
При кон'югації в клітину-реципієнт переноситься звичайно менше половини
 Молекулярно-генетические механизмы развития корня
Молекулярно-генетические механизмы развития корня Интерсекс. Терминология и идентичность. Гендерная идентичность
Интерсекс. Терминология и идентичность. Гендерная идентичность Хромосомная теория наследственности. Сцепленное наследование
Хромосомная теория наследственности. Сцепленное наследование Теория И.П. Павлова о типах высшей нервной деятельности
Теория И.П. Павлова о типах высшей нервной деятельности Строение семян двудольных и однодольных растений
Строение семян двудольных и однодольных растений Тонкий кишечник
Тонкий кишечник The muscular system
The muscular system Мы - друзья пернатых
Мы - друзья пернатых Вода - основной источник жизни и здоровья
Вода - основной источник жизни и здоровья Асқазан. Дамуы
Асқазан. Дамуы Різноманітність ссавців
Різноманітність ссавців Биологические ритмы
Биологические ритмы 7 Б класс. Принял эстафету
7 Б класс. Принял эстафету 1 апреля – Международный день птиц
1 апреля – Международный день птиц Проверка знаний
Проверка знаний Внутрішня будова ссавців
Внутрішня будова ссавців Физиология сенсорных систем
Физиология сенсорных систем Квіткові рослини
Квіткові рослини Урок биологии Многообразие ракообразных с презентацией в 7 классе по учебнику Н.И.Сонина
Урок биологии Многообразие ракообразных с презентацией в 7 классе по учебнику Н.И.Сонина Водоросли. Особенности строения, питания, размножения
Водоросли. Особенности строения, питания, размножения Внеклассное мероприятие по биологии в 7-8 классах на тему: Мы- за здоровье!
Внеклассное мероприятие по биологии в 7-8 классах на тему: Мы- за здоровье! Репликация ДНК. (Лекция 3)
Репликация ДНК. (Лекция 3) Витамины
Витамины Фиалка домашняя
Фиалка домашняя Мегаспорогенез және микроспорогенез
Мегаспорогенез және микроспорогенез Бактериофаги. Основная роль
Бактериофаги. Основная роль Урок-игра по теме Системы органов животных
Урок-игра по теме Системы органов животных Полевые цветы. Фото
Полевые цветы. Фото