- Механизмы газообмена. Лекция 23

Содержание
- 2. Диффузия газов в легких и транспорт газов кровью. Переносчиком кислорода из альвеолярного воздуха к тканям тела
- 3. Приведенные цифры показывают, что венозная кровь, пройдя по капиллярам легкого, обогащается кислородом и теряет углекислый газ.
- 4. Диффузия газов через аэрогематический барьер. В организме газообмен О2 и СО2 через альвеолярно-капиллярную мембрану происходит с
- 5. В легких ΔР является градиентом давлений газа в альвеолах и в крови легочных капилляров. Проницаемость альвеолярно-капиллярной
- 6. Содержание газов в альвеолярном воздухе. Выше было указано парциальное давление газов в альвеолярной газовой смеси, которое
- 7. 23. 2. Транспорт О2 и СО2 кровью. Газообмен между кровью и тканями. Связывание кислорода гемоглобином. Транспорт
- 8. Гемоглобин переносит О2 от легких к тканям. Эта функция зависит от двух свойств гемоглобина: 1) способности
- 9. Графическая зависимость насыщения гемоглобина кислородом от напряжения О2 называется кривой диссоциации оксигемоглобина. Эта кривая имеет S-образную
- 10. Артерио-венозная разница по О2. Так как содержание кислорода в артериальной крови составляет 0,2 л на 1
- 11. Под кислородной емкостью крови понимают количество О2, которое связывается кровью до полного насыщения гемоглобина. При содержании
- 12. Перенос углекислого газа. Поступление СО2 в легких из крови в альвеолы обеспечивается из следующих источников: 1)
- 13. Диффузия СО2 из тканей в кровь. Обмен СО2 между клетками тканей с кровью тканевых капилляров осуществляется
- 14. Ионы НСО3– выходят в плазму, а вместо них в эритроцит поступают анионы хлора – так сохраняется
- 16. Скачать презентацию

Диффузия газов в легких и транспорт газов кровью. Переносчиком кислорода
Диффузия газов в легких и транспорт газов кровью. Переносчиком кислорода

Приведенные цифры показывают, что венозная кровь, пройдя по капиллярам легкого, обогащается
Приведенные цифры показывают, что венозная кровь, пройдя по капиллярам легкого, обогащается

Диффузия газов через аэрогематический барьер.
В организме газообмен О2 и СО2
Диффузия газов через аэрогематический барьер.
В организме газообмен О2 и СО2

В легких ΔР является градиентом давлений газа в альвеолах и в
В легких ΔР является градиентом давлений газа в альвеолах и в

Содержание газов в альвеолярном воздухе.
Выше было указано парциальное давление газов
Содержание газов в альвеолярном воздухе.
Выше было указано парциальное давление газов

23. 2. Транспорт О2 и СО2 кровью. Газообмен между кровью и
23. 2. Транспорт О2 и СО2 кровью. Газообмен между кровью и

Гемоглобин переносит О2 от легких к тканям. Эта функция зависит от
Гемоглобин переносит О2 от легких к тканям. Эта функция зависит от

Графическая зависимость насыщения гемоглобина кислородом от напряжения О2 называется кривой
Графическая зависимость насыщения гемоглобина кислородом от напряжения О2 называется кривой

Артерио-венозная разница по О2.
Так как содержание кислорода в артериальной крови
Артерио-венозная разница по О2.
Так как содержание кислорода в артериальной крови

Под кислородной емкостью крови понимают количество О2, которое связывается кровью до
Под кислородной емкостью крови понимают количество О2, которое связывается кровью до

Перенос углекислого газа.
Поступление СО2 в легких из крови в альвеолы
Перенос углекислого газа.
Поступление СО2 в легких из крови в альвеолы

Диффузия СО2 из тканей в кровь. Обмен СО2 между клетками тканей
Диффузия СО2 из тканей в кровь. Обмен СО2 между клетками тканей

Ионы НСО3– выходят в плазму, а вместо них в эритроцит поступают
Ионы НСО3– выходят в плазму, а вместо них в эритроцит поступают
 Мінеральні речовини в організмі тварин
Мінеральні речовини в організмі тварин Класс Брюхоногие моллюски, или Улитки
Класс Брюхоногие моллюски, или Улитки Динозавр. Гигантские монстры
Динозавр. Гигантские монстры Классификация мутаций
Классификация мутаций Қазіргі заманғы иммунология, оның мақсаттары мен жетістіктері. Иммундық жүйе. Гуморалдық иммунитет
Қазіргі заманғы иммунология, оның мақсаттары мен жетістіктері. Иммундық жүйе. Гуморалдық иммунитет Прицел на старение человека – Можем ли мы продлить жизнь человека?
Прицел на старение человека – Можем ли мы продлить жизнь человека? Виділення. Функції нирок
Виділення. Функції нирок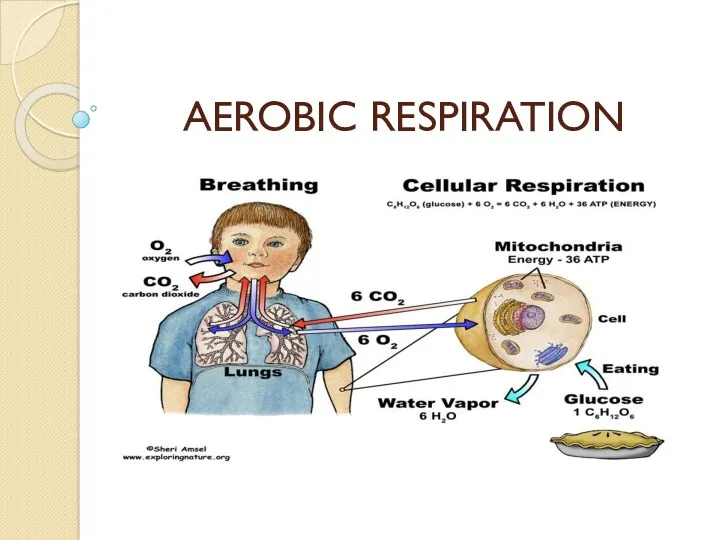 Aerobic respiration
Aerobic respiration Предшественники человека
Предшественники человека Квітка як орган статевого розмноження рослин
Квітка як орган статевого розмноження рослин Возможность зимовки и гнездования лебедей-шипунов вместе с утками на речке Черноголовка в городской зоне
Возможность зимовки и гнездования лебедей-шипунов вместе с утками на речке Черноголовка в городской зоне Млекопитающие отряд приматы
Млекопитающие отряд приматы Молекулярные и клеточные механизмы старения: современные концепции
Молекулярные и клеточные механизмы старения: современные концепции Закон гомологических рядов
Закон гомологических рядов Химическая эволюция и биогенез
Химическая эволюция и биогенез Презентация по биологии Бесполое размножение организмов
Презентация по биологии Бесполое размножение организмов Физиология сенсорных рецепторов. Лекция № 26
Физиология сенсорных рецепторов. Лекция № 26 Голосеменные растения (лат. Gymnospérmae)
Голосеменные растения (лат. Gymnospérmae)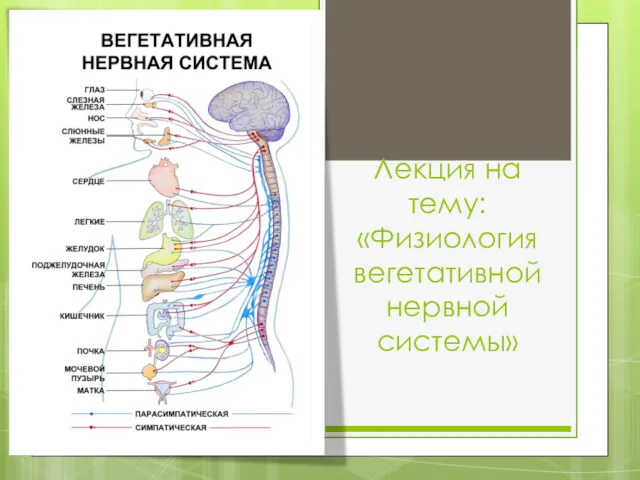 Физиология вегетативной нервной системы. Вегетативная нервная система
Физиология вегетативной нервной системы. Вегетативная нервная система Влияние биоритмов на физическую работоспособность учащихся
Влияние биоритмов на физическую работоспособность учащихся Tardigrades ( also known as water bears)
Tardigrades ( also known as water bears) Познавательная игра для 7-х классов Микромир
Познавательная игра для 7-х классов Микромир Презентация Наследственные и врожденные заболевания.
Презентация Наследственные и врожденные заболевания. Генетична психологія
Генетична психологія Белки, жиры, углеводы
Белки, жиры, углеводы Зоология беспозвоночных
Зоология беспозвоночных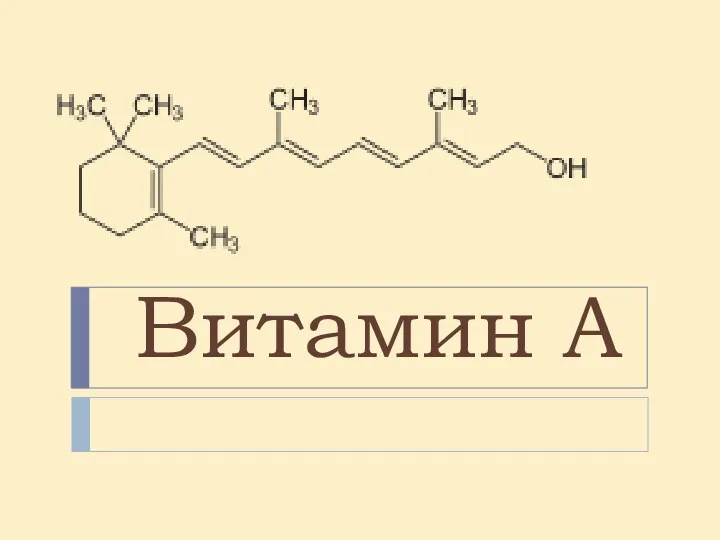 Витамин А
Витамин А Исследование биомассы водорослей озера Байкал
Исследование биомассы водорослей озера Байкал