- Морфология и ультраструктура бактериальных клеток. Простые и сложные методы окраски

Содержание
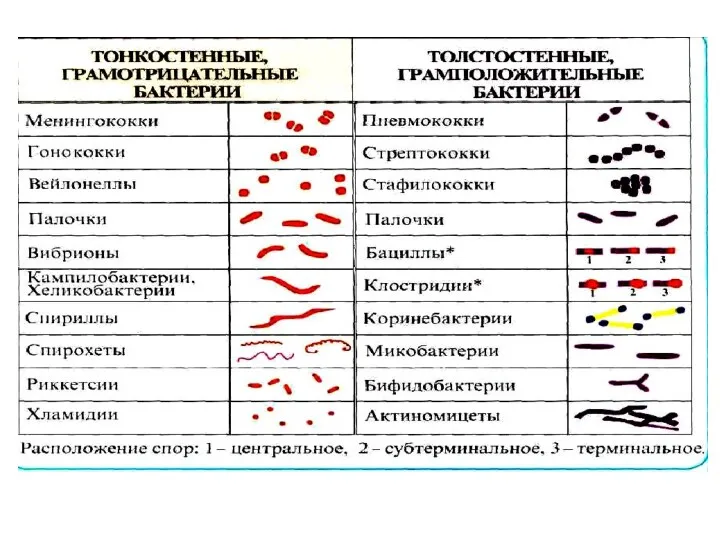
- 2. Основные морфологические группы бактерий Морфология бактерий - размер, форма и взаимное расположение бактериальных клеток. Формы бактерий
- 3. Кокковидные патогенные бактерии обычно имеют форму правильного шара диаметром 1,0— 1,5 мкм; некоторые — бобовидную, ланцетовидную,
- 4. Палочковидные (цилиндрические) формы бактерий. Палочки бывают длинными — более З мкм, короткими — 1,5—3,0 мкм и
- 5. По взаиморасположению бактерий их подразделяют на три группы: 1) монобактерии — палочки располагаются одиночно и беспорядочно,
- 6. Извитые (спиралевидные) бактерии по количеству и характеру завитков, а также по диаметру клеток подразделяют на две
- 7. Нитевидные формы бактерий. Различают два типа нитевидных бактерий: образующие временные нити и постоянные. Временные нити, иногда
- 9. сарцины актиномицеты
- 10. Этапы приготовления мазков-препаратов. 1.Подготовка стекла: исследуемый материал наносят на чистое обезжиренное смесью Никифорова предметное стекло (смесь
- 11. Тинкториальные свойства - свойства бактерий, грибов и простейших, характеризующие их способность вступать в реакцию с красителями
- 12. Структурные компоненты бактериальной клетки: 1. Поверхностные структуры: - капсула - клеточная стенка - ЦПМ - жгутики,
- 13. Постоянные структуры: - клеточная стенка, - ЦПМ, - нуклеоид, рибосомы, мезосомы 2. Непостоянные структуры: - ворсинки,
- 14. Функции облигатных и факультативных компонентов бактериальной клетки Клеточная стенка — структурный компонент, присущий только бактериям (кроме
- 15. Строение клеточной стенки В составе клеточной стенки имеется два слоя: наружный — пластичный и внутренний —
- 16. Пептидогликан обладает следующими важнейшими иммунобиологическими свойствами: 1. В его составе обнаружены родоспецифические антигенные детерминанты. 2. Пептидогликан
- 17. Особенности клеточной стенки грамположительных бактерий Основную массу стенки составляет пептидогликан. Он представлен не 1-2 слоями, как
- 18. Особенности клеточной стенки грамотрицательных бактерий Основная особенность клеточной стенки грамотрицательных бактерий: ригидный слой тонкий, представлен одним
- 19. Принцип метода окраски по Граму: Генцианвиолет связывается с пептидогликаном клеточной стенки. Толстый слой пептидогликана грамположительных бактерий
- 21. Смесь Грам+ и Грам- бактерий
- 22. Цитоплазматическая мембрана: строение (кле́точная мембра́на, или цитолемма, или плазмолемма) ЦПМ содержит 25—40% фосфолипидов, образующих два слоя,
- 23. Функции цитоплазматической мембраны барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой.
- 24. Нуклеоид — эквивалент ядра. Здесь находится генетический материал (хранение наследственной информации) Расположен в центральной зоне бактерий
- 25. Цитоплазма занимает основной объем бактериальной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных
- 26. МЕЗОСОМЫ Внутрицитоплазматические мембранные структуры бактерий везикулярной и трубчатой формы, образующиеся путём впячивания ЦПМ внутрь цитоплазмы. Предполагается,
- 27. Ворсинки, или пили (фимбрии) — нитевидные образования, более тонкие и короткие, чем жгутики. Пили отходят от
- 28. Пили, E.coli (эл.микроскопия)
- 29. Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны,
- 30. Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и
- 31. Капсула, микрокапсула, слизь. Капсула — слизистая структура толщиной более 0,2 мкм, прочно связанная с клеточной стенкой
- 32. Споры — своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных
- 33. Форма спор может быть овальной, шаровидной; расположение в клетке — терминальное, т. е. на конце палочки
- 34. В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной кислоты и полифосфатов (волютин). Они
- 35. Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по
- 36. СЛОЖНЫЕ МЕТОДЫ ОКРАСКИ ВЫЯВЛЕНИЕ КАПСУЛ ПО БУРРИ И БУРРИ-ГИНСУ. На середину предметного стекла наносят каплю черной
- 37. Мазок из чистой культуры Klebsiella pneumoniae, окраска по Бурри-Гинсу. Видны капсулы — светлые ореолы вокруг палочковидных
- 38. Механизм и этапы окраски по Цилю-Нельсону (выявление свойства кислотоустойчивости) 1. На фиксированный мазок наносят карболовый р-р
- 39. Mycobacterium tuberculosis, окраска по Цилю — Нельсену
- 40. ОКРАСКА СПОР ПО ОЖЕШКО нанести несколько капель 0,5% соляной кислоты на нефиксированный мазок и нагреть до
- 41. Окрашенный препарат Bacillus subtilis. Вегетативные клетки выделены красным, споры — зелёным.
- 42. ОКРАСКА ВОЛЮТИНА ПО НЕЙССЕРУ Окрасить мазок уксуснокислой синькой Нейссера (2-3 минуты); Промыть; Окрасить раствором Люголя (30
- 43. Рисунок мазка из чистой культуры С diphtheriae. Окраска по Нейссеру
- 45. Скачать презентацию

Основные морфологические группы бактерий
Морфология бактерий - размер, форма и взаимное расположение
Основные морфологические группы бактерий
Морфология бактерий - размер, форма и взаимное расположение

Кокковидные патогенные бактерии обычно имеют форму правильного шара диаметром 1,0— 1,5
Кокковидные патогенные бактерии обычно имеют форму правильного шара диаметром 1,0— 1,5

Палочковидные (цилиндрические) формы бактерий.
Палочки бывают длинными — более З мкм, короткими
Палочковидные (цилиндрические) формы бактерий.
Палочки бывают длинными — более З мкм, короткими

По взаиморасположению бактерий их подразделяют на три группы:
1) монобактерии —
По взаиморасположению бактерий их подразделяют на три группы:
1) монобактерии —

Извитые (спиралевидные) бактерии по количеству и характеру завитков, а также по
Извитые (спиралевидные) бактерии по количеству и характеру завитков, а также по

Нитевидные формы бактерий. Различают два типа нитевидных бактерий: образующие временные нити
Нитевидные формы бактерий. Различают два типа нитевидных бактерий: образующие временные нити

сарцины актиномицеты
сарцины актиномицеты

Этапы приготовления мазков-препаратов.
1.Подготовка стекла: исследуемый материал наносят на чистое обезжиренное смесью
Этапы приготовления мазков-препаратов.
1.Подготовка стекла: исследуемый материал наносят на чистое обезжиренное смесью

Тинкториальные свойства - свойства бактерий, грибов и простейших, характеризующие их способность
Тинкториальные свойства - свойства бактерий, грибов и простейших, характеризующие их способность

Структурные компоненты бактериальной клетки:
1. Поверхностные структуры:
- капсула
- клеточная стенка
-
Структурные компоненты бактериальной клетки:
1. Поверхностные структуры:
- капсула
- клеточная стенка
-

Постоянные структуры:
- клеточная стенка,
- ЦПМ,
- нуклеоид, рибосомы, мезосомы
2. Непостоянные
Постоянные структуры:
- клеточная стенка,
- ЦПМ,
- нуклеоид, рибосомы, мезосомы
2. Непостоянные

Функции облигатных и факультативных компонентов бактериальной клетки
Клеточная стенка — структурный компонент,
Функции облигатных и факультативных компонентов бактериальной клетки
Клеточная стенка — структурный компонент,

Строение клеточной стенки
В составе клеточной стенки имеется два слоя: наружный —
Строение клеточной стенки
В составе клеточной стенки имеется два слоя: наружный —

Пептидогликан обладает следующими важнейшими иммунобиологическими свойствами:
1. В его составе обнаружены родоспецифические
Пептидогликан обладает следующими важнейшими иммунобиологическими свойствами:
1. В его составе обнаружены родоспецифические

Особенности клеточной стенки грамположительных бактерий
Основную массу стенки составляет пептидогликан. Он
Особенности клеточной стенки грамположительных бактерий
Основную массу стенки составляет пептидогликан. Он

Особенности клеточной стенки грамотрицательных бактерий
Основная особенность клеточной стенки грамотрицательных бактерий:
Особенности клеточной стенки грамотрицательных бактерий
Основная особенность клеточной стенки грамотрицательных бактерий:

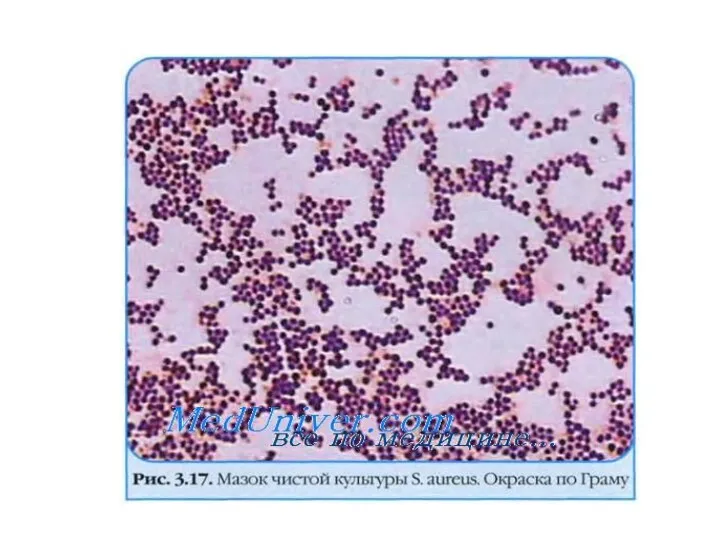
Принцип метода окраски по Граму:
Генцианвиолет связывается с пептидогликаном клеточной стенки. Толстый
Принцип метода окраски по Граму:
Генцианвиолет связывается с пептидогликаном клеточной стенки. Толстый
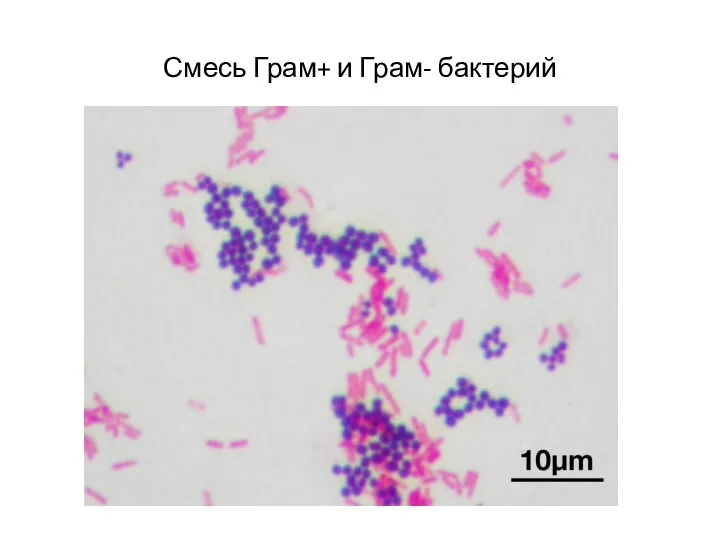
Смесь Грам+ и Грам- бактерий
Смесь Грам+ и Грам- бактерий

Цитоплазматическая мембрана: строение
(кле́точная мембра́на, или цитолемма, или плазмолемма)
ЦПМ содержит 25—40%
Цитоплазматическая мембрана: строение
(кле́точная мембра́на, или цитолемма, или плазмолемма)
ЦПМ содержит 25—40%

Функции цитоплазматической мембраны
барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен
Функции цитоплазматической мембраны
барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен

Нуклеоид — эквивалент ядра. Здесь находится генетический материал (хранение наследственной информации)
Нуклеоид — эквивалент ядра. Здесь находится генетический материал (хранение наследственной информации)

Цитоплазма занимает основной объем бактериальной клетки и состоит из растворимых белков,
Цитоплазма занимает основной объем бактериальной клетки и состоит из растворимых белков,

МЕЗОСОМЫ
Внутрицитоплазматические мембранные структуры бактерий везикулярной и трубчатой формы, образующиеся путём впячивания
МЕЗОСОМЫ
Внутрицитоплазматические мембранные структуры бактерий везикулярной и трубчатой формы, образующиеся путём впячивания

Ворсинки, или пили (фимбрии) — нитевидные образования, более тонкие и короткие,
Ворсинки, или пили (фимбрии) — нитевидные образования, более тонкие и короткие,
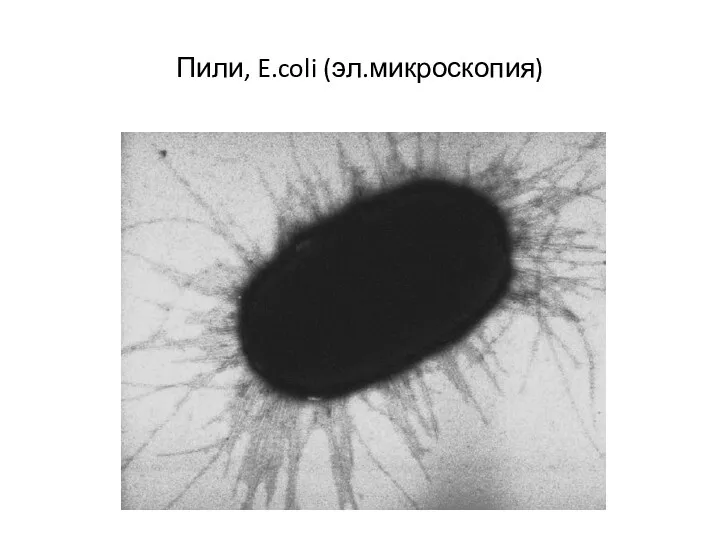
Пили, E.coli (эл.микроскопия)
Пили, E.coli (эл.микроскопия)

Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити,
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити,

Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у
Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у

Капсула, микрокапсула, слизь.
Капсула — слизистая структура толщиной более 0,2 мкм,
Капсула, микрокапсула, слизь.
Капсула — слизистая структура толщиной более 0,2 мкм,

Споры — своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной
Споры — своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной

Форма спор может быть овальной, шаровидной;
расположение в клетке — терминальное,
Форма спор может быть овальной, шаровидной;
расположение в клетке — терминальное,

В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, бета-оксимасляной

Волютин обладает сродством к основным красителям и легко выявляется с помощью
Волютин обладает сродством к основным красителям и легко выявляется с помощью

СЛОЖНЫЕ МЕТОДЫ ОКРАСКИ
ВЫЯВЛЕНИЕ КАПСУЛ ПО БУРРИ И БУРРИ-ГИНСУ.
На середину предметного стекла
СЛОЖНЫЕ МЕТОДЫ ОКРАСКИ
ВЫЯВЛЕНИЕ КАПСУЛ ПО БУРРИ И БУРРИ-ГИНСУ.
На середину предметного стекла
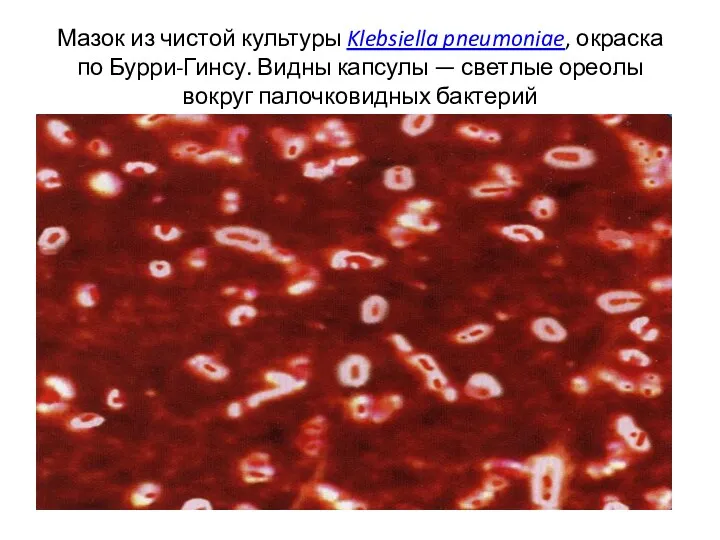
Мазок из чистой культуры Klebsiella pneumoniae, окраска по Бурри-Гинсу. Видны капсулы — светлые
Мазок из чистой культуры Klebsiella pneumoniae, окраска по Бурри-Гинсу. Видны капсулы — светлые

Механизм и этапы окраски по Цилю-Нельсону (выявление свойства кислотоустойчивости)
1. На фиксированный
Механизм и этапы окраски по Цилю-Нельсону (выявление свойства кислотоустойчивости)
1. На фиксированный
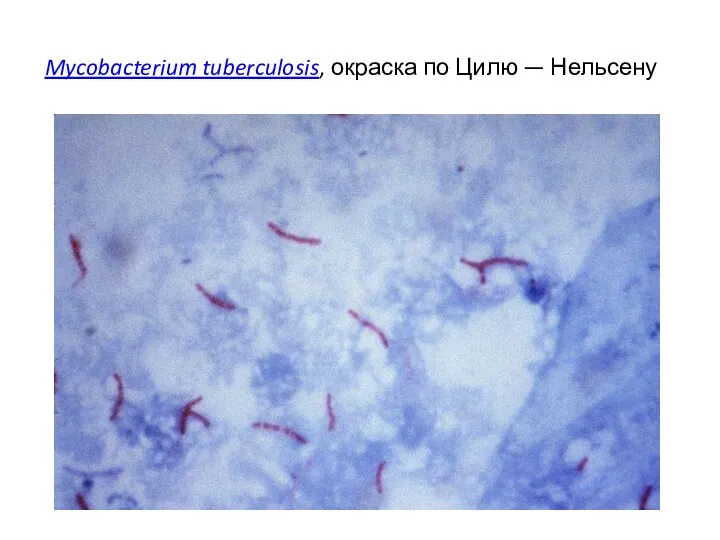
Mycobacterium tuberculosis, окраска по Цилю — Нельсену
Mycobacterium tuberculosis, окраска по Цилю — Нельсену

ОКРАСКА СПОР ПО ОЖЕШКО
нанести несколько капель 0,5% соляной кислоты на нефиксированный
ОКРАСКА СПОР ПО ОЖЕШКО
нанести несколько капель 0,5% соляной кислоты на нефиксированный
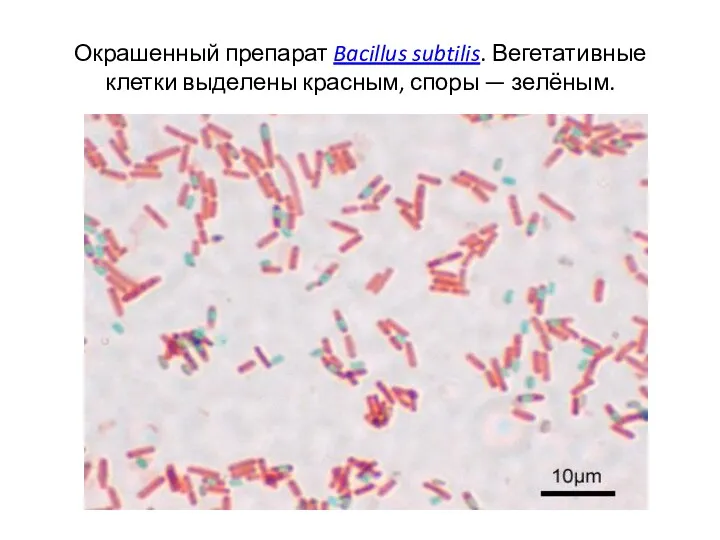
Окрашенный препарат Bacillus subtilis. Вегетативные клетки выделены красным, споры — зелёным.
Окрашенный препарат Bacillus subtilis. Вегетативные клетки выделены красным, споры — зелёным.

ОКРАСКА ВОЛЮТИНА ПО НЕЙССЕРУ
Окрасить мазок уксуснокислой синькой Нейссера (2-3 минуты);
Промыть;
Окрасить
ОКРАСКА ВОЛЮТИНА ПО НЕЙССЕРУ
Окрасить мазок уксуснокислой синькой Нейссера (2-3 минуты);
Промыть;
Окрасить

Рисунок мазка из чистой культуры С diphtheriae. Окраска по Нейссеру
Рисунок мазка из чистой культуры С diphtheriae. Окраска по Нейссеру
 Генетические аспекты спортивного отбора
Генетические аспекты спортивного отбора АТФ и другие органические соединения клетки
АТФ и другие органические соединения клетки Вторичный метаболизм высших растений. Алкалоиды. Часть 1
Вторичный метаболизм высших растений. Алкалоиды. Часть 1 Класс Млекопитающие или Звери. Отряды: Однопроходные, Сумчатые, Насекомоядные, Рукокрылые
Класс Млекопитающие или Звери. Отряды: Однопроходные, Сумчатые, Насекомоядные, Рукокрылые Происхождение современного человека. (Тема 3)
Происхождение современного человека. (Тема 3) Механизмы старения и его профилактика
Механизмы старения и его профилактика Поток энергии и цепи питания
Поток энергии и цепи питания Бактериология. Классификация бактерий. Морфология бактерий и методы её изучения. (Лекция 9)
Бактериология. Классификация бактерий. Морфология бактерий и методы её изучения. (Лекция 9) Материальные основы наследственности
Материальные основы наследственности Свойства ферментов. Механизм действия. Лекция 2
Свойства ферментов. Механизм действия. Лекция 2 Растения Мотовилихинского пруда
Растения Мотовилихинского пруда Структурная организация белка
Структурная организация белка Сообщество, экосистема, биогеоценоз
Сообщество, экосистема, биогеоценоз Биофизика фоторецепции
Биофизика фоторецепции Грибы, одно из царств живых организмов (ранее относились к низшим растениям)
Грибы, одно из царств живых организмов (ранее относились к низшим растениям) 373acb0e4cd14e079a5c6c48d3803fbf
373acb0e4cd14e079a5c6c48d3803fbf Строение и физиология слуховой системы человека
Строение и физиология слуховой системы человека песчаная эфа
песчаная эфа Биология клетки
Биология клетки Ферменты в генетической инженерии. Тема 2
Ферменты в генетической инженерии. Тема 2 Нервная система
Нервная система Презентация к уроку биологии с использованием модульных технологий по теме Органические вещества клетки. нуклеиновые кислоты.
Презентация к уроку биологии с использованием модульных технологий по теме Органические вещества клетки. нуклеиновые кислоты. Основные взаимосвязи в организме, образующиеся при мышечной деятельности. Тема 1
Основные взаимосвязи в организме, образующиеся при мышечной деятельности. Тема 1 Выращивание кактуса в домашних условиях
Выращивание кактуса в домашних условиях Царство животные
Царство животные Чудо-доктора. Лекарственные растения Лутугинского района
Чудо-доктора. Лекарственные растения Лутугинского района Органогенез. Постэмбриональное развитие.
Органогенез. Постэмбриональное развитие. Биохимия нервной ткани. Биологические мембраны
Биохимия нервной ткани. Биологические мембраны