- Моторика (регуляція рухів)

Содержание
- 2. МОТОРИКА - 1 (регуляція рухів) Пропріорецептори. Моторні рефлекси спинного мозку. Вестибулярний аналізатор. Моторні рефлекси ствола мозку.
- 3. Це найбільший орган - 30-40% маси тіла. Завдяки появі скелетних м'язів тварини стали відрізнятися від рослин.
- 4. Моторні центры мозку У міру формування мозку в кожному з його відділів з'являлися моторні центри
- 5. Скорочення скелетних м'язів спрямоване не тільки на виконання справжніх цілеспрямованих локомоций, а й на протидію гравітаційним
- 6. пропріорецептори, вестибулярний, рецептори шкіри, зоровий, слуховий та інші. Рецептори, які приймають участьв регуляції моторики
- 7. Нервні центри Багаторівневий (розташований в різних структурах ЦНС) нервовий центр забезпечує людині можливість виконання всієї різноманітної
- 8. Спинний мозок У сегментах спинного мозку пул мотонейронів кожного м'яза знаходиться в певному місці, причому: згиначі
- 9. Сухожильний рефлекс При ударі молоточком по сухожиль (будь-якого м'яза) м'яз скорочується. Цей ефект обумовлений рефлексом, замикається
- 10. Пропріорецептори Схема сухожильного органу (а), м`язового веретена (б). 1, 12- афферентні волокна, 2 – інтрафузальне м`язове
- 11. Кількість м'язових веретен і сухожильних органів в різних м'язах неоднаково. Чим більше складну, більш значущу для
- 12. Взаємодія нейронів в спинном мозку Рецептори збуджуються при їх розтягуванні. Імпульси від веретен в спинному мозку
- 13. Сухожильний орган Cухожильні органи - контролюють напругу м'язів (силу скорочення). А якщо скорочення занадто сильне (можна
- 14. М'язові веретена - контролюють довжину м'яза. Ця інформація надходить у головний мозок (ми завжди знаємо в
- 15. У природних умовах збудження α- і γ -мотонейронів однієї м'язи виникає найчастіше одночасно. Але внаслідок різної
- 16. Гамма-регуляція м’язового скорочення (γ - петля): механізм усилення скорочення м’язів Вплив ретикулярної формації стовбура мозку вказано
- 17. Рефлекси спинного мозку (Починаються з різних рецепторів або автономних програм) Сегментарні: моносинаптичні полісинаптичні. Межсегментарних: згинальний, перехресний
- 18. Центри верхніх відділів мозку координують рефлекси спинного мозку. А ретикулярна формація стовбура ще і тонізує центри
- 19. Центри ствола мозку
- 20. Моторні центри стволу головного мозку. 1 - червоне ядро, 2 - ядро ретикулярної формації моста, 3
- 21. В орієнтації людини в просторі крім пропріорецепторов велику роль відіграє вестибулярна сенсорна система. Вона разом з
- 22. Лабіринт 1 - Ендолімфатичний мішечок, 2, 3, 4 - півкруглі канали, 5 - равлик, 6 -
- 23. Рецепторна клітина вестибулярного аппарату Вії двох типів: Одна довга - кіноцілій і багато коротких - стероцілій.
- 24. Схема будови, механізм збудження і гальмування рецепторних клітин Клітка має пейсмекерного властивостями (спонтанне збудження близько 200
- 25. Сприйняття зміни руху голови рецепторною кліткою вестибулярного аналізатора і поява електричних відповідей в закінченні сенсорного нейрона
- 26. Схема будови купули і отолитового апарату
- 27. Стимулом для рецепторів півколових каналів є кутові прискорення. Тут рецепторні клітини в кожному каналі згруповані в
- 28. Дратуються вони при зміні швидкості руху (початок і кінець рухів), при так званих рівнях лінійного прискорення.
- 29. Рецептори маточки реагують на зміну швидкості горизонтального руху, а мішечка - вертикального (ліфтні рефлекси) Механізм виникнення
- 30. Перш за все: всі центри стовбура тісно пов'язані між собою. Рефлекси стовбурових моторних ядер реалізуються через
- 31. Стовбурові центри забезпечують збереження рівноваги і нормальне вертикальне положення тіла в умовах дії гравітаційного поля Землі
- 32. Зміна тонусу м'язів при порушенні взаємодії центрів стовбура
- 33. Рефлекси стовбура мозку Позні рефлекси: шийні, вестибулярні, очі-рухові. Статокинетічеськие рефлекси: статичні, тонічні, Орієнтовні. Починаються вони з:
- 34. При «неправильному» положенні голови з пропріорецепторов м'язів шиї запускаються руху м'язів тулуба спрямовані на збереження більш
- 35. Прояв позних рефлексів у кішки при падінні Починаються руху з повороту голови, тобто спочатку включаються шийні
- 36. Перерозподіл тонусу м'язів кішки при нахилах голови У тварин при нахилі голови вниз (рис. Б) підвищується
- 37. При нахилі голови вперед підвищується тонус м'язів-згиначів і в результаті покращується можливість угруповання. При відкиданні голови
- 38. При повороті голови підвищується тонус м'язів згиначів на стороні повороту, а розгиначів на протилежному боці. В
- 39. Участь лабіринту в збереженні горизонтального положення очей при нахилах голови Крім того, ці ж структури беруть
- 41. Скачать презентацию

МОТОРИКА - 1
(регуляція рухів)
Пропріорецептори.
Моторні рефлекси спинного мозку.
Вестибулярний аналізатор.
Моторні рефлекси ствола
МОТОРИКА - 1
(регуляція рухів)
Пропріорецептори.
Моторні рефлекси спинного мозку.
Вестибулярний аналізатор.
Моторні рефлекси ствола

Це найбільший орган - 30-40% маси тіла.
Завдяки появі скелетних м'язів тварини
Це найбільший орган - 30-40% маси тіла.
Завдяки появі скелетних м'язів тварини
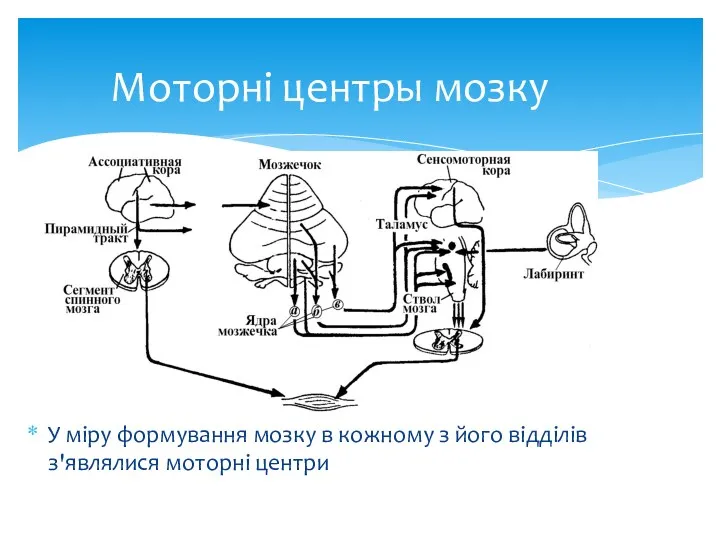
Моторні центры мозку
У міру формування мозку в кожному з його відділів
Моторні центры мозку
У міру формування мозку в кожному з його відділів

Скорочення скелетних м'язів спрямоване не тільки на виконання справжніх цілеспрямованих локомоций,
Скорочення скелетних м'язів спрямоване не тільки на виконання справжніх цілеспрямованих локомоций,

пропріорецептори,
вестибулярний,
рецептори шкіри,
зоровий,
слуховий
та інші.
Рецептори, які приймають участьв регуляції моторики
пропріорецептори,
вестибулярний,
рецептори шкіри,
зоровий,
слуховий
та інші.
Рецептори, які приймають участьв регуляції моторики
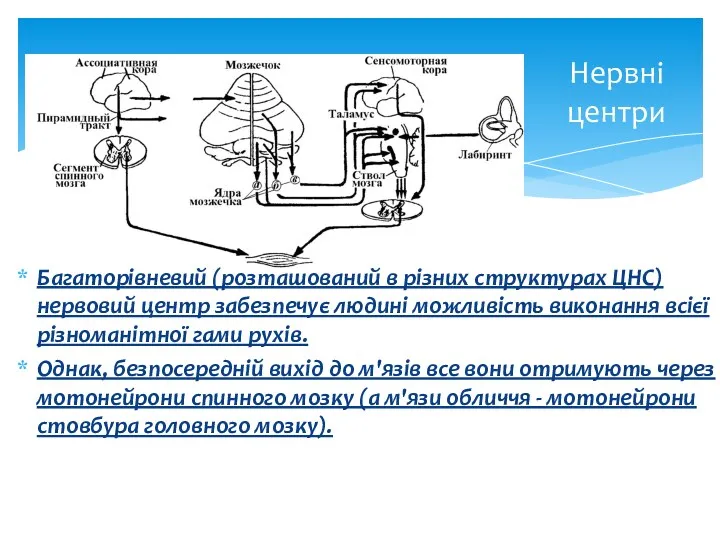
Нервні центри
Багаторівневий (розташований в різних структурах ЦНС) нервовий центр забезпечує людині
Нервні центри
Багаторівневий (розташований в різних структурах ЦНС) нервовий центр забезпечує людині
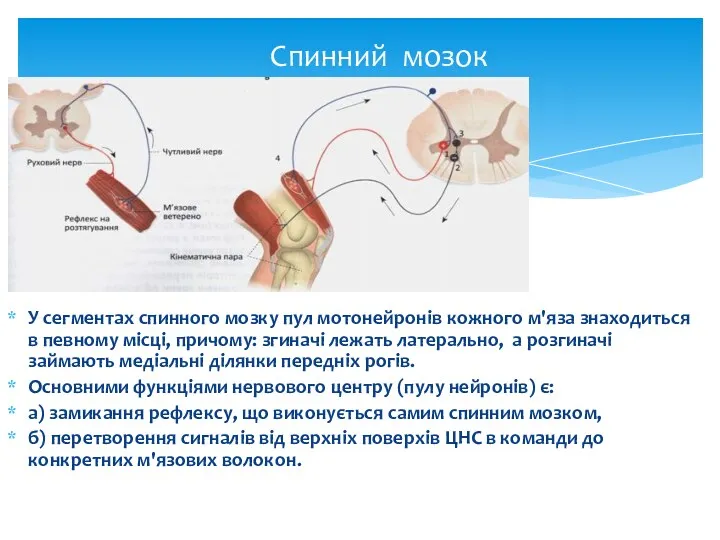
Спинний мозок
У сегментах спинного мозку пул мотонейронів кожного м'яза знаходиться в
Спинний мозок
У сегментах спинного мозку пул мотонейронів кожного м'яза знаходиться в
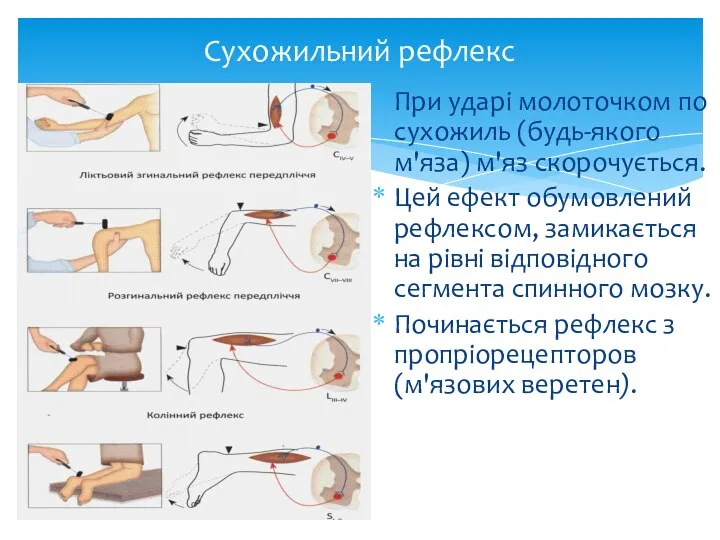
Сухожильний рефлекс
При ударі молоточком по сухожиль (будь-якого м'яза) м'яз скорочується.
Цей ефект
Сухожильний рефлекс
При ударі молоточком по сухожиль (будь-якого м'яза) м'яз скорочується.
Цей ефект
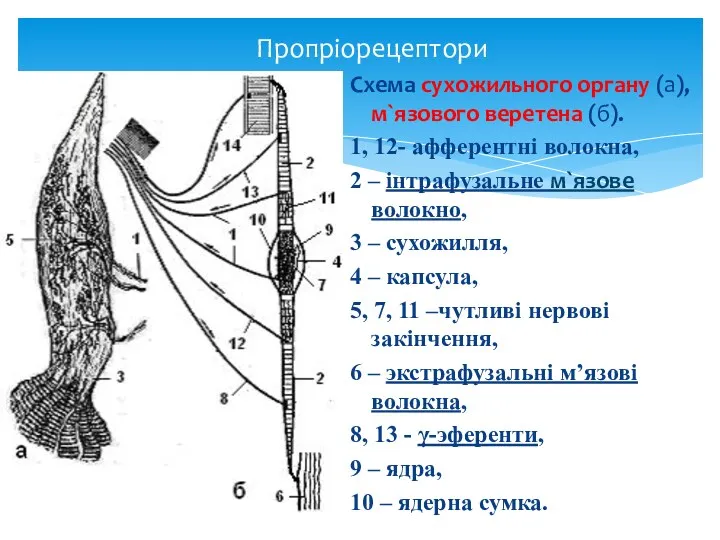
Пропріорецептори
Схема сухожильного органу (а),
м`язового веретена (б).
1, 12- афферентні
Пропріорецептори
Схема сухожильного органу (а),
м`язового веретена (б).
1, 12- афферентні

Кількість м'язових веретен і сухожильних органів в різних м'язах неоднаково. Чим
Кількість м'язових веретен і сухожильних органів в різних м'язах неоднаково. Чим

Взаємодія нейронів в спинном мозку
Рецептори збуджуються при їх розтягуванні.
Імпульси від веретен
Взаємодія нейронів в спинном мозку
Рецептори збуджуються при їх розтягуванні.
Імпульси від веретен
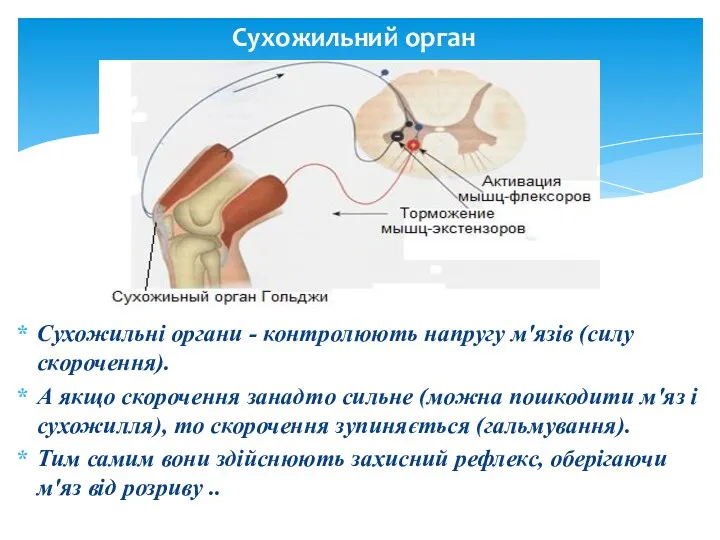
Сухожильний орган
Cухожильні органи - контролюють напругу м'язів (силу скорочення).
А якщо скорочення
Сухожильний орган
Cухожильні органи - контролюють напругу м'язів (силу скорочення).
А якщо скорочення
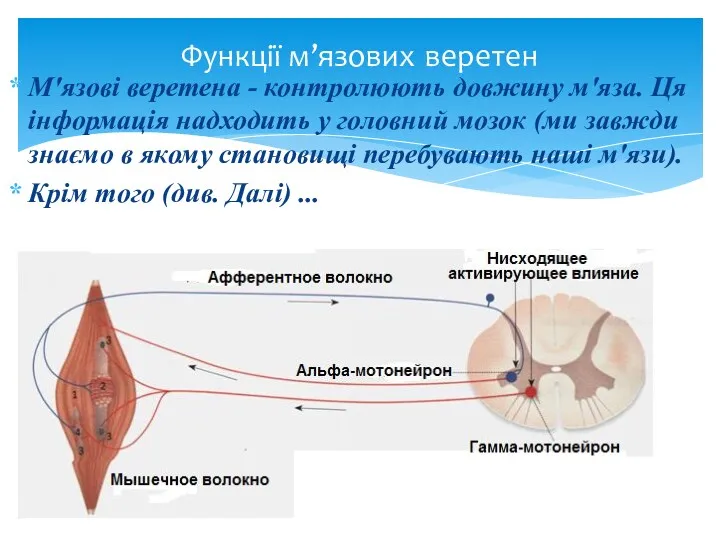
М'язові веретена - контролюють довжину м'яза. Ця інформація надходить у головний
М'язові веретена - контролюють довжину м'яза. Ця інформація надходить у головний

У природних умовах збудження α- і γ -мотонейронів однієї м'язи виникає
У природних умовах збудження α- і γ -мотонейронів однієї м'язи виникає
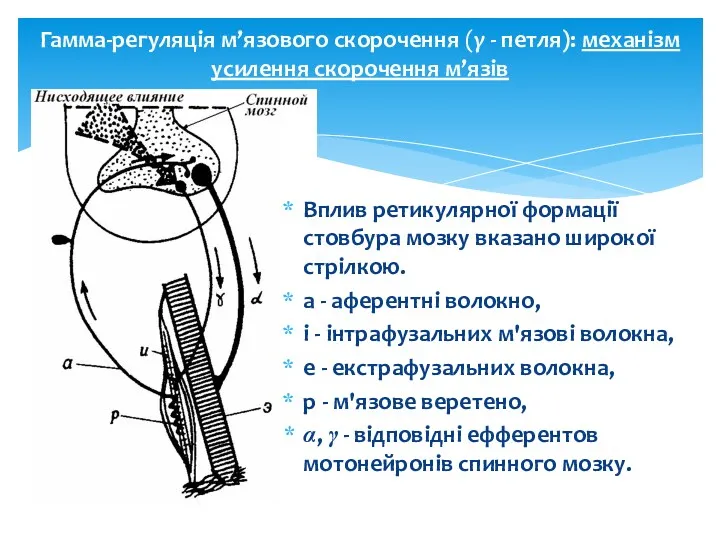
Гамма-регуляція м’язового скорочення (γ - петля): механізм усилення скорочення м’язів
Вплив ретикулярної
Гамма-регуляція м’язового скорочення (γ - петля): механізм усилення скорочення м’язів
Вплив ретикулярної

Рефлекси спинного мозку (Починаються з різних рецепторів або автономних програм)
Сегментарні:
моносинаптичні
полісинаптичні.
Межсегментарних:
згинальний,
перехресний
розгинальний,
чесательний,
Шагательном.
Крім того,
Рефлекси спинного мозку (Починаються з різних рецепторів або автономних програм)
Сегментарні:
моносинаптичні
полісинаптичні.
Межсегментарних:
згинальний,
перехресний
розгинальний,
чесательний,
Шагательном.
Крім того,

Центри верхніх відділів мозку координують рефлекси спинного мозку.
А ретикулярна формація стовбура
Центри верхніх відділів мозку координують рефлекси спинного мозку.
А ретикулярна формація стовбура
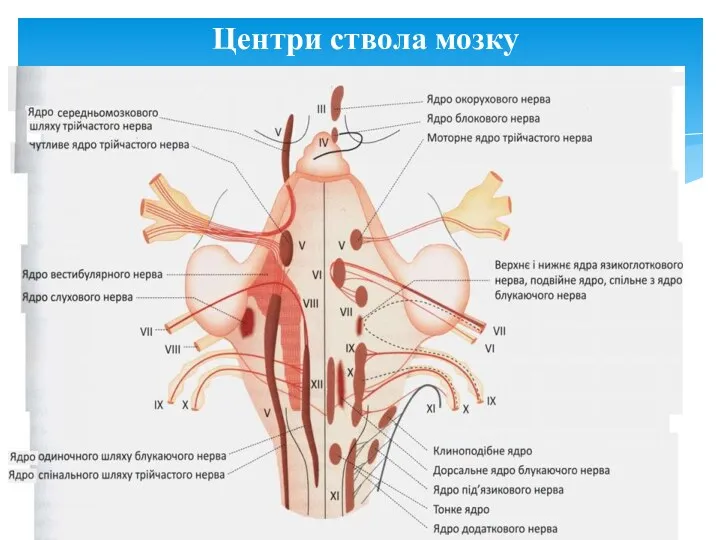
Центри ствола мозку
Центри ствола мозку
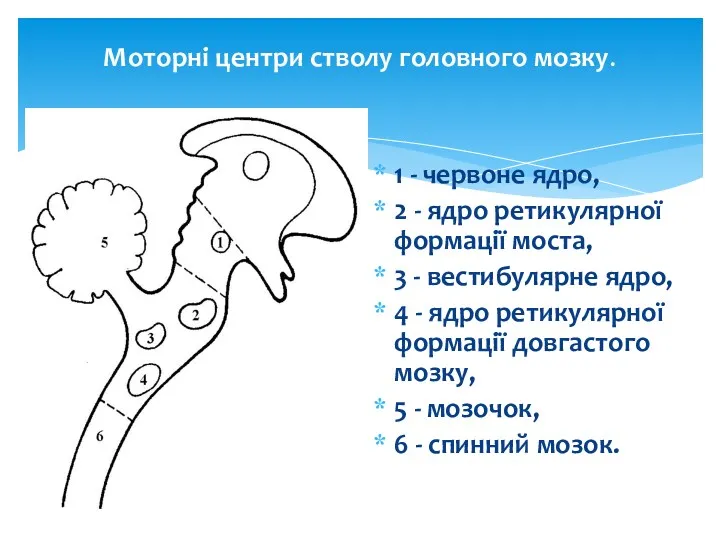
Моторні центри стволу головного мозку.
1 - червоне ядро,
2 - ядро
Моторні центри стволу головного мозку.
1 - червоне ядро,
2 - ядро

В орієнтації людини в просторі крім пропріорецепторов велику роль відіграє вестибулярна
В орієнтації людини в просторі крім пропріорецепторов велику роль відіграє вестибулярна

Лабіринт
1 - Ендолімфатичний мішечок,
2, 3, 4 - півкруглі канали,
5 - равлик,
Лабіринт
1 - Ендолімфатичний мішечок,
2, 3, 4 - півкруглі канали,
5 - равлик,
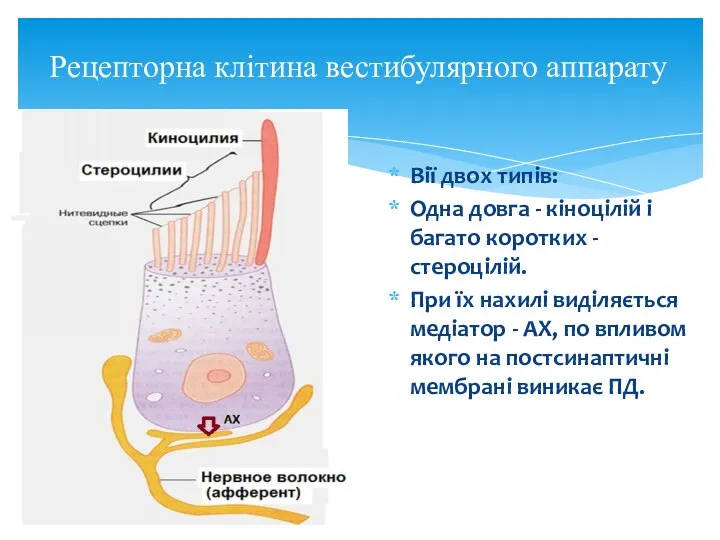
Рецепторна клітина вестибулярного аппарату
Вії двох типів:
Одна довга - кіноцілій і
Рецепторна клітина вестибулярного аппарату
Вії двох типів:
Одна довга - кіноцілій і
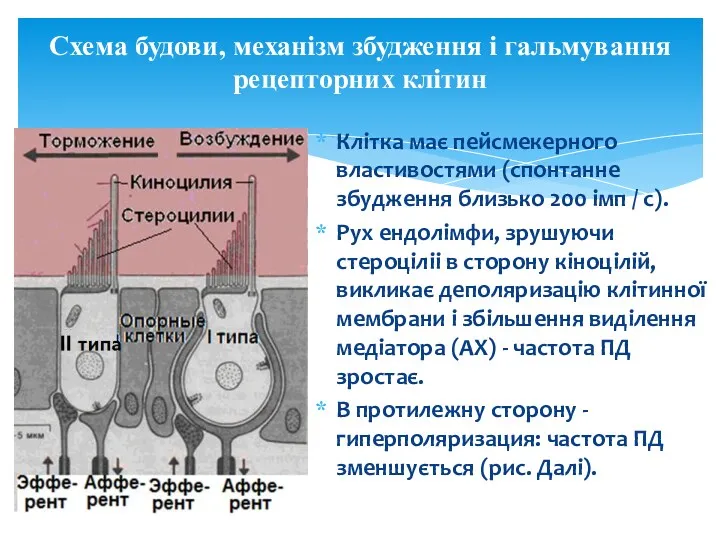
Схема будови, механізм збудження і гальмування рецепторних клітин
Клітка має пейсмекерного властивостями
Схема будови, механізм збудження і гальмування рецепторних клітин
Клітка має пейсмекерного властивостями
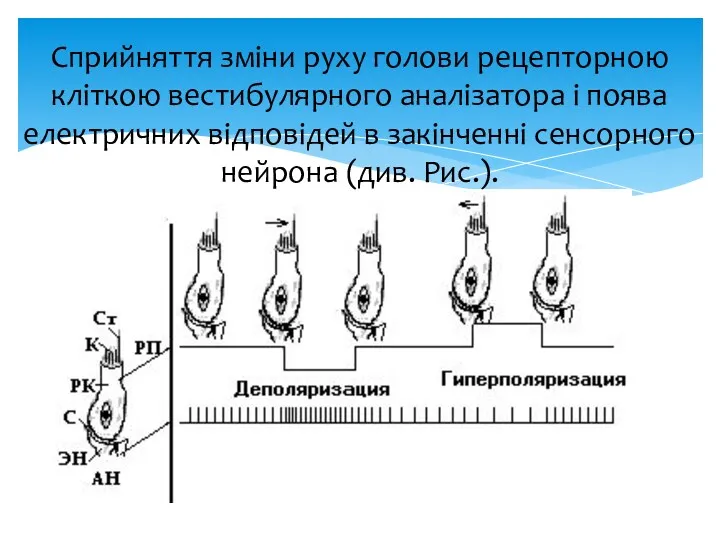
Сприйняття зміни руху голови рецепторною кліткою вестибулярного аналізатора і поява електричних
Сприйняття зміни руху голови рецепторною кліткою вестибулярного аналізатора і поява електричних
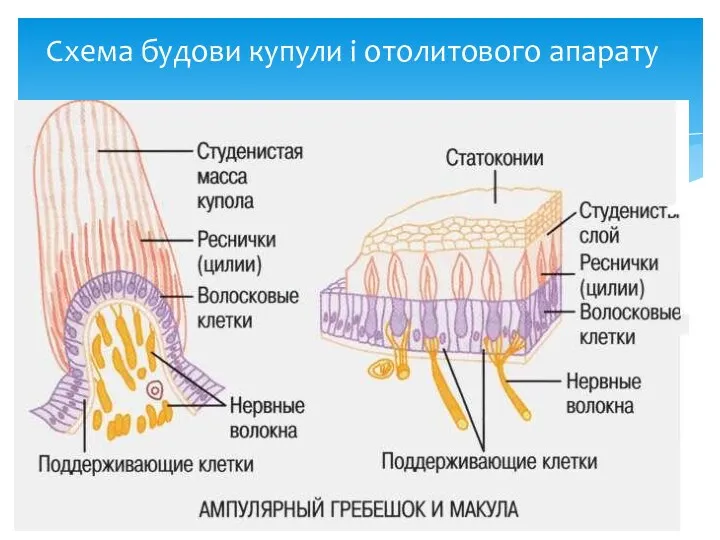
Схема будови купули і отолитового апарату
Схема будови купули і отолитового апарату
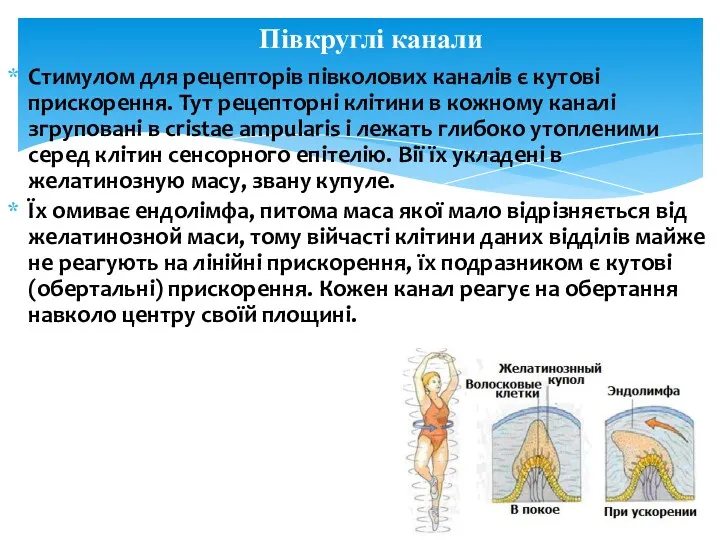
Стимулом для рецепторів півколових каналів є кутові прискорення. Тут рецепторні клітини
Стимулом для рецепторів півколових каналів є кутові прискорення. Тут рецепторні клітини

Дратуються вони при зміні швидкості руху (початок і кінець рухів), при
Дратуються вони при зміні швидкості руху (початок і кінець рухів), при
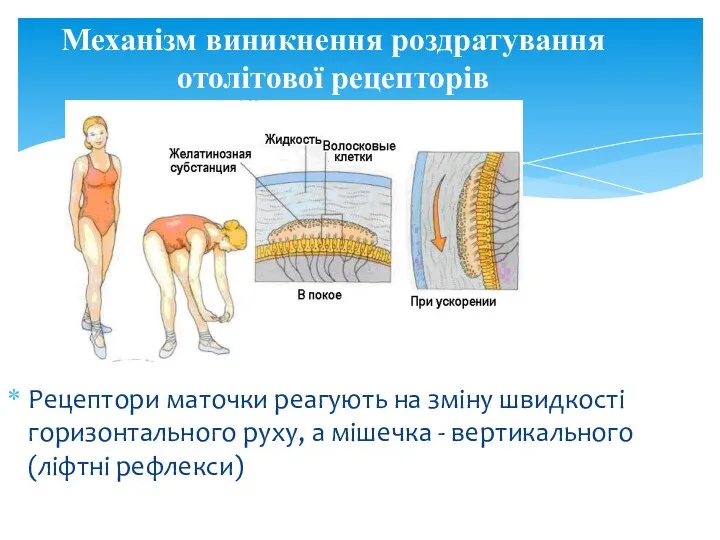
Рецептори маточки реагують на зміну швидкості горизонтального руху, а мішечка -
Рецептори маточки реагують на зміну швидкості горизонтального руху, а мішечка -

Перш за все: всі центри стовбура тісно пов'язані між собою.
Рефлекси стовбурових
Перш за все: всі центри стовбура тісно пов'язані між собою.
Рефлекси стовбурових

Стовбурові центри забезпечують збереження рівноваги і нормальне вертикальне положення тіла в
Стовбурові центри забезпечують збереження рівноваги і нормальне вертикальне положення тіла в
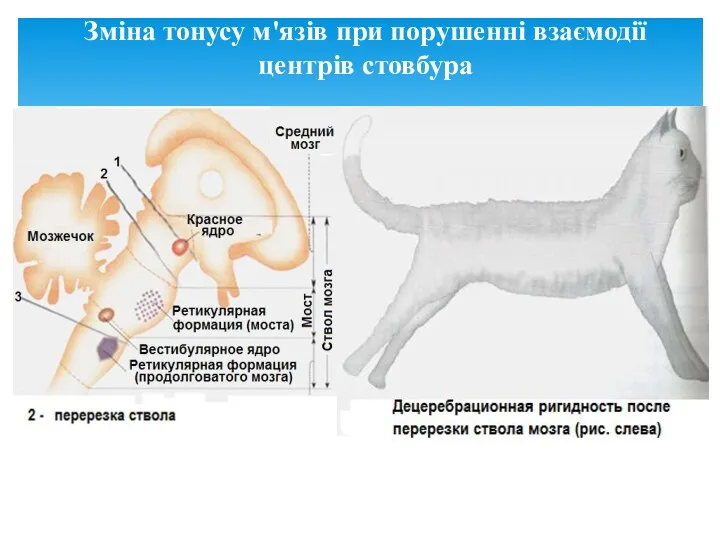
Зміна тонусу м'язів при порушенні взаємодії центрів стовбура
Зміна тонусу м'язів при порушенні взаємодії центрів стовбура

Рефлекси стовбура мозку
Позні рефлекси:
шийні,
вестибулярні,
очі-рухові.
Статокинетічеськие рефлекси:
статичні,
тонічні,
Орієнтовні.
Починаються вони з: пропріорецепторов
рецепторів м'язів шиї,
вестибулярного аналізатора.
Рефлекси стовбура мозку
Позні рефлекси:
шийні,
вестибулярні,
очі-рухові.
Статокинетічеськие рефлекси:
статичні,
тонічні,
Орієнтовні.
Починаються вони з: пропріорецепторов
рецепторів м'язів шиї,
вестибулярного аналізатора.

При «неправильному» положенні голови з пропріорецепторов м'язів шиї запускаються руху м'язів
При «неправильному» положенні голови з пропріорецепторов м'язів шиї запускаються руху м'язів

Прояв позних рефлексів у кішки при падінні
Починаються руху з повороту голови,
Прояв позних рефлексів у кішки при падінні
Починаються руху з повороту голови,

Перерозподіл тонусу м'язів кішки при нахилах голови
У тварин при нахилі голови
Перерозподіл тонусу м'язів кішки при нахилах голови
У тварин при нахилі голови

При нахилі голови вперед підвищується тонус м'язів-згиначів і в результаті покращується
При нахилі голови вперед підвищується тонус м'язів-згиначів і в результаті покращується

При повороті голови підвищується тонус м'язів згиначів на стороні повороту, а
При повороті голови підвищується тонус м'язів згиначів на стороні повороту, а
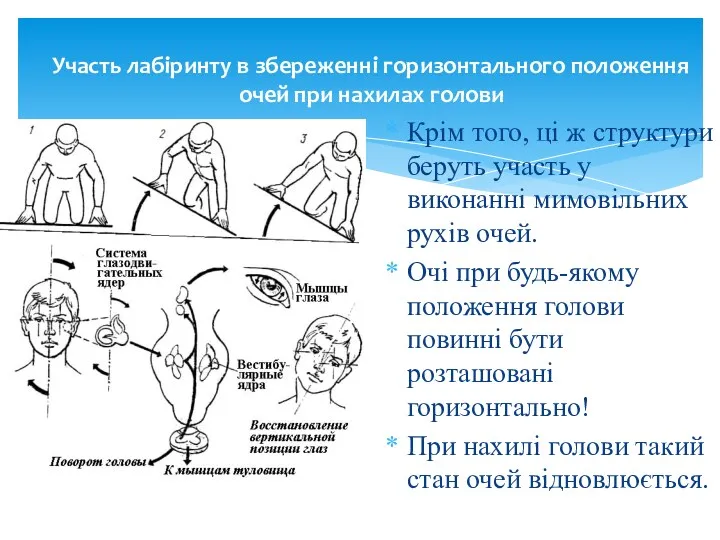
Участь лабіринту в збереженні горизонтального положення очей при нахилах голови
Крім того,
Участь лабіринту в збереженні горизонтального положення очей при нахилах голови
Крім того,
 Половое размножение.
Половое размножение. Методи розведення сільськогосподарських тварин, добір та підбір тварин
Методи розведення сільськогосподарських тварин, добір та підбір тварин Высокопроизводительные оmics-технологии, применяемые в системной биологии
Высокопроизводительные оmics-технологии, применяемые в системной биологии изменчивость
изменчивость Лекарственные и съедобные растения,произрастающие в Якутии
Лекарственные и съедобные растения,произрастающие в Якутии Проект по биологии, как вырастить годецию
Проект по биологии, как вырастить годецию Мастер класс по теме:Виды моделирования на уроках биологии
Мастер класс по теме:Виды моделирования на уроках биологии Аюлар
Аюлар Углеводы
Углеводы Везикулярлық тасымалдау
Везикулярлық тасымалдау Genetic enginnering
Genetic enginnering Клиническая биохимия надпочечников
Клиническая биохимия надпочечников Экстерьер и конституция собак
Экстерьер и конституция собак Биохимия печени
Биохимия печени Тип плоские черви
Тип плоские черви Конопля. Ботаническое описание
Конопля. Ботаническое описание Биология клетки
Биология клетки Цели и задачи биотехнологии. История развития биотехнологии. Слагаемые биотехнологического процесса
Цели и задачи биотехнологии. История развития биотехнологии. Слагаемые биотехнологического процесса Пищеварительная система. Полость рта, глотка, пищевод, желудок
Пищеварительная система. Полость рта, глотка, пищевод, желудок Хто такі комахи? Урок №118. Я досліджую світ
Хто такі комахи? Урок №118. Я досліджую світ Культивирование двустворчатых моллюсков мидий и их выращивание
Культивирование двустворчатых моллюсков мидий и их выращивание Удивительные свойства деревьев
Удивительные свойства деревьев Рыбы. Приспособленность к обитанию в водной среде. 1
Рыбы. Приспособленность к обитанию в водной среде. 1 Анализ красителей и консервантов, входящих в состав безалкогольных газированных и негазированных напитков
Анализ красителей и консервантов, входящих в состав безалкогольных газированных и негазированных напитков Рекордсмени в природі
Рекордсмени в природі Как звери готовятся к зиме
Как звери готовятся к зиме Правда ли, что растения кормят всех, даже хищников?
Правда ли, что растения кормят всех, даже хищников? Трансгенные животные
Трансгенные животные