- Нехромосомное наследование

Содержание
- 2. Генетика хлоропластов Материнский тип наследования Отцовский тип наследования Мутации пластидных генов у Chlamydomonas: Неспособность к фотосинтезу
- 4. Как определить внеядерную наследственность: Различия в результатах реципрокных скрещиваний Наличие связи между наследованием определенных признаков и
- 5. Цитоплазматическая мужская стерильность Мужская стерильность - явление появления форм, которые не образуют пыльцу или образуют пыльцу,
- 6. Восстановление фертильности пыльцы используют на практике для получения гетерозисных двойных межлинейных гибридов кукурузы
- 7. Гельминтоспориоз листьев
- 8. Генетика митохондрий Длина 19659 п.о. 13 белков аэробного дыхания, 22 тРНК, 2 типа рРНК
- 9. MERRF заболевание
- 10. MERRF
- 11. MELAS
- 12. Синдром Пирсона
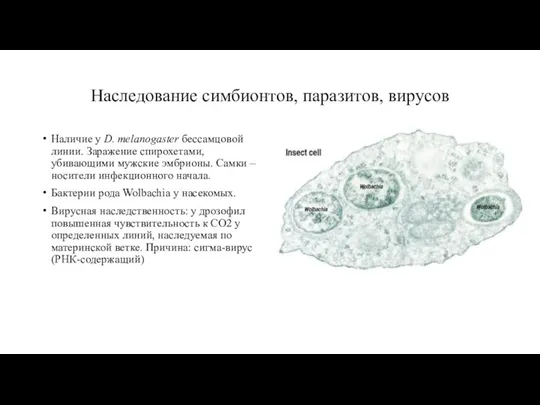
- 17. Наследование симбионтов, паразитов, вирусов Наличие у D. melanogaster бессамцовой линии. Заражение спирохетами, убивающими мужские эмбрионы. Самки
- 18. Собственно цитоплазматическое наследование Проявление признака – направление закручивания раковины у прудовика – как бы отстает на
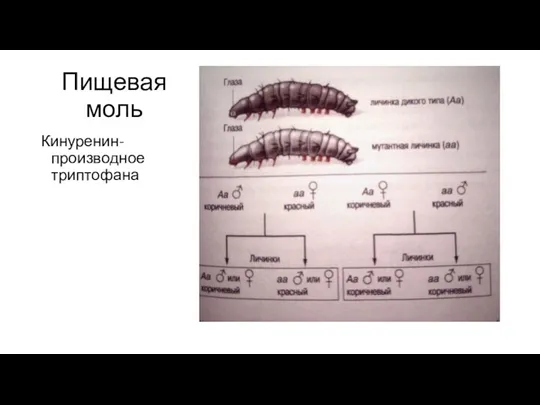
- 19. Пищевая моль Кинуренин-производное триптофана
- 20. Прионы В 1967 была предложена так называемая "белковая" гипотеза, согласно которой инфекционный агент, вызывающий эти заболевания
- 23. Скачать презентацию

Генетика хлоропластов
Материнский тип наследования
Отцовский тип наследования
Мутации пластидных генов у Chlamydomonas:
Неспособность к
Генетика хлоропластов
Материнский тип наследования
Отцовский тип наследования
Мутации пластидных генов у Chlamydomonas:
Неспособность к


Как определить внеядерную наследственность:
Различия в результатах реципрокных скрещиваний
Наличие связи между наследованием
Как определить внеядерную наследственность:
Различия в результатах реципрокных скрещиваний
Наличие связи между наследованием

Цитоплазматическая мужская стерильность
Мужская стерильность - явление появления форм, которые не образуют
Цитоплазматическая мужская стерильность
Мужская стерильность - явление появления форм, которые не образуют

Восстановление фертильности пыльцы используют на практике для получения гетерозисных двойных межлинейных
Восстановление фертильности пыльцы используют на практике для получения гетерозисных двойных межлинейных

Гельминтоспориоз листьев
Гельминтоспориоз листьев
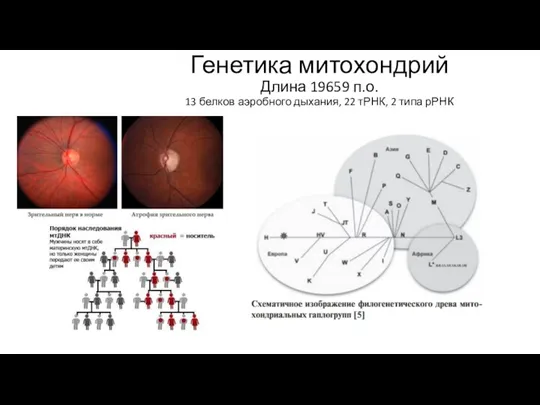
Генетика митохондрий
Длина 19659 п.о.
13 белков аэробного дыхания, 22 тРНК, 2
Генетика митохондрий Длина 19659 п.о. 13 белков аэробного дыхания, 22 тРНК, 2
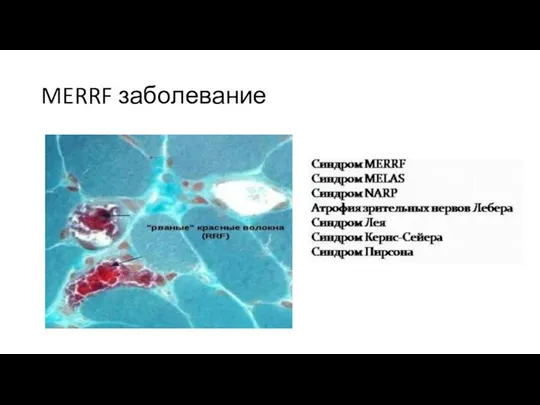
MERRF заболевание
MERRF заболевание
MERRF
MERRF

MELAS
MELAS
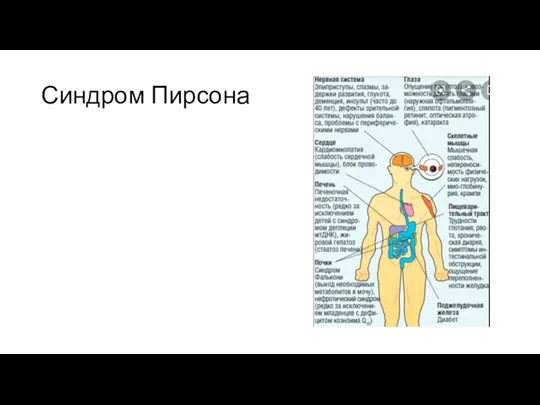
Синдром Пирсона
Синдром Пирсона


Наследование симбионтов, паразитов, вирусов
Наличие у D. melanogaster бессамцовой линии. Заражение спирохетами,
Наследование симбионтов, паразитов, вирусов
Наличие у D. melanogaster бессамцовой линии. Заражение спирохетами,

Собственно цитоплазматическое наследование
Проявление признака – направление закручивания раковины у прудовика –
Собственно цитоплазматическое наследование
Проявление признака – направление закручивания раковины у прудовика –
Пищевая моль
Кинуренин-производное триптофана
Пищевая моль
Кинуренин-производное триптофана

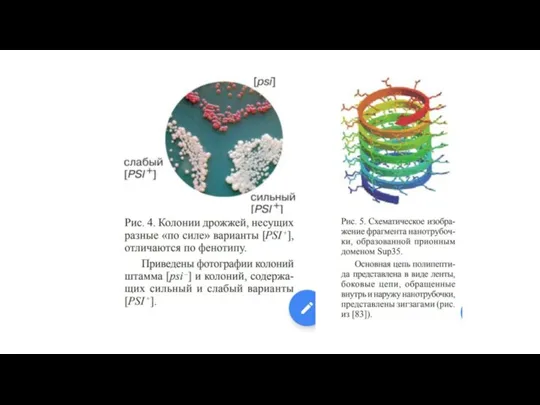
Прионы
В 1967 была предложена так называемая "белковая" гипотеза, согласно которой инфекционный
Прионы
В 1967 была предложена так называемая "белковая" гипотеза, согласно которой инфекционный
 Науки о человеке и их методы
Науки о человеке и их методы Семейство пингвины
Семейство пингвины Неделя экологии
Неделя экологии Презентация к викторине Мир пернатых.
Презентация к викторине Мир пернатых. Среда обитания организмов
Среда обитания организмов Зрительный анализатор
Зрительный анализатор Отделы головного мозга
Отделы головного мозга Травы и цветы полей и лугов
Травы и цветы полей и лугов Развитие проектно-исследовательской компетентности учащихся при изучении курса Решение проектных задач по биологии посредством освоения ими методов научного познания и умений учебно-исследовательской и проектной деятельности.
Развитие проектно-исследовательской компетентности учащихся при изучении курса Решение проектных задач по биологии посредством освоения ими методов научного познания и умений учебно-исследовательской и проектной деятельности. Экзотические плоды. Ягоды. Орехи
Экзотические плоды. Ягоды. Орехи Комбикорма. Основные факты, влияющие на продуктивность и здоровье птицы
Комбикорма. Основные факты, влияющие на продуктивность и здоровье птицы Происхождение птиц
Происхождение птиц Внешнее строение листа. Общие и отличительные признаки простых и сложных листьев
Внешнее строение листа. Общие и отличительные признаки простых и сложных листьев Световые и темновые реакции фотосинтеза
Световые и темновые реакции фотосинтеза Оборудование для культивирования микроорганизмов. Биореакторы непрерывного действия (лекция 3)
Оборудование для культивирования микроорганизмов. Биореакторы непрерывного действия (лекция 3) Дріжджі – це одноклітинні гриби
Дріжджі – це одноклітинні гриби Клетка как открытая метаболическая система
Клетка как открытая метаболическая система Ферменты-3
Ферменты-3 Родина гормону росту і пролактину (соматомамотропіни)
Родина гормону росту і пролактину (соматомамотропіни) Пищевые продукты, питательные вещества и их превращения в организме
Пищевые продукты, питательные вещества и их превращения в организме Өсімдіктер селекциясы
Өсімдіктер селекциясы Циклы споровых растений. Задачи на моногибридное скрещивание
Циклы споровых растений. Задачи на моногибридное скрещивание Что такое биоинформатика? Банк SwissProt
Что такое биоинформатика? Банк SwissProt Многообразие живых организмов. Царство Растения. 7 класс
Многообразие живых организмов. Царство Растения. 7 класс Отдел моховидные
Отдел моховидные Рост и развитие растений
Рост и развитие растений Обучение с использованием технологической карты.
Обучение с использованием технологической карты. Зеленые насаждения в городе
Зеленые насаждения в городе