- Нервная ткань

Содержание
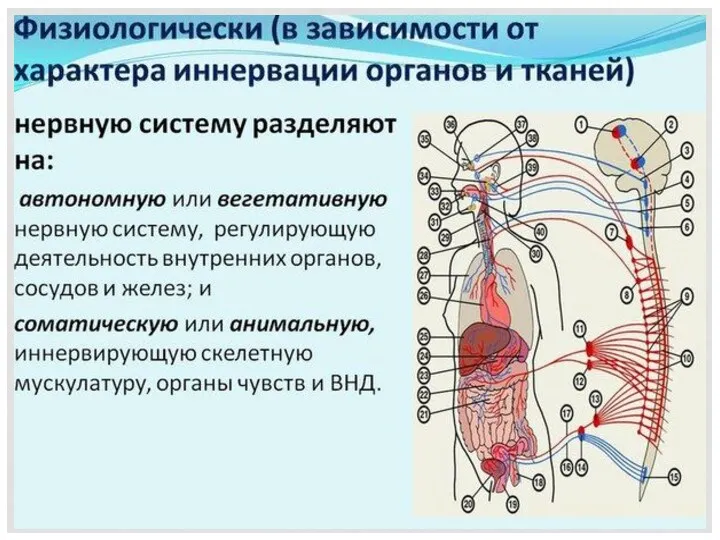
- 2. НЕРВНАЯ ТКАНЬ Нервная ткань — это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия
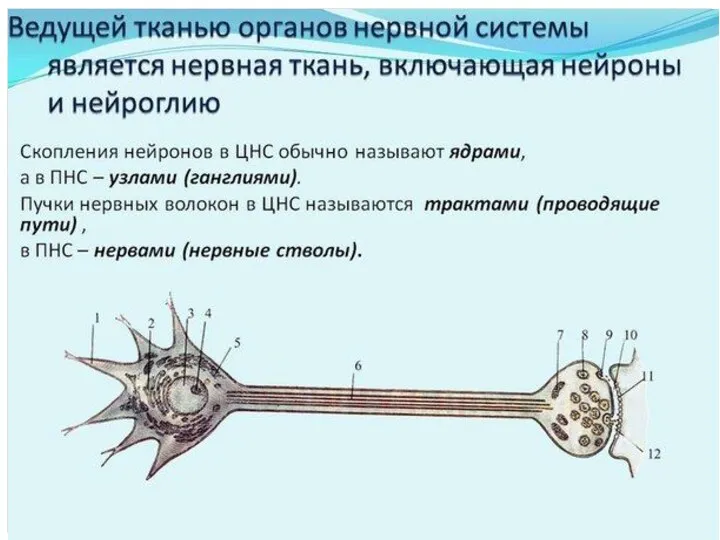
- 3. НЕРВНАЯ ТКАНЬ Нервные клетки (нейроны, нейроциты) — основные структурные компоненты нервной ткани, выполняющие специфическую функцию. Нейроглия
- 4. РАЗВИТИЕ Нервная ткань развивается из дорсальной эктодермы. У 18-дневного эмбриона человека эктодерма формирует нервную пластинку, латеральные
- 5. РАЗВИТИЕ Нервная ткань развивается из дорсальной эктодермы. У 18-дневного эмбриона человека эктодерма формирует нервную пластинку, латеральные
- 6. РАЗВИТИЕ
- 7. РАЗВИТИЕ Эмбриональные источники развития А. Нейроэктодерма Нервная пластинка ↓ Нервная трубка ↓ Ганглиозные пластинки - Нервные
- 8. НЕЙРОНЫ. Специализированные клетки нервной системы, ответственные за рецепцию, обработку стимулов, проведение импульса и влияние на другие
- 9. Нейроны Нейроны состоят из тела (перикариона) и отростков, среди которых выделяют дендриты и аксон (нейрит). Дендритов
- 10. В цитоплазме перикариона находятся хорошо развитые органеллы: ЭПС, комплекс Гольджи, митохондрии, лизосомы. Из элементов цитоскелета в
- 11. ТИГРОИДНОЕ ВЕЩЕТВО Хроматофильное вещество цитоплазмы (субстанция Ниссля, тигроид, базофильное вещество) проявляется при окрашивании нервных клеток основными
- 12. КЛАССИФИКАЦИЯ НЕЙРОНОВ Функциональная Чувствительные (рецепторные, сенсорные, афферентные, аффекторные) – на дендрите располагается рецептор, воспринимают раздражение и
- 13. КЛАССИФИКАЦИЯ НЕЙРОНОВ Нейроны состоят из тела и отростков: аксона и различного числа ветвящихся дендритов. По количеству
- 14. МОРФОЛОГИЯ НЕЙРОНА I
- 15. МОРФОЛОГИЯ НЕЙРОНА II
- 16. МОРФОЛОГИЯ НЕЙРОНА III
- 17. Классификация клеток нервной ткани
- 18. ГЛИОЦИТЫ (НЕЙРОГЛИЯ) Не проводят нервный импульс. Функции: опорная – поддержание тела и отростки нейронов, обеспечивая их
- 19. ВИДЫ ГЛИОЦИТОВ Использование методов импрегнации серебром и золотом по методу Рамон-и-Кахала и дель Рио-Ортега позволило подразделить
- 21. ВОЛОКНИСТЫЕ АСТРОЦИТЫ АСТРОЦИТЫ – присутствуют во всех отделах НС. Крупные к-ки со светлыми ядрами, небольшим количеством
- 22. ЭПЕНДИМОЦИТЫ Эпендимная глия выстилает центральный канал спинного мозга. Эпендимоциты участвуют в выработке спинномозговой жидкости (ликвора). От
- 23. Олигодендроциты II ОЛИГОДЕНДРОГЛИАЦИТЫ – относят олигодендроциты серого и белого в-ва спинного мозга, Шванновские к-ки (лемоциты), сотелиты
- 24. СВЯЗЬ АСТРОЦИТОВ С НЕЙРОНАМИ
- 25. МИКРОГЛИЯ Развивается из мезенхимы, относится к моноцито – макрофагальной системе. Мелкие отросчатые к-ки, разбросанные в сером
- 26. ВЗАИМООТНОШЕНИЯ НЕЙРОНОВ И ГЛИОЦИТОВ
- 27. В основе нервного волокна лежит отросток нервной клетки (чаще аксон) – осевой цилиндр. Каждое периферическое нервное
- 28. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА СТРОЕНИЯ МИЕЛИНОВЫХ И БЕЗМИЕЛИНОВЫХ НЕРВНЫХ ВОЛОКОН. СТРУКТУРНЫЕ ОСНОВЫ ПРОВЕДЕНИЯ НЕРВНОГО ИМПУЛЬСА ПО НЕРВНЫМ ВОЛОКНАМ
- 29. НЕМИЕЛИНИЗИРОВАННОЕ ВОЛОКНО Серые, не имеют миелиновой оболочки. Защищены шванновскими клетками: пучки волокон расположены так, что каждое
- 30. Миелин покрывает нервное волокно не сплошь, а прерывается через регулярные промежутки так называемыми перехватами Ранвье. В
- 31. ОБРАЗОВАНИЕ МИЕЛИНОВОГО ВОЛОКНА Глиоцит сначала обхватывает аксон, так что он оказывается лежащим в длинном желобке. Затем
- 32. Миелинизация в центральной и периферической нервной системах идет несколько разными механизмами. В периферической нервной системе шванновские
- 33. НЕРВНЫЕ ВОЛОКНА
- 34. НЕРВНЫЕ ВОЛОКНА
- 35. Нервные окончания
- 36. Чувствительные (рецепторные) нервные окончания представляют собой концевые аппараты дендритов афферентных нейронов, тела которых располагаются в спинальных,
- 37. Рецепторные окончания кожи
- 38. Клетка Меркеля (осязательный мениск)
- 39. Нервно-мышечное веретено
- 40. Эффекторные (двигательные) нервные окончания, или нейроорганные синапсы, имеются во всех тканях, обеспечивая передачу управляющего сигнала от
- 41. Иннервация скелетной мускулатуры I
- 42. Иннервация скелетной мускулатуры II
- 43. Моторная бляшка
- 44. Структура моторной бляшки
- 45. Функционирование моторной бляшки
- 46. НЕРВНЫЕ ОКОНЧАНИЯ
- 47. НЕРВНЫЕ ОКОНЧАНИЯ
- 48. СИНАПСЫ Если передача импульса совершается с помощью медиатора ацетилхолина, - синапсы называют холинергическими, если медиатором служит
- 49. СИНАПСЫ В зависимости от локализации окончаний терминальных веточек аксона, межнейрональные синапсы различают: аксо-дендритические - синапсы между
- 50. МЕЖНЕЙРОНАЛЬНЫЕ СИНАПСЫ Синапсы — это структуры, предназначенные для передачи импульса с одного нейрона на другой или
- 51. ХИМИЧЕСКИЕ СИНАПСЫ Передают импульс на другую клетку с помощью специальных биологически активных веществ — нейромедиаторов, находящихся
- 52. ХИМИЧЕСКИЕ СИНАПСЫ Химические синапсы передают импульс на другую клетку с помощью специальных биологически активных веществ —
- 53. ЭЛЕКТРИЧЕСКИЕ, ИЛИ ЭЛЕКТРОТОНИЧЕСКИЕ, СИНАПСЫ В нервной системе млекопитающих встречаются относительно редко. В области таких синапсов цитоплазмы
- 55. Электрический и химический типы синапса
- 56. Механизм синаптической передачи 1 . Волна деполяризации доходит до пресинаптической мембраны. 2. При этом открываются кальциевые
- 57. Нейрофибриллы образуют сеть в теле нейрона, а в отростках расположены параллельно. Нейротубулы и нейрофиламенты участвуют в
- 58. АКСОНАЛЬНЫЙ ТРАНСПОРТ Нейротубулы — органеллы, ответственные за быстрый транспорт, который называется также нейротубулозависимым. Каждая нейротубула содержит
- 59. НЕЙРОМЕДИАТОРЫ Отдельной разновидностью нейронов являются секреторные нейроны. Способность синтезировать и секретировать биологически активные вещества, в частности
- 60. НЕЙРОННАЯ ТЕОРИЯ КОНТАКТОВ Нейронная теория. Теория контакта, утверждающая, что нервная система построена из обособленных, контактирующих между
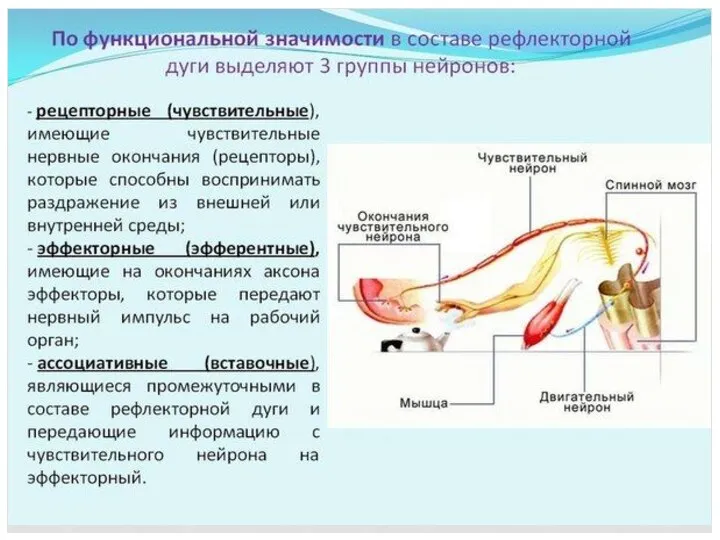
- 62. НЕЙРОНЫ В зависимости от функции в рефлекторной дуге различают рецепторные (чувствительные, афферентные), ассоциативные и эфферентные (эффекторные)
- 63. РЕФЛЕКТОРНЫЕ ДУГИ Наиболее простыми рефлекторными дугами являются двух- и трехнейронные, замыкающиеся на уровне одного сегмента спинного
- 65. РЕФЛЕКТОРНАЯ ДУГА ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ Рефлекторная дуга вегетативной нервной системы состоит из чувствительного (афферентного, сенсорного), ассоциативного
- 66. СОМАТИЧЕСКАЯ РД
- 67. 1-ый нейрон оканчивается в боковых рогах спинного мозга. 2-ой нейрон выходит за пределы сегмента и оканчиваются
- 68. СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА СТРУКТУРНОЙ ОРГАНИЗАЦИИ РЕФЛЕКТОРНЫХ ДУГ СОМАТИЧЕСКОЙ И ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ. Рефлекторная дуга — это цепь
- 69. КЛАССИФИКАЦИЯ НЕЙРОНОВ ПО МЕСТУ В РЕФЛЕКТОРНОЙ ДУГЕ
- 73. Гемато-энцефалический барьер включает в себя следующие компоненты: • эндотелий кровеносных капилляров (с непрерывной выстилкой) главный компонент
- 74. СПИННОЙ МОЗГ Спинной мозг состоит из 2-х половин, соединенных узкой полоской, содержащей центральный канал (остаток полости
- 75. РАЗВИТИЕ: В процессе развития спинного мозга из нервной трубки нейроны группируются в 10 слоях, или пластинах
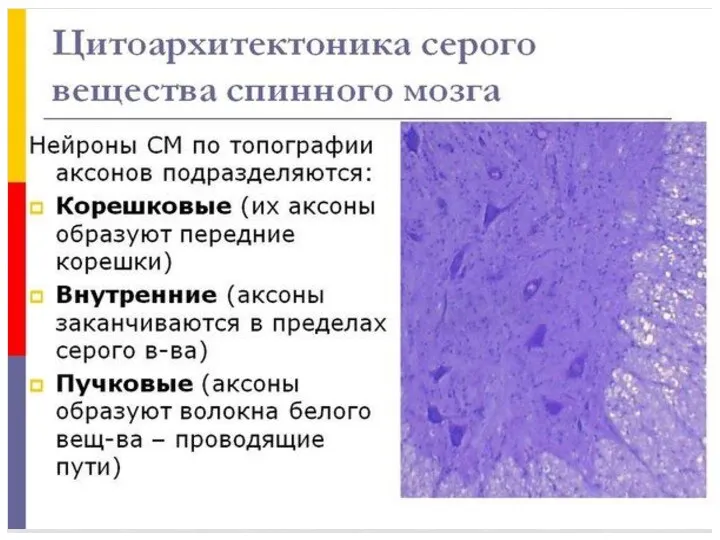
- 81. Среди нейронов спинного мозга можно выделить три вида клеток: *корешковые, *внутренние, *пучковые. Аксоны корешковых клеток покидают
- 82. В задних рогах различают: * губчатый слой, *желатинозное вещество, *собственное ядро заднего рога и *грудное ядро
- 83. Глия спинного мозга Основную часть глиального остова серого вещества составляют протоплазматические и волокнистые астроциты. Отростки волокнистых
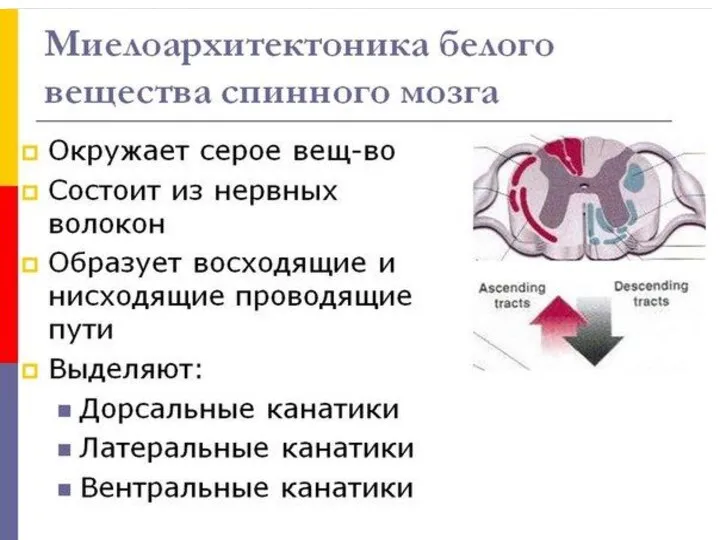
- 84. СЕРОЕ ВЕЩЕСТВО В сером веществе имеются выступы (столбы) получили название рога - с вентральной стороны обьемистые
- 86. СЕРОЕ ВЕЩЕСТВО К мягкой мозговой оболочке примыкает паутинная оболочка (arachnoidea), состоящая из рыхло расположенных пучков Соед.тк.,
- 87. СЕРОЕ ВЕЩЕСТВО Чувствительные клетки располагаются в спинальных ганглиях. Нейриты нервных клеток спинального ганглия вступают в спинной
- 88. СЕРОЕ И БЕЛОЕ ВЕЩЕСТВО ЦНС
- 89. Микроструктура серого вещества
- 93. РЕГЕНЕРАЦИЯ ЗАВИСИТ ОТ МЕСТА ТРАВМЫ Как в центральной, так и в периферической нервной системе погибшие нейроны
- 94. РЕАКЦИЯ НЕЙРОНОВ И ИХ ВОЛОКОН НА ТРАВМУ Перерезка нервного волокна вызывает различные реакции в теле нейрона,
- 95. ПОВРЕЖДЕННЫЕ НЕРВНЫЕ ВОЛОКНА ГОЛОВНОГО И СПИННОГО МОЗГА Поврежденные нервные волокна головного и спинного мозгане регенерируют. Возможно,
- 97. Скачать презентацию

НЕРВНАЯ ТКАНЬ
Нервная ткань — это система взаимосвязанных нервных клеток и нейроглии,
НЕРВНАЯ ТКАНЬ
Нервная ткань — это система взаимосвязанных нервных клеток и нейроглии,

НЕРВНАЯ ТКАНЬ
Нервные клетки (нейроны, нейроциты) — основные структурные компоненты нервной ткани,
НЕРВНАЯ ТКАНЬ
Нервные клетки (нейроны, нейроциты) — основные структурные компоненты нервной ткани,

РАЗВИТИЕ
Нервная ткань развивается из дорсальной эктодермы.
У 18-дневного эмбриона человека эктодерма
РАЗВИТИЕ
Нервная ткань развивается из дорсальной эктодермы.
У 18-дневного эмбриона человека эктодерма
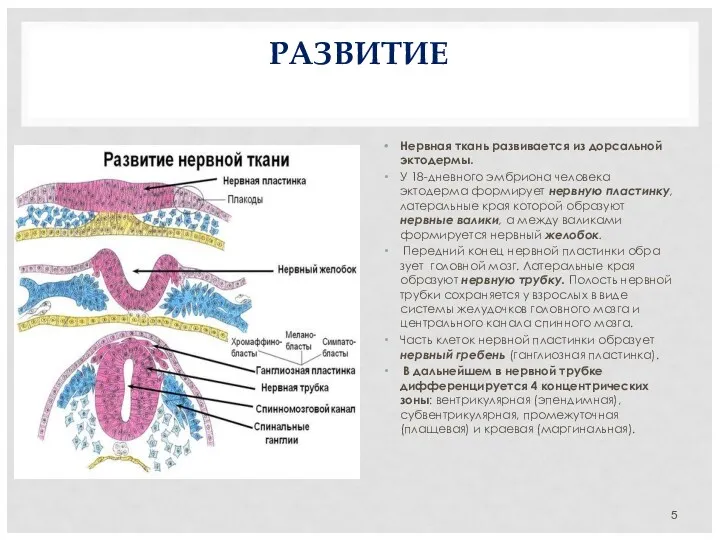
РАЗВИТИЕ
Нервная ткань развивается из дорсальной эктодермы.
У 18-дневного эмбриона человека эктодерма
РАЗВИТИЕ
Нервная ткань развивается из дорсальной эктодермы.
У 18-дневного эмбриона человека эктодерма
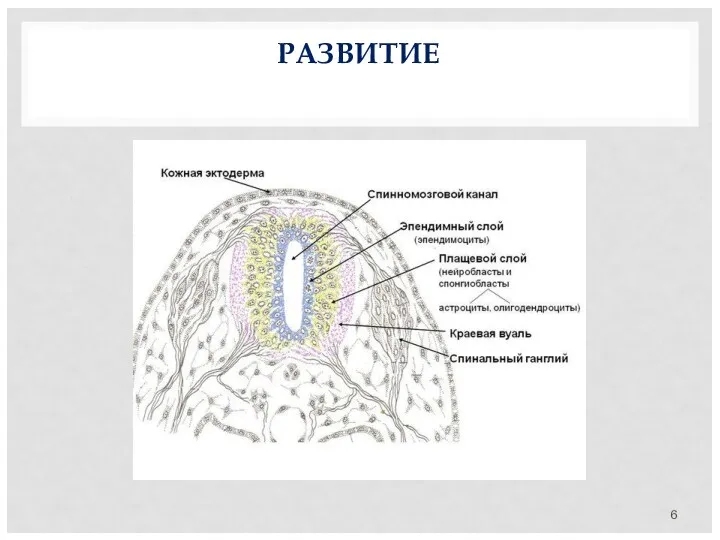
РАЗВИТИЕ
РАЗВИТИЕ

РАЗВИТИЕ
Эмбриональные источники развития
А. Нейроэктодерма
Нервная пластинка
↓
Нервная трубка
↓
РАЗВИТИЕ
Эмбриональные источники развития
А. Нейроэктодерма
Нервная пластинка
↓
Нервная трубка
↓

НЕЙРОНЫ.
Специализированные клетки нервной системы, ответственные за рецепцию, обработку стимулов, проведение импульса
НЕЙРОНЫ.
Специализированные клетки нервной системы, ответственные за рецепцию, обработку стимулов, проведение импульса
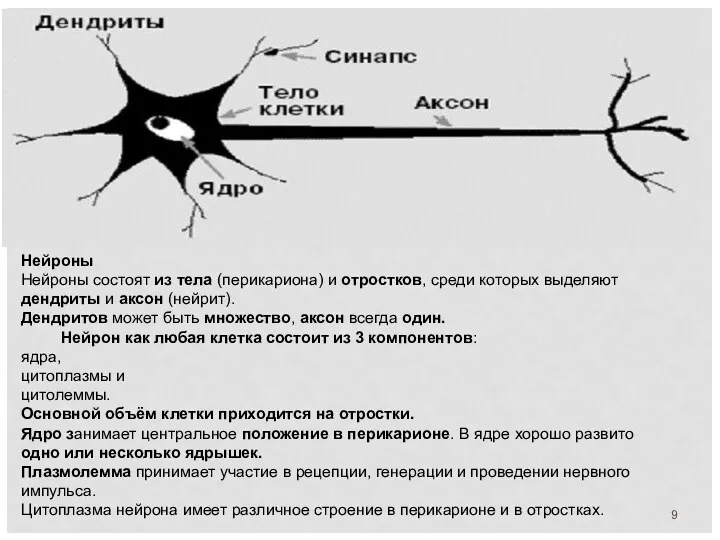
Нейроны
Нейроны состоят из тела (перикариона) и отростков, среди которых выделяют дендриты
Нейроны
Нейроны состоят из тела (перикариона) и отростков, среди которых выделяют дендриты
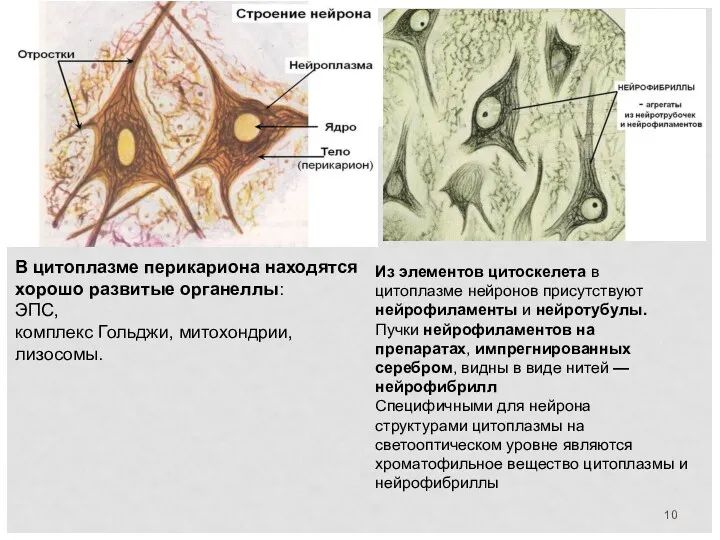
В цитоплазме перикариона находятся хорошо развитые органеллы:
ЭПС,
комплекс Гольджи, митохондрии,
В цитоплазме перикариона находятся хорошо развитые органеллы:
ЭПС,
комплекс Гольджи, митохондрии,
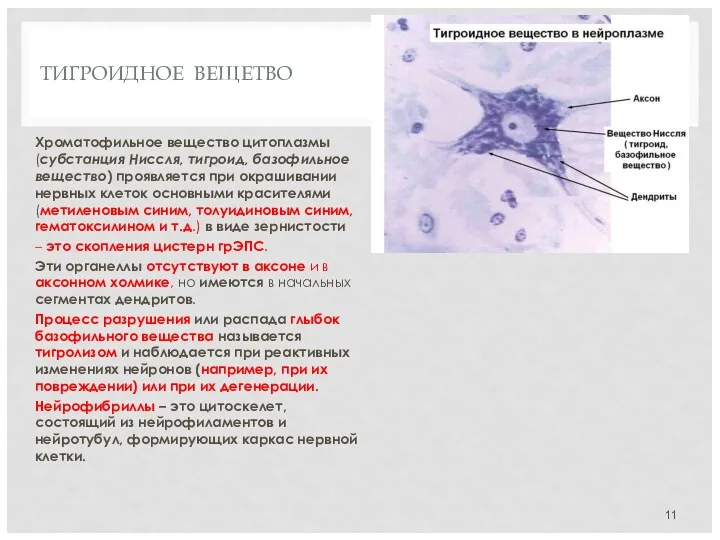
ТИГРОИДНОЕ ВЕЩЕТВО
Хроматофильное вещество цитоплазмы (субстанция Ниссля, тигроид, базофильное вещество) проявляется при
ТИГРОИДНОЕ ВЕЩЕТВО
Хроматофильное вещество цитоплазмы (субстанция Ниссля, тигроид, базофильное вещество) проявляется при
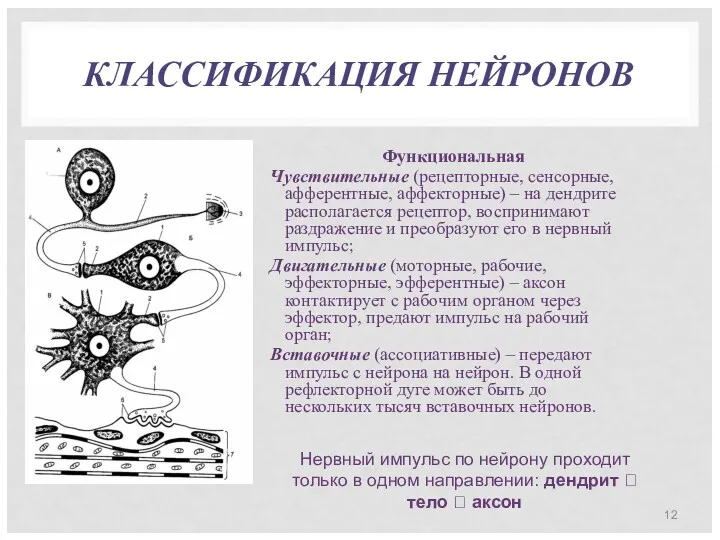
КЛАССИФИКАЦИЯ НЕЙРОНОВ
Функциональная
Чувствительные (рецепторные, сенсорные, афферентные, аффекторные) – на дендрите располагается рецептор,
КЛАССИФИКАЦИЯ НЕЙРОНОВ
Функциональная
Чувствительные (рецепторные, сенсорные, афферентные, аффекторные) – на дендрите располагается рецептор,
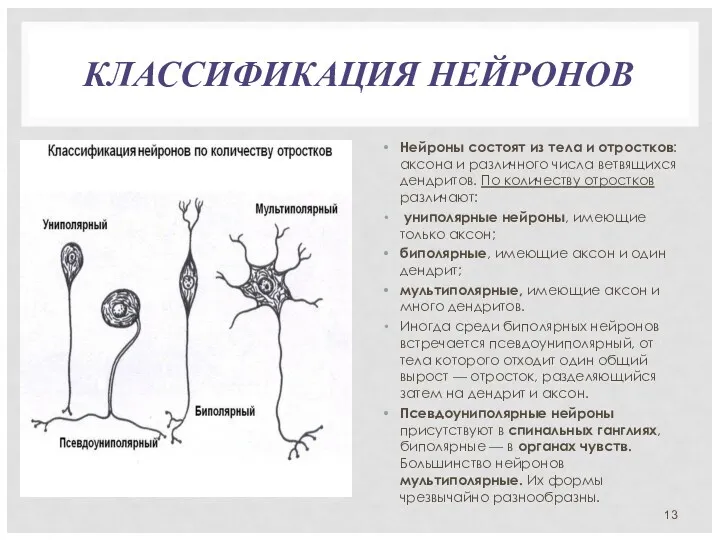
КЛАССИФИКАЦИЯ НЕЙРОНОВ
Нейроны состоят из тела и отростков: аксона и различного числа
КЛАССИФИКАЦИЯ НЕЙРОНОВ
Нейроны состоят из тела и отростков: аксона и различного числа

МОРФОЛОГИЯ НЕЙРОНА I
МОРФОЛОГИЯ НЕЙРОНА I
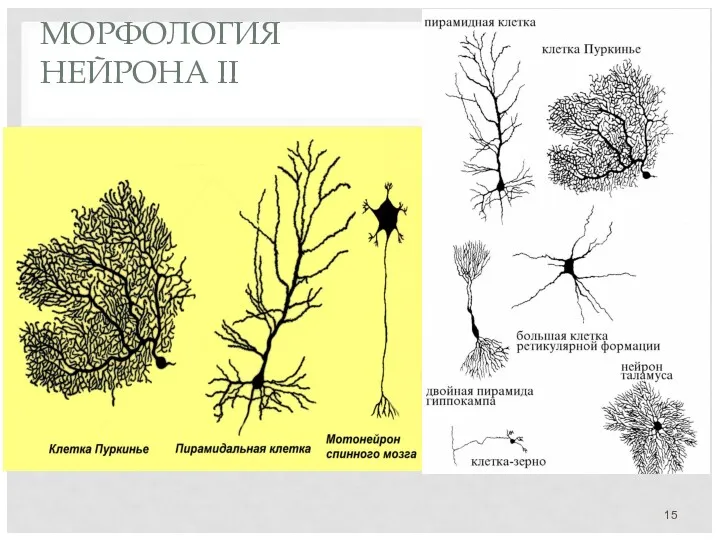
МОРФОЛОГИЯ
НЕЙРОНА II
МОРФОЛОГИЯ
НЕЙРОНА II
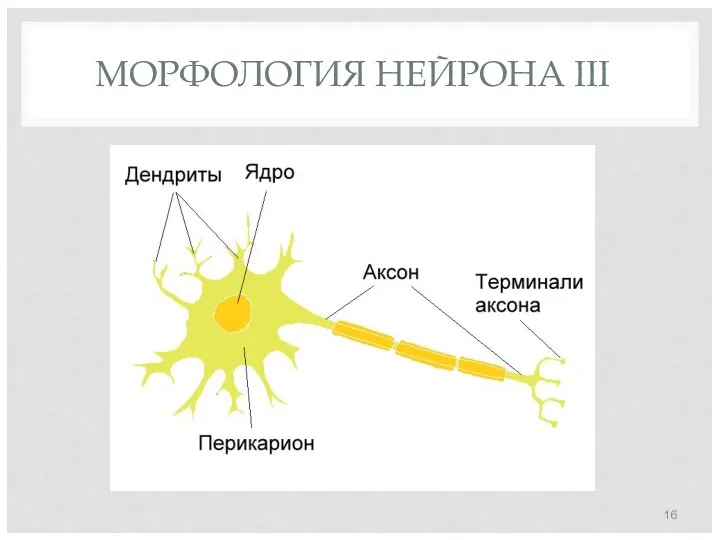
МОРФОЛОГИЯ НЕЙРОНА III
МОРФОЛОГИЯ НЕЙРОНА III
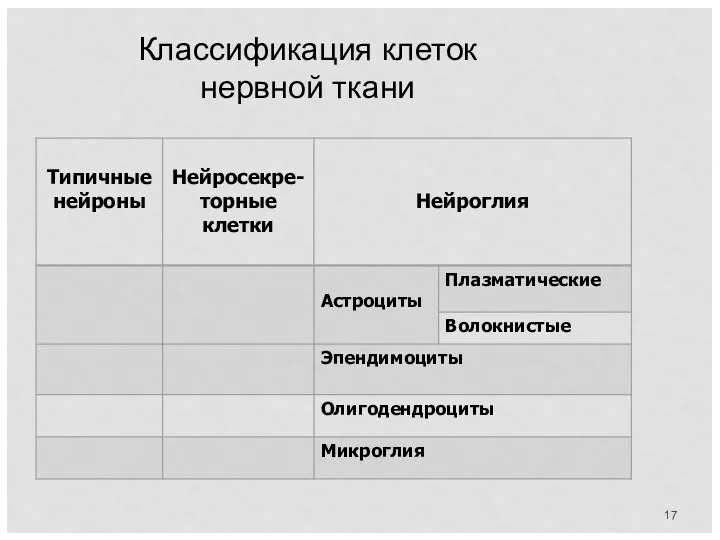
Классификация клеток нервной ткани
Классификация клеток нервной ткани
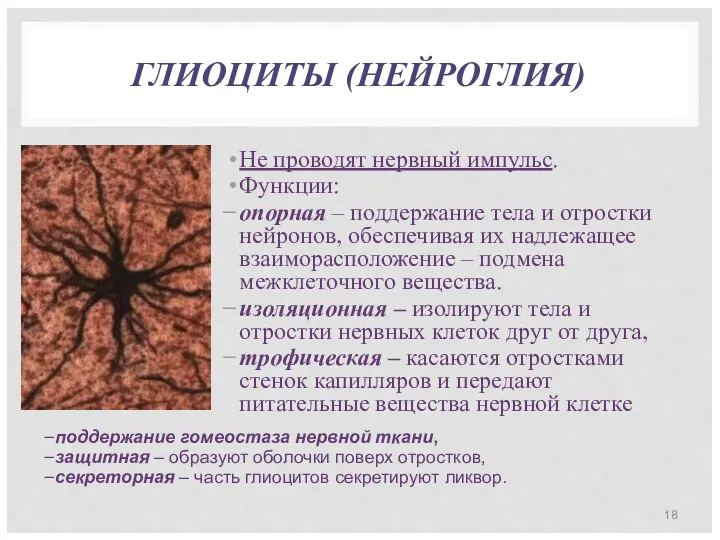
ГЛИОЦИТЫ (НЕЙРОГЛИЯ)
Не проводят нервный импульс.
Функции:
опорная – поддержание тела и
ГЛИОЦИТЫ (НЕЙРОГЛИЯ)
Не проводят нервный импульс.
Функции:
опорная – поддержание тела и
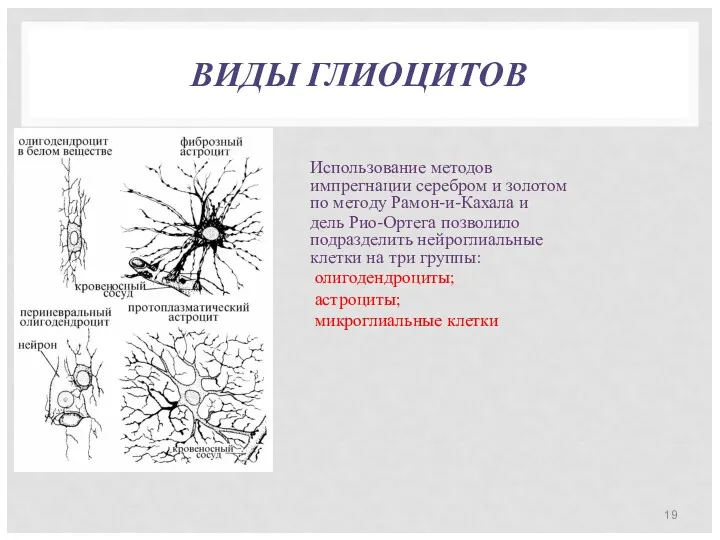
ВИДЫ ГЛИОЦИТОВ
Использование методов импрегнации серебром и золотом по методу Рамон-и-Кахала и
ВИДЫ ГЛИОЦИТОВ
Использование методов импрегнации серебром и золотом по методу Рамон-и-Кахала и
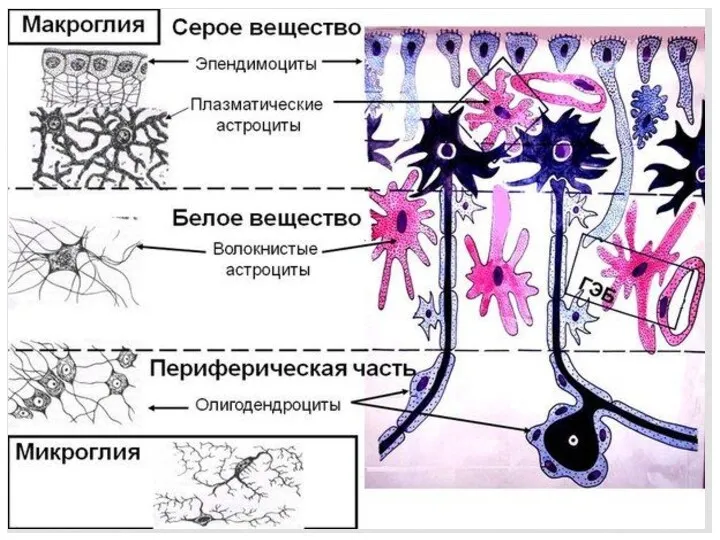
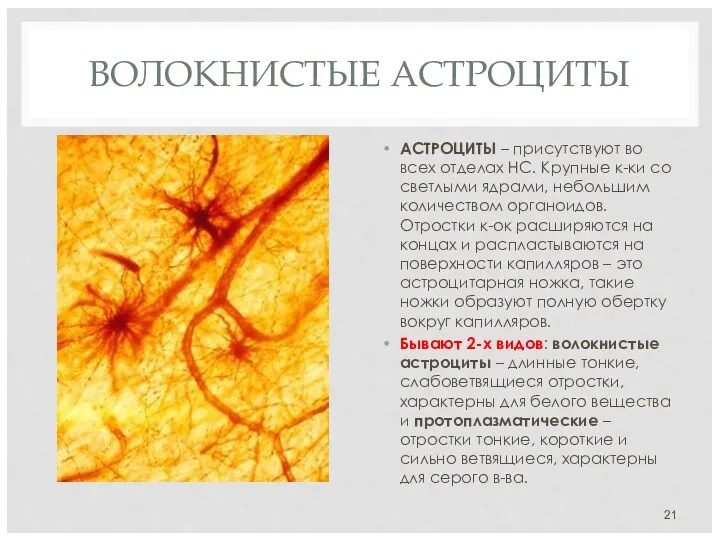
ВОЛОКНИСТЫЕ АСТРОЦИТЫ
АСТРОЦИТЫ – присутствуют во всех отделах НС. Крупные к-ки со
ВОЛОКНИСТЫЕ АСТРОЦИТЫ
АСТРОЦИТЫ – присутствуют во всех отделах НС. Крупные к-ки со

ЭПЕНДИМОЦИТЫ
Эпендимная глия выстилает центральный канал спинного мозга. Эпендимоциты участвуют в выработке
ЭПЕНДИМОЦИТЫ
Эпендимная глия выстилает центральный канал спинного мозга. Эпендимоциты участвуют в выработке
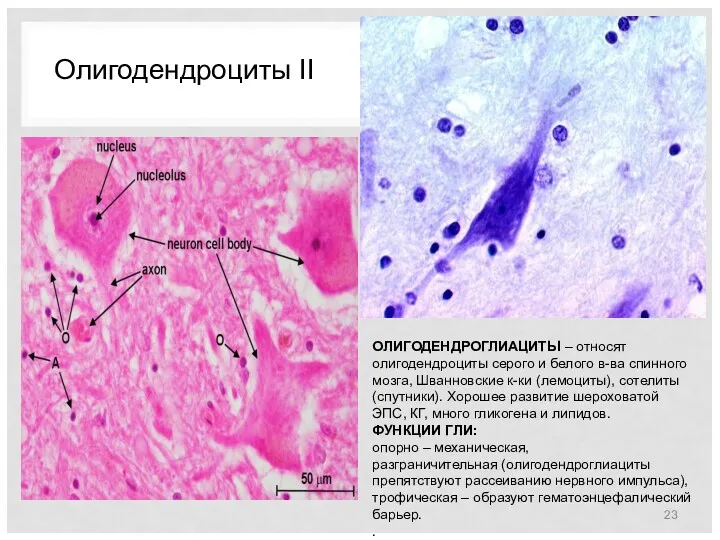
Олигодендроциты II
ОЛИГОДЕНДРОГЛИАЦИТЫ – относят олигодендроциты серого и белого в-ва спинного
Олигодендроциты II
ОЛИГОДЕНДРОГЛИАЦИТЫ – относят олигодендроциты серого и белого в-ва спинного
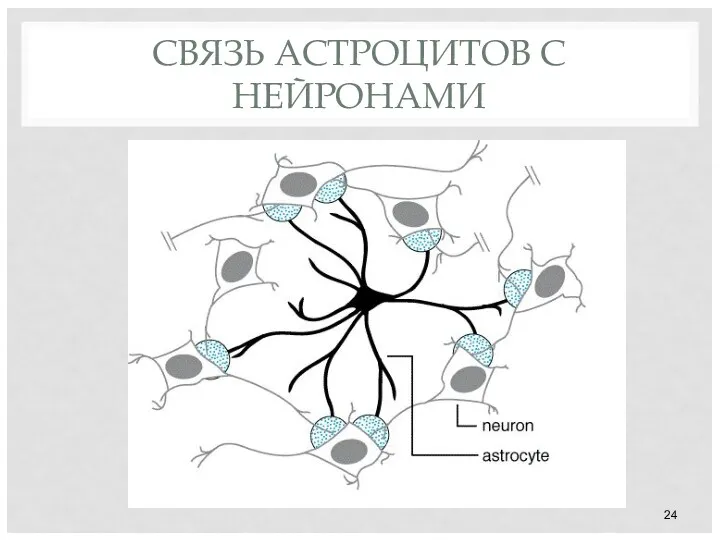
СВЯЗЬ АСТРОЦИТОВ С НЕЙРОНАМИ
СВЯЗЬ АСТРОЦИТОВ С НЕЙРОНАМИ
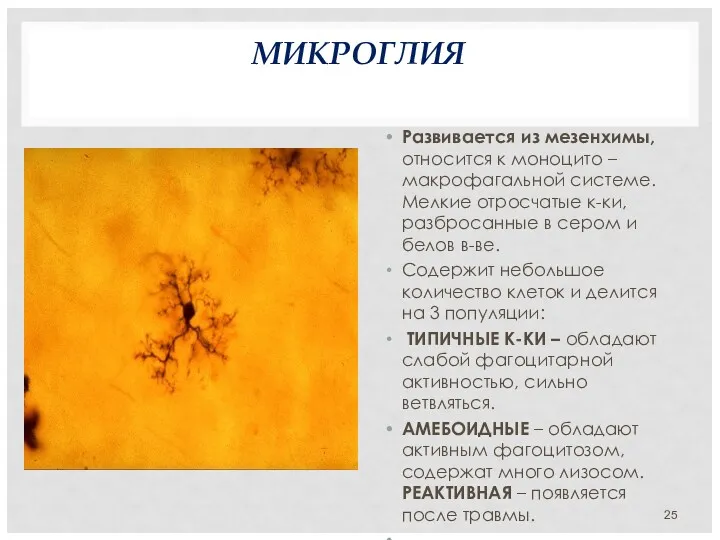
МИКРОГЛИЯ
Развивается из мезенхимы, относится к моноцито – макрофагальной системе. Мелкие отросчатые
МИКРОГЛИЯ
Развивается из мезенхимы, относится к моноцито – макрофагальной системе. Мелкие отросчатые
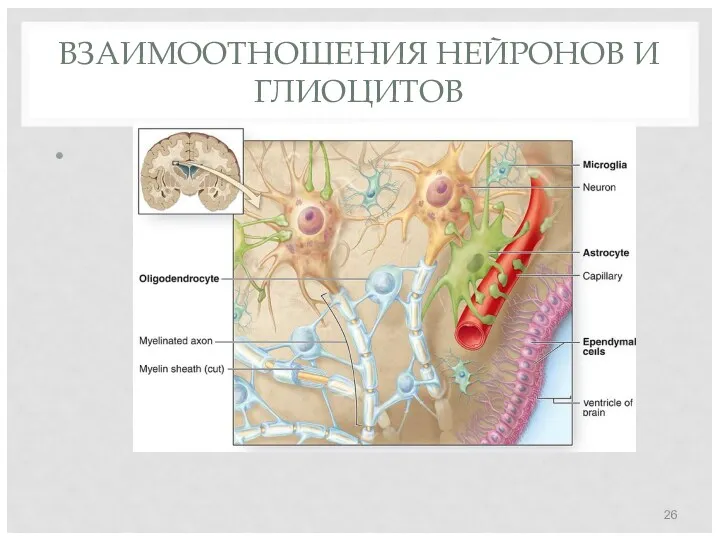
ВЗАИМООТНОШЕНИЯ НЕЙРОНОВ И ГЛИОЦИТОВ
ВЗАИМООТНОШЕНИЯ НЕЙРОНОВ И ГЛИОЦИТОВ
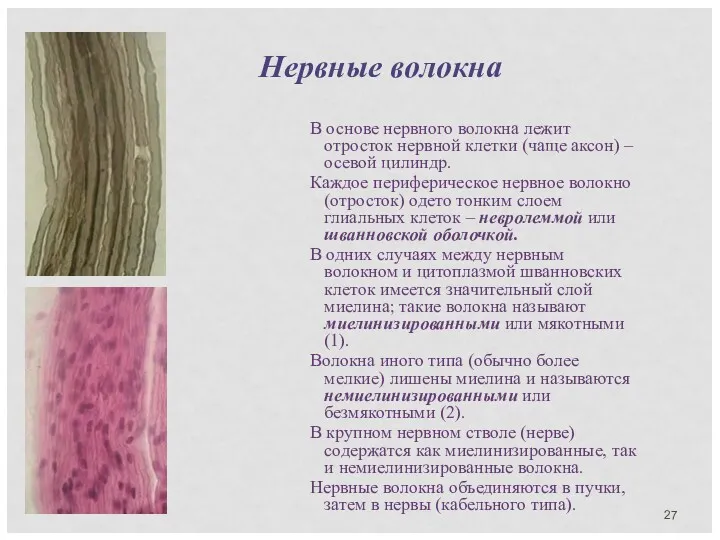
В основе нервного волокна лежит отросток нервной клетки (чаще аксон) –
В основе нервного волокна лежит отросток нервной клетки (чаще аксон) –
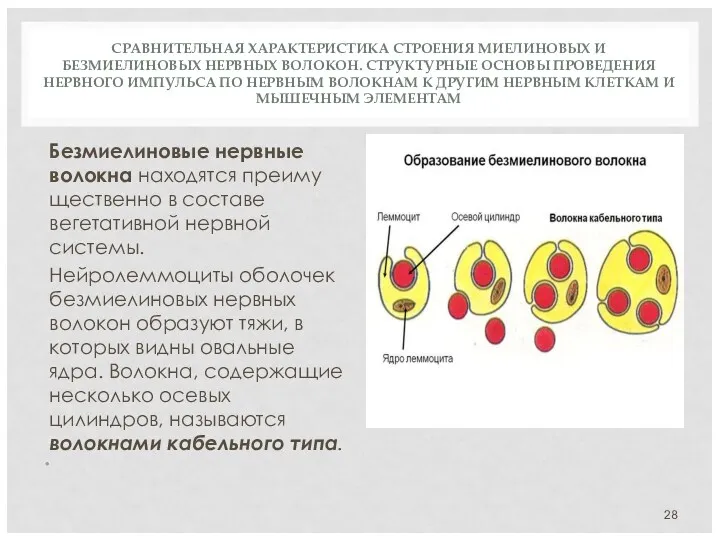
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА СТРОЕНИЯ МИЕЛИНОВЫХ И БЕЗМИЕЛИНОВЫХ НЕРВНЫХ ВОЛОКОН. СТРУКТУРНЫЕ ОСНОВЫ ПРОВЕДЕНИЯ
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА СТРОЕНИЯ МИЕЛИНОВЫХ И БЕЗМИЕЛИНОВЫХ НЕРВНЫХ ВОЛОКОН. СТРУКТУРНЫЕ ОСНОВЫ ПРОВЕДЕНИЯ
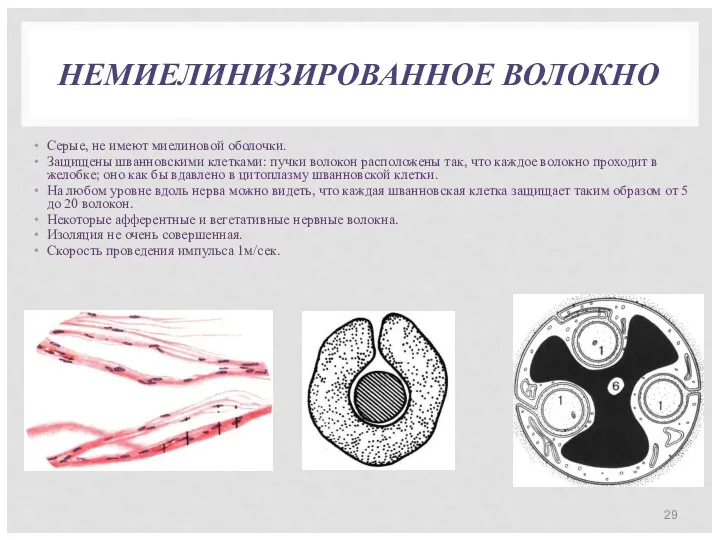
НЕМИЕЛИНИЗИРОВАННОЕ ВОЛОКНО
Серые, не имеют миелиновой оболочки.
Защищены шванновскими клетками: пучки волокон
НЕМИЕЛИНИЗИРОВАННОЕ ВОЛОКНО
Серые, не имеют миелиновой оболочки.
Защищены шванновскими клетками: пучки волокон
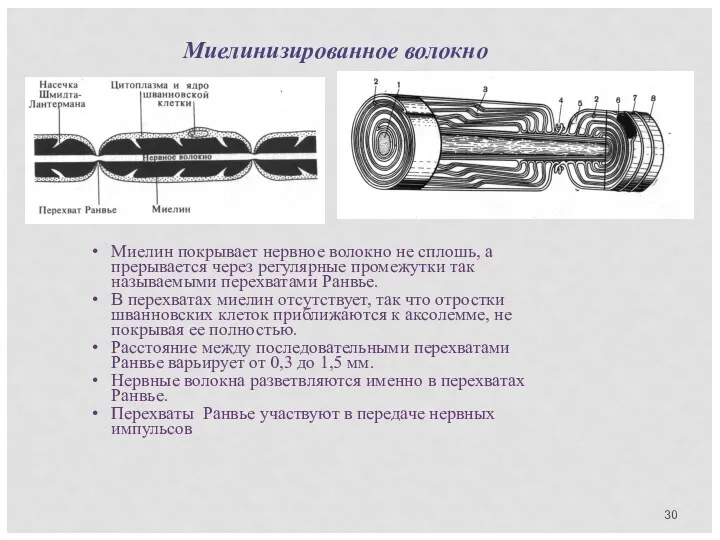
Миелин покрывает нервное волокно не сплошь, а прерывается через регулярные промежутки
Миелин покрывает нервное волокно не сплошь, а прерывается через регулярные промежутки
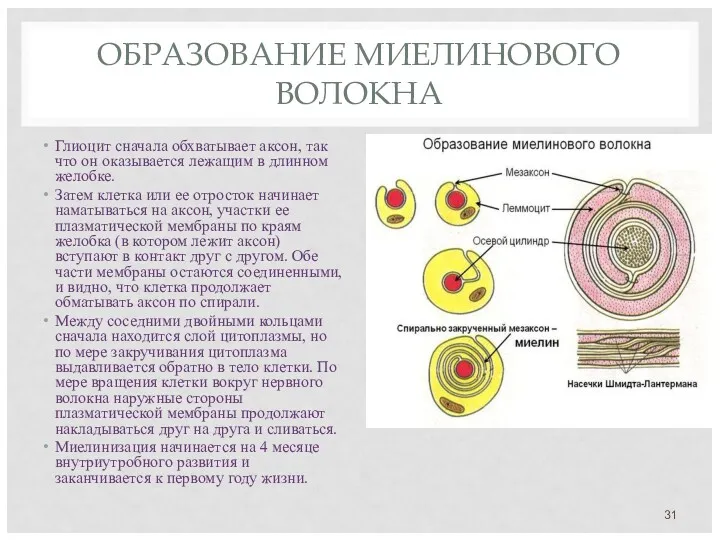
ОБРАЗОВАНИЕ МИЕЛИНОВОГО ВОЛОКНА
Глиоцит сначала обхватывает аксон, так что он оказывается лежащим
ОБРАЗОВАНИЕ МИЕЛИНОВОГО ВОЛОКНА
Глиоцит сначала обхватывает аксон, так что он оказывается лежащим
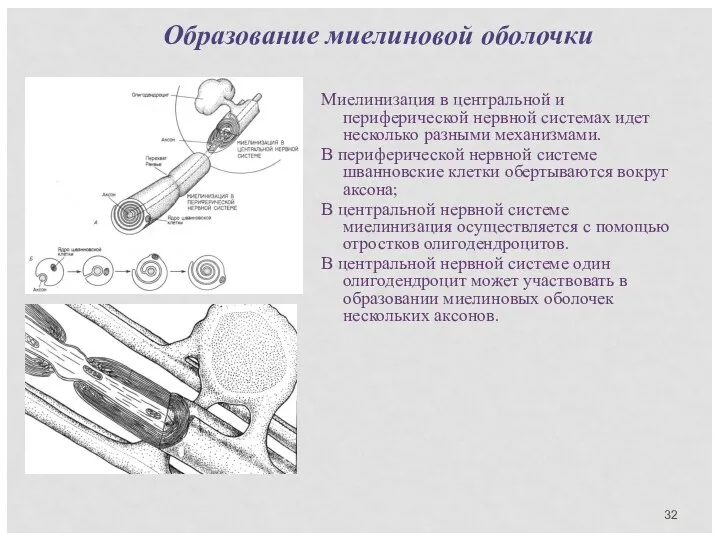
Миелинизация в центральной и периферической нервной системах идет несколько разными механизмами.
В
Миелинизация в центральной и периферической нервной системах идет несколько разными механизмами.
В
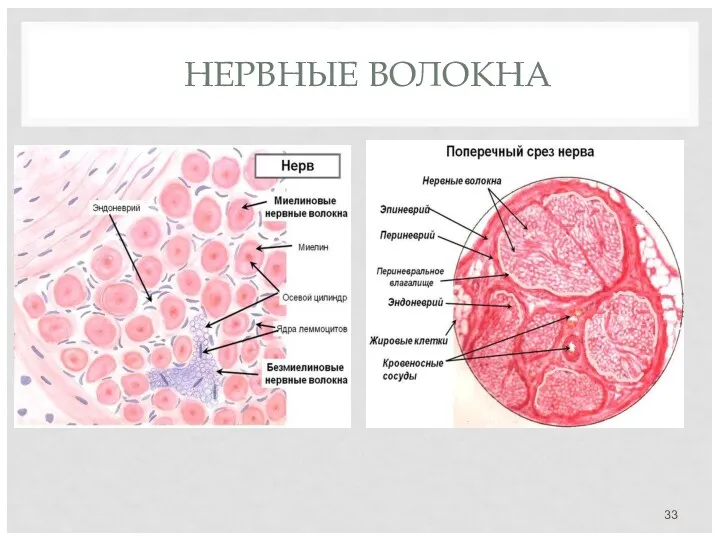
НЕРВНЫЕ ВОЛОКНА
НЕРВНЫЕ ВОЛОКНА
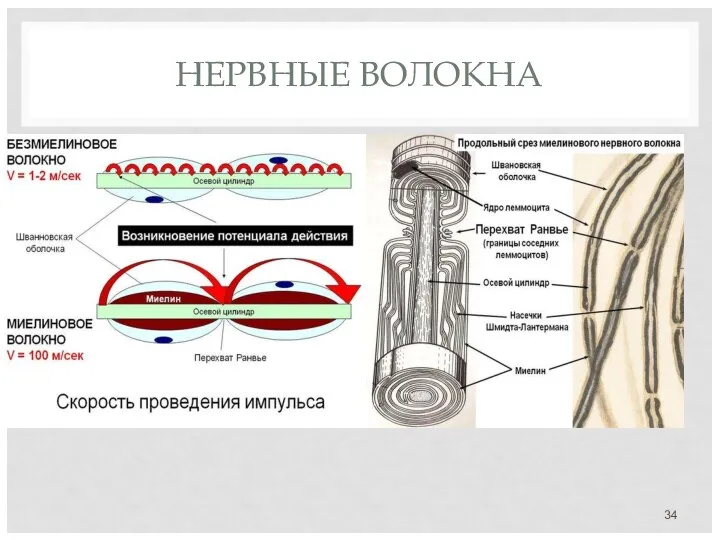
НЕРВНЫЕ ВОЛОКНА
НЕРВНЫЕ ВОЛОКНА
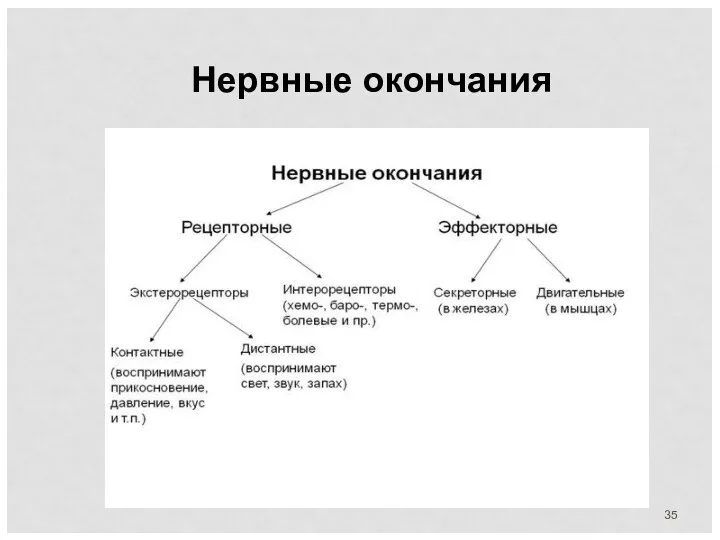
Нервные окончания
Нервные окончания

Чувствительные (рецепторные) нервные окончания представляют собой концевые аппараты дендритов афферентных нейронов,
Чувствительные (рецепторные) нервные окончания представляют собой концевые аппараты дендритов афферентных нейронов,
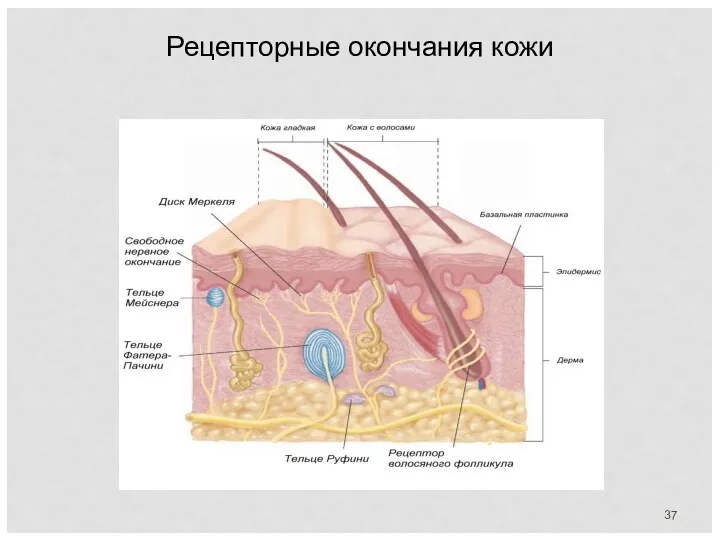
Рецепторные окончания кожи
Рецепторные окончания кожи
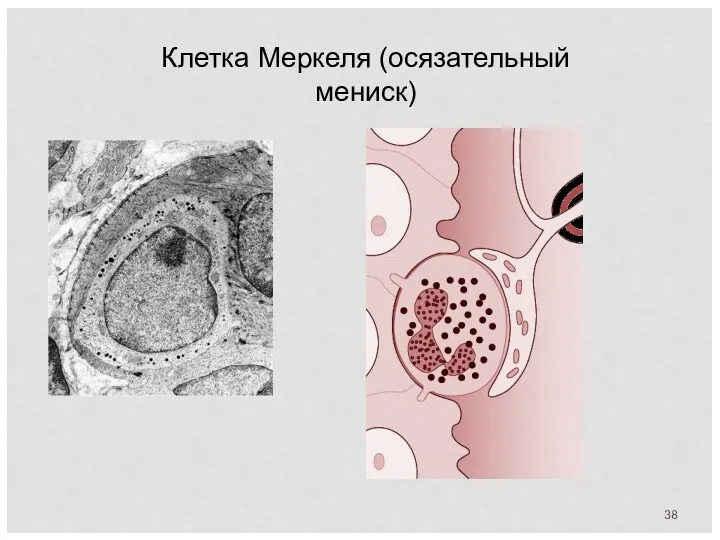
Клетка Меркеля (осязательный мениск)
Клетка Меркеля (осязательный мениск)
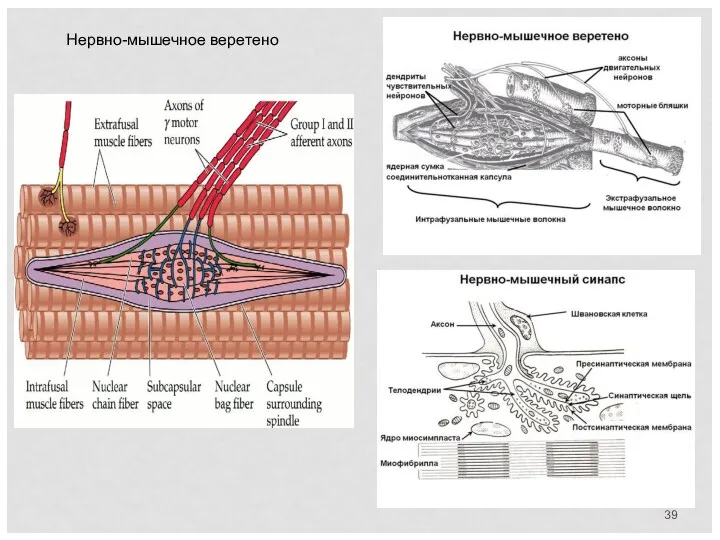
Нервно-мышечное веретено
Нервно-мышечное веретено

Эффекторные (двигательные) нервные окончания, или нейроорганные синапсы, имеются во всех
Эффекторные (двигательные) нервные окончания, или нейроорганные синапсы, имеются во всех
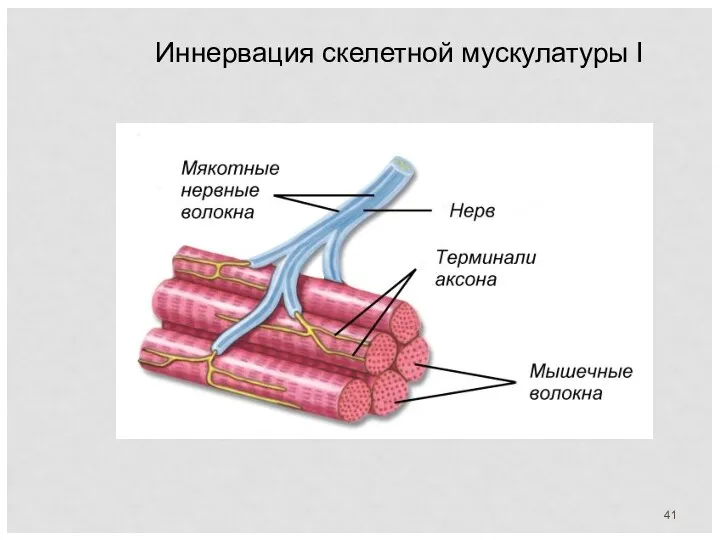
Иннервация скелетной мускулатуры I
Иннервация скелетной мускулатуры I
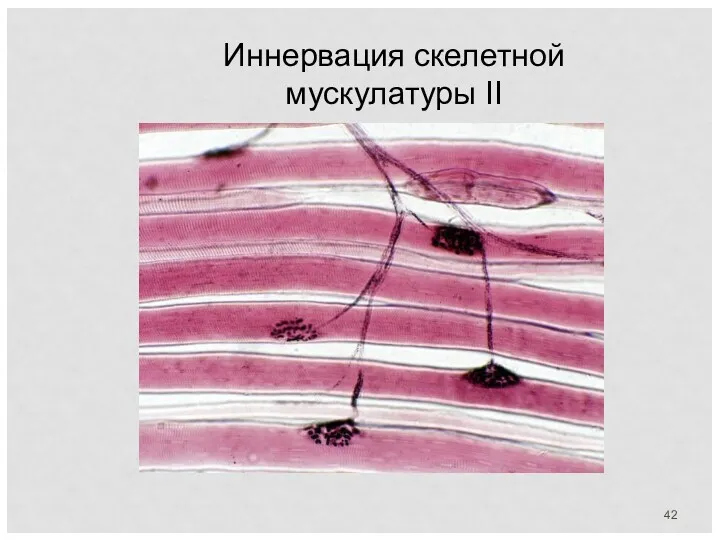
Иннервация скелетной мускулатуры II
Иннервация скелетной мускулатуры II
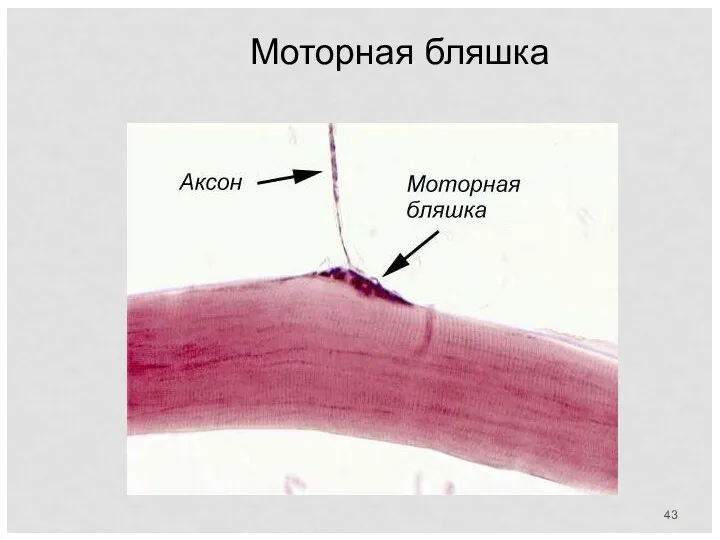
Моторная бляшка
Моторная бляшка
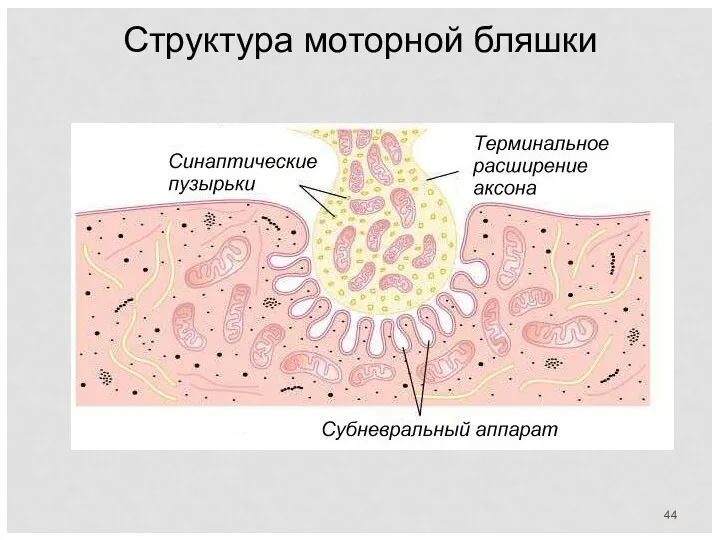
Структура моторной бляшки
Структура моторной бляшки
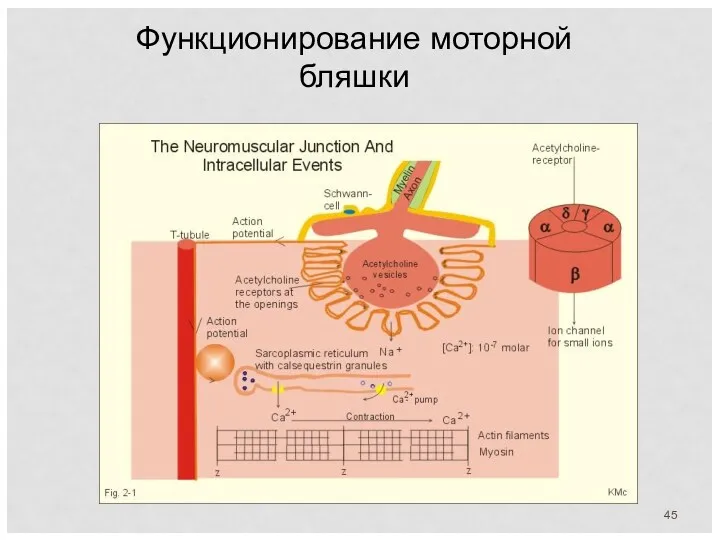
Функционирование моторной бляшки
Функционирование моторной бляшки
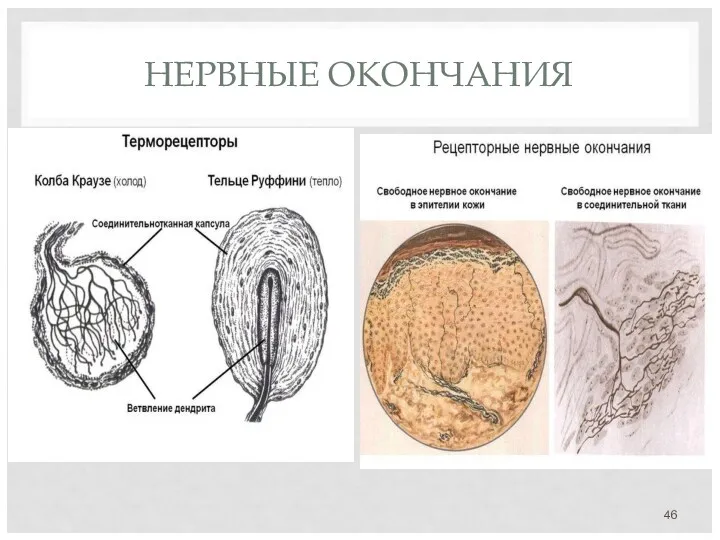
НЕРВНЫЕ ОКОНЧАНИЯ
НЕРВНЫЕ ОКОНЧАНИЯ
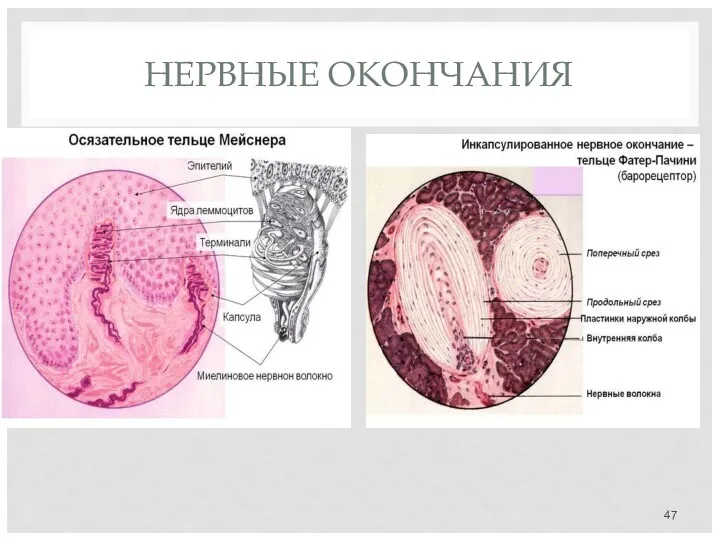
НЕРВНЫЕ ОКОНЧАНИЯ
НЕРВНЫЕ ОКОНЧАНИЯ

СИНАПСЫ
Если передача импульса совершается с помощью медиатора ацетилхолина, - синапсы называют
СИНАПСЫ
Если передача импульса совершается с помощью медиатора ацетилхолина, - синапсы называют
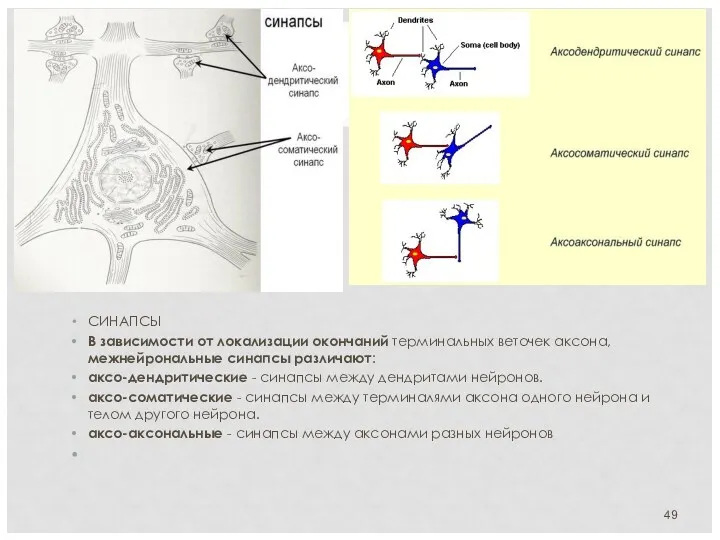
СИНАПСЫ
В зависимости от локализации окончаний терминальных веточек аксона, межнейрональные синапсы
СИНАПСЫ
В зависимости от локализации окончаний терминальных веточек аксона, межнейрональные синапсы

МЕЖНЕЙРОНАЛЬНЫЕ СИНАПСЫ
Синапсы — это структуры, предназначенные для передачи импульса с одного
МЕЖНЕЙРОНАЛЬНЫЕ СИНАПСЫ
Синапсы — это структуры, предназначенные для передачи импульса с одного
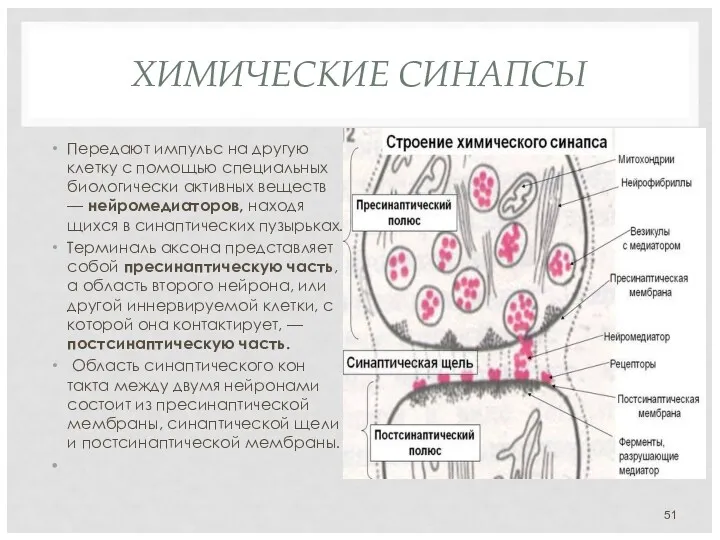
ХИМИЧЕСКИЕ СИНАПСЫ
Передают импульс на другую клетку с помощью специальных биологически
ХИМИЧЕСКИЕ СИНАПСЫ
Передают импульс на другую клетку с помощью специальных биологически

ХИМИЧЕСКИЕ СИНАПСЫ
Химические синапсы передают импульс на другую клетку с помощью
ХИМИЧЕСКИЕ СИНАПСЫ
Химические синапсы передают импульс на другую клетку с помощью

ЭЛЕКТРИЧЕСКИЕ, ИЛИ ЭЛЕКТРОТОНИЧЕСКИЕ, СИНАПСЫ
В нервной системе млекопитающих встречаются относительно редко.
ЭЛЕКТРИЧЕСКИЕ, ИЛИ ЭЛЕКТРОТОНИЧЕСКИЕ, СИНАПСЫ
В нервной системе млекопитающих встречаются относительно редко.
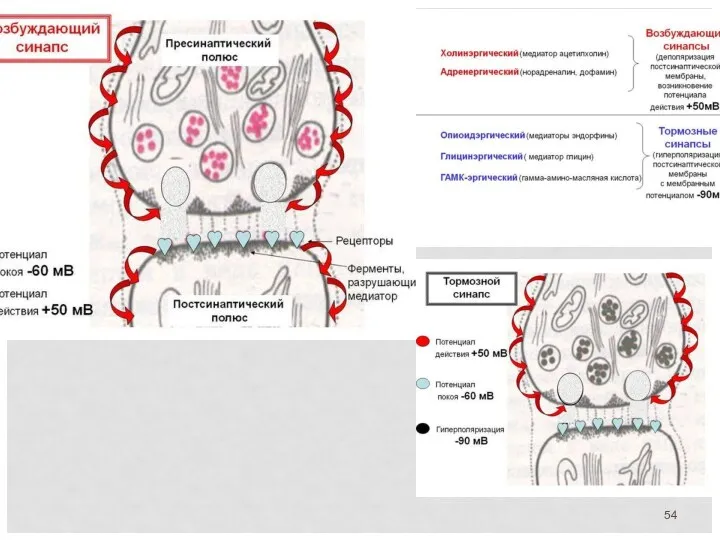
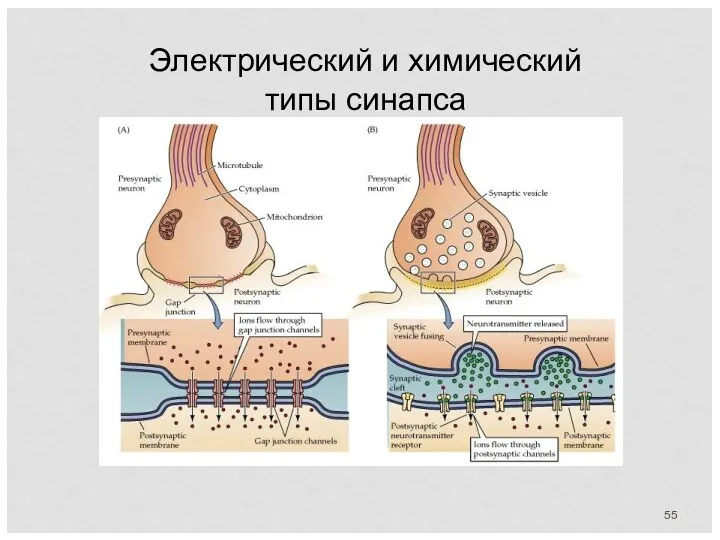
Электрический и химический типы синапса
Электрический и химический типы синапса
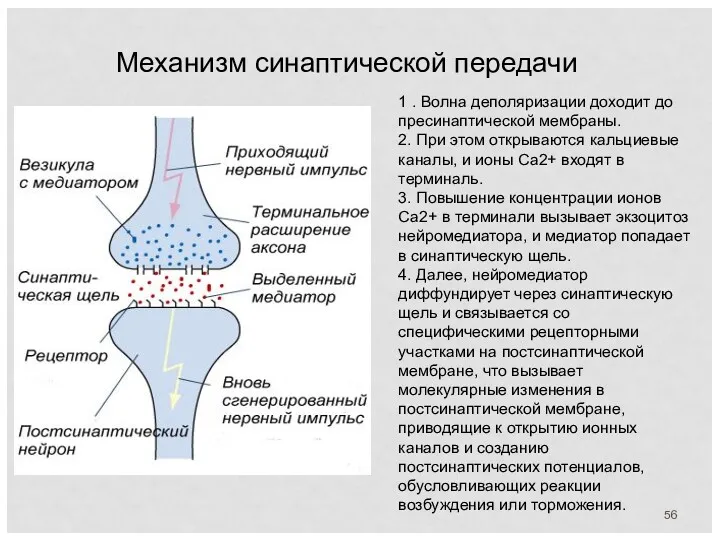
Механизм синаптической передачи
1 . Волна деполяризации доходит до пресинаптической мембраны.
2.
Механизм синаптической передачи
1 . Волна деполяризации доходит до пресинаптической мембраны.
2.

Нейрофибриллы образуют сеть в теле нейрона, а в отростках расположены параллельно.
Нейрофибриллы образуют сеть в теле нейрона, а в отростках расположены параллельно.

АКСОНАЛЬНЫЙ ТРАНСПОРТ
Нейротубулы — органеллы, ответственные за быстрый транспорт, который называется
АКСОНАЛЬНЫЙ ТРАНСПОРТ
Нейротубулы — органеллы, ответственные за быстрый транспорт, который называется

НЕЙРОМЕДИАТОРЫ
Отдельной разновидностью нейронов являются секреторные нейроны. Способность синтезировать и секретировать биологически
НЕЙРОМЕДИАТОРЫ
Отдельной разновидностью нейронов являются секреторные нейроны. Способность синтезировать и секретировать биологически
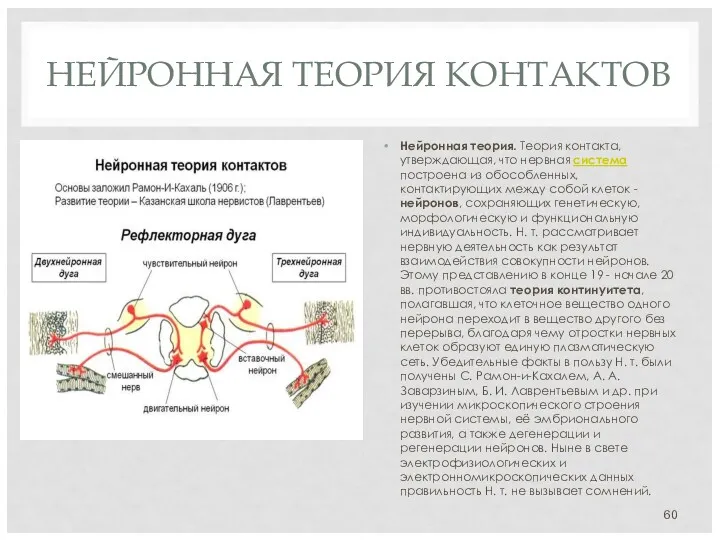
НЕЙРОННАЯ ТЕОРИЯ КОНТАКТОВ
Нейронная теория. Теория контакта, утверждающая, что нервная система построена
НЕЙРОННАЯ ТЕОРИЯ КОНТАКТОВ
Нейронная теория. Теория контакта, утверждающая, что нервная система построена


НЕЙРОНЫ
В зависимости от функции в рефлекторной дуге различают рецепторные
(чувствительные,
НЕЙРОНЫ
В зависимости от функции в рефлекторной дуге различают рецепторные
(чувствительные,
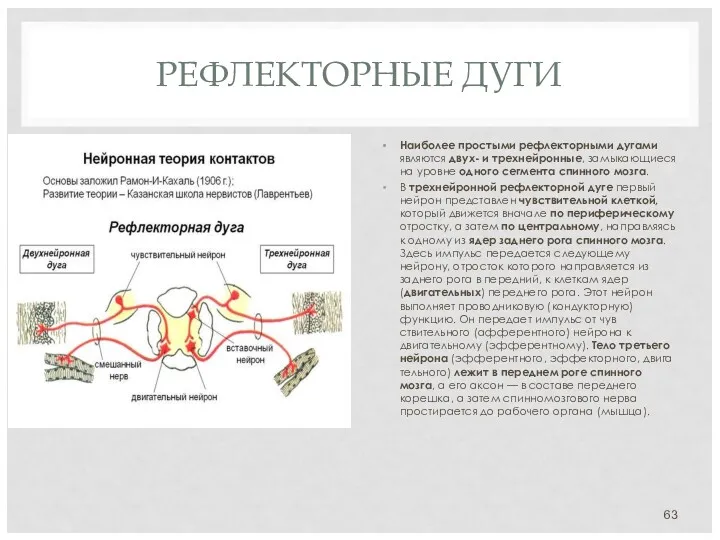
РЕФЛЕКТОРНЫЕ ДУГИ
Наиболее простыми рефлекторными дугами являются двух- и трехнейронные, замыкающиеся на
РЕФЛЕКТОРНЫЕ ДУГИ
Наиболее простыми рефлекторными дугами являются двух- и трехнейронные, замыкающиеся на

РЕФЛЕКТОРНАЯ ДУГА ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Рефлекторная дуга вегетативной нервной системы состоит из
РЕФЛЕКТОРНАЯ ДУГА ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Рефлекторная дуга вегетативной нервной системы состоит из
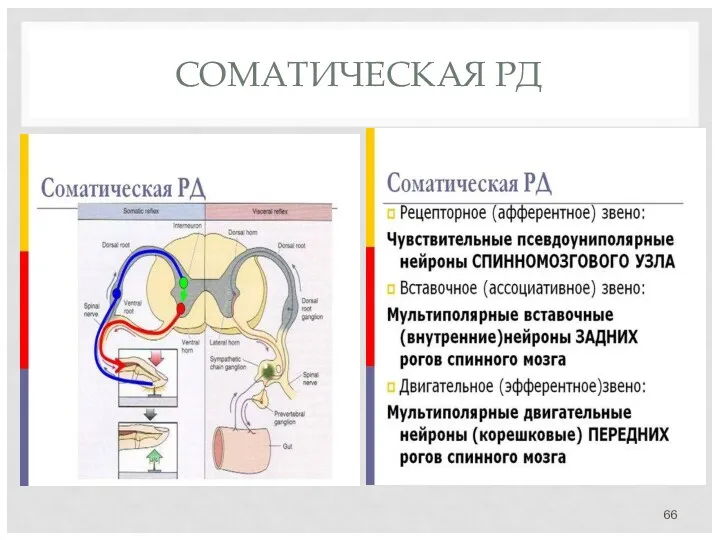
СОМАТИЧЕСКАЯ РД
СОМАТИЧЕСКАЯ РД

1-ый нейрон оканчивается в боковых рогах спинного мозга.
2-ой нейрон выходит за
1-ый нейрон оканчивается в боковых рогах спинного мозга.
2-ой нейрон выходит за

СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА СТРУКТУРНОЙ ОРГАНИЗАЦИИ РЕФЛЕКТОРНЫХ ДУГ СОМАТИЧЕСКОЙ И ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ.
Рефлекторная
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА СТРУКТУРНОЙ ОРГАНИЗАЦИИ РЕФЛЕКТОРНЫХ ДУГ СОМАТИЧЕСКОЙ И ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ.
Рефлекторная
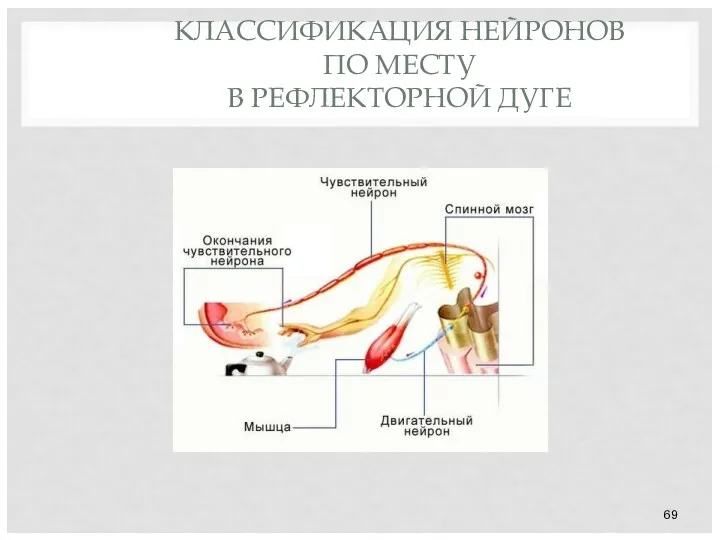
КЛАССИФИКАЦИЯ НЕЙРОНОВ
ПО МЕСТУ
В РЕФЛЕКТОРНОЙ ДУГЕ
КЛАССИФИКАЦИЯ НЕЙРОНОВ
ПО МЕСТУ
В РЕФЛЕКТОРНОЙ ДУГЕ

Гемато-энцефалический барьер включает в себя следующие компоненты:
• эндотелий кровеносных капилляров (с
Гемато-энцефалический барьер включает в себя следующие компоненты:
• эндотелий кровеносных капилляров (с

СПИННОЙ МОЗГ
Спинной мозг состоит из 2-х половин, соединенных узкой полоской, содержащей
СПИННОЙ МОЗГ
Спинной мозг состоит из 2-х половин, соединенных узкой полоской, содержащей

РАЗВИТИЕ: В процессе развития спинного мозга из нервной трубки нейроны группируются
РАЗВИТИЕ: В процессе развития спинного мозга из нервной трубки нейроны группируются




Среди нейронов спинного мозга можно выделить три вида клеток:
*корешковые,
*внутренние,
*пучковые.
Среди нейронов спинного мозга можно выделить три вида клеток:
*корешковые,
*внутренние,
*пучковые.

В задних рогах различают:
* губчатый слой,
*желатинозное вещество,
*собственное ядро заднего
В задних рогах различают:
* губчатый слой,
*желатинозное вещество,
*собственное ядро заднего

Глия спинного мозга
Основную часть глиального остова серого вещества составляют протоплазматические и
Глия спинного мозга
Основную часть глиального остова серого вещества составляют протоплазматические и

СЕРОЕ ВЕЩЕСТВО
В сером веществе имеются выступы (столбы) получили название рога -
СЕРОЕ ВЕЩЕСТВО
В сером веществе имеются выступы (столбы) получили название рога -


СЕРОЕ ВЕЩЕСТВО
К мягкой мозговой оболочке примыкает паутинная оболочка (arachnoidea), состоящая из
СЕРОЕ ВЕЩЕСТВО
К мягкой мозговой оболочке примыкает паутинная оболочка (arachnoidea), состоящая из

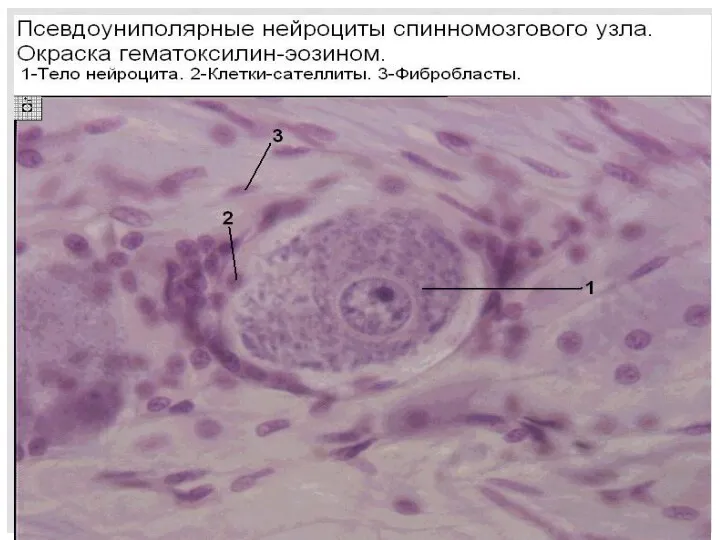
СЕРОЕ ВЕЩЕСТВО
Чувствительные клетки располагаются в спинальных ганглиях. Нейриты нервных клеток спинального
СЕРОЕ ВЕЩЕСТВО
Чувствительные клетки располагаются в спинальных ганглиях. Нейриты нервных клеток спинального
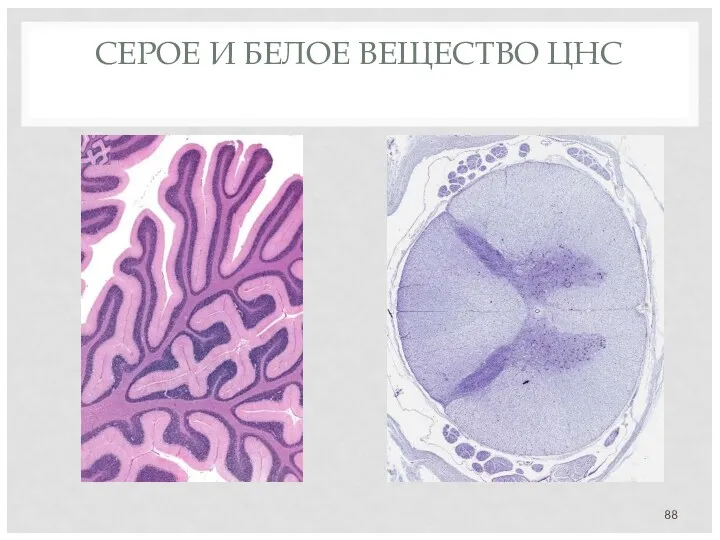
СЕРОЕ И БЕЛОЕ ВЕЩЕСТВО ЦНС
СЕРОЕ И БЕЛОЕ ВЕЩЕСТВО ЦНС
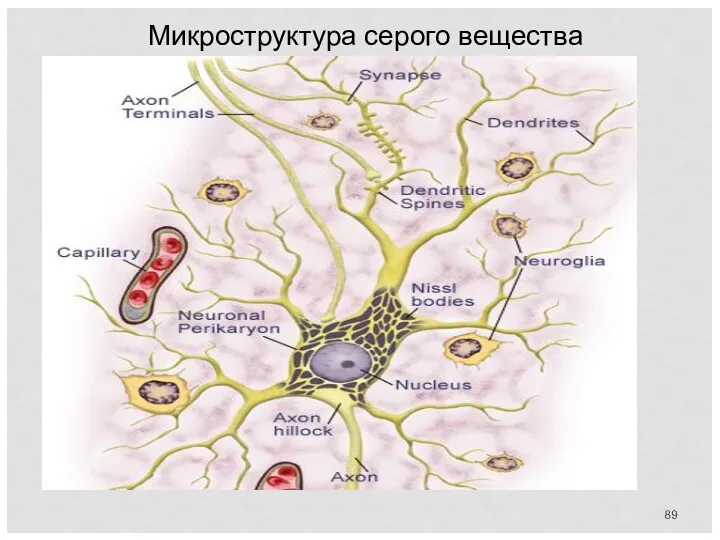
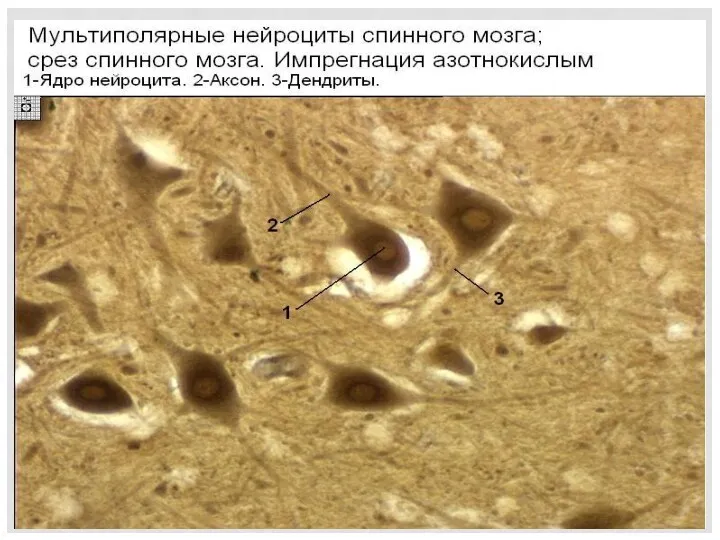
Микроструктура серого вещества
Микроструктура серого вещества
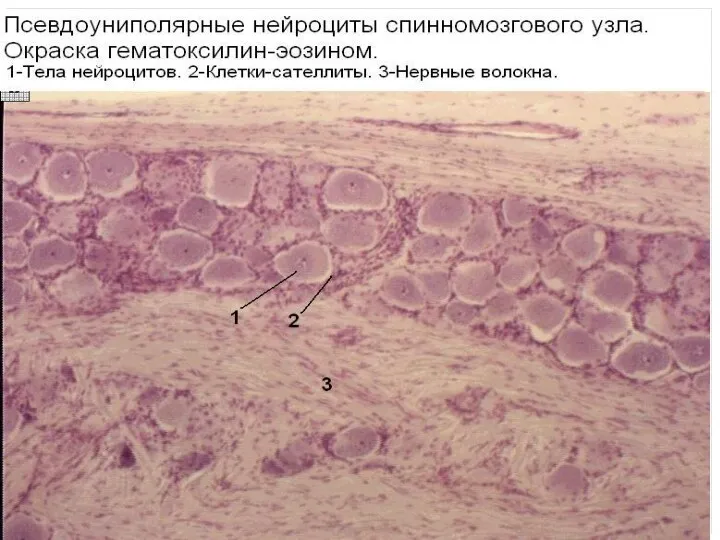

РЕГЕНЕРАЦИЯ ЗАВИСИТ ОТ МЕСТА ТРАВМЫ
Как в центральной, так и в периферической
РЕГЕНЕРАЦИЯ ЗАВИСИТ ОТ МЕСТА ТРАВМЫ
Как в центральной, так и в периферической

РЕАКЦИЯ НЕЙРОНОВ И ИХ ВОЛОКОН НА ТРАВМУ
Перерезка нервного волокна вызывает различные
РЕАКЦИЯ НЕЙРОНОВ И ИХ ВОЛОКОН НА ТРАВМУ
Перерезка нервного волокна вызывает различные

ПОВРЕЖДЕННЫЕ НЕРВНЫЕ ВОЛОКНА ГОЛОВНОГО И СПИННОГО МОЗГА
Поврежденные нервные волокна головного и
ПОВРЕЖДЕННЫЕ НЕРВНЫЕ ВОЛОКНА ГОЛОВНОГО И СПИННОГО МОЗГА
Поврежденные нервные волокна головного и
 Формирование УУД на уроках биологии и химии.
Формирование УУД на уроках биологии и химии. Презентация к уроку биологии в 6 классе по темеЦветок.Соцветия
Презентация к уроку биологии в 6 классе по темеЦветок.Соцветия Введение в паразитологию
Введение в паразитологию Клеточная оболочка и цитоплазма
Клеточная оболочка и цитоплазма Класс двудольные. Семейство Крестоцветные
Класс двудольные. Семейство Крестоцветные Презентация по теме : Подземное видоизменение побегов
Презентация по теме : Подземное видоизменение побегов Влияние макро- и микроэлементов на здоровье человека и животных
Влияние макро- и микроэлементов на здоровье человека и животных Чому мігрують тварини
Чому мігрують тварини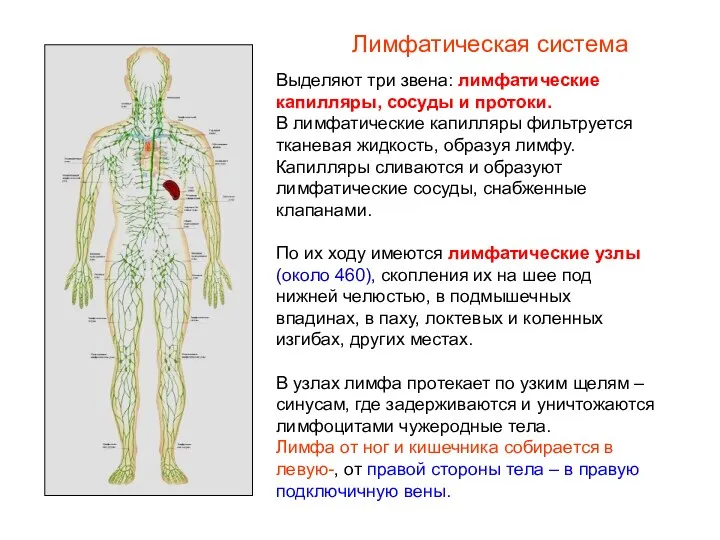 Лимфатическая система в организме человека
Лимфатическая система в организме человека Жануар және өсімдік майлары
Жануар және өсімдік майлары Радиацияның биологиялық әсері
Радиацияның биологиялық әсері Урок Развитие эволюционного учения. Ч.Дарвин.
Урок Развитие эволюционного учения. Ч.Дарвин. Болезни и травмы органов дыхания
Болезни и травмы органов дыхания Хищные звери
Хищные звери Половое размножение
Половое размножение Торможение условных рефлексов. Лекция № 29
Торможение условных рефлексов. Лекция № 29 Оплодотворенное и неоплодотворенное яйцо
Оплодотворенное и неоплодотворенное яйцо Высшая нервная деятельность
Высшая нервная деятельность СРС на тему: “Везикулярный транспорт”
СРС на тему: “Везикулярный транспорт” Мир насекомых
Мир насекомых Черепные нервы: топографическая и клиническая анатомия
Черепные нервы: топографическая и клиническая анатомия Особенности строения и функционирования гладких мышц
Особенности строения и функционирования гладких мышц Такие разные цветы
Такие разные цветы Лекция 1. Растениеводство
Лекция 1. Растениеводство Різноманітний світ тварин
Різноманітний світ тварин Буферные системы. Классификация буферных растворов
Буферные системы. Классификация буферных растворов Эмбриогенез человека. Оплодотворение, дробление. Первая фаза гаструляции. Часть 1
Эмбриогенез человека. Оплодотворение, дробление. Первая фаза гаструляции. Часть 1 Plant Diversity I: How Plants Colonized Land
Plant Diversity I: How Plants Colonized Land