- Нейромедиаторы. Классификация медиаторов

Содержание
- 2. Высокомолекулрные, долговременного действия: НЕЙРОПЕПТИДЫ Гипоталамические Гипофизарные Гастро-интестинальные и другие Низкомолекулярные, кратковременного действия: АЦЕТИЛХОЛИН АМИНЫ АМИНОКИСЛОТЫ ОКСИД
- 3. МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ ПОСТСИНАПТИЧЕСКИХ МЕМБРАН ИОНОТРОПНЫЕ (связанные с хемочувствительными ионными каналами) Р Р ИОННЫЙ КАНАЛ ЗАКРЫТ ИОННЫЙ
- 4. МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ ПОСТСИНАПТИЧЕСКИХ МЕМБРАН МЕТАБОТРОПНЫЕ (связанные с системой вторых посредников) Р АДЕНИЛАТ- ЦИКЛАЗА G-белок ц АМФ
- 5. АКТИВАЦИЯ СИСТЕМЫ ВТОРЫХ ПОСРЕДНИКОВ Медиатор (первый посредник) химически связывается с рецептором постсинаптической мембраны. Происходит активация G-белка.
- 6. СИСТЕМА ВТОРЫХ ПОСРЕДНИКОВ – каскад биохимических реакций, который работает как высоко-эффективный усилитель медиатор рецептор G-белок Аденилат-
- 7. НЕРВНЫЕ ЦЕНТРЫ – скопления нейронов, которые участвуют в регуляции одной и той же функции организма.
- 8. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ ЗАВИСЯТ: От свойств нервных клеток. От свойств химических синапсов. От сочетания возбуждающих и
- 9. ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО РЕФЛЕКТОРНОЙ ДУГЕ (по сравнению с нервным волокном) Одностороннее проведение: за счёт 1-стороннего
- 10. СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ Суммация возбуждения (временная и пространственная) Инерционность Последействие Пластичность Фоновая активность, тонус Высокая утомляемость
- 11. ЭЛЕМЕНТАРНЫЕ НЕЙРОННЫЕ КОНТУРЫ ДИВЕРГЕНЦИЯ Дивергенция приводит к распространению (иррадиации) возбуждения по всем отделам ЦНС
- 12. КОНВЕРГЕНЦИЯ Конвергенция лежит в основе простран- ственной суммации возбуждения, ведёт к трансформации ритма возбуждения
- 13. КОЛЬЦЕВЫЕ НЕЙРОННЫЕ ЦЕПИ Благодаря циркуляции возбуждения по замкнутым нейронным цепям, происходит усиление импульсации. Возбуждение нейронов продолжается
- 14. ВОЗВРАТНОЕ ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ В замкнутой цепи нейронов с возвратным постсинаптическим торможением не только снижается возбудимость нейрона
- 15. Суммация ВПСП и ТПСП при возвратном постсинаптическом торможении нейрона ВПСП ТПСП Результат суммации Импульсация в нервном
- 16. ВОЗВРАТНОЕ ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ С помощью возвратного пресинаптического торможения происходит ограничение потока импульсов, поступающих к нервному центру
- 17. РЕЦИПРОКНОЕ ТОРМОЖЕНИЕ К сгибательным мышцам К разгибательным мышцам Нет импульсов Благодаря реципрокному (сопряжённому) торможению, при возбуждении
- 20. КООРДИНАЦИЯ ДЕЯТЕЛЬНОСТИ ЦНС I. ПРИНЦИП СУБОРДИНАЦИИ СПИННОЙ МОЗГ СТВОЛ ГОЛОВНОГО МОЗГА КОРА БОЛЬШИХ ПОЛУШАРИЙ
- 21. II. ПРИНЦИП ОБРАТНОЙ СВЯЗИ КОРА БОЛЬШИХ ПОЛУШАРИЙ СТВОЛ ГОЛОВНОГО МОЗГА СПИННОЙ МОЗГ ВТОРИЧНАЯ АФФЕРЕНТАЦИЯ КОПИЯ ЭФФЕРЕНТАЦИИ
- 22. III. ПРИНЦИП ОБЛЕГЧЕНИЯ И ОККЛЮЗИИ (СИНЕРГИЧЕСКИХ НЕРВНЫХ ЦЕНТРОВ) ОБЛЕГЧЕНИЕ Афферентный вход А (допороговый) Афферентный вход Б
- 23. ОККЛЮЗИЯ Афферентный вход А (сверхпорогоый) Афферентный вход Б (сверхпороговый) Суммарный ответ меньше, чем простая сумма двух
- 24. IV. ПРИНЦИП РЕЦИПРОКНОЙ ИННЕРВАЦИИ СПИННОЙ МОЗГ вход возбуждающий К сгибательным мышцам Отсутствие импульсации к разгибательным мышцам
- 25. V. ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ (ОКП) ФАКТОР СИЛЫ В БОРЬБЕ ЗА ОКП Конвергенция импульсов от разных
- 26. Доминантный центр «притягивает» к себе возбуждение, возникшее в других центрах. Из всех возможных ответных реакций в
- 27. МЕХАНИЗМ ФОРМИРОВАНИЯ ДОМИНАНТЫ (СХЕМА ФУНКЦИОНАЛЬНОЙ СИСТЕМЫ) П.К. АНОХИН ОБСТАНОВОЧ- НАЯ АФФЕРЕНТАЦИЯ ВНУТРЕННЯЯ МОТИВАЦИЯ ПУСКОВАЯ АФФЕРЕНТАЦИЯ ПАМЯТЬ
- 28. КОНЕЦ
- 30. Скачать презентацию

Высокомолекулрные, долговременного действия:
НЕЙРОПЕПТИДЫ
Гипоталамические
Гипофизарные
Гастро-интестинальные
и другие
Низкомолекулярные, кратковременного действия:
АЦЕТИЛХОЛИН
АМИНЫ
АМИНОКИСЛОТЫ
ОКСИД АЗОТА (NO)
КЛАССИФИКАЦИЯ МЕДИАТОРОВ
Высокомолекулрные, долговременного действия:
НЕЙРОПЕПТИДЫ
Гипоталамические
Гипофизарные
Гастро-интестинальные
и другие
Низкомолекулярные, кратковременного действия:
АЦЕТИЛХОЛИН
АМИНЫ
АМИНОКИСЛОТЫ
ОКСИД АЗОТА (NO)
КЛАССИФИКАЦИЯ МЕДИАТОРОВ
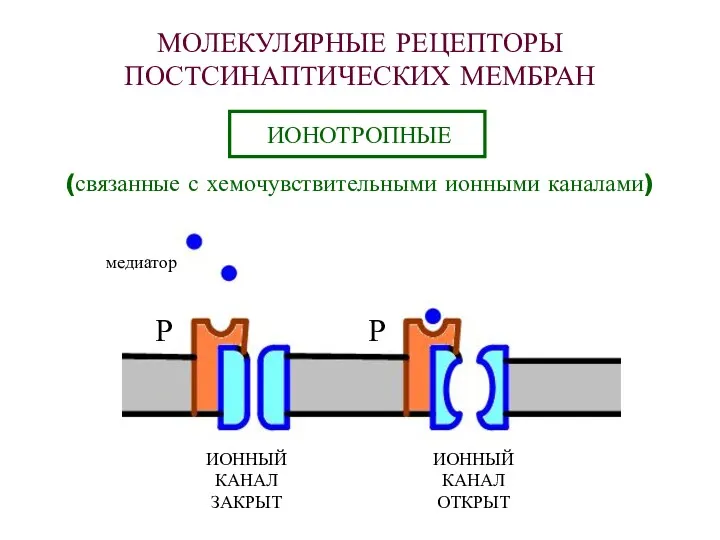
МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ
ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
ИОНОТРОПНЫЕ
(связанные с хемочувствительными ионными каналами)
Р
Р
ИОННЫЙ
КАНАЛ
ЗАКРЫТ
ИОННЫЙ
КАНАЛ
ОТКРЫТ
медиатор
МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ
ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
ИОНОТРОПНЫЕ
(связанные с хемочувствительными ионными каналами)
Р
Р
ИОННЫЙ
КАНАЛ
ЗАКРЫТ
ИОННЫЙ
КАНАЛ
ОТКРЫТ
медиатор
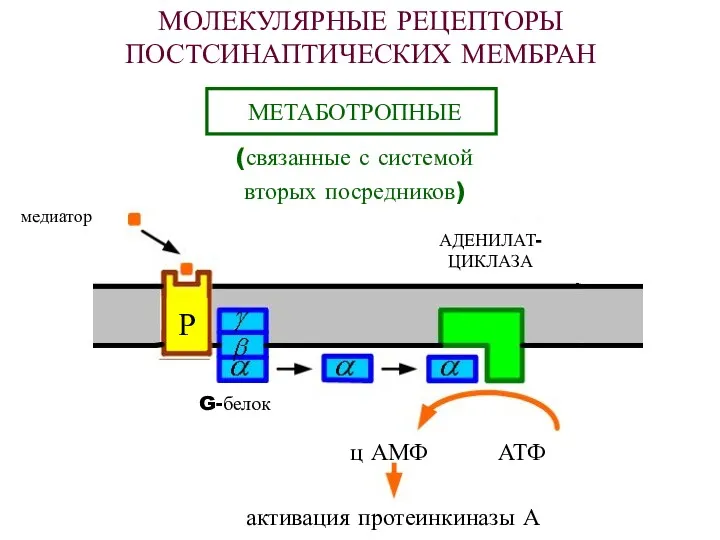
МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ
ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
МЕТАБОТРОПНЫЕ
(связанные с системой
вторых посредников)
Р
АДЕНИЛАТ-
ЦИКЛАЗА
G-белок
ц АМФ АТФ
активация протеинкиназы А
медиатор
МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ
ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
МЕТАБОТРОПНЫЕ
(связанные с системой
вторых посредников)
Р
АДЕНИЛАТ-
ЦИКЛАЗА
G-белок
ц АМФ АТФ
активация протеинкиназы А
медиатор

АКТИВАЦИЯ
СИСТЕМЫ ВТОРЫХ ПОСРЕДНИКОВ
Медиатор (первый посредник) химически связывается с рецептором постсинаптической
АКТИВАЦИЯ
СИСТЕМЫ ВТОРЫХ ПОСРЕДНИКОВ
Медиатор (первый посредник) химически связывается с рецептором постсинаптической
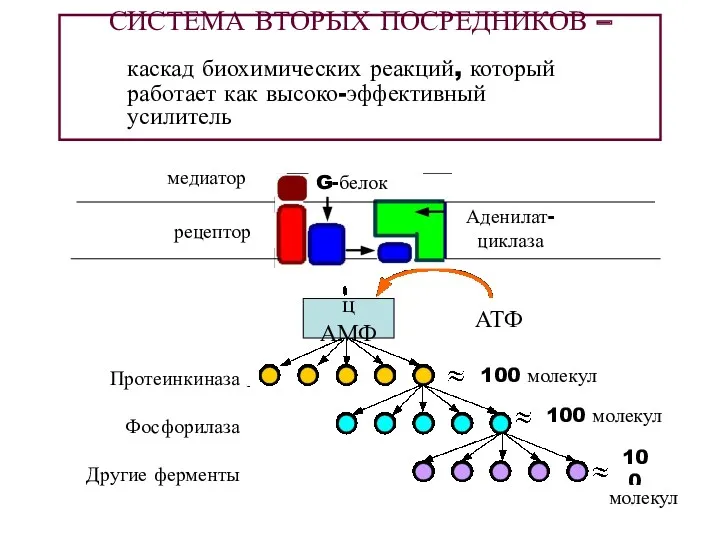
СИСТЕМА ВТОРЫХ ПОСРЕДНИКОВ –
каскад биохимических реакций, который работает как высоко-эффективный
СИСТЕМА ВТОРЫХ ПОСРЕДНИКОВ –
каскад биохимических реакций, который работает как высоко-эффективный

НЕРВНЫЕ ЦЕНТРЫ –
скопления нейронов, которые участвуют в регуляции одной и
НЕРВНЫЕ ЦЕНТРЫ –
скопления нейронов, которые участвуют в регуляции одной и

СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
ЗАВИСЯТ:
От свойств нервных клеток.
От свойств химических синапсов.
От сочетания возбуждающих
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
ЗАВИСЯТ:
От свойств нервных клеток.
От свойств химических синапсов.
От сочетания возбуждающих

ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО РЕФЛЕКТОРНОЙ ДУГЕ
(по сравнению с нервным волокном)
Одностороннее проведение:
за
ОСОБЕННОСТИ ПРОВЕДЕНИЯ ВОЗБУЖДЕНИЯ ПО РЕФЛЕКТОРНОЙ ДУГЕ
(по сравнению с нервным волокном)
Одностороннее проведение:
за

СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Суммация возбуждения (временная и пространственная)
Инерционность
Последействие
Пластичность
Фоновая активность, тонус
Высокая утомляемость
Высокая
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Суммация возбуждения (временная и пространственная)
Инерционность
Последействие
Пластичность
Фоновая активность, тонус
Высокая утомляемость
Высокая
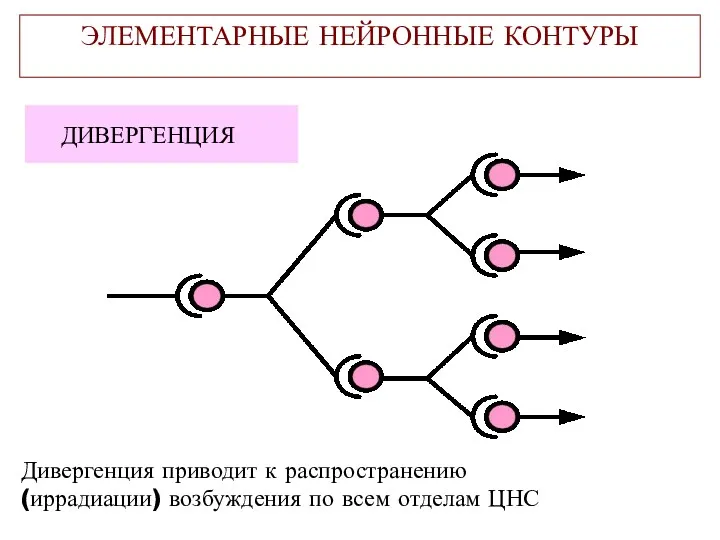
ЭЛЕМЕНТАРНЫЕ НЕЙРОННЫЕ КОНТУРЫ
ДИВЕРГЕНЦИЯ
Дивергенция приводит к распространению
(иррадиации) возбуждения по всем отделам ЦНС
ЭЛЕМЕНТАРНЫЕ НЕЙРОННЫЕ КОНТУРЫ
ДИВЕРГЕНЦИЯ
Дивергенция приводит к распространению
(иррадиации) возбуждения по всем отделам ЦНС
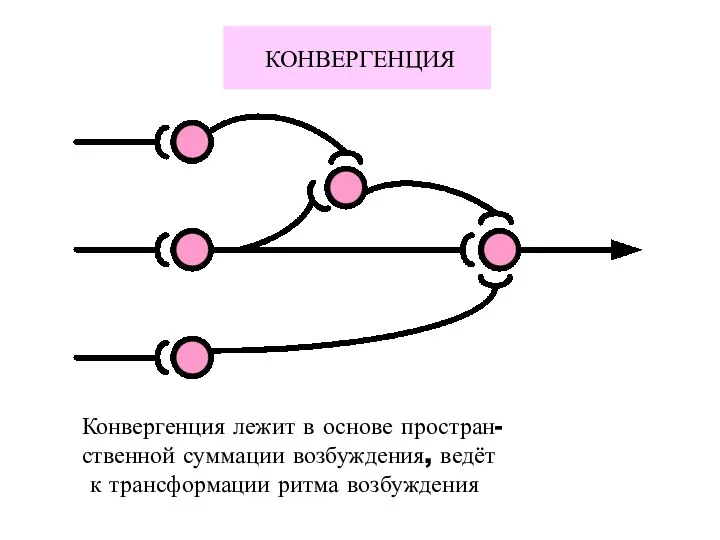
КОНВЕРГЕНЦИЯ
Конвергенция лежит в основе простран-
ственной суммации возбуждения, ведёт
к трансформации ритма
КОНВЕРГЕНЦИЯ
Конвергенция лежит в основе простран-
ственной суммации возбуждения, ведёт
к трансформации ритма
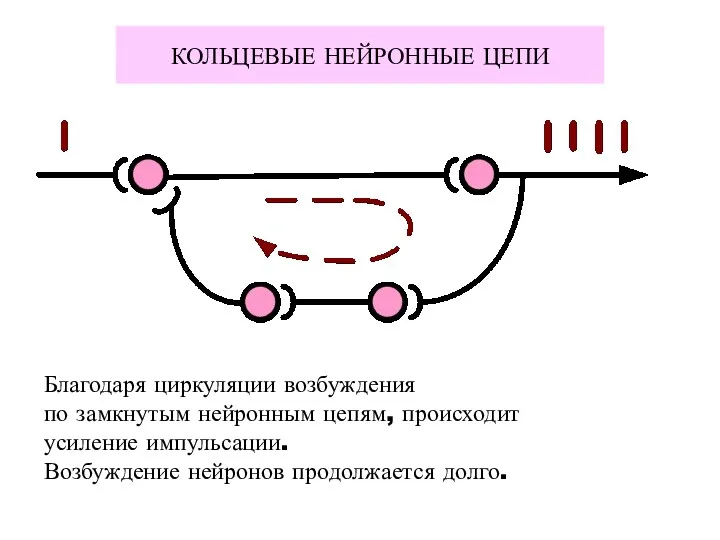
КОЛЬЦЕВЫЕ НЕЙРОННЫЕ ЦЕПИ
Благодаря циркуляции возбуждения
по замкнутым нейронным цепям, происходит
усиление импульсации.
Возбуждение
КОЛЬЦЕВЫЕ НЕЙРОННЫЕ ЦЕПИ
Благодаря циркуляции возбуждения
по замкнутым нейронным цепям, происходит
усиление импульсации.
Возбуждение
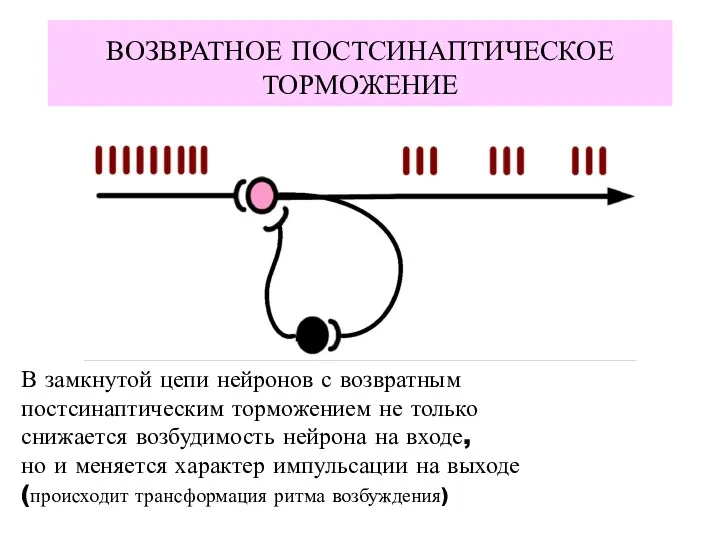
ВОЗВРАТНОЕ ПОСТСИНАПТИЧЕСКОЕ
ТОРМОЖЕНИЕ
В замкнутой цепи нейронов с возвратным
постсинаптическим торможением не только
снижается
ВОЗВРАТНОЕ ПОСТСИНАПТИЧЕСКОЕ
ТОРМОЖЕНИЕ
В замкнутой цепи нейронов с возвратным
постсинаптическим торможением не только
снижается
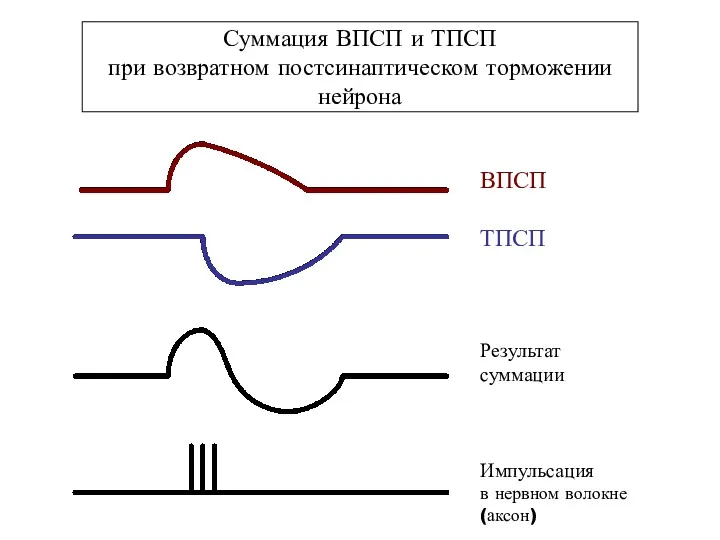
Суммация ВПСП и ТПСП
при возвратном постсинаптическом торможении нейрона
ВПСП
ТПСП
Результат
суммации
Импульсация
в нервном волокне
(аксон)
Суммация ВПСП и ТПСП
при возвратном постсинаптическом торможении нейрона
ВПСП
ТПСП
Результат
суммации
Импульсация
в нервном волокне
(аксон)
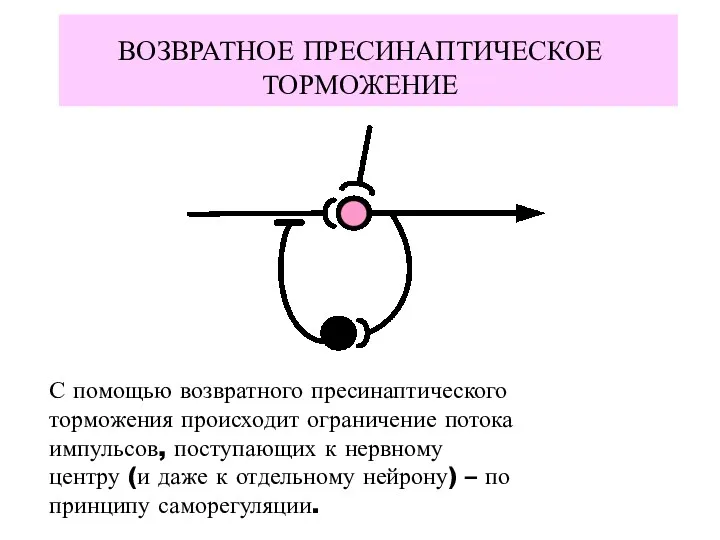
ВОЗВРАТНОЕ ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
С помощью возвратного пресинаптического
торможения происходит ограничение потока
импульсов, поступающих к
ВОЗВРАТНОЕ ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
С помощью возвратного пресинаптического
торможения происходит ограничение потока
импульсов, поступающих к
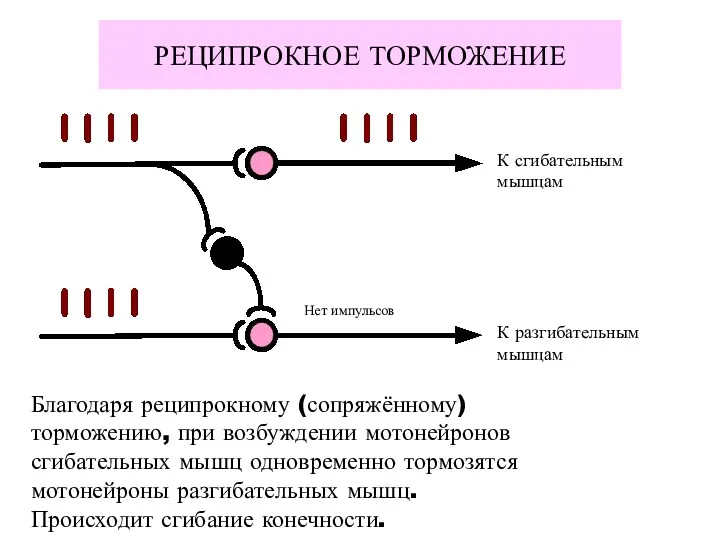
РЕЦИПРОКНОЕ ТОРМОЖЕНИЕ
К сгибательным
мышцам
К разгибательным
мышцам
Нет импульсов
Благодаря реципрокному (сопряжённому)
торможению, при возбуждении мотонейронов
сгибательных
РЕЦИПРОКНОЕ ТОРМОЖЕНИЕ
К сгибательным
мышцам
К разгибательным
мышцам
Нет импульсов
Благодаря реципрокному (сопряжённому)
торможению, при возбуждении мотонейронов
сгибательных
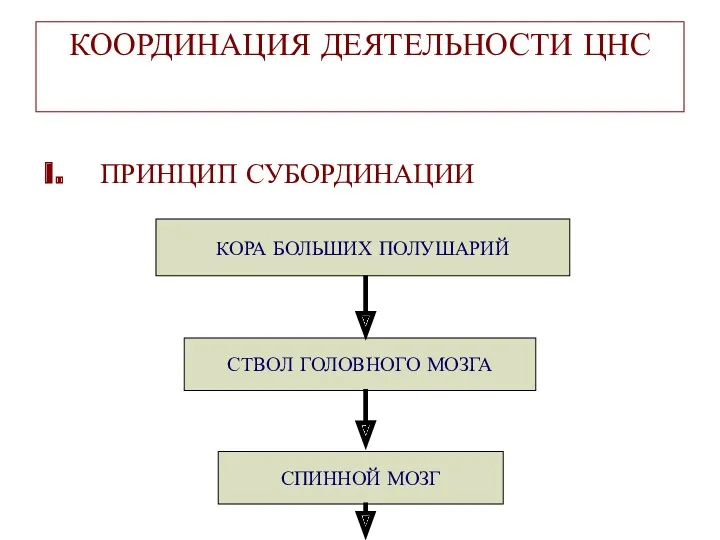
КООРДИНАЦИЯ ДЕЯТЕЛЬНОСТИ ЦНС
I. ПРИНЦИП СУБОРДИНАЦИИ
СПИННОЙ МОЗГ
СТВОЛ ГОЛОВНОГО МОЗГА
КОРА БОЛЬШИХ ПОЛУШАРИЙ
КООРДИНАЦИЯ ДЕЯТЕЛЬНОСТИ ЦНС
I. ПРИНЦИП СУБОРДИНАЦИИ
СПИННОЙ МОЗГ
СТВОЛ ГОЛОВНОГО МОЗГА
КОРА БОЛЬШИХ ПОЛУШАРИЙ
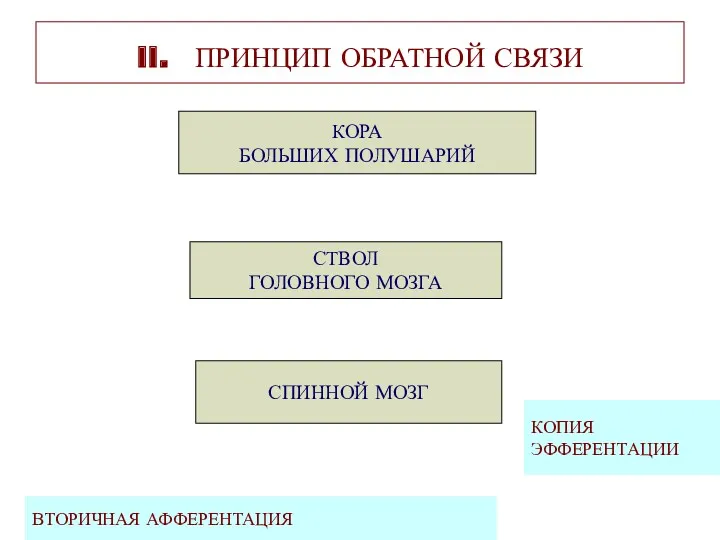
II. ПРИНЦИП ОБРАТНОЙ СВЯЗИ
КОРА
БОЛЬШИХ ПОЛУШАРИЙ
СТВОЛ
ГОЛОВНОГО МОЗГА
СПИННОЙ МОЗГ
ВТОРИЧНАЯ АФФЕРЕНТАЦИЯ
КОПИЯ
ЭФФЕРЕНТАЦИИ
II. ПРИНЦИП ОБРАТНОЙ СВЯЗИ
КОРА
БОЛЬШИХ ПОЛУШАРИЙ
СТВОЛ
ГОЛОВНОГО МОЗГА
СПИННОЙ МОЗГ
ВТОРИЧНАЯ АФФЕРЕНТАЦИЯ
КОПИЯ
ЭФФЕРЕНТАЦИИ
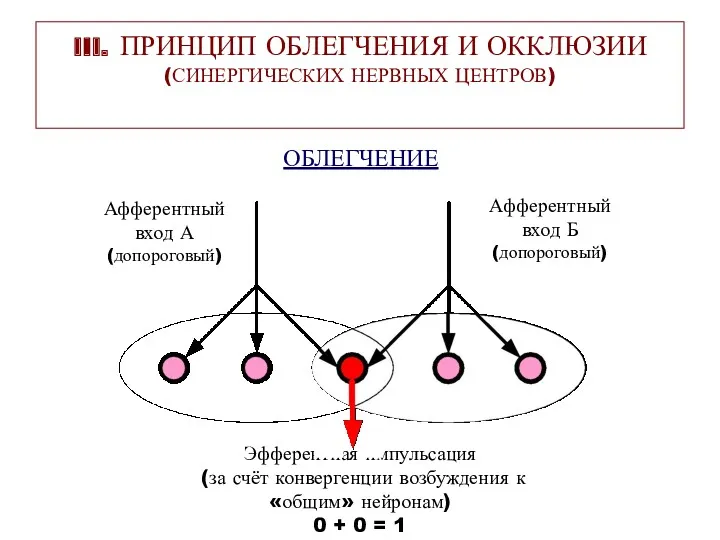
III. ПРИНЦИП ОБЛЕГЧЕНИЯ И ОККЛЮЗИИ (СИНЕРГИЧЕСКИХ НЕРВНЫХ ЦЕНТРОВ)
ОБЛЕГЧЕНИЕ
Афферентный
вход А
(допороговый)
Афферентный
вход Б
(допороговый)
Эфферентная импульсация
(за
III. ПРИНЦИП ОБЛЕГЧЕНИЯ И ОККЛЮЗИИ (СИНЕРГИЧЕСКИХ НЕРВНЫХ ЦЕНТРОВ)
ОБЛЕГЧЕНИЕ
Афферентный
вход А
(допороговый)
Афферентный
вход Б
(допороговый)
Эфферентная импульсация
(за
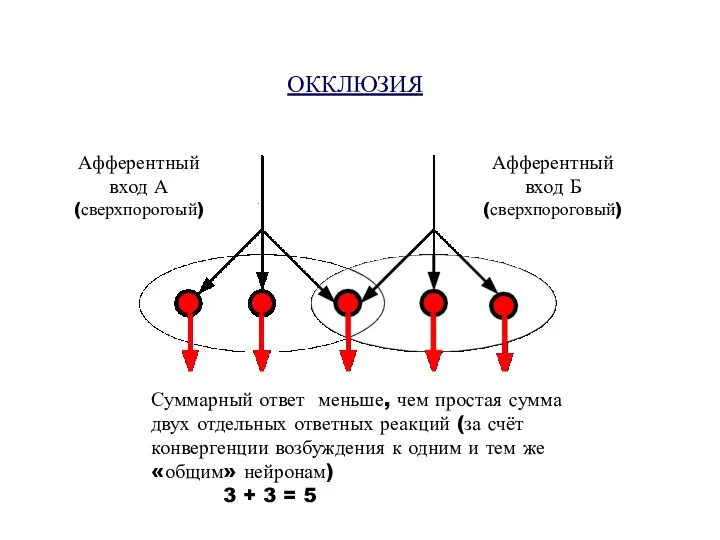
ОККЛЮЗИЯ
Афферентный
вход А
(сверхпорогоый)
Афферентный
вход Б
(сверхпороговый)
Суммарный ответ меньше, чем простая сумма двух отдельных ответных
ОККЛЮЗИЯ
Афферентный
вход А
(сверхпорогоый)
Афферентный
вход Б
(сверхпороговый)
Суммарный ответ меньше, чем простая сумма двух отдельных ответных
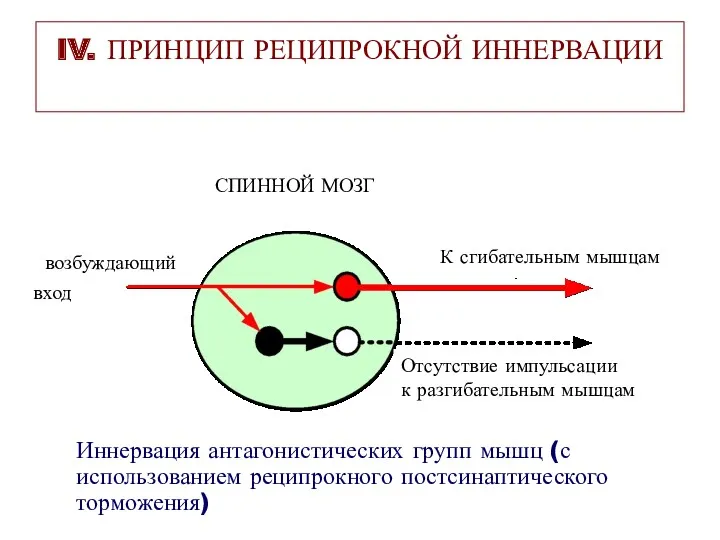
IV. ПРИНЦИП РЕЦИПРОКНОЙ ИННЕРВАЦИИ
СПИННОЙ МОЗГ
вход
возбуждающий
К сгибательным мышцам
Отсутствие импульсации
к разгибательным мышцам
Иннервация
IV. ПРИНЦИП РЕЦИПРОКНОЙ ИННЕРВАЦИИ
СПИННОЙ МОЗГ
вход
возбуждающий
К сгибательным мышцам
Отсутствие импульсации
к разгибательным мышцам
Иннервация
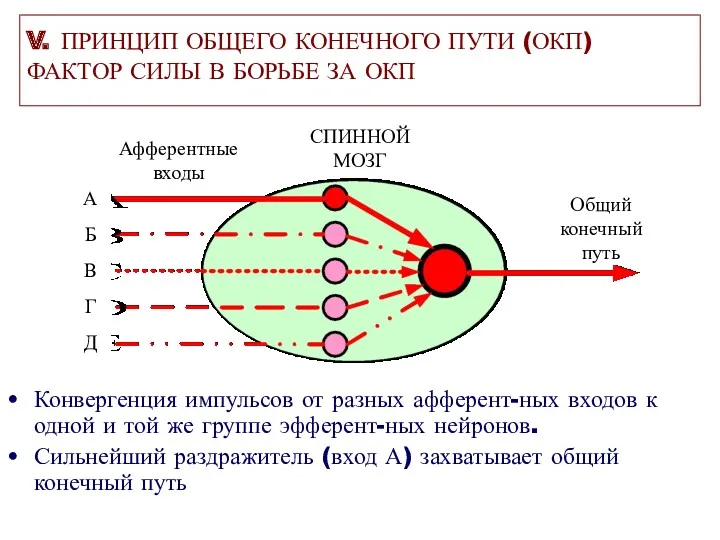
V. ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ (ОКП)
ФАКТОР СИЛЫ В БОРЬБЕ ЗА ОКП
Конвергенция
V. ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ (ОКП)
ФАКТОР СИЛЫ В БОРЬБЕ ЗА ОКП
Конвергенция
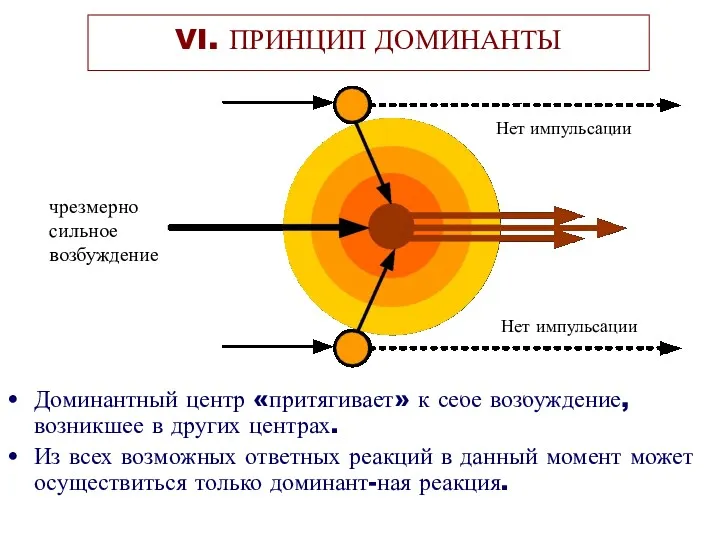
Доминантный центр «притягивает» к себе возбуждение, возникшее в других центрах.
Из всех
Доминантный центр «притягивает» к себе возбуждение, возникшее в других центрах.
Из всех
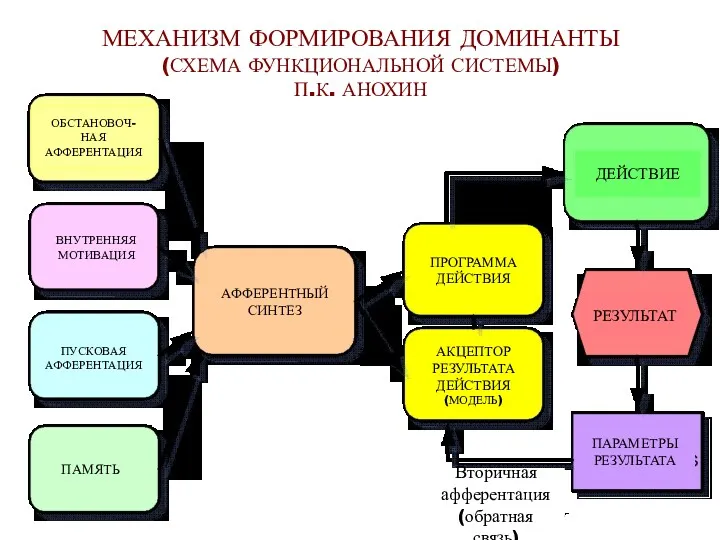
МЕХАНИЗМ ФОРМИРОВАНИЯ ДОМИНАНТЫ
(СХЕМА ФУНКЦИОНАЛЬНОЙ СИСТЕМЫ)
П.К. АНОХИН
ОБСТАНОВОЧ-
НАЯ
АФФЕРЕНТАЦИЯ
ВНУТРЕННЯЯ
МОТИВАЦИЯ
ПУСКОВАЯ
АФФЕРЕНТАЦИЯ
ПАМЯТЬ
АФФЕРЕНТНЫЙ
СИНТЕЗ
ПРОГРАММА
ДЕЙСТВИЯ
АКЦЕПТОР
РЕЗУЛЬТАТА
ДЕЙСТВИЯ
(МОДЕЛЬ)
ДЕЙСТВИЕ
РЕЗУЛЬТАТ
ПАРАМЕТРЫ
РЕЗУЛЬТАТА
Вторичная
афферентация
(обратная связь)
МЕХАНИЗМ ФОРМИРОВАНИЯ ДОМИНАНТЫ
(СХЕМА ФУНКЦИОНАЛЬНОЙ СИСТЕМЫ)
П.К. АНОХИН
ОБСТАНОВОЧ-
НАЯ
АФФЕРЕНТАЦИЯ
ВНУТРЕННЯЯ
МОТИВАЦИЯ
ПУСКОВАЯ
АФФЕРЕНТАЦИЯ
ПАМЯТЬ
АФФЕРЕНТНЫЙ
СИНТЕЗ
ПРОГРАММА
ДЕЙСТВИЯ
АКЦЕПТОР
РЕЗУЛЬТАТА
ДЕЙСТВИЯ
(МОДЕЛЬ)
ДЕЙСТВИЕ
РЕЗУЛЬТАТ
ПАРАМЕТРЫ
РЕЗУЛЬТАТА
Вторичная
афферентация
(обратная связь)

КОНЕЦ
КОНЕЦ
 Размножение организмов
Размножение организмов Видоизмененные корни растений
Видоизмененные корни растений Водоросли -их значение в жизни человека
Водоросли -их значение в жизни человека Углеводы. Функции углеводов
Углеводы. Функции углеводов Исчезающие растения и животные России
Исчезающие растения и животные России презентация к уроку в 9 классе ФОТОСИНТЕЗ
презентация к уроку в 9 классе ФОТОСИНТЕЗ Птахи, які розмножуються взимку
Птахи, які розмножуються взимку Физиология дыхания
Физиология дыхания Луговые травы
Луговые травы Эволюционное учение Ч. Дарвина
Эволюционное учение Ч. Дарвина Эмбриогенез печени и желчевыводящих путей
Эмбриогенез печени и желчевыводящих путей Синквейн Жизнь
Синквейн Жизнь Витамин C
Витамин C Биология. Метаболизм
Биология. Метаболизм Презентация по теме Доказательства эволюции органического мира
Презентация по теме Доказательства эволюции органического мира охорона первоцвітів. Збереження ранніх квітучих рослин
охорона первоцвітів. Збереження ранніх квітучих рослин Строение тела человека (2 класс)
Строение тела человека (2 класс) Отряд Китообразные
Отряд Китообразные Eukaryotic microorganisms. Fungi
Eukaryotic microorganisms. Fungi Гортань. Функции гортани
Гортань. Функции гортани Буферные системы. Классификация буферных растворов
Буферные системы. Классификация буферных растворов Презентация к уроку биологии в 8 классе по теме Скелет человека. Осевой скелет по программе Д.В. Колесова,Р.Д. Маша,И. Н. Беляева.
Презентация к уроку биологии в 8 классе по теме Скелет человека. Осевой скелет по программе Д.В. Колесова,Р.Д. Маша,И. Н. Беляева. Классификация животных
Классификация животных Обмен веществ и энергии в клетке
Обмен веществ и энергии в клетке Систематика растений. Урок биологии в 7 классе
Систематика растений. Урок биологии в 7 классе Сосуды малого и большого кругов кровообращения
Сосуды малого и большого кругов кровообращения Цветковые растения
Цветковые растения Строение и работа сердца
Строение и работа сердца