- О способности к эволюции

Содержание
- 2. Evolvability (способность к эволюции) – способность популяции создавать генетическое разнообразие, и, как следствие, новые более адаптивные
- 3. Ландшафты приспособленности Ландшафт приспособленности – математическая модель, позволяющая анализировать относительную приспособленность различных генов. Спускаться вниз по
- 4. Как найти улучшенный фенотип, при этом не потеряв предыдущего? Иметь большую выборку различных связанных нейтральных генотипов
- 5. Робустность Робустность – способность биологической системы поддерживать стабильность определенного фенотипа при действии различных факторов.
- 6. Фенотипические переходы, которые в норме блокирует робустность Чем больше робустность, тем больше соседей у данного генотипа
- 7. Шапероны – один из механизмов мутационной робустности Шапероны способствуют мутационной робустности пытаясь «маскировать» мутантные пептиды, способствуя
- 8. Всегда ли робустность уменьшает фенотипическую вариацию? Робустность расширяет пространство возможных генотипов, т.к. способствует образованию нейтральных генотипов.
- 9. Нейтральные генотипы более разнообразны, т.к. у находящихся выше на ландшафте «полезных» генотипов меньше возможностей для эволюции.
- 10. Скрытая вариация (cryptic variation) Адаптация этих популяций к изменению РНК-субстрата.
- 11. Способы ускорения поиска нужного генотипа среди скрытых В стрессовой ситуации (например, изменение среды) преимущество получают организмы,
- 12. Сначала гипермутатор способствует поднятию по ландшафту, затем, при приближении к пику выгоднее становится антимутатор. Мутаторы Barrick
- 13. Мутаторы нужны только короткий период времени a. Работа мутаторов различной частоты после изменения условий среды. b.
- 14. SOS-ответ SOS-ответ – бактериальная система, приводящая к активации неточной системы репарации при стрессовых условиях.
- 15. Белки UmuC и UmuD позволяют ДНК полимеразе III пропускать многие ошибки в ДНК, в том числе
- 16. Помогает ли стресс-индуцированный мутагенез адаптироваться? SIMe – фенотип, способствующий стресс-индуцированному мутагенезу. NM – фенотип с нормальным
- 17. Повтор-индуцированный мутагенез (RIP; repeat-induced point mutation) RIP – способ защиты грибов отделов Ascomycota и Basidiomycota от
- 18. Прионы и эволюция Прион – это белок, который имеет несколько стабильных конформаций, как минимум одна из
- 19. Прион дрожжей PSI+ True et al., 2004 При трансформации белка-регулятора терминации трансляции Sup35 в прионную форму
- 20. Регуляция метаболизма – еще один дрожжевой прион GAR
- 21. Прионы выгоднее мутаций из-за большей гибкости Masel & Bergmann, 2003
- 22. Взаимоотношения бактерий и дрожжей Jarosz et al., 2014 Бактерии способны активировать дрожжевой прион GAR, уменьшая таким
- 23. Прионы эволюционируют Прионы – глобальные минимумы свободной энергии для данной аминокислотной последовательности.
- 24. Как минимум для приона млекопитающих PrP была показана конформационная гетерогенность популяции прионов. Отбор среди этих разных
- 25. Было показано, что прионы могут вырабатывать устойчивость к препаратам (сваинзонин). В результате эксперимента также возникли прионы,
- 27. Скачать презентацию

Evolvability (способность к эволюции) – способность популяции создавать генетическое разнообразие, и,
Evolvability (способность к эволюции) – способность популяции создавать генетическое разнообразие, и,

Ландшафты приспособленности
Ландшафт приспособленности – математическая модель, позволяющая анализировать относительную приспособленность различных
Ландшафты приспособленности
Ландшафт приспособленности – математическая модель, позволяющая анализировать относительную приспособленность различных

Как найти улучшенный фенотип, при этом не потеряв предыдущего?
Иметь большую выборку
Как найти улучшенный фенотип, при этом не потеряв предыдущего?
Иметь большую выборку

Робустность
Робустность – способность биологической системы поддерживать стабильность определенного фенотипа при действии
Робустность
Робустность – способность биологической системы поддерживать стабильность определенного фенотипа при действии
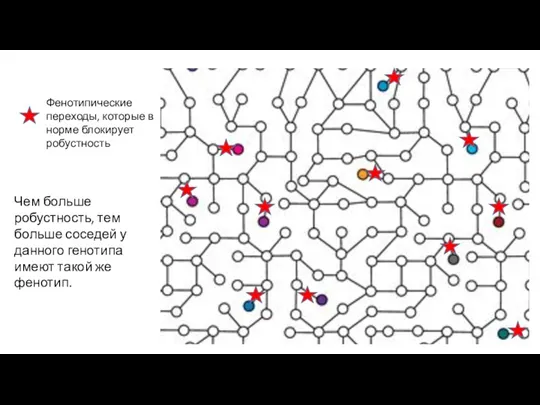
Фенотипические переходы, которые в норме блокирует робустность
Чем больше робустность, тем больше
Фенотипические переходы, которые в норме блокирует робустность
Чем больше робустность, тем больше

Шапероны – один из механизмов мутационной робустности
Шапероны способствуют мутационной робустности пытаясь
Шапероны – один из механизмов мутационной робустности
Шапероны способствуют мутационной робустности пытаясь
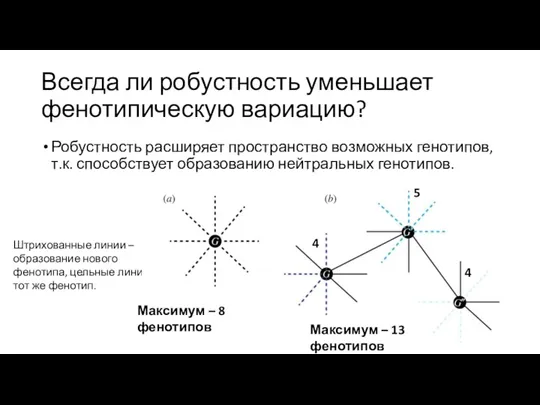
Всегда ли робустность уменьшает фенотипическую вариацию?
Робустность расширяет пространство возможных генотипов, т.к.
Всегда ли робустность уменьшает фенотипическую вариацию?
Робустность расширяет пространство возможных генотипов, т.к.
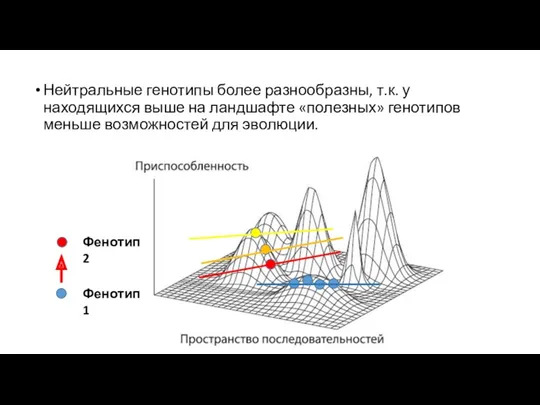
Нейтральные генотипы более разнообразны, т.к. у находящихся выше на ландшафте «полезных»
Нейтральные генотипы более разнообразны, т.к. у находящихся выше на ландшафте «полезных»
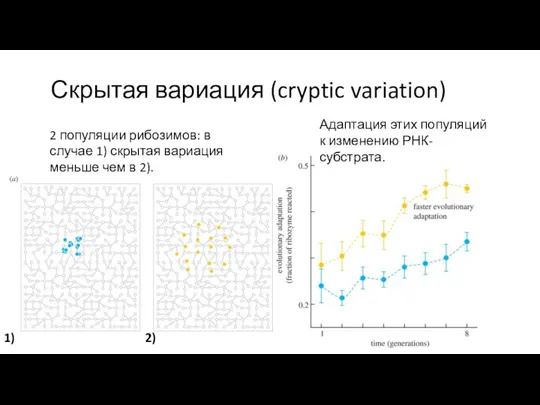
Скрытая вариация (cryptic variation)
Адаптация этих популяций к изменению РНК-субстрата.
Скрытая вариация (cryptic variation)
Адаптация этих популяций к изменению РНК-субстрата.

Способы ускорения поиска нужного генотипа среди скрытых
В стрессовой ситуации (например, изменение
Способы ускорения поиска нужного генотипа среди скрытых
В стрессовой ситуации (например, изменение
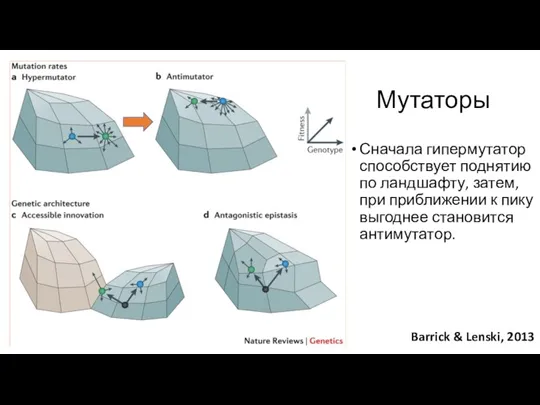
Сначала гипермутатор способствует поднятию по ландшафту, затем, при приближении к пику
Сначала гипермутатор способствует поднятию по ландшафту, затем, при приближении к пику
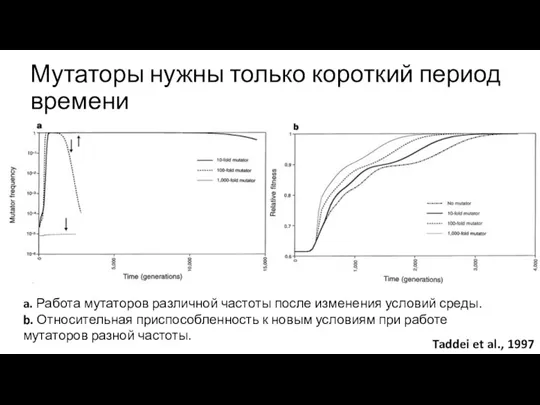
Мутаторы нужны только короткий период времени
a. Работа мутаторов различной частоты после
Мутаторы нужны только короткий период времени
a. Работа мутаторов различной частоты после
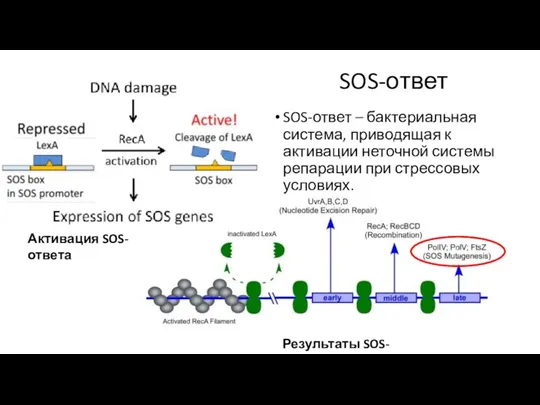
SOS-ответ
SOS-ответ – бактериальная система, приводящая к активации неточной системы репарации при
SOS-ответ
SOS-ответ – бактериальная система, приводящая к активации неточной системы репарации при
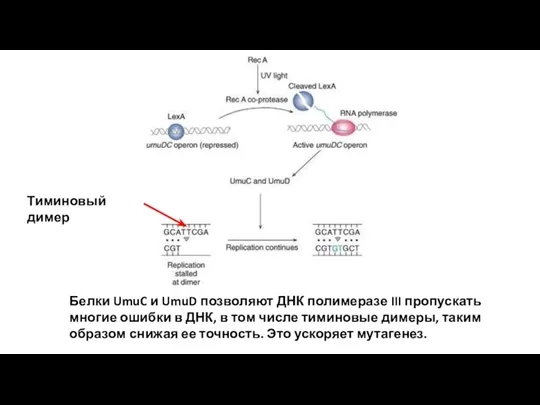
Белки UmuC и UmuD позволяют ДНК полимеразе III пропускать многие ошибки
Белки UmuC и UmuD позволяют ДНК полимеразе III пропускать многие ошибки
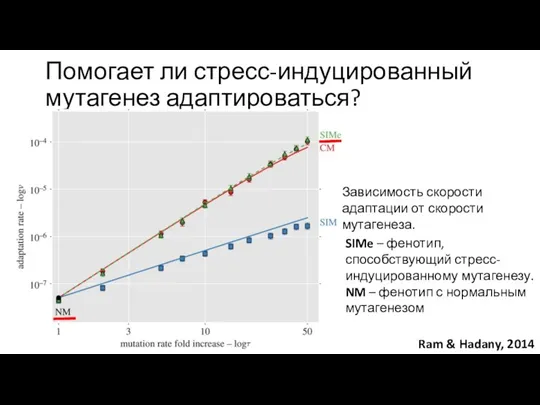
Помогает ли стресс-индуцированный мутагенез адаптироваться?
SIMe – фенотип, способствующий стресс-индуцированному мутагенезу.
NM –
Помогает ли стресс-индуцированный мутагенез адаптироваться?
SIMe – фенотип, способствующий стресс-индуцированному мутагенезу.
NM –

Повтор-индуцированный мутагенез (RIP; repeat-induced point mutation)
RIP – способ защиты грибов отделов
Повтор-индуцированный мутагенез (RIP; repeat-induced point mutation)
RIP – способ защиты грибов отделов
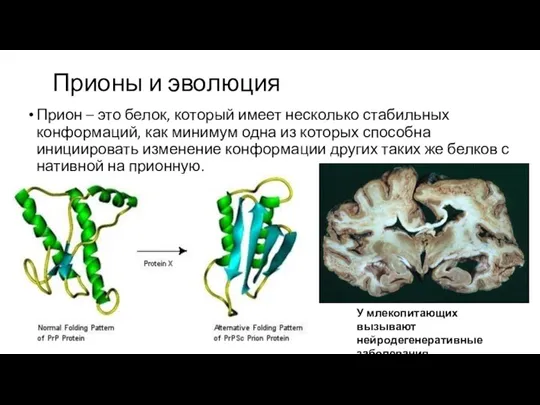
Прионы и эволюция
Прион – это белок, который имеет несколько стабильных конформаций,
Прионы и эволюция
Прион – это белок, который имеет несколько стабильных конформаций,
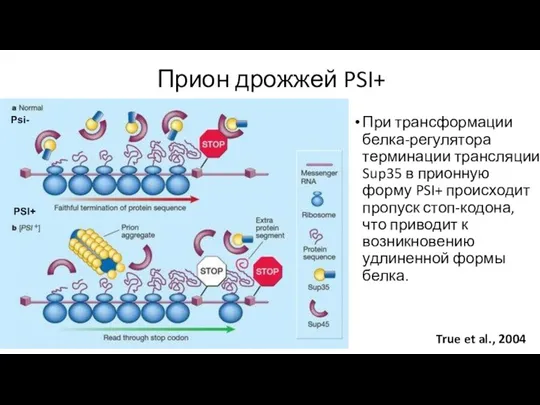
Прион дрожжей PSI+
True et al., 2004
При трансформации белка-регулятора терминации трансляции Sup35
Прион дрожжей PSI+
True et al., 2004
При трансформации белка-регулятора терминации трансляции Sup35
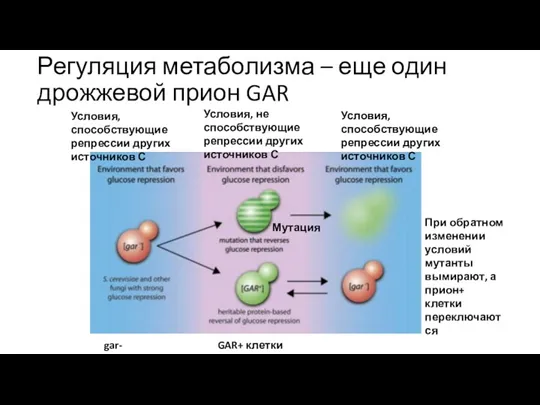
Регуляция метаболизма – еще один дрожжевой прион GAR
Регуляция метаболизма – еще один дрожжевой прион GAR
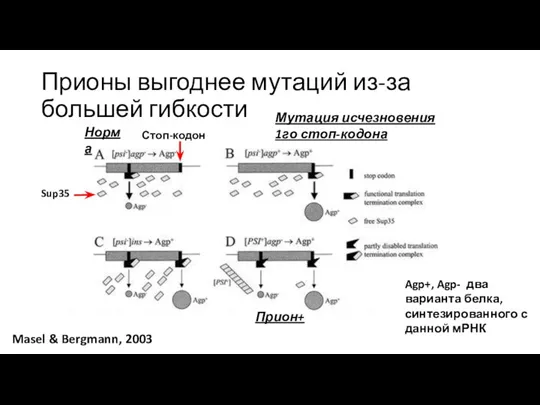
Прионы выгоднее мутаций из-за большей гибкости
Masel & Bergmann, 2003
Прионы выгоднее мутаций из-за большей гибкости
Masel & Bergmann, 2003
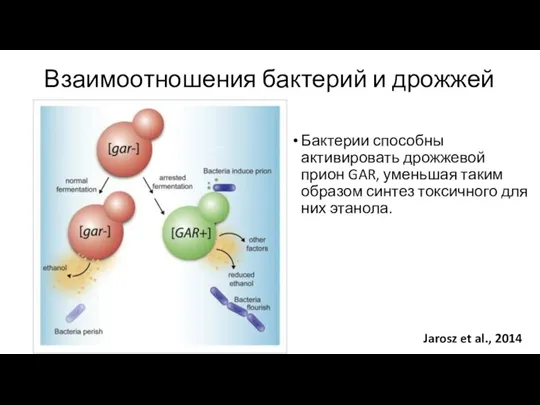
Взаимоотношения бактерий и дрожжей
Jarosz et al., 2014
Бактерии способны активировать дрожжевой прион
Взаимоотношения бактерий и дрожжей
Jarosz et al., 2014
Бактерии способны активировать дрожжевой прион
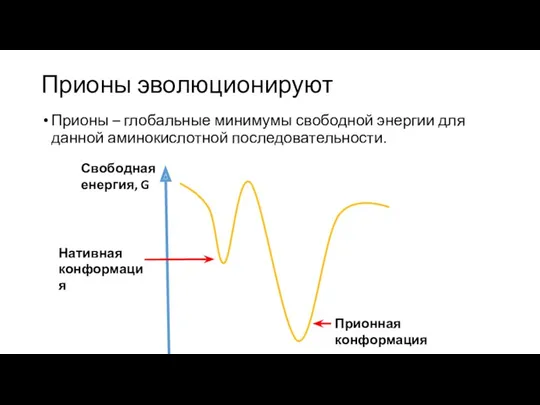
Прионы эволюционируют
Прионы – глобальные минимумы свободной энергии для данной аминокислотной последовательности.
Прионы эволюционируют
Прионы – глобальные минимумы свободной энергии для данной аминокислотной последовательности.
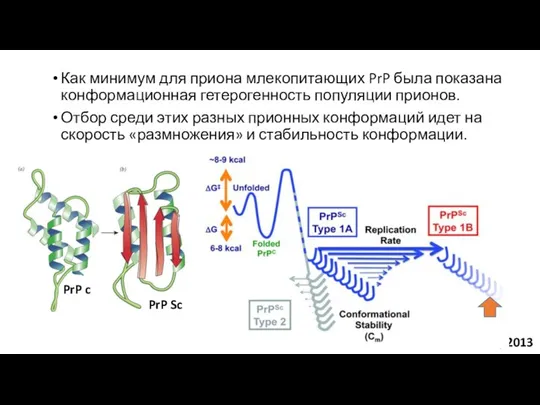
Как минимум для приона млекопитающих PrP была показана конформационная гетерогенность популяции
Как минимум для приона млекопитающих PrP была показана конформационная гетерогенность популяции
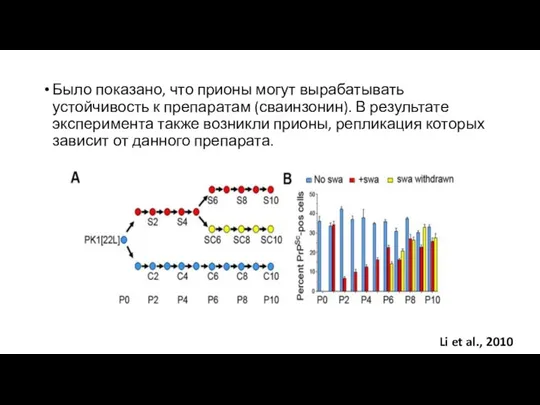
Было показано, что прионы могут вырабатывать устойчивость к препаратам (сваинзонин). В
Было показано, что прионы могут вырабатывать устойчивость к препаратам (сваинзонин). В
 Исследовательский проект на тему: Лук – зеленый друг
Исследовательский проект на тему: Лук – зеленый друг Многообразие грибов. Значение грибов в природе и жизни человека
Многообразие грибов. Значение грибов в природе и жизни человека Строение и свойства белков.
Строение и свойства белков. Размножение организмов
Размножение организмов Улучшение семейного бюджета на примере разведения пчелиного хозяйства
Улучшение семейного бюджета на примере разведения пчелиного хозяйства Регуляторные системы организма. Биохимия эндокринной системы
Регуляторные системы организма. Биохимия эндокринной системы Генетика пола. Наследование сцепленное с полом
Генетика пола. Наследование сцепленное с полом Фотосинтез. Световая фаза фотосинтеза
Фотосинтез. Световая фаза фотосинтеза Введение в современную биотехнологию БИООБЪЕКТ
Введение в современную биотехнологию БИООБЪЕКТ Пищеварение в тонком и толстом кишечнике. Барьерная роль печени
Пищеварение в тонком и толстом кишечнике. Барьерная роль печени Воздушное питание растений. Фотосинтез
Воздушное питание растений. Фотосинтез Обмен веществ и энергии
Обмен веществ и энергии Хайваннар. Йорт хайваннары. Кыргый хайваннар
Хайваннар. Йорт хайваннары. Кыргый хайваннар Лабораторная диагностика паразитарных заболеваний. Занятие 11
Лабораторная диагностика паразитарных заболеваний. Занятие 11 Разработка урока по теме Клетка. Клеточная теория строения организмов 10 класс
Разработка урока по теме Клетка. Клеточная теория строения организмов 10 класс Дидактическая игра Корзина грецких орехов
Дидактическая игра Корзина грецких орехов Инженерия антител
Инженерия антител Строение цветка. Опыление растений
Строение цветка. Опыление растений Загадка человека. Наследственность. Урок 3
Загадка человека. Наследственность. Урок 3 Циклы развития растений. Подготовка к выполнению заданий ЕГЭ С5
Циклы развития растений. Подготовка к выполнению заданий ЕГЭ С5 Фитопатогенді бактериялардың таксономикалық бөлімдері
Фитопатогенді бактериялардың таксономикалық бөлімдері Возрастные особенности температурной сенсорной системы. Механизмы закаливания
Возрастные особенности температурной сенсорной системы. Механизмы закаливания Возникновение жизненного цикла и онтогенеза metazoa
Возникновение жизненного цикла и онтогенеза metazoa Апоптоз. Исследования феномена программируемой клеточной смерти
Апоптоз. Исследования феномена программируемой клеточной смерти Кобчик - птица 2021 года
Кобчик - птица 2021 года Цитологические основы наследования признаков при моногибридном скрещивании
Цитологические основы наследования признаков при моногибридном скрещивании Антибиотики и их влияние на болезнетворные бактерии
Антибиотики и их влияние на болезнетворные бактерии ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Антропогенез. Эволюция человека. (В одиннадцати частях). 9 класс
ПРЕЗЕНТАЦИЯ ДЛЯ ИНТЕРАКТИВНОЙ ДОСКИ. Антропогенез. Эволюция человека. (В одиннадцати частях). 9 класс