- Обмен углеводов

Содержание
- 5. Всасывание моносахаридов
- 8. Аэробный дихотомический путь катаболизма глюкозы Глюкоза в тканях организма человека и животных может разрушаться двумя путями
- 11. Окисление пирувата
- 12. Окисление пирувата Далее две молекулы пирувата в митохондриях подвергаются окислительному декарбоксилированию. В этой реакции образуются две
- 13. Анаэробный дихотомический распад глюкозы В организме человека и животных при недостаточном поступлении кислорода в клетки возможен,
- 14. Брожение и гликолитичекая оксиредукция Анаэробные механизм дихотомического пути распада глюкозы имеет место и у некоторых бактерий.
- 17. Глюконеогенез
- 18. Глюконеогенез
- 19. Эффект Пастера Для того чтобы аэробные организмы могли покрыть свои энергетические потребности в анаэробных условиях, необходима
- 20. Пентозофосфатный путь катаболизма глюкозы Этот путь катаболизма широко представлен в различных тканях человека и животных (печень,
- 21. Биологическая роль пентозного цикла Биологическая роль пентозного цикла состоит в следующем: 1. В этом пути при
- 22. Фазы пентозного цикла Пентозный цикл катаболизма глюкозы идет в две фазы: Первая фаза - окисление шести
- 24. далее:
- 25. 6(глюкозо-6-фосфат)+12НАДФ+7НОН 5(глюкозо-6-фосфат)+6СО2+12НАДФН2+Н3РО4
- 26. Пентозный путь оксиления глюкозы Пентозофосфатный путь катаболизма углеводов активен прежде всего, в тех органах и тканях,
- 27. Биосинтез гликогена Фермент синтеза гликогена гликогенсинтетаза "а" (активная форма ), под воздействием цАМФ-зависимой протеинкиназы переходит в
- 28. Мобилизация гликогена Мобилизация гликогена начинается с возникновения готовности и необходимости организма выполнять мышечную работу, например, при
- 29. Регуляция синтеза и распада гликогена. Ключевую роль в регуляции синтеза и распада гликогена играют ферменты гликогенсинтетаза
- 30. Частные пути углеводного обмена. Галактоземии. Особый интерес вызывает метаболизм галактозы в связи с существованием наследственного заболевания
- 31. Превращение галактозы в глюкозу Превращение галактозы в глюкозу может иным путем при участии фермента катализирующего синтез
- 32. Галактоза Галактоза необходима для образования цереброзидов, протеогликанов и гликопротеинов. В кишечнике и печени галактоза превращается в
- 33. фруктоза Изомер глюкозы фруктоза (левулоза) в крови присутствует в очень низких количествах (55.5-333,0 мкмоль/л). При врожденной
- 34. лактоза Основной углевод молока лактоза образуется путем переноса галактозильного остатка от УДФ-галактозы непосредственно на глюкозу под
- 35. Структурные компоненты глюкозамингликанов Из глюкозы в организме человека и животных синтезируются структурные компоненты глюкозамингликанов: глюкуроновая кислота,
- 37. Гетерополисахариды К углеводам относятся гетерополисахариды (мукополисахариды), которые в основном находятся в межклеточном веществе в комплексе с
- 38. Гиалуроновая кислота Гиалуроновая кислота - несульфатированный гетерополисахарид с линейной структурой и самой большой молекулярной массой из
- 40. Разрушение гиалуроновой кислоты Разрушение этого биополимера осуществляет специальный фермент - гиалуронидаза. Гиалуронидаза, разрушая гиалуроновую кислоту, повышает
- 41. Хондроитинсульфаты Хондроитинсульфаты - наиболее распространенные кислые гетерополисахариды. Мономер этих полисахаридов состоит из глюкуроновой кислоты соединенный 1,3-бета-
- 42. Дерматансульфаты Мономер дерматансульфатов образован альфа-L-идуроновой кислотой (изомер глюкуроновой кислоты) соединенной 1,3 альфа-гликозидной связью с N-ацетилгалактозамин-4-сульфатом. Дерматансульфаты
- 43. Кератансульфаты Кератансульфаты состоят из мономеров образованных бета-галактозой соединенной 1,4-бета-гликозидной связью с N-ацетилглюкозамин-6 сульфатом. Кератансульфаты находятся в
- 44. Гепарин Гепарин в отличие от остальных кислых гетерополисахаридов не являются структурными компонентами соединительной ткани. Он синтезируется
- 45. Гликопротеины Углеводная часть гликопротеинов, к которым относятся многие ферменты, гормоны, транспортные белки крови, структурные белки, является
- 46. Гликопротеины - клеточные рецепторы Например,белок плазмы церулоплазмин содержит на конце углеводной части сиаловую кислоту. Если удалить
- 47. Гликопротеины плазмы крови В плазме крови содержится много разных гликопротеинов, и если они утрачивают концевую сиаловую
- 48. Гликозидозы Синтез гетерополисахаридов происходит при участии очень большой группы ферментов гликозилтрансфераз, связанных в клетках с мембранами
- 49. Регуляция и нарушения обмена углеводов. Уровень глюкозы в крови (3,5 - 5,7 ммоль/л) зависит от многих
- 50. Гипергликемии По происхождению различают гипергликемии физиологические и патологические. К физиологическим гипергликемиям относятся алиментарные, возникающие при одномоментном
- 52. Сахарный диабет Значительное повышение содержания глюкозы в крови отмечается при сахарном диабете, возникающем в результате недостаточной
- 53. Диагностическое значение определения гексоз Гексозы (галактоза, манноза, фукоза и др.) входят в состав различных гликопротеидов. Диагностическое
- 54. Молочная кислота Значительное количество молочной кислоты образуется при усиленной физической работе в мышцах, откуда она током
- 55. Пируват Пируват (пировиноградная кислота) является одним из центральных метаболитов углеводного обмена. При окислительном декарбоксилировании пирувата образуется
- 56. Сиаловые кислоты Сиаловые кислоты являются структурными компонентами гетерополисахаридов, входящих в состав гликопротеидов. При поражении соединительной ткани
- 58. Скачать презентацию
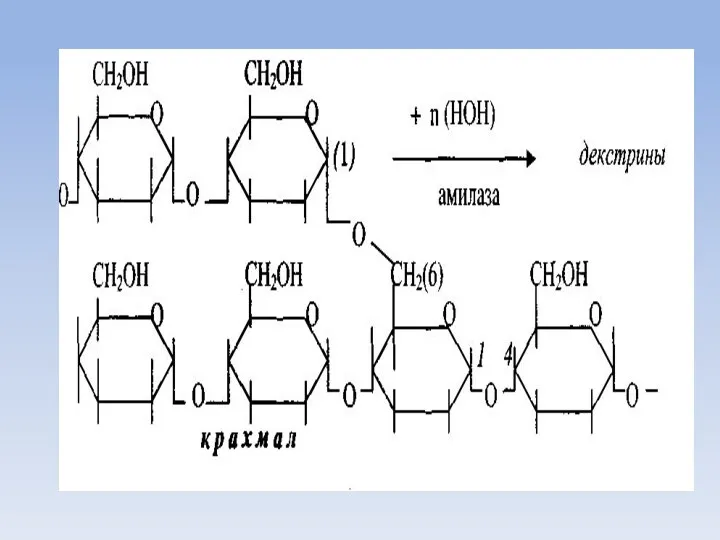
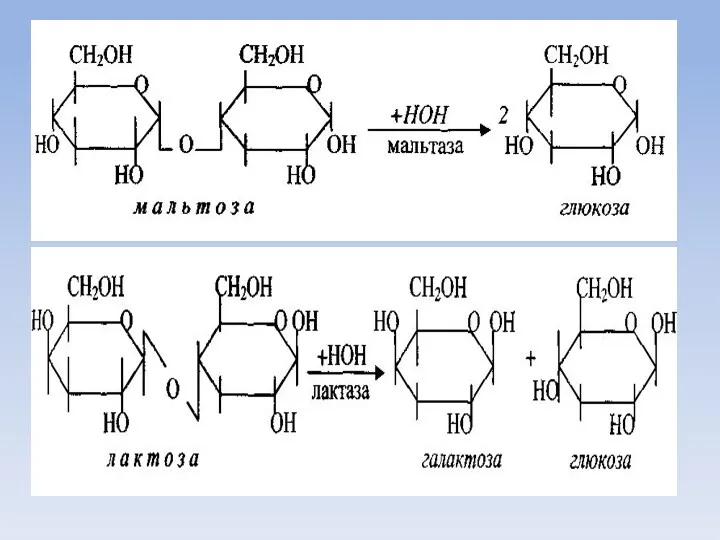
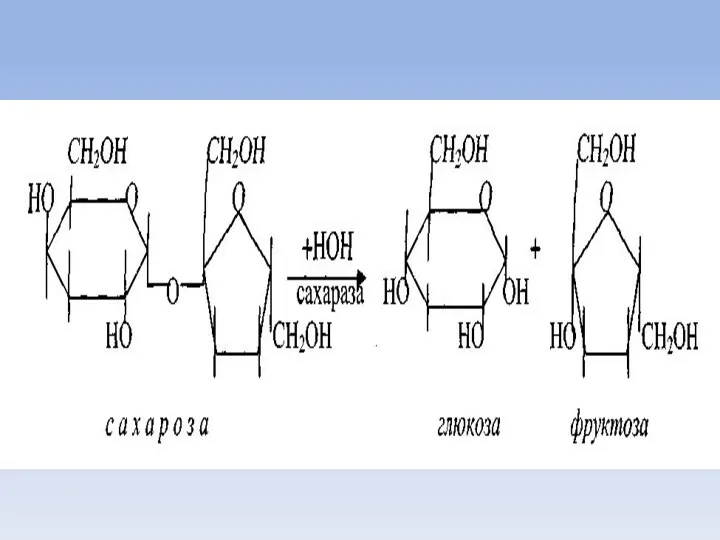

Всасывание моносахаридов
Всасывание моносахаридов
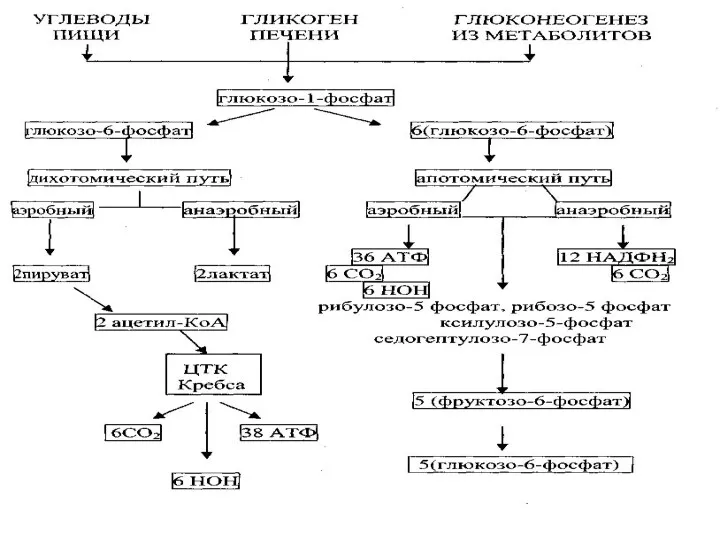
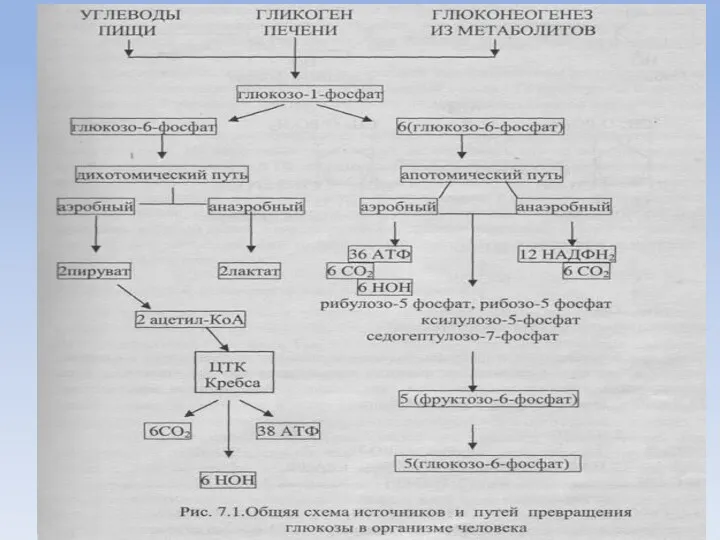

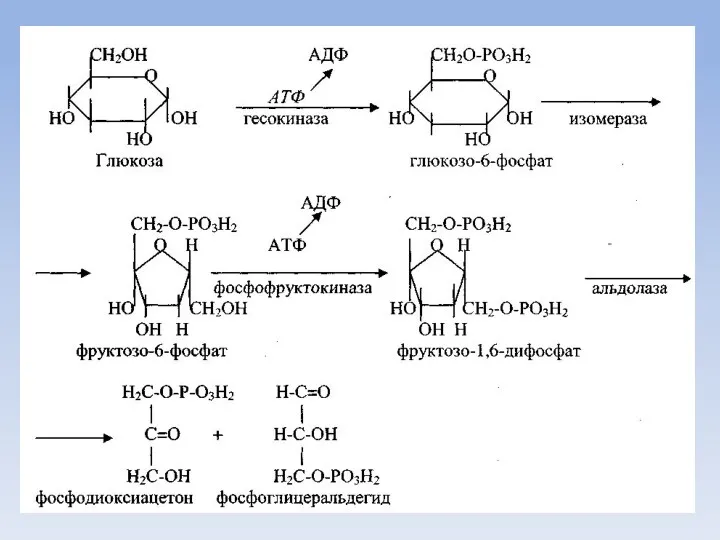
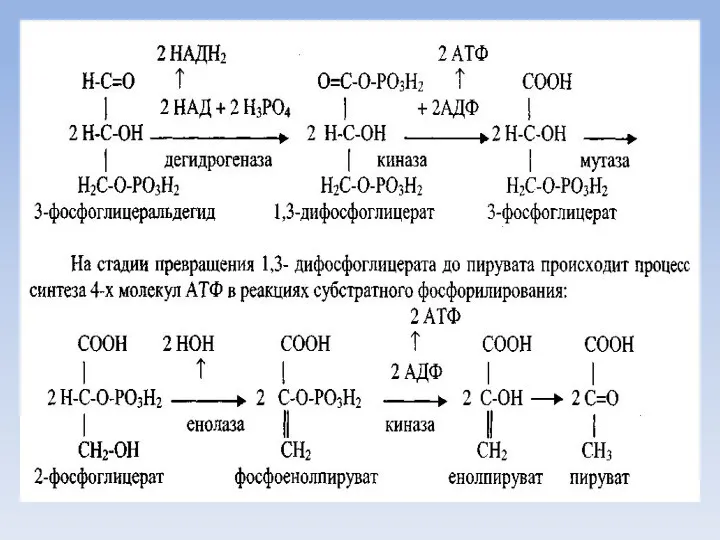
Аэробный дихотомический путь катаболизма глюкозы
Глюкоза в тканях организма человека и
Аэробный дихотомический путь катаболизма глюкозы
Глюкоза в тканях организма человека и
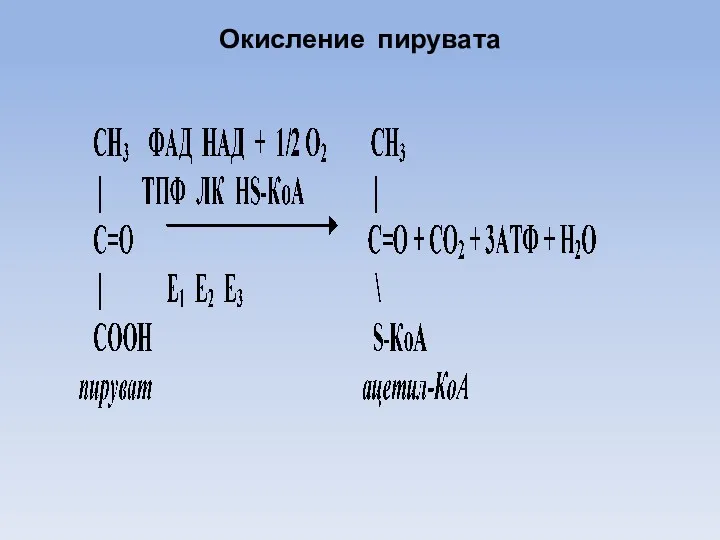
Окисление пирувата
Окисление пирувата

Окисление пирувата
Далее две молекулы пирувата в митохондриях подвергаются окислительному декарбоксилированию.
В
Окисление пирувата
Далее две молекулы пирувата в митохондриях подвергаются окислительному декарбоксилированию.
В

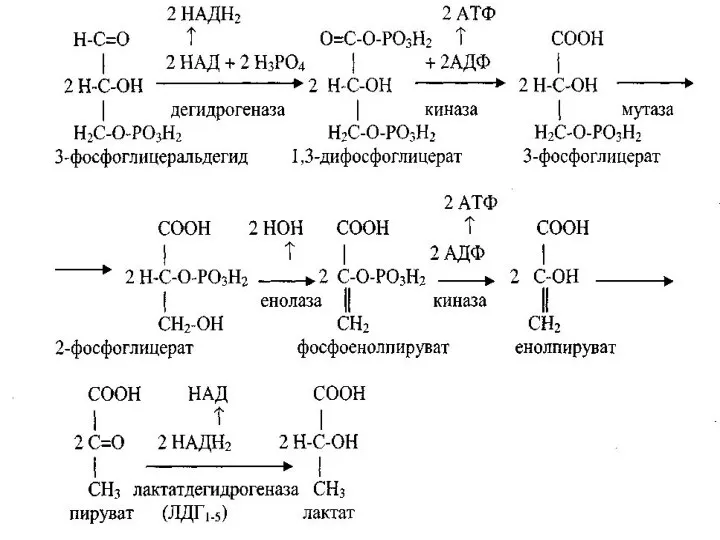
Анаэробный дихотомический распад глюкозы
В организме человека и животных при недостаточном поступлении
Анаэробный дихотомический распад глюкозы
В организме человека и животных при недостаточном поступлении

Брожение и гликолитичекая оксиредукция
Анаэробные механизм дихотомического пути распада глюкозы имеет место
Брожение и гликолитичекая оксиредукция
Анаэробные механизм дихотомического пути распада глюкозы имеет место
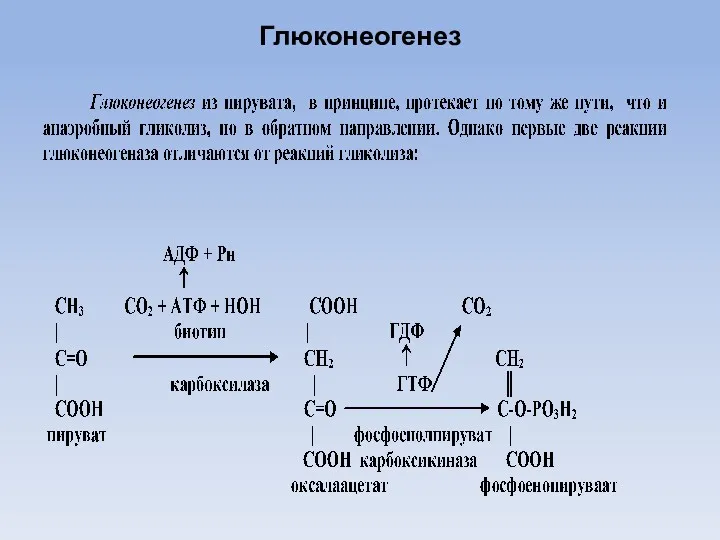
Глюконеогенез
Глюконеогенез
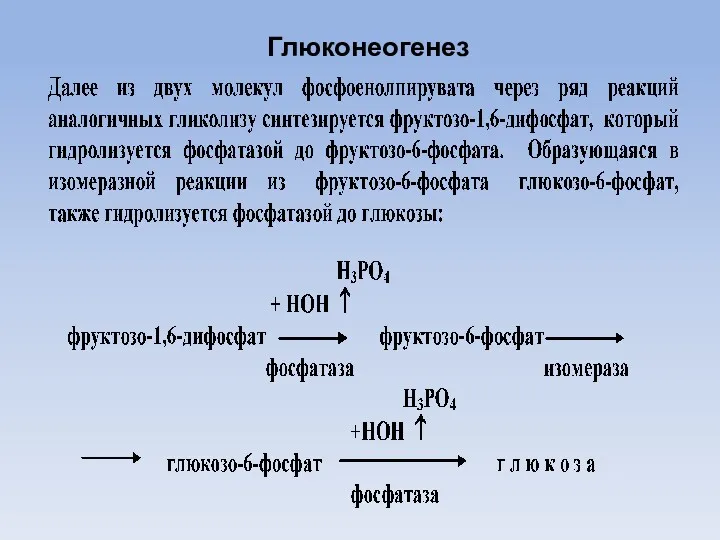
Глюконеогенез
Глюконеогенез

Эффект Пастера
Для того чтобы аэробные организмы могли покрыть свои энергетические потребности
Эффект Пастера
Для того чтобы аэробные организмы могли покрыть свои энергетические потребности

Пентозофосфатный путь катаболизма глюкозы
Этот путь катаболизма широко представлен в различных
Пентозофосфатный путь катаболизма глюкозы
Этот путь катаболизма широко представлен в различных

Биологическая роль пентозного цикла
Биологическая роль пентозного цикла состоит в следующем:
1.
Биологическая роль пентозного цикла
Биологическая роль пентозного цикла состоит в следующем:
1.

Фазы пентозного цикла
Пентозный цикл катаболизма глюкозы идет в две фазы:
Первая фаза
Фазы пентозного цикла
Пентозный цикл катаболизма глюкозы идет в две фазы:
Первая фаза
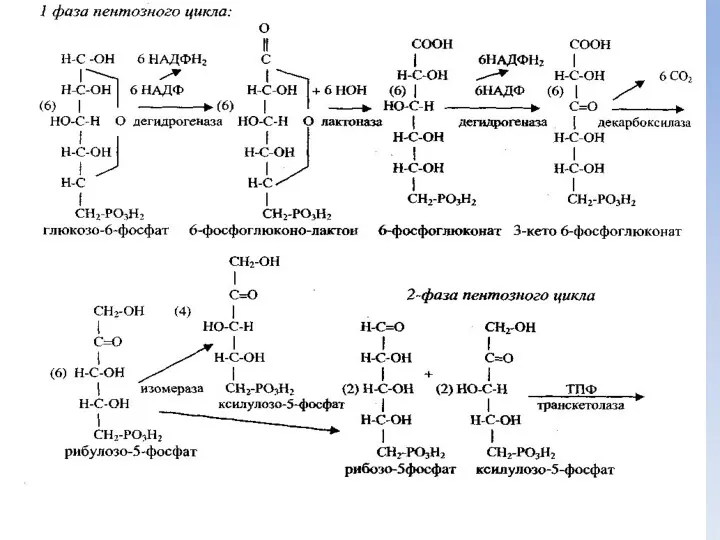
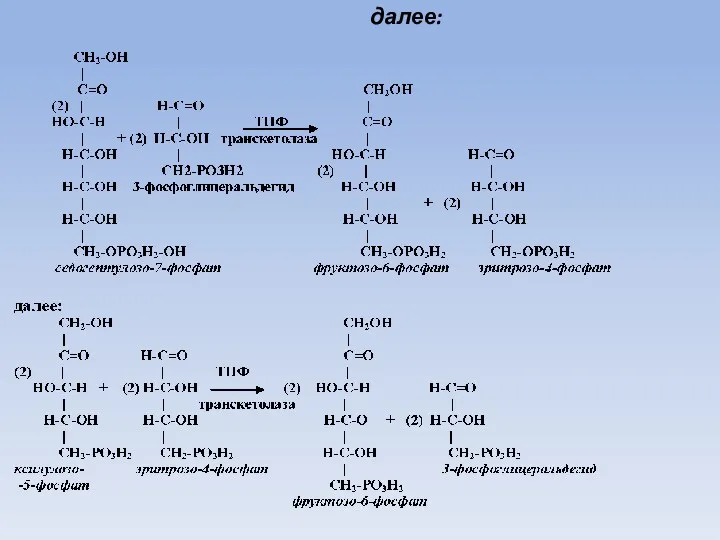
далее:
далее:

6(глюкозо-6-фосфат)+12НАДФ+7НОН
5(глюкозо-6-фосфат)+6СО2+12НАДФН2+Н3РО4
6(глюкозо-6-фосфат)+12НАДФ+7НОН
5(глюкозо-6-фосфат)+6СО2+12НАДФН2+Н3РО4

Пентозный путь оксиления глюкозы
Пентозофосфатный путь катаболизма углеводов активен прежде всего, в
Пентозный путь оксиления глюкозы
Пентозофосфатный путь катаболизма углеводов активен прежде всего, в

Биосинтез гликогена
Фермент синтеза гликогена гликогенсинтетаза "а" (активная форма ), под
Биосинтез гликогена
Фермент синтеза гликогена гликогенсинтетаза "а" (активная форма ), под

Мобилизация гликогена
Мобилизация гликогена начинается с возникновения готовности и необходимости организма выполнять
Мобилизация гликогена
Мобилизация гликогена начинается с возникновения готовности и необходимости организма выполнять

Регуляция синтеза и распада гликогена.
Ключевую роль в регуляции синтеза и распада
Регуляция синтеза и распада гликогена.
Ключевую роль в регуляции синтеза и распада

Частные пути углеводного обмена. Галактоземии.
Особый интерес вызывает метаболизм галактозы в
Частные пути углеводного обмена. Галактоземии.
Особый интерес вызывает метаболизм галактозы в

Превращение галактозы в глюкозу
Превращение галактозы в глюкозу может иным путем при
Превращение галактозы в глюкозу
Превращение галактозы в глюкозу может иным путем при

Галактоза
Галактоза необходима для образования цереброзидов, протеогликанов и гликопротеинов. В кишечнике и
Галактоза
Галактоза необходима для образования цереброзидов, протеогликанов и гликопротеинов. В кишечнике и

фруктоза
Изомер глюкозы фруктоза (левулоза) в крови присутствует в очень низких количествах
фруктоза
Изомер глюкозы фруктоза (левулоза) в крови присутствует в очень низких количествах

лактоза
Основной углевод молока лактоза образуется путем переноса галактозильного остатка от УДФ-галактозы
лактоза
Основной углевод молока лактоза образуется путем переноса галактозильного остатка от УДФ-галактозы

Структурные компоненты глюкозамингликанов
Из глюкозы в организме человека и животных синтезируются структурные
Структурные компоненты глюкозамингликанов
Из глюкозы в организме человека и животных синтезируются структурные
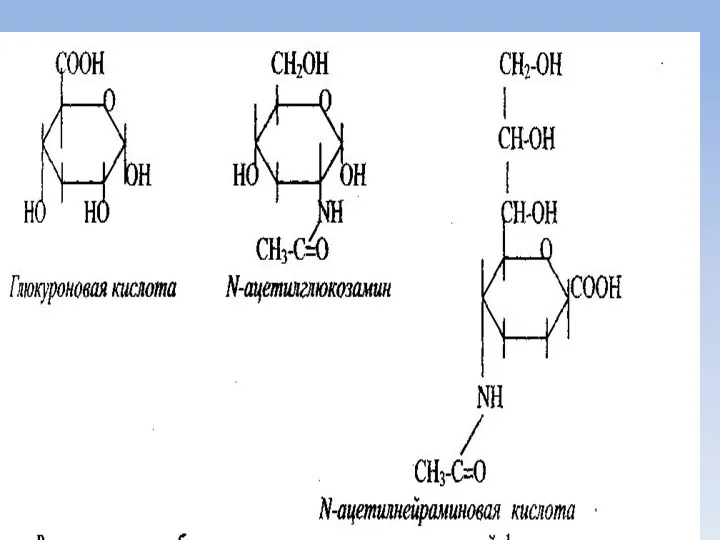

Гетерополисахариды
К углеводам относятся гетерополисахариды (мукополисахариды), которые в основном находятся в межклеточном
Гетерополисахариды
К углеводам относятся гетерополисахариды (мукополисахариды), которые в основном находятся в межклеточном

Гиалуроновая кислота
Гиалуроновая кислота - несульфатированный гетерополисахарид с линейной структурой и самой
Гиалуроновая кислота
Гиалуроновая кислота - несульфатированный гетерополисахарид с линейной структурой и самой
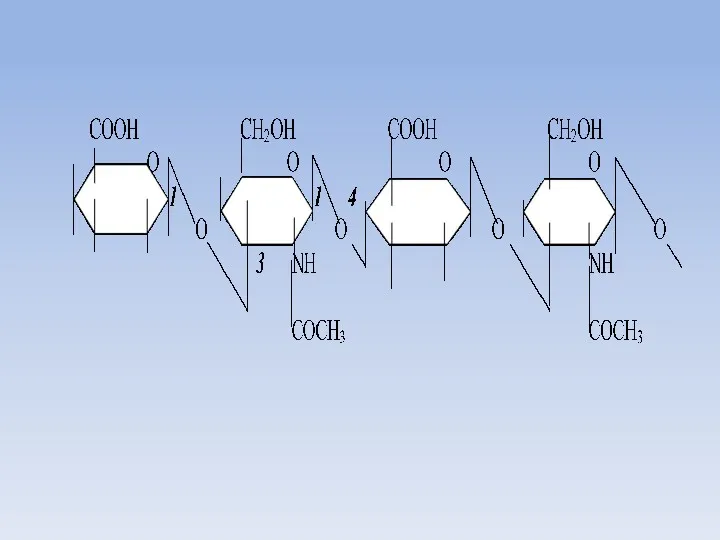

Разрушение гиалуроновой кислоты
Разрушение этого биополимера осуществляет специальный фермент - гиалуронидаза.
Гиалуронидаза,
Разрушение гиалуроновой кислоты
Разрушение этого биополимера осуществляет специальный фермент - гиалуронидаза.
Гиалуронидаза,

Хондроитинсульфаты
Хондроитинсульфаты - наиболее распространенные кислые гетерополисахариды.
Мономер этих полисахаридов состоит из
Хондроитинсульфаты
Хондроитинсульфаты - наиболее распространенные кислые гетерополисахариды.
Мономер этих полисахаридов состоит из

Дерматансульфаты
Мономер дерматансульфатов образован альфа-L-идуроновой кислотой (изомер глюкуроновой кислоты) соединенной 1,3 альфа-гликозидной
Дерматансульфаты
Мономер дерматансульфатов образован альфа-L-идуроновой кислотой (изомер глюкуроновой кислоты) соединенной 1,3 альфа-гликозидной

Кератансульфаты
Кератансульфаты состоят из мономеров образованных бета-галактозой соединенной 1,4-бета-гликозидной связью с N-ацетилглюкозамин-6
Кератансульфаты
Кератансульфаты состоят из мономеров образованных бета-галактозой соединенной 1,4-бета-гликозидной связью с N-ацетилглюкозамин-6

Гепарин
Гепарин в отличие от остальных кислых гетерополисахаридов не являются структурными компонентами
Гепарин
Гепарин в отличие от остальных кислых гетерополисахаридов не являются структурными компонентами

Гликопротеины
Углеводная часть гликопротеинов, к которым относятся многие ферменты, гормоны, транспортные белки
Гликопротеины
Углеводная часть гликопротеинов, к которым относятся многие ферменты, гормоны, транспортные белки

Гликопротеины - клеточные рецепторы
Например,белок плазмы церулоплазмин содержит на конце углеводной части
Гликопротеины - клеточные рецепторы
Например,белок плазмы церулоплазмин содержит на конце углеводной части

Гликопротеины плазмы крови
В плазме крови содержится много разных гликопротеинов, и
Гликопротеины плазмы крови
В плазме крови содержится много разных гликопротеинов, и

Гликозидозы
Синтез гетерополисахаридов происходит при участии очень большой группы ферментов гликозилтрансфераз, связанных
Гликозидозы
Синтез гетерополисахаридов происходит при участии очень большой группы ферментов гликозилтрансфераз, связанных

Регуляция и нарушения обмена углеводов.
Уровень глюкозы в крови (3,5 - 5,7
Регуляция и нарушения обмена углеводов.
Уровень глюкозы в крови (3,5 - 5,7

Гипергликемии
По происхождению различают гипергликемии физиологические и патологические.
К физиологическим гипергликемиям относятся
Гипергликемии
По происхождению различают гипергликемии физиологические и патологические.
К физиологическим гипергликемиям относятся
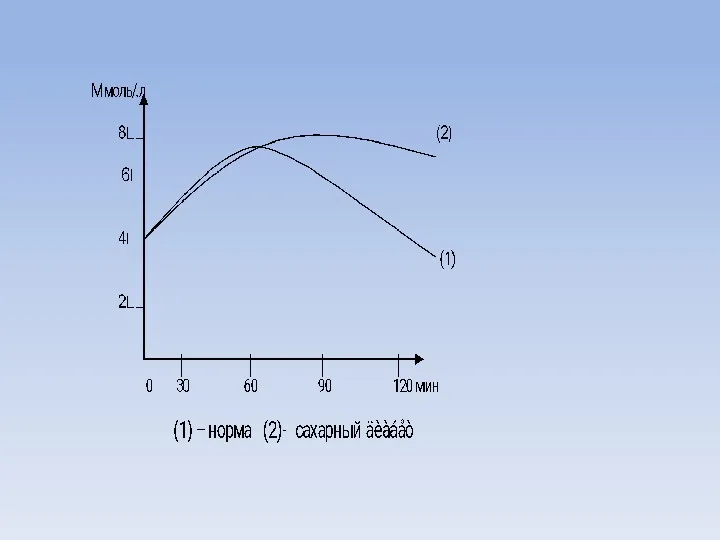

Сахарный диабет
Значительное повышение содержания глюкозы в крови отмечается при сахарном диабете,
Сахарный диабет
Значительное повышение содержания глюкозы в крови отмечается при сахарном диабете,

Диагностическое значение определения гексоз
Гексозы (галактоза, манноза, фукоза и др.) входят в
Диагностическое значение определения гексоз
Гексозы (галактоза, манноза, фукоза и др.) входят в

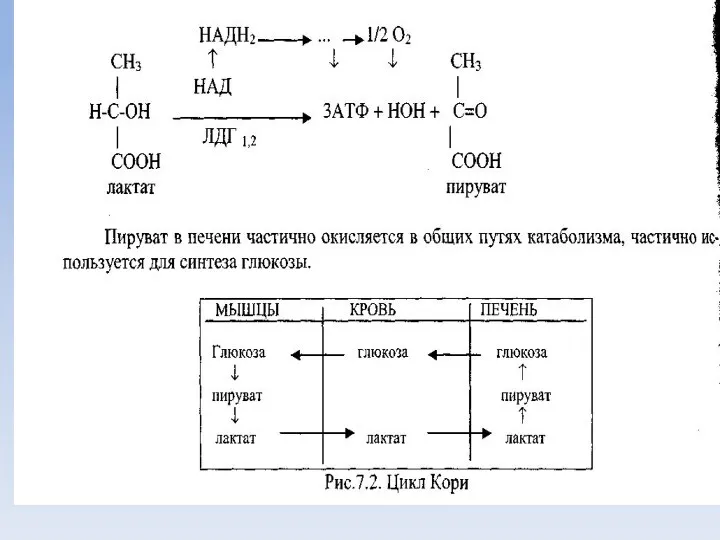
Молочная кислота
Значительное количество молочной кислоты образуется при усиленной физической работе в
Молочная кислота
Значительное количество молочной кислоты образуется при усиленной физической работе в

Пируват
Пируват (пировиноградная кислота) является одним из центральных метаболитов углеводного обмена.
При
Пируват
Пируват (пировиноградная кислота) является одним из центральных метаболитов углеводного обмена.
При

Сиаловые кислоты
Сиаловые кислоты являются структурными компонентами гетерополисахаридов, входящих в состав гликопротеидов.
Сиаловые кислоты
Сиаловые кислоты являются структурными компонентами гетерополисахаридов, входящих в состав гликопротеидов.
 Класс Головоногие моллюски
Класс Головоногие моллюски Орган слуха
Орган слуха Внешнее строение листа
Внешнее строение листа Цитоплазма и органеллы
Цитоплазма и органеллы Человек. Загадка человека
Человек. Загадка человека Пищеварительная система
Пищеварительная система Основные объекты аквакультуры РФ
Основные объекты аквакультуры РФ Гигантский короткомордый медведь
Гигантский короткомордый медведь Определение пола. 9 класс.
Определение пола. 9 класс. Водная среда обитания
Водная среда обитания Понятие об онтогенезе
Понятие об онтогенезе Происхождение жизни и развитие органического мира
Происхождение жизни и развитие органического мира Спинной мозг
Спинной мозг Драцена - это удивительное комнатное растение
Драцена - это удивительное комнатное растение Мал азығын жіктеу шығу тегі, химиялық құрамы мен қоректілігі бойынша
Мал азығын жіктеу шығу тегі, химиялық құрамы мен қоректілігі бойынша Многообразие млекопитающих. Отряд китообразные
Многообразие млекопитающих. Отряд китообразные Строение и функции головного мозга
Строение и функции головного мозга Нервная система. Спинной мозг
Нервная система. Спинной мозг Органы дыхания, строение и функции
Органы дыхания, строение и функции Designin a photobioreactor for growing microalgae at home
Designin a photobioreactor for growing microalgae at home 27 сентября - Международный день кролика
27 сентября - Международный день кролика Побеги. Морфология побега. Строение почек
Побеги. Морфология побега. Строение почек Размножение. Митоз и мейоз
Размножение. Митоз и мейоз Размножение ягодных и декоративных кустарников разными видами черенков
Размножение ягодных и декоративных кустарников разными видами черенков БГЦ как особый уровень организации жизни
БГЦ как особый уровень организации жизни Pathologic Protozoa (Lesson 1)
Pathologic Protozoa (Lesson 1) Культурные растения
Культурные растения Световая микроскопия. Занятие 1
Световая микроскопия. Занятие 1