- Ориентиры эмбрионов дрозофилы на ранних стадиях развития

Содержание
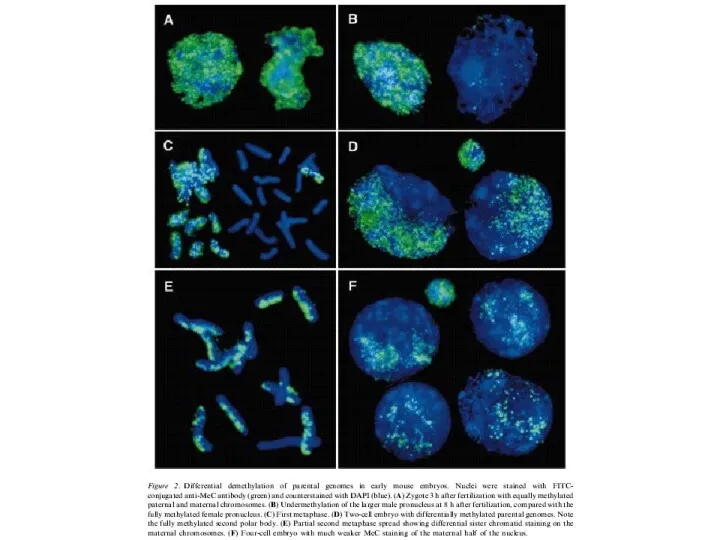
- 2. Схема развития пронуклеусов и образование зиготического генома мыши (Adenot et al., 1997) Мужской (серый) и женский
- 3. Сингамия – слияние пронуклеусов (A,B,D,E) и первый митоз (C,F). A-C – иммунофлуоресценция гистона H4-L5; D-F –
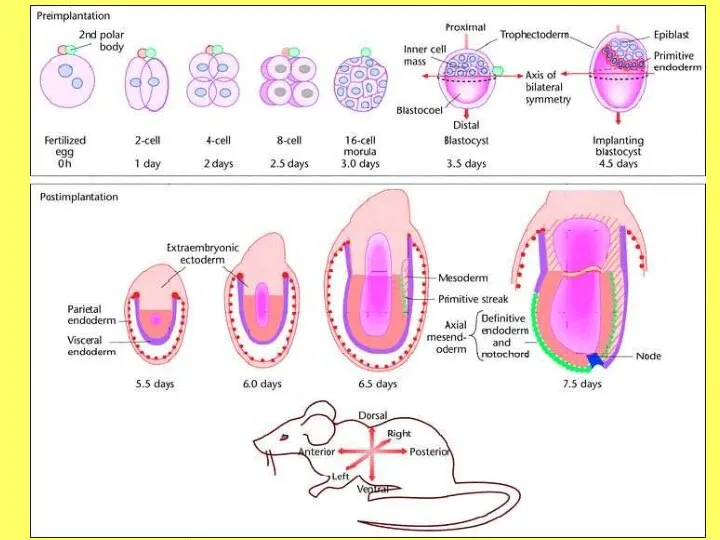
- 4. Стадии развития предимплантационных эмбрионов человека и мыши. А – после 3-х делений-дроблений зиготы 8-ми клеточный эмбрион
- 5. Поляризация предимплантационных эмбрионов мыши. А – 8-ми клеточной стадии все бластомеры поляризованы вдоль оси клеточного контакта,
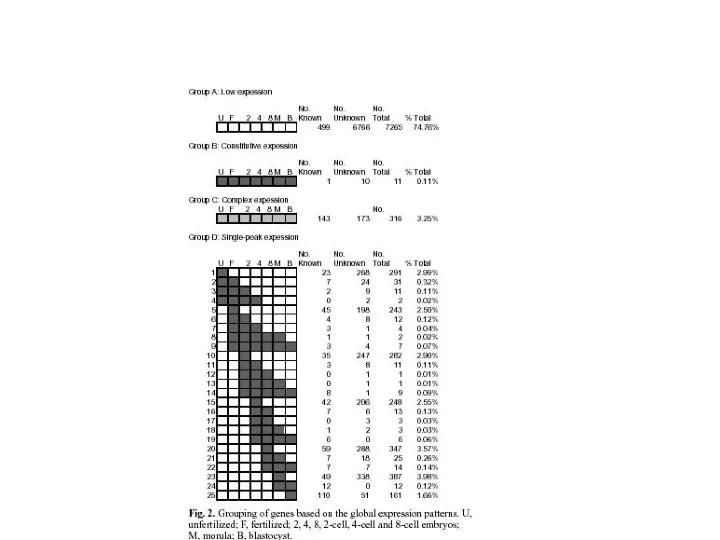
- 6. Важнейшие события с момента оплодотворения: Синтез белка происходит на материнских мРНК; Зиготические гены активируются в конце
- 7. Модели образования ТЭ. А – внутренние и внешние клетки имеют разное количество контактов, что определяет различия
- 8. Коммитирование апикальных клеток к дифференцировке в ТЭ, первое реальное событие дифференцировки в развитии млекопитающих. Предполагается что
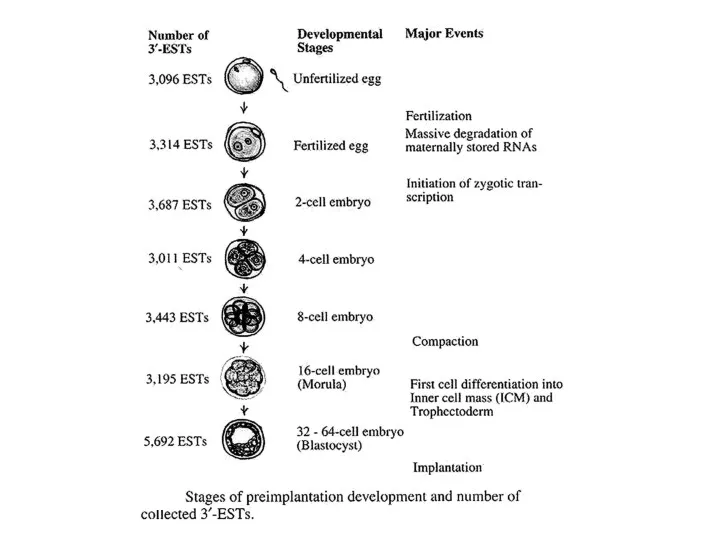
- 10. Временные клеточного цикла в первые часы после оплодотворения. М – микроинъекции конструкции с геном репортером, альфа-аматин
- 11. Анализ активности промотора конструкции с геном репортером (люцифераза). hCG –время введения хориогонина Синтез м РНК Синтез
- 12. Экспрессия транспозоновых элементов в зрелых ооцитах, 2-х клеточных эмбрионах и бластоцистах по данным анализа сДНК библиотек
- 13. Экспрессия ретротранспозонов на стадии зиготы и 2-хклеточной стадии В зиготах экспрессируются эндогенные ретровирусы, LINE-1 и SINE
- 14. 2-х клеточный эмбрион (d) и бластоциста (е) окрашенные антителами против MuERV-L-Gag. Зиготы и 2-х клеточные эмбрионы
- 19. Метилирование ДНК. Полуконсервативный способ репликации метилированных динуклеотидов CpG в геномной ДНК дает дочернюю нить, которая гемиметилирована
- 21. Иммунодетекция 5mC в мышиных и овечьих нормальных эмбрионах или полученных оплодовторением in vitro. Видно, что у
- 22. Схема развития ооцита и раннее развитие мыши. Метилирование ДНК материнского (красный цвет) и отцовского (синий) геномов.
- 23. Метилирование ДНК материнского (красный цвет) и отцовского (синий) геномов в раннем развитии коров в сравнении с
- 24. Эпигенетическое репрограммирование. Ремоделирование мужского и женского пронуклеусов. Транскрипционные (зеленый цвет) и репрессионные (красный) модификации с разной
- 25. Метилирование ДНК de novo в ВКМ, но значительно менее это выражено вТЭ мышей, коров, овец. H3K9ac
- 31. Скачать презентацию
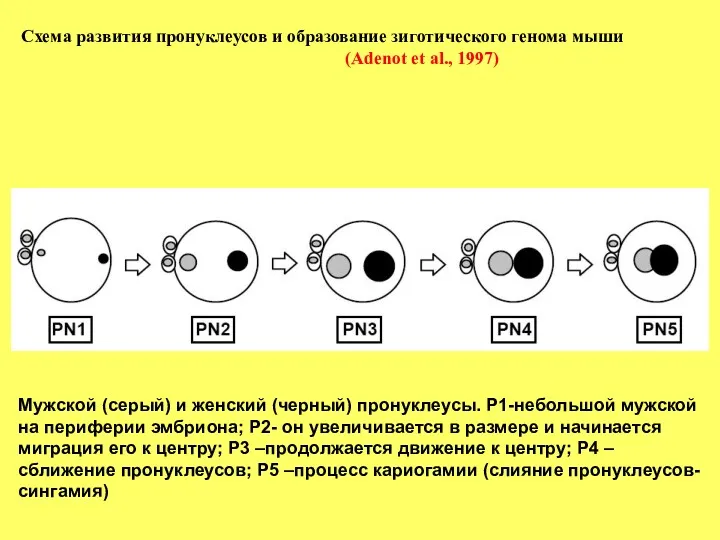
Схема развития пронуклеусов и образование зиготического генома мыши
(Adenot et al.,
Схема развития пронуклеусов и образование зиготического генома мыши
(Adenot et al.,
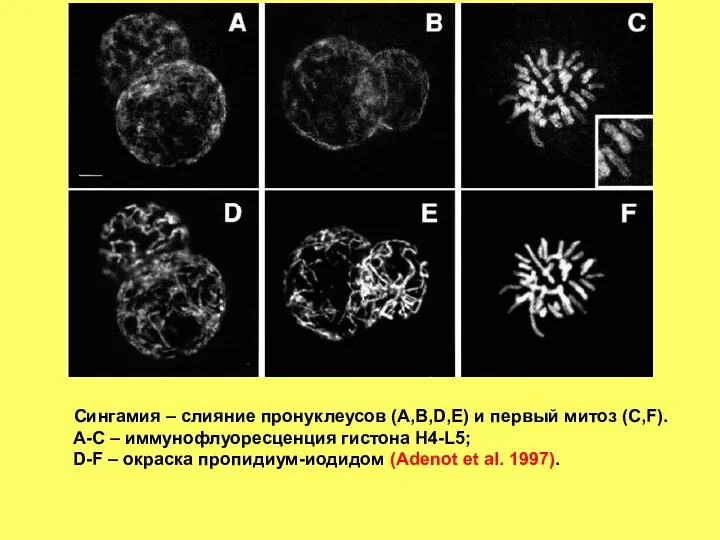
Сингамия – слияние пронуклеусов (A,B,D,E) и первый митоз (C,F).
A-C
Сингамия – слияние пронуклеусов (A,B,D,E) и первый митоз (C,F).
A-C
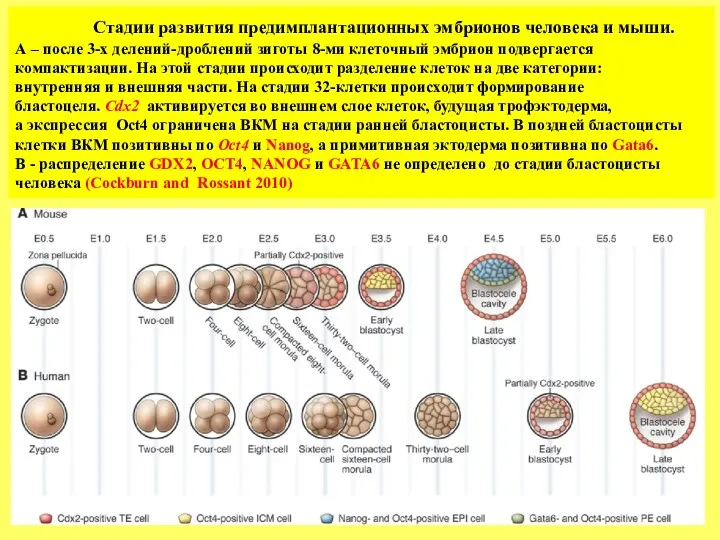
Стадии развития предимплантационных эмбрионов человека и мыши.
А – после 3-х
Стадии развития предимплантационных эмбрионов человека и мыши.
А – после 3-х
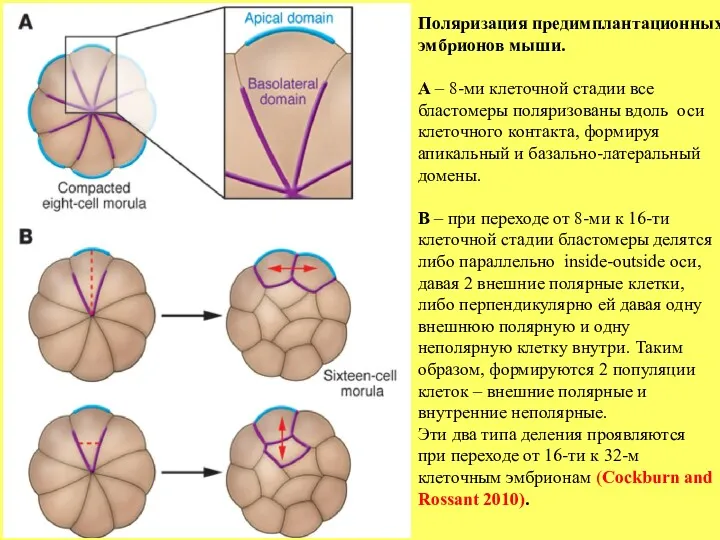
Поляризация предимплантационных
эмбрионов мыши.
А – 8-ми клеточной стадии все
бластомеры поляризованы вдоль
Поляризация предимплантационных
эмбрионов мыши.
А – 8-ми клеточной стадии все
бластомеры поляризованы вдоль

Важнейшие события с момента оплодотворения:
Синтез белка происходит на материнских мРНК;
Зиготические
Важнейшие события с момента оплодотворения:
Синтез белка происходит на материнских мРНК;
Зиготические
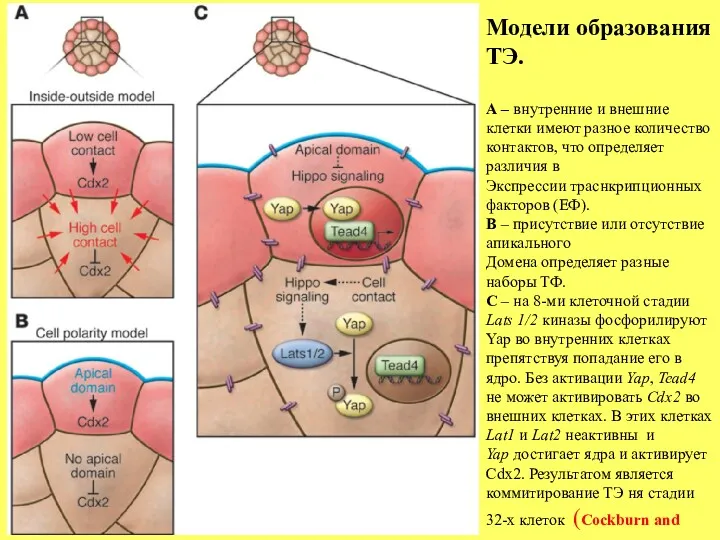
Модели образования ТЭ.
А – внутренние и внешние
клетки имеют разное количество
Модели образования ТЭ.
А – внутренние и внешние
клетки имеют разное количество

Коммитирование апикальных клеток к дифференцировке в ТЭ, первое реальное событие дифференцировки
Коммитирование апикальных клеток к дифференцировке в ТЭ, первое реальное событие дифференцировки
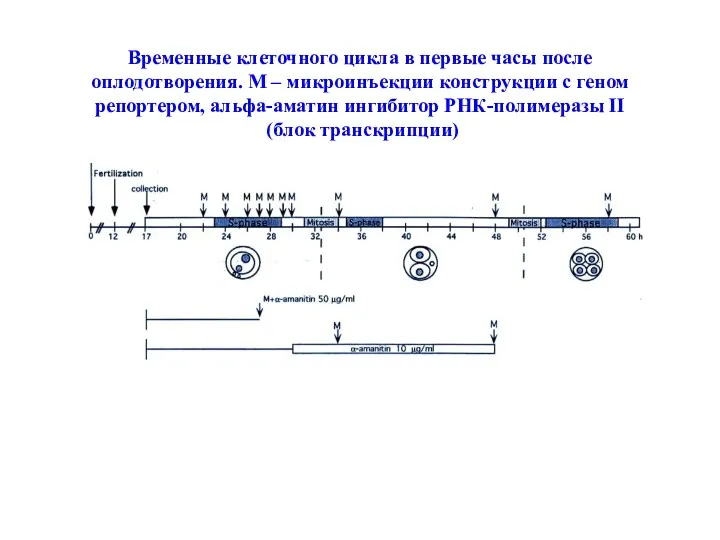
Временные клеточного цикла в первые часы после оплодотворения. М – микроинъекции
Временные клеточного цикла в первые часы после оплодотворения. М – микроинъекции
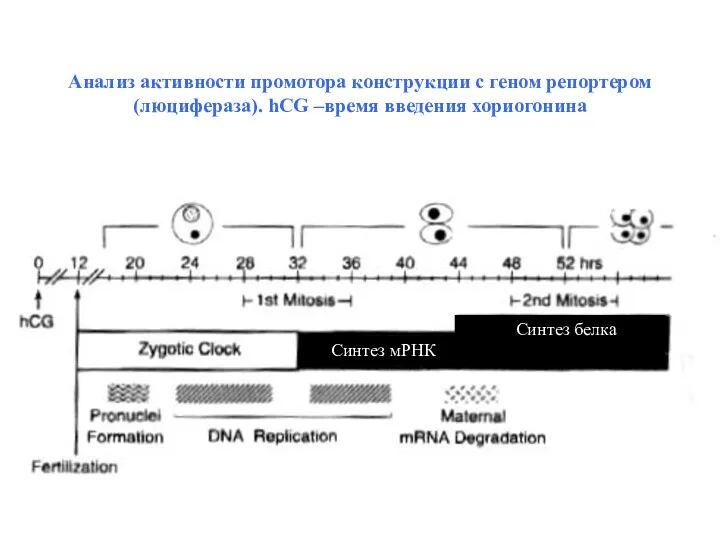
Анализ активности промотора конструкции с геном репортером (люцифераза). hCG –время введения
Анализ активности промотора конструкции с геном репортером (люцифераза). hCG –время введения
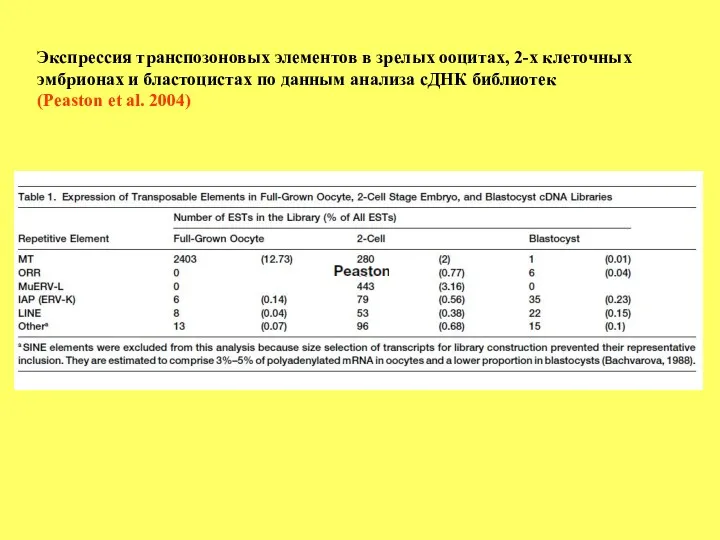
Экспрессия транспозоновых элементов в зрелых ооцитах, 2-х клеточных
эмбрионах и бластоцистах
Экспрессия транспозоновых элементов в зрелых ооцитах, 2-х клеточных
эмбрионах и бластоцистах

Экспрессия ретротранспозонов на стадии зиготы и 2-хклеточной стадии
В
Экспрессия ретротранспозонов на стадии зиготы и 2-хклеточной стадии
В
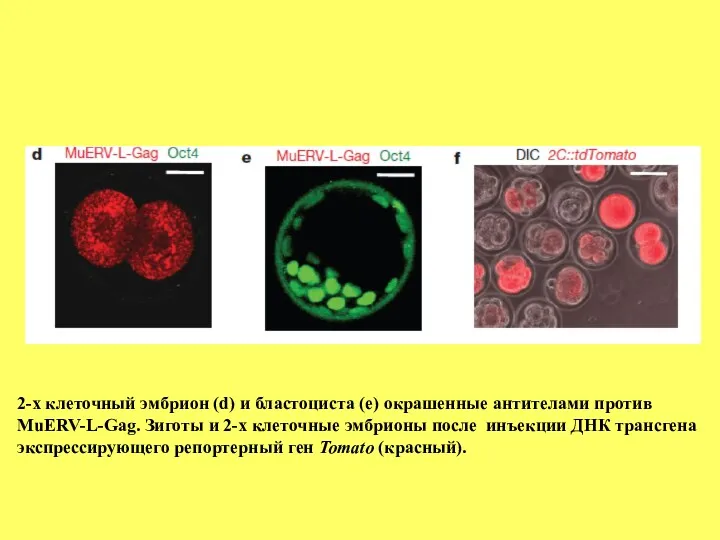
2-х клеточный эмбрион (d) и бластоциста (е) окрашенные антителами против
MuERV-L-Gag.
2-х клеточный эмбрион (d) и бластоциста (е) окрашенные антителами против
MuERV-L-Gag.

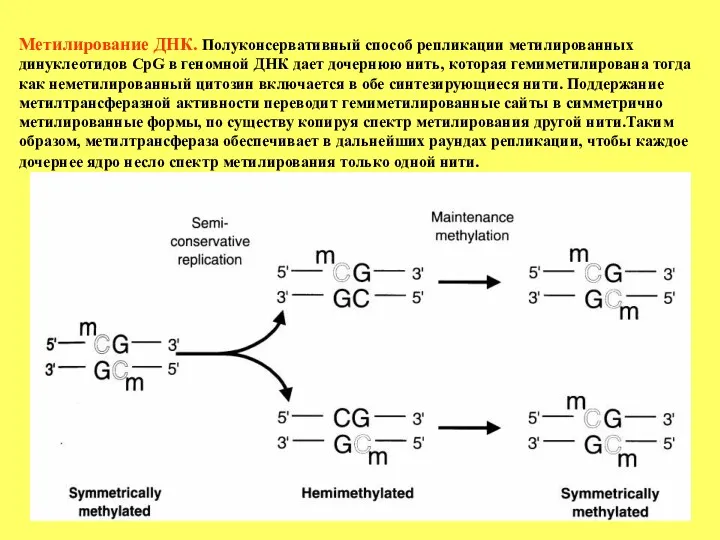
Метилирование ДНК. Полуконсервативный способ репликации метилированных динуклеотидов CpG в геномной ДНК
Метилирование ДНК. Полуконсервативный способ репликации метилированных динуклеотидов CpG в геномной ДНК
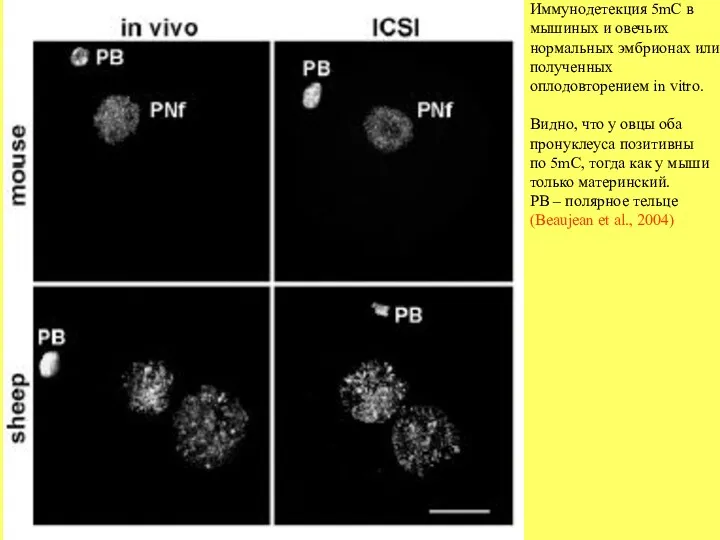
Иммунодетекция 5mC в
мышиных и овечьих
нормальных эмбрионах или
полученных
оплодовторением in vitro.
Видно, что
Иммунодетекция 5mC в
мышиных и овечьих
нормальных эмбрионах или
полученных
оплодовторением in vitro.
Видно, что
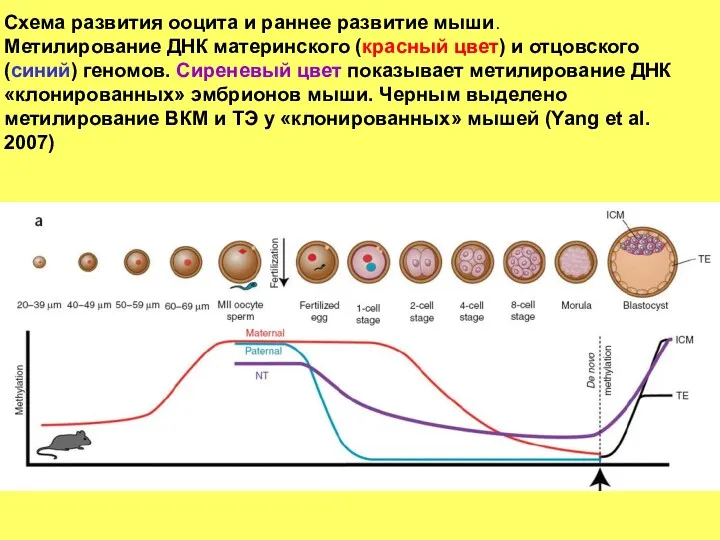
Схема развития ооцита и раннее развитие мыши.
Метилирование ДНК материнского (красный
Схема развития ооцита и раннее развитие мыши.
Метилирование ДНК материнского (красный
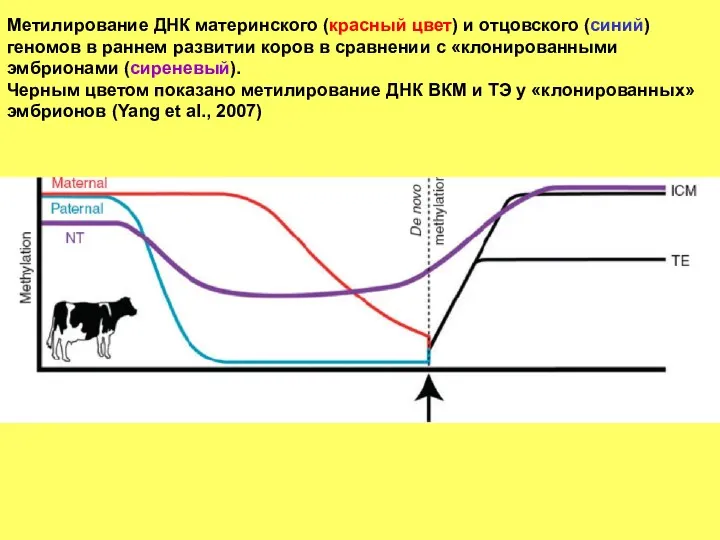
Метилирование ДНК материнского (красный цвет) и отцовского (синий) геномов в раннем
Метилирование ДНК материнского (красный цвет) и отцовского (синий) геномов в раннем
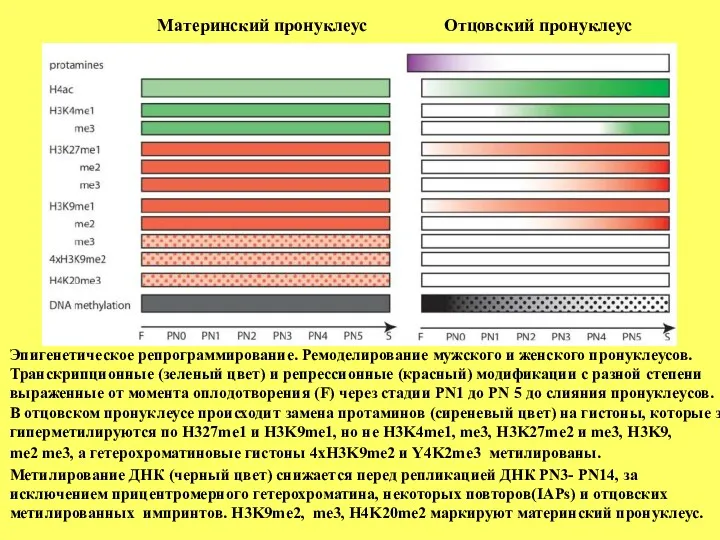
Эпигенетическое репрограммирование. Ремоделирование мужского и женского пронуклеусов. Транскрипционные (зеленый цвет) и
Эпигенетическое репрограммирование. Ремоделирование мужского и женского пронуклеусов. Транскрипционные (зеленый цвет) и
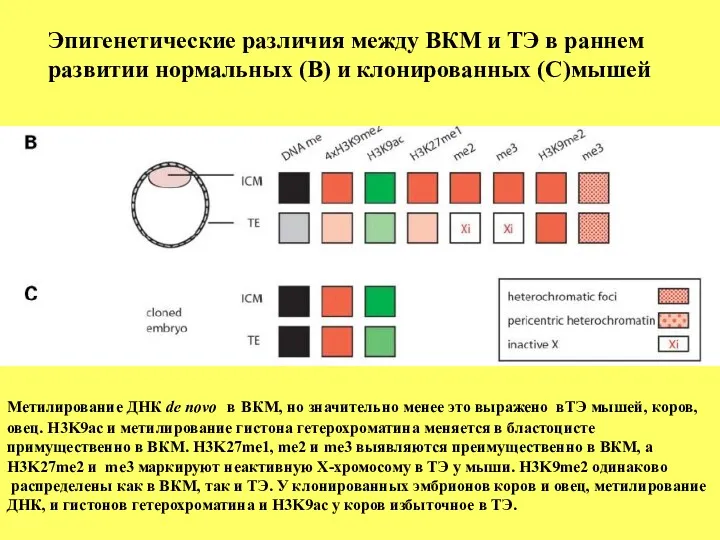
Метилирование ДНК de novo в ВКМ, но значительно менее это выражено
Метилирование ДНК de novo в ВКМ, но значительно менее это выражено

 Human genetics
Human genetics Соседи по планете живущие в океанских глубинах
Соседи по планете живущие в океанских глубинах Этапы развития жизни на Земле
Этапы развития жизни на Земле Семейство Розоцветные
Семейство Розоцветные Сохранение видов лекарственных растений моей малой Родины. Тамбовская область
Сохранение видов лекарственных растений моей малой Родины. Тамбовская область Шкіра має 3 прошарки
Шкіра має 3 прошарки Leaf as a lateral organ of stalk. Morphology and anatomy. Metamorphosis of leaf
Leaf as a lateral organ of stalk. Morphology and anatomy. Metamorphosis of leaf Проект экологической игры Моя планета - человеческий дом
Проект экологической игры Моя планета - человеческий дом Технология возделывания чечевицы
Технология возделывания чечевицы мхи
мхи Бактерии. Грибы. Общая характеристика грибов
Бактерии. Грибы. Общая характеристика грибов Восприятие цвета глазом. Аддитивное и субтрактивное смешение цветов
Восприятие цвета глазом. Аддитивное и субтрактивное смешение цветов Примитивные черты организации типа Плоские черви класса сосальщики
Примитивные черты организации типа Плоские черви класса сосальщики Презентация Хартия Земли
Презентация Хартия Земли Катаболизм. Энергетический обмен и всё, что с ним связано
Катаболизм. Энергетический обмен и всё, что с ним связано Невидимый мир, работа с микроскопом
Невидимый мир, работа с микроскопом Санитарная микробиология
Санитарная микробиология Санитарно-показательные микроорганизмы
Санитарно-показательные микроорганизмы Бұлшық ет құрылысы
Бұлшық ет құрылысы Тура қанаттылар отряды. Шегіртке тұқымдасы
Тура қанаттылар отряды. Шегіртке тұқымдасы Анатомия и физиология мужской половой системы
Анатомия и физиология мужской половой системы Органи розмноження
Органи розмноження Бактерії - найменші одноклітинні організми. Будова, поширення, розмноження бактерій
Бактерії - найменші одноклітинні організми. Будова, поширення, розмноження бактерій Продление рода. Органы размножения. 7 класс
Продление рода. Органы размножения. 7 класс Исследование экологической среды жилого помещения
Исследование экологической среды жилого помещения Роль бактерий в природе и жизни человека
Роль бактерий в природе и жизни человека Нужны ли минеральные соли животным и человеку
Нужны ли минеральные соли животным и человеку Биотехнология. Генетические векторы для клонирования ДНК
Биотехнология. Генетические векторы для клонирования ДНК