- Основные стратегии регуляции метаболических путей

Содержание
- 2. Живые системы включают в свой состав все хими-ческие элементы, которые находятся в окружающей его среде. Наибольшая
- 3. Живые системы – открытые системы, способные к саморегуляции, самоорганизации и самовоспроизве-дению. Они обладают, в частности, свойством
- 4. Схема внутриклеточной регуляции действия ферментов
- 5. I стратегия: Быстрое изменение каталитической активности ключевых (регуляторных) ферментов под влиянием аллостерических регуляторов. Ключевые (регуляторные) ферменты
- 6. В ферментативном катализе принципиально важен этап образования комплекса фермент-субстрат (ES): E + S ?? ES ?
- 7. Vmax1 VmaxK Vmax2 Km1 KmK Km2 Влияние аллостерических регуляторов на Vmax In vivo [S] почти всегда
- 8. II стратегия: А. Ковалентная модификация фермен-тов путём фосфорилирования – дефосфорилирова-ния. Эта модификация обратима. Действует параллельно с
- 9. Серин, треонин и тирозин, как мишени для протеинкиназ
- 10. Протеинкиназы (ПК) классифицируют по остаткам фосфорилируемых ими аминокислот Тирозиновые ПК: 1. Цитоплазматические тиро-зиновые ПК: (Src) –
- 11. Дефосфорилирование молекулы фермента или другого белка происходит с участием протеинфосфа-таз. Протеинфосфатазы, PP (protein phosphatase) Тирозин-специфичные РР:
- 12. II стратегия: Б. Нековалентная модификация фермен-тов. Реализуется путем ограниченного (лимитирован-ного) протеолиза. Как правило, носит каскадный харак-тер
- 13. Каскадный механизм, приводящий к свертыванию крови Последовательность из 5 ферментов, в которой каждый фермент активирует следующий
- 14. Внешний и внутренний пути инициирования апоптоза. Конечный эффект – активация исполнительной каспазы-3 благодаря работе каскада инициирующих
- 15. Каспазы (англ. caspase + cysteine-dependent aspar-tate specific protease) — семейство внутриклеточных цистеиновых протеаз, расщепляющих пептидные связи
- 16. Общая схема «созревания» / активации каспаз Зимоген каспазы α β α β Catalytical dyad = каталитическая
- 17. Очевидно, что I и II стратегии регуляции обеспечивают не только быстрый, но и очень точный механизм
- 18. III стратегия: Изменение количества фермента – уси-ление его биосинтеза, либо разрушение уже имеющих-ся молекул фермента. Это
- 19. Б. Ферменты индуцируемые – скорость их синтеза изменяется в зависимости от условий существования организма. Регуляция синтеза
- 20. Синтез индуцируемых ферментов - одно из проявле-ний биохимической адаптации метаболизма клетки к изменившимся условиям существования. Итог:
- 21. IV стратегия: Компартментализация ферментов и метаболических путей. Пространственное разделение метаболических путей позволяет согласованно и одновременно протекать
- 22. Основные компартменты клетки: Плазматическая мембрана Ядро Цитоскелет Митохондрии: внутренняя мембрана и матрикс ЭПР (микросомы): мембрана и
- 23. Существуют ферменты (киназы), которые при переходе из неактивной в активную форму изменяют своё местонахождение (компартмен-тализацию) 1.
- 24. 2. Протеинкиназа С (ПКС) – серин-треониновая ПК. В неактивной конформации ПК находится в цитоплазме. Многочисленные лиганды
- 25. V стратегия: Гормональная (эндокринная) регуляция. Под действием гормонов (первичных мессендже-ров) внутри клетки синтезируются вторичные мессен-джеры, которые
- 26. Каждый тип клетки содержит специфическую комби- нацию различных рецепторов, что даёт ей возмож- ность по разному
- 27. Мультиферментные комплексы (как пример нековалентной модификации ферментов) При работе мультиферментного комплекса: продукт первого фермента в составе
- 28. Модель пируватдегидрогеназного комплекса Е1 – пируват дегидрогеназа (кофактор: тиаминпиро- фосфат) Е2 – дигидролипоил трансацетилаза (кофакторы: ли-
- 29. Изоферменты (изоэнзимы) Изоферменты – молекулярные формы (изотипы) одного фермента. Изоферменты отличаются по их первичной струк-туре, что
- 30. ЛДГ существует в виде 5 изоферментов. Все они являются вариантами комбинации в различных соотношениях двух типов
- 31. В зависимости от степени подвижности при электро-форезе в крахмальном геле, изоферменты ЛДГ нумеруют: ЛДГ-1, обладающий наибольшей
- 32. Биологический смысл существования изоферментов: В разных тканях существует разная концентрация субстрата для одного и того же
- 33. Электрофоретическая оценка содержания изофер-ментов ЛДГ в сыворотке крови в норме и на различ-ных сроках от начала
- 35. Скачать презентацию

Живые системы включают в свой состав все хими-ческие элементы, которые
Живые системы включают в свой состав все хими-ческие элементы, которые

Живые системы – открытые системы, способные к саморегуляции, самоорганизации и
Живые системы – открытые системы, способные к саморегуляции, самоорганизации и
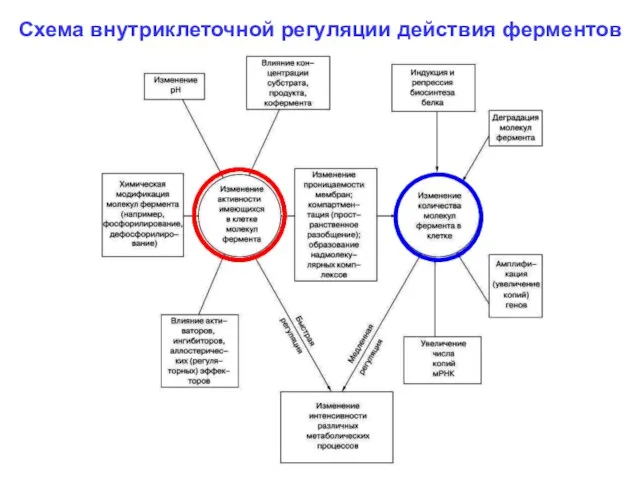
Схема внутриклеточной регуляции действия ферментов
Схема внутриклеточной регуляции действия ферментов

I стратегия: Быстрое изменение каталитической активности ключевых (регуляторных) ферментов под
I стратегия: Быстрое изменение каталитической активности ключевых (регуляторных) ферментов под

В ферментативном катализе принципиально важен
этап образования комплекса фермент-субстрат (ES):
В ферментативном катализе принципиально важен
этап образования комплекса фермент-субстрат (ES):
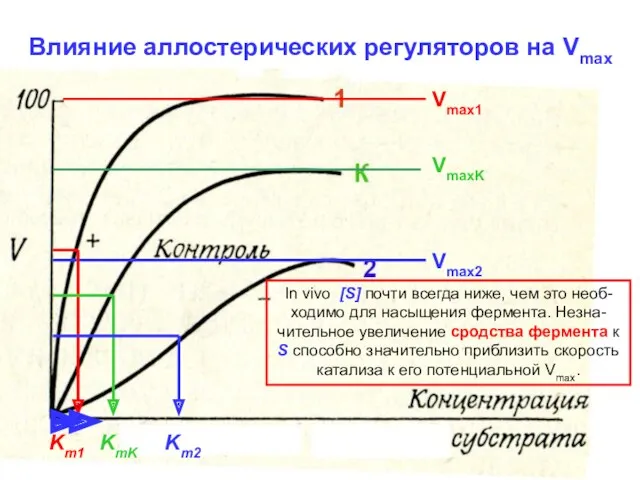
Vmax1
VmaxK
Vmax2
Km1
KmK
Km2
Влияние аллостерических регуляторов на Vmax
In vivo [S] почти всегда ниже, чем
Vmax1
VmaxK
Vmax2
Km1
KmK
Km2
Влияние аллостерических регуляторов на Vmax
In vivo [S] почти всегда ниже, чем

II стратегия: А. Ковалентная модификация фермен-тов путём фосфорилирования – дефосфорилирова-ния.
II стратегия: А. Ковалентная модификация фермен-тов путём фосфорилирования – дефосфорилирова-ния.
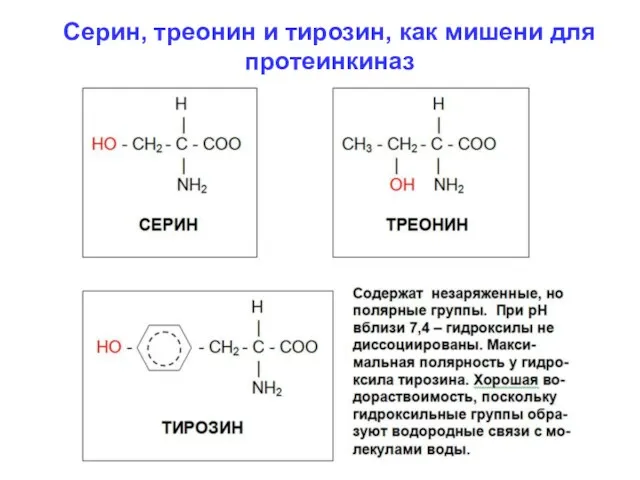
Серин, треонин и тирозин, как мишени для
протеинкиназ
Серин, треонин и тирозин, как мишени для
протеинкиназ
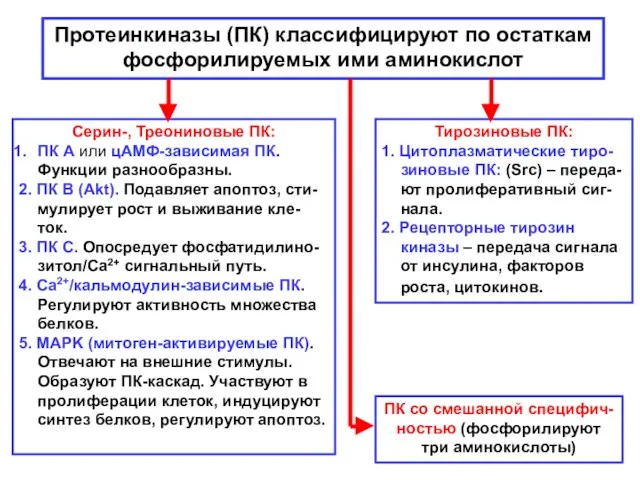
Протеинкиназы (ПК) классифицируют по остаткам
фосфорилируемых ими аминокислот
Тирозиновые ПК:
1. Цитоплазматические
Протеинкиназы (ПК) классифицируют по остаткам
фосфорилируемых ими аминокислот
Тирозиновые ПК:
1. Цитоплазматические
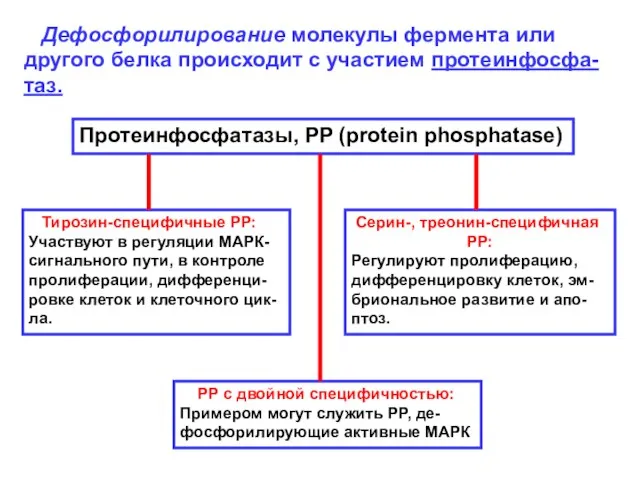
Дефосфорилирование молекулы фермента или другого белка происходит с участием протеинфосфа-таз.
Протеинфосфатазы,
Дефосфорилирование молекулы фермента или другого белка происходит с участием протеинфосфа-таз.
Протеинфосфатазы,

II стратегия: Б. Нековалентная модификация фермен-тов. Реализуется путем ограниченного (лимитирован-ного)
II стратегия: Б. Нековалентная модификация фермен-тов. Реализуется путем ограниченного (лимитирован-ного)
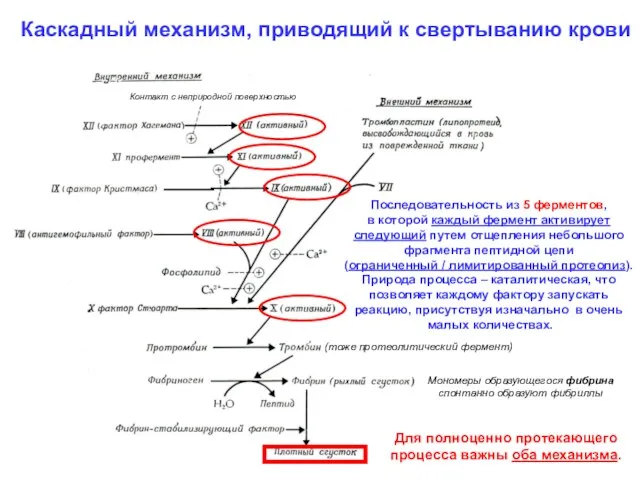
Каскадный механизм, приводящий к свертыванию крови
Последовательность из 5 ферментов,
в которой
Каскадный механизм, приводящий к свертыванию крови
Последовательность из 5 ферментов,
в которой
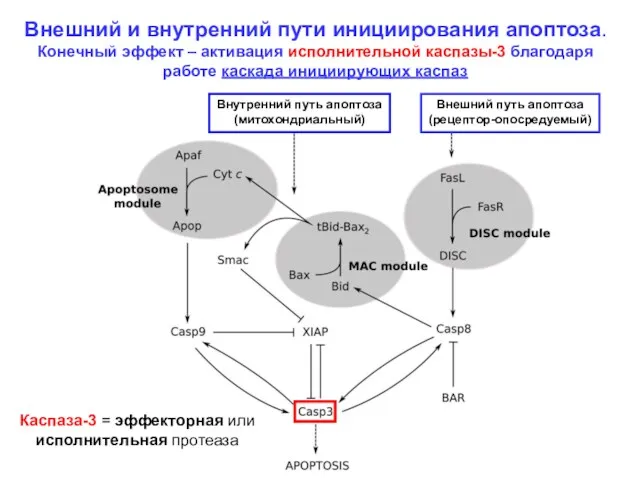
Внешний и внутренний пути инициирования апоптоза.
Конечный эффект – активация исполнительной каспазы-3
Внешний и внутренний пути инициирования апоптоза.
Конечный эффект – активация исполнительной каспазы-3

Каспазы (англ. caspase + cysteine-dependent aspar-tate specific protease) — семейство внутриклеточных
Каспазы (англ. caspase + cysteine-dependent aspar-tate specific protease) — семейство внутриклеточных
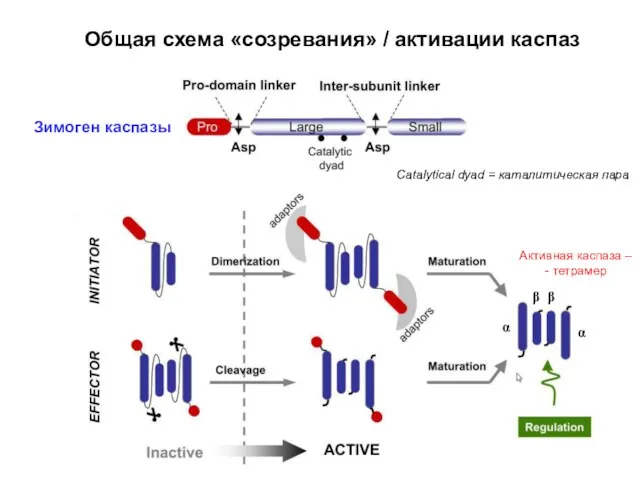
Общая схема «созревания» / активации каспаз
Зимоген каспазы
α
β
α
β
Catalytical dyad = каталитическая пара
Активная
Общая схема «созревания» / активации каспаз
Зимоген каспазы
α
β
α
β
Catalytical dyad = каталитическая пара
Активная

Очевидно, что I и II стратегии регуляции
обеспечивают не только быстрый,
Очевидно, что I и II стратегии регуляции
обеспечивают не только быстрый,

III стратегия: Изменение количества фермента – уси-ление его биосинтеза, либо
III стратегия: Изменение количества фермента – уси-ление его биосинтеза, либо

Б. Ферменты индуцируемые – скорость их синтеза
изменяется в зависимости от условий существования
организма.
Регуляция синтеза происходит на генетическом
уровне под действием индукторов (соответствующие
субстраты или метаболиты).
Ген «выключен», пока с
Б. Ферменты индуцируемые – скорость их синтеза
изменяется в зависимости от условий существования
организма.
Регуляция синтеза происходит на генетическом
уровне под действием индукторов (соответствующие
субстраты или метаболиты).
Ген «выключен», пока с

Синтез индуцируемых ферментов - одно из проявле-ний биохимической адаптации метаболизма клетки к
изменившимся условиям существования.
Итог: либо увеличивается количество уже имеюще-гося фермента, что обеспечивает более быстрое
протекание определённой реакции, либо вырабатыва-ются новые ферменты, ранее отсутствовавшие в
клетке
Синтез индуцируемых ферментов - одно из проявле-ний биохимической адаптации метаболизма клетки к
изменившимся условиям существования.
Итог: либо увеличивается количество уже имеюще-гося фермента, что обеспечивает более быстрое
протекание определённой реакции, либо вырабатыва-ются новые ферменты, ранее отсутствовавшие в
клетке

IV стратегия: Компартментализация ферментов и
метаболических путей. Пространственное разделение метаболических
IV стратегия: Компартментализация ферментов и
метаболических путей. Пространственное разделение метаболических

Основные компартменты клетки:
Плазматическая мембрана
Ядро
Цитоскелет
Митохондрии: внутренняя мембрана и матрикс
ЭПР (микросомы): мембрана и
Основные компартменты клетки:
Плазматическая мембрана
Ядро
Цитоскелет
Митохондрии: внутренняя мембрана и матрикс
ЭПР (микросомы): мембрана и

Существуют ферменты (киназы), которые при
переходе из неактивной в активную форму
Существуют ферменты (киназы), которые при
переходе из неактивной в активную форму

2. Протеинкиназа С (ПКС) – серин-треониновая ПК. В неактивной конформации
2. Протеинкиназа С (ПКС) – серин-треониновая ПК. В неактивной конформации

V стратегия: Гормональная (эндокринная) регуляция.
Под действием гормонов (первичных
V стратегия: Гормональная (эндокринная) регуляция.
Под действием гормонов (первичных

Каждый тип клетки содержит специфическую комби-
нацию различных рецепторов, что даёт
Каждый тип клетки содержит специфическую комби-
нацию различных рецепторов, что даёт

Мультиферментные комплексы
(как пример нековалентной модификации ферментов)
При работе мультиферментного комплекса: продукт
Мультиферментные комплексы
(как пример нековалентной модификации ферментов)
При работе мультиферментного комплекса: продукт
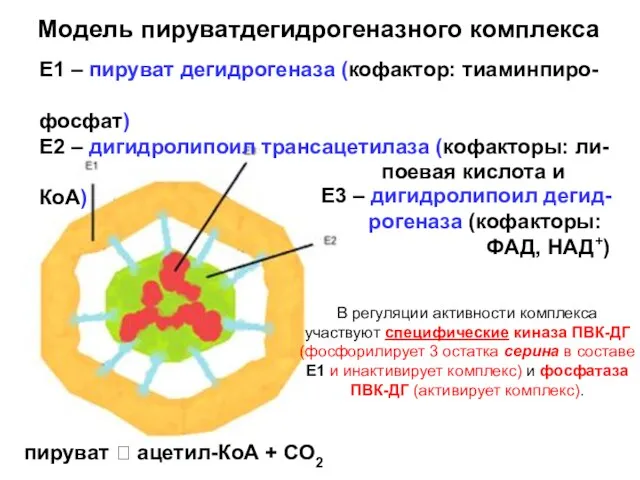
Модель пируватдегидрогеназного комплекса
Е1 – пируват дегидрогеназа (кофактор: тиаминпиро-
фосфат)
Е2 – дигидролипоил
Модель пируватдегидрогеназного комплекса
Е1 – пируват дегидрогеназа (кофактор: тиаминпиро-
фосфат)
Е2 – дигидролипоил

Изоферменты (изоэнзимы)
Изоферменты – молекулярные формы (изотипы) одного фермента.
Изоферменты
Изоферменты (изоэнзимы)
Изоферменты – молекулярные формы (изотипы) одного фермента.
Изоферменты
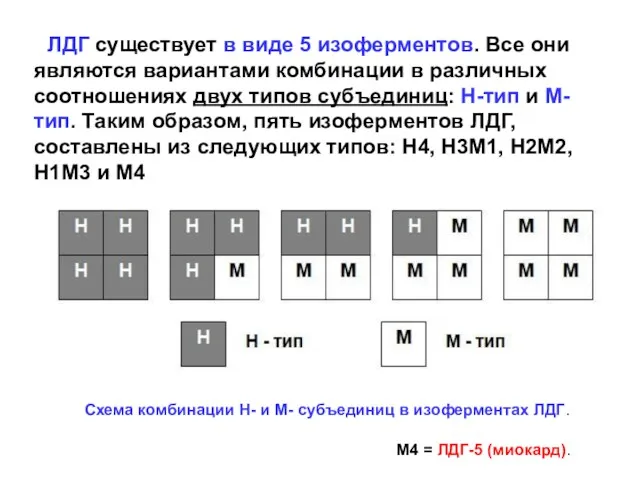
ЛДГ существует в виде 5 изоферментов. Все они являются вариантами
ЛДГ существует в виде 5 изоферментов. Все они являются вариантами

В зависимости от степени подвижности при электро-форезе в крахмальном геле,
В зависимости от степени подвижности при электро-форезе в крахмальном геле,

Биологический смысл существования изоферментов:
В разных тканях существует разная концентрация субстрата
Биологический смысл существования изоферментов:
В разных тканях существует разная концентрация субстрата
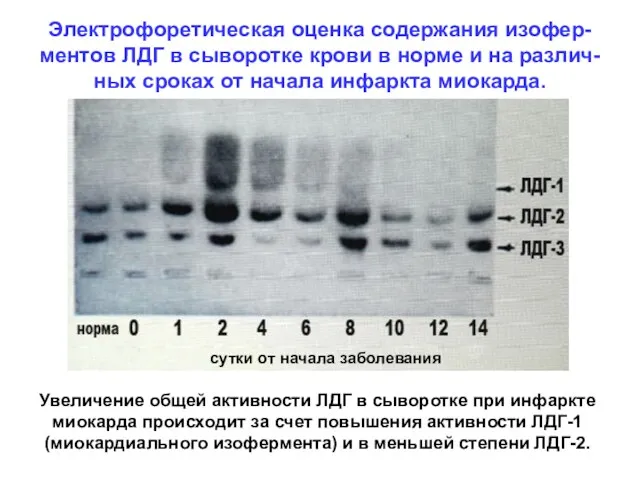
Электрофоретическая оценка содержания изофер-ментов ЛДГ в сыворотке крови в норме и
Электрофоретическая оценка содержания изофер-ментов ЛДГ в сыворотке крови в норме и
 Сенсорні функції організму
Сенсорні функції організму Сорта огурцов
Сорта огурцов Простейшие и кишечнополостные организмы
Простейшие и кишечнополостные организмы Видоизменения побегов
Видоизменения побегов Лес и его голоса
Лес и его голоса Клетка. 10 класс
Клетка. 10 класс Всасывание. Роль печени. Функции толстого кишечника
Всасывание. Роль печени. Функции толстого кишечника Систематика растений
Систематика растений Характеристика основних типів ієрархії у тварин
Характеристика основних типів ієрархії у тварин Царство Растения
Царство Растения Палеонтологическая консультация для подготовки к открытой Березниковский олимпиаде по геологии
Палеонтологическая консультация для подготовки к открытой Березниковский олимпиаде по геологии Биология как наука, её история. Методы исследования в биологии
Биология как наука, её история. Методы исследования в биологии Особенности питания пожилых людей
Особенности питания пожилых людей Бионика
Бионика Гаметаларды in vivo культивирлеу
Гаметаларды in vivo культивирлеу Тип Хордовые
Тип Хордовые Здоровая пища-залог здоровья
Здоровая пища-залог здоровья Транспорт веществ через биологические мембраны
Транспорт веществ через биологические мембраны Породы уток и гусей
Породы уток и гусей Посттрансляционные модификации белков
Посттрансляционные модификации белков Многообразие водных биогеоценозов
Многообразие водных биогеоценозов Мышечная система. Кафедра анатомии человека
Мышечная система. Кафедра анатомии человека Мидии. Места обитания
Мидии. Места обитания Солнце, растение и мы с вами
Солнце, растение и мы с вами Строение и развитие женских половых клеток. Лекция 2
Строение и развитие женских половых клеток. Лекция 2 Обмін речовин та перетворення енергії в організмі людини – основна властивість живого
Обмін речовин та перетворення енергії в організмі людини – основна властивість живого Growth and Development
Growth and Development Объекты биотехнологии. Биообъекты, применяемые на практике
Объекты биотехнологии. Биообъекты, применяемые на практике