- Построение и анализ молекулярных сетей. Лекция 4

Содержание
- 2. Сложная биологическая система может быть представлена в виде сети взаимодействующих объектов: организмы, клетки, белки, гены, РНК,
- 3. Сигнальные сети Основная функция – распознавание и передача сигналов с рецепторов в ядро или к другим
- 4. Сигнальные пути Активация рецепторов запускает сигнальные каскады, что приводит к изменению транскрипции генов и ответу клетки
- 5. Адренергический сигнальный путь в кардиомиоцитах Стимуляция бета-адренорецепторов активирует киназы, которые фосфорилируют транспортеры и ионные каналы, что
- 6. Свойства сигнальных сетей Большое количество перекрестов между сигнальными путями. Один и тот же белок участвует в
- 7. Метаболические пути Метаболический путь – последовательность химических реакций, осуществляющих синтез продукта (или нескольких продуктов) из исходных
- 8. Биосинтез стероидов
- 9. Реконструкция метаболической сети клетки
- 10. Базы данных по сигнальным и метаболическим путям
- 11. KEGG (Kyoto Encyclopedia of Genes and Genomes) 321 сигнальных, регуляторных, метаболических путей для разных видов животных
- 12. REACTOME 1886 сигнальных и метаболических пути человека, организованные иерархически: от отдельных реакций до крупных путей. Позволяет
- 13. SIGNOR
- 14. Сети белок-белковых взаимодействий (ББВ) Вершины данной сети представляют собой белки, а ребра показывают наличие прямых, физических
- 15. Подходы к оценке белок-белковых взаимодействий Ко-иммунопреципитация Аффинная хроматография Перекресная сшивка при помощи реактивных соединений или УФ
- 16. Базы данных по белок-белковым взаимодействиям
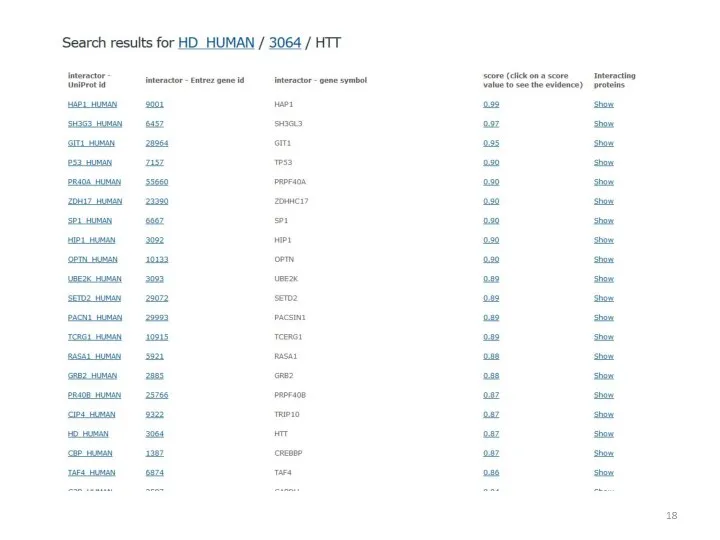
- 17. HIPPIE Содержит информацию о более чем 18000 белках человека и 400000 взаимодействий. Каждому взаимодействию присвоена оценка,
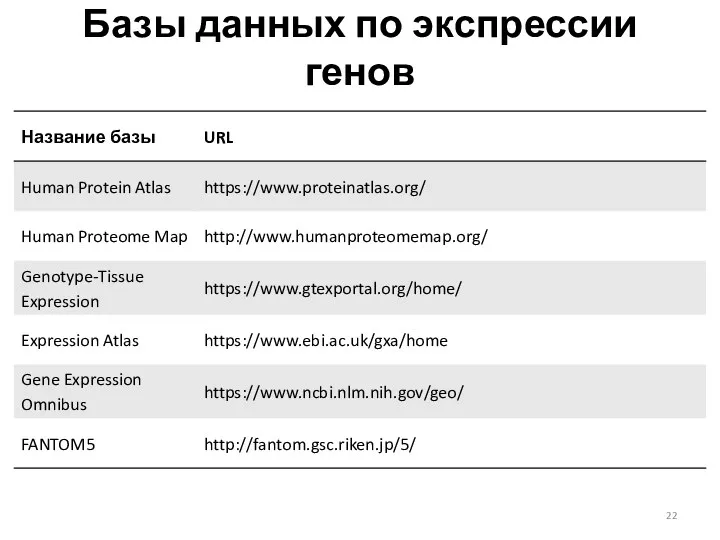
- 22. Базы данных по экспрессии генов
- 23. Сети ко-экспрессии Разные условия, разные эксперименты Для каждой пары генов Коэффициент корреляции Пирсона Ранговый коэффициент корреляции
- 25. Сети ко-экспрессии Определяется порог коэффициента корреляции r. Если r выше порога, то два гена соединяются ребром,
- 26. COXPRESdb
- 27. Результаты поиска для гена ADRB1 в COXPRESdb
- 28. Кластерный анализ в COXPRESdb
- 29. Сети на основе интеграции информации Сигнальные сети построены на основе интеграции данных о белок-белковых взаимодействиях, регуляции
- 30. Примеры средств для визуализации и анализа сетей
- 31. Cytoscape (http://www.cytoscape.org/)
- 32. Cytoscape Платформа с открытым исходным кодом для визуализации и анализа биологических сетей. Возможность использования дополнительных данных,
- 33. Анализ топологии сетей
- 34. Связные компоненты 7 связных компонент
- 35. Путь в сети (графе) Путь – это последовательность вершин {x1, x2, …, xn} такая, что (x1,
- 36. Кратчайший путь между вершинами
- 37. Характеристики сети – степень вершины Степень вершины (k) – число взаимодействий с другими вершинами. Для направленных
- 38. Распределение степени Распределение имеет вид степенной зависимости P(k) ~ k-γ Большинство вершин сети имеет небольшое число
- 39. Свойства белков-«хабов» Белки с большим числом взаимодействий называются белками- «хабами». Многие «хабы» являются наиболее важными, ключевыми
- 40. Свойства белков-«хабов» Белки-«хабы» чаще всего не взаимодействуют непосредственно друг с другом Белки-«хабы» более консервативны у разных
- 41. Мотивы и графлеты Мотивами называются цепочки или контуры из 3-6 вершин в направленной сети (например в
- 42. Модули в биологических сетях Структура биологических сетей неоднородна. Сети содержат модули, состоящие из вершин, которые имею
- 43. Модули в биологических сетях Межмодулярные хабы – «хабы», которые соединяют два соседних модуля. Мосты (bridge) –
- 44. Модули в биологических сетях Межмодулярные «хабы» представляют собой временные «хабы», для которых количество и состав взаимодействий
- 45. Предсказание маркеров заболеваний Белки (гены), связанные с определенным заболеванием, обладают тенденцией формировать один или несколько перекрывающихся
- 46. Сеть белок-белковых взаимодействий Белки с известной связью с заболеванием Белки-кандидаты
- 47. Методы оценки связи белков (генов) с заболеванием Метод ближайшего соседа n – число взаимодействий с белками
- 48. Случайное блуждание с возвратом Методы оценки связи белков (генов) с заболеванием Вычисляется вероятность p∞ оказаться в
- 49. Оценка связи белка (гена) с заболеванием Широкомасштабные генетические исследования. Оценка корреляции полиморфизма гена с наличием/отсутствием заболевания.
- 50. Базы данных по связи белков/генов с заболеваниями
- 51. DisGeNET Включает информацию, интегрированную из многих источников, в том числе из литературы. Эту информацию можно представить
- 52. Результаты поиска для ADRB1 Score отражает достоверность связи между геном и заболеванием. Наиболее достоверна связь с
- 53. Связь ADRB1 с гипертензией Указан источник данных. Показаны предложения из Abstarcts PubMed, в которых выделены ключевые
- 54. «Мастер-регуляторы» в сети «Мастер-регулятор» – ключевой белок, ответственный за изменение экспрессии генов. Гиперэкспрессированные гены Гипоэкспрессированные гены
- 55. Этапы поиска «мастер-регуляторов» Идентификация дифференциально экспрессирующихся генов (норма-патология, лекарство-растворитель и др.); Поиск экспериментальной информации о транскрипционных
- 56. Моделирование динамики сетей
- 57. Моделирование динамики сетей Моделирование изменения концентраций субстратов/продуктов реакций во времени. В сигнальной сети изменение концентраций активных
- 58. Моделирование динамики сетей Дифференциальные уравнения Логические сети Сети Петри Клеточный автомат Гибридные методы
- 59. Дифференциальные уравнения Система дифференциальных уравнений, описывающих изменение концентраций РНК и белков во времени: Xi – концентрация
- 61. Численные методы решения. Метод Эйлера Начальное состояние:
- 62. Логические сети Простейший вариант – булевы сети Каждая молекула (ген или белок) активна (1) или неактивна
- 63. Модель апоптоза у дрожжей Сеть регуляции апоптоза
- 64. Для каждого белка записываются булевы функции Плюсом обозначено логическое «И».
- 65. Различные внешние воздействия приводят к индукции апоптоза
- 66. Сети Петри Впервые описаны Карлом Петри в 1962 году. Сеть Петри – это граф с двумя
- 67. Примеры описания взаимодействий A+B?C C?A+B A+B?C (кат. ферментом E) A+B?C (ингибитор) A B C A B
- 68. Расширение сетей Петри Временная сеть Петри - такая сеть, где переходы обладают весом, определяющим продолжительность срабатывания
- 69. Клеточный автомат Состоит из регулярной решетки ячеек, каждая из которых может находится в одном из конечного
- 71. Моделирование апоптоза, индуцированного FASL FLIP и IAP – ингибиторы апоптоза. А siRNA и SMAC, в свою
- 72. Описание взаимодействий
- 74. Скачать презентацию

Сложная биологическая система может быть представлена в виде сети взаимодействующих объектов:
Сложная биологическая система может быть представлена в виде сети взаимодействующих объектов:

Сигнальные сети
Основная функция – распознавание и передача сигналов с рецепторов в
Сигнальные сети
Основная функция – распознавание и передача сигналов с рецепторов в
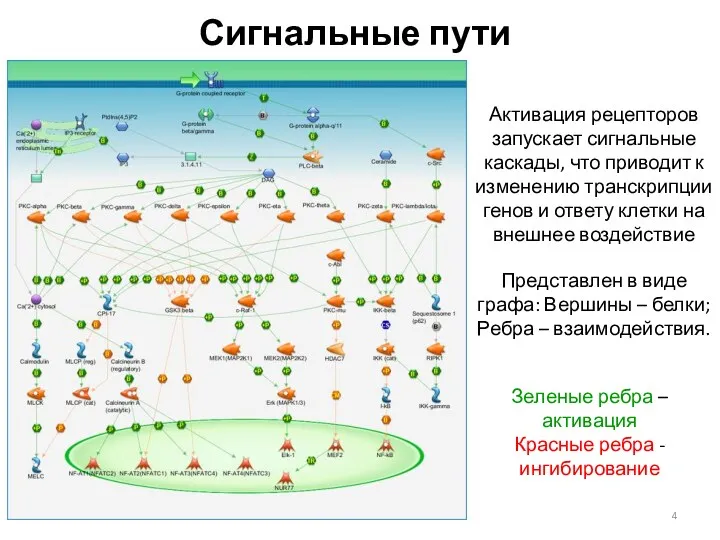
Сигнальные пути
Активация рецепторов запускает сигнальные каскады, что приводит к изменению транскрипции
Сигнальные пути
Активация рецепторов запускает сигнальные каскады, что приводит к изменению транскрипции
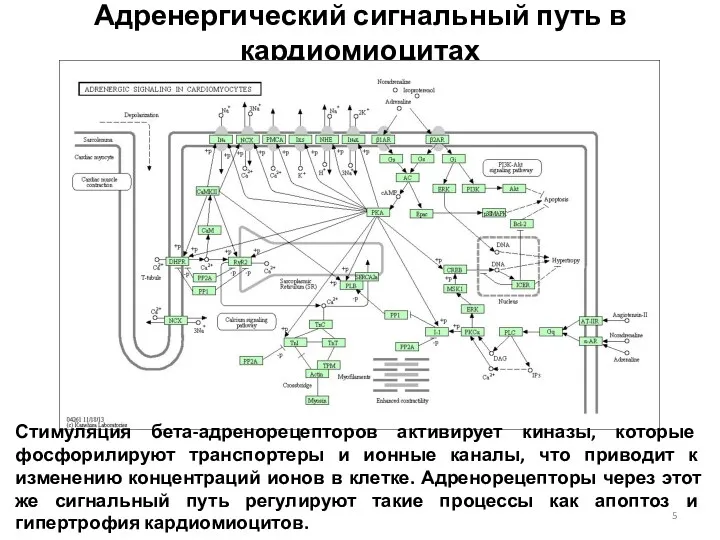
Адренергический сигнальный путь в кардиомиоцитах
Стимуляция бета-адренорецепторов активирует киназы, которые фосфорилируют транспортеры
Адренергический сигнальный путь в кардиомиоцитах
Стимуляция бета-адренорецепторов активирует киназы, которые фосфорилируют транспортеры
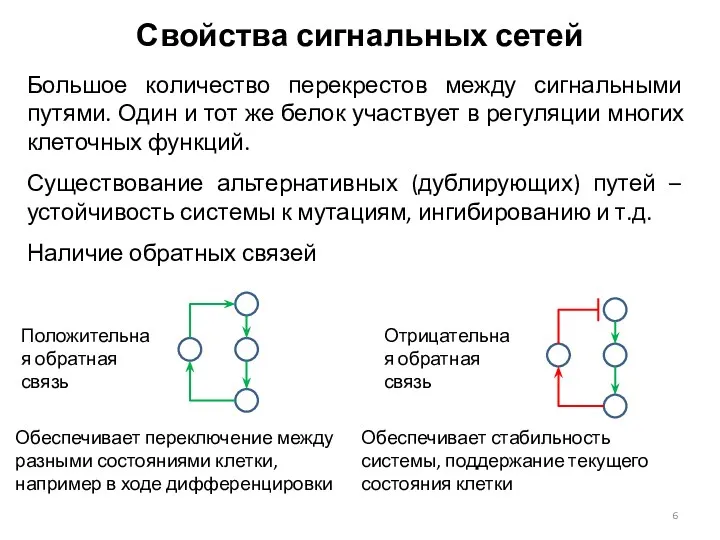
Свойства сигнальных сетей
Большое количество перекрестов между сигнальными путями. Один и тот
Свойства сигнальных сетей
Большое количество перекрестов между сигнальными путями. Один и тот

Метаболические пути
Метаболический путь – последовательность химических реакций, осуществляющих синтез продукта (или
Метаболические пути
Метаболический путь – последовательность химических реакций, осуществляющих синтез продукта (или
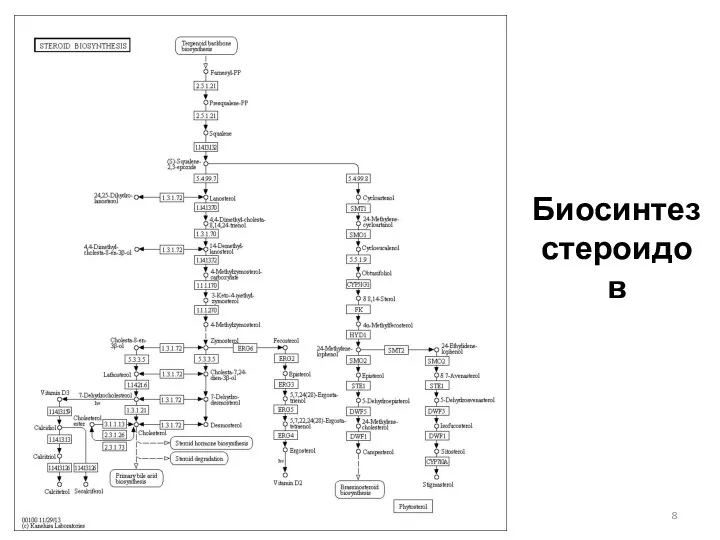
Биосинтез стероидов
Биосинтез стероидов

Реконструкция метаболической сети клетки
Реконструкция метаболической сети клетки

Базы данных по сигнальным и метаболическим путям
Базы данных по сигнальным и метаболическим путям
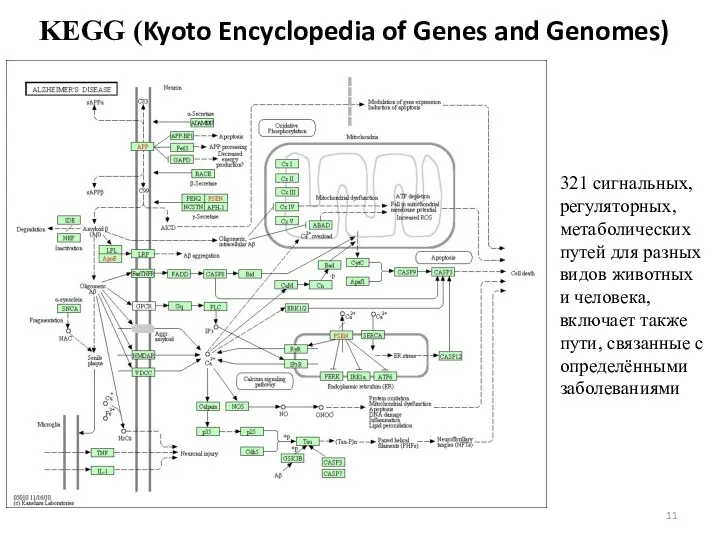
KEGG (Kyoto Encyclopedia of Genes and Genomes)
321 сигнальных, регуляторных, метаболических путей
KEGG (Kyoto Encyclopedia of Genes and Genomes)
321 сигнальных, регуляторных, метаболических путей
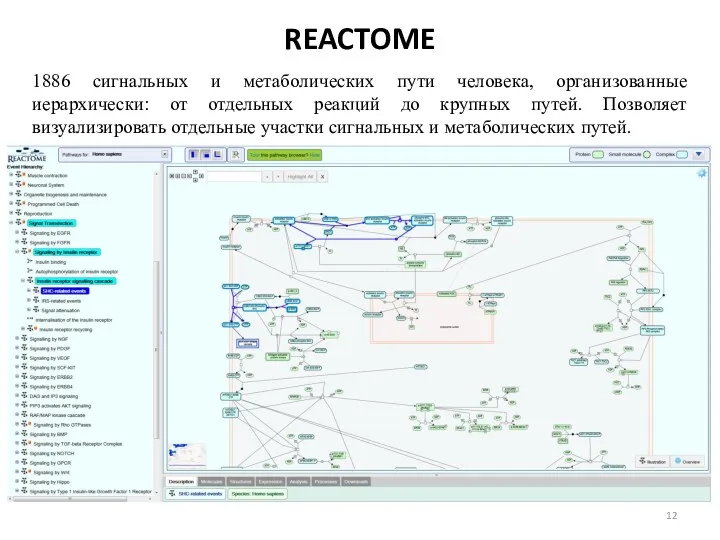
REACTOME
1886 сигнальных и метаболических пути человека, организованные иерархически: от отдельных реакций
REACTOME
1886 сигнальных и метаболических пути человека, организованные иерархически: от отдельных реакций
SIGNOR
SIGNOR

Сети белок-белковых взаимодействий (ББВ)
Вершины данной сети представляют собой белки, а ребра
Сети белок-белковых взаимодействий (ББВ)
Вершины данной сети представляют собой белки, а ребра

Подходы к оценке белок-белковых взаимодействий
Ко-иммунопреципитация
Аффинная хроматография
Перекресная сшивка при помощи реактивных соединений
Подходы к оценке белок-белковых взаимодействий
Ко-иммунопреципитация
Аффинная хроматография
Перекресная сшивка при помощи реактивных соединений

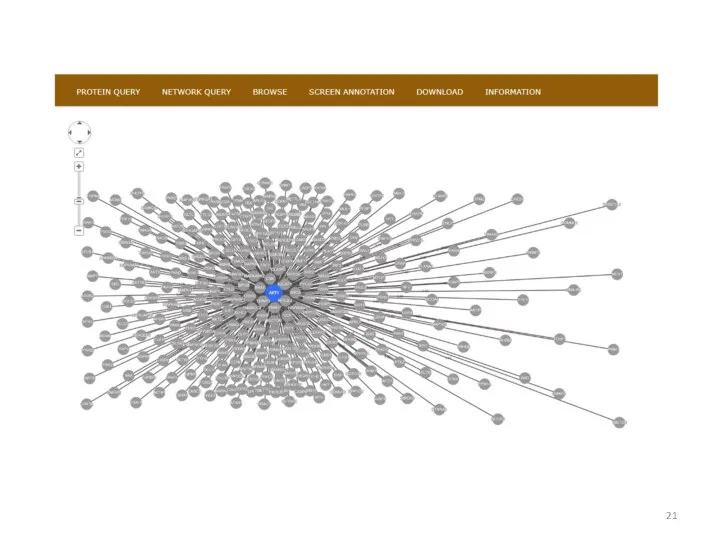
Базы данных по белок-белковым взаимодействиям
Базы данных по белок-белковым взаимодействиям
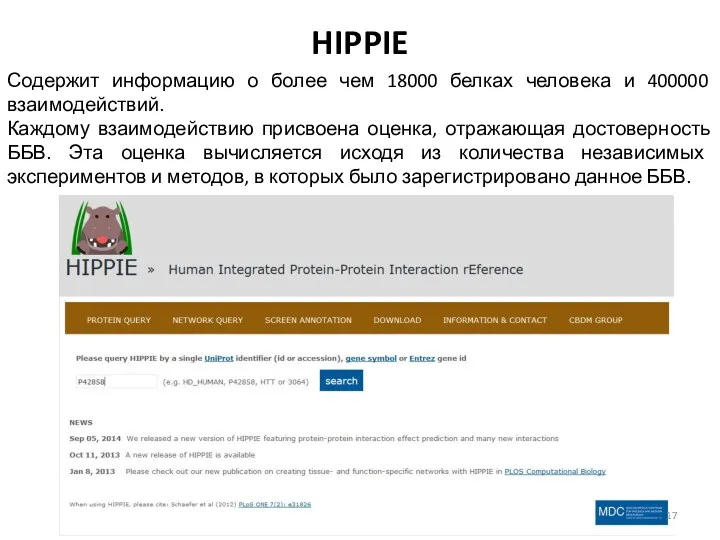
HIPPIE
Содержит информацию о более чем 18000 белках человека и 400000 взаимодействий.
Каждому
HIPPIE
Содержит информацию о более чем 18000 белках человека и 400000 взаимодействий.
Каждому

Базы данных по экспрессии генов
Базы данных по экспрессии генов
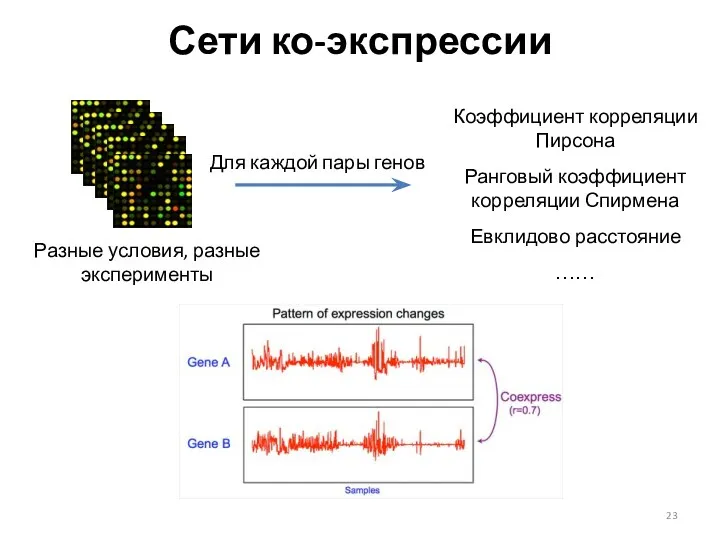
Сети ко-экспрессии
Разные условия, разные эксперименты
Для каждой пары генов
Коэффициент корреляции Пирсона
Ранговый коэффициент
Сети ко-экспрессии
Разные условия, разные эксперименты
Для каждой пары генов
Коэффициент корреляции Пирсона
Ранговый коэффициент
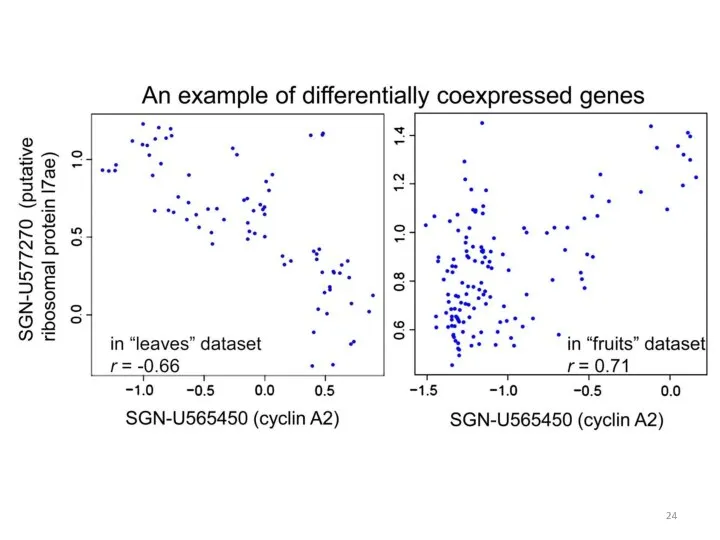
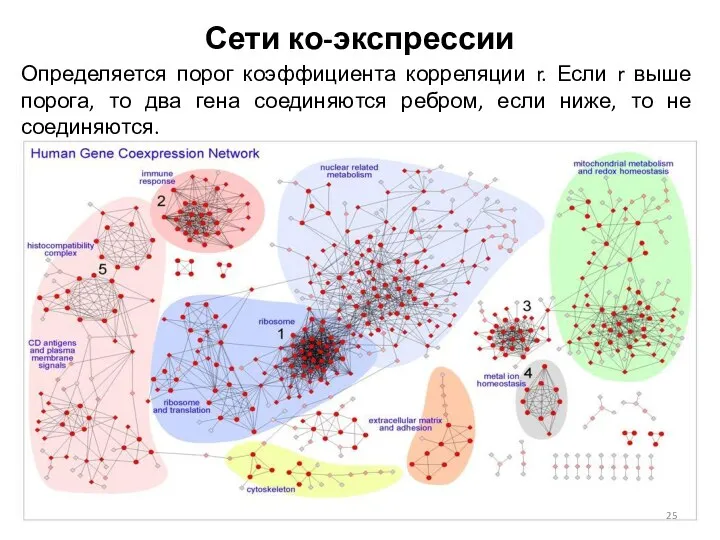
Сети ко-экспрессии
Определяется порог коэффициента корреляции r. Если r выше порога, то
Сети ко-экспрессии
Определяется порог коэффициента корреляции r. Если r выше порога, то
COXPRESdb
COXPRESdb
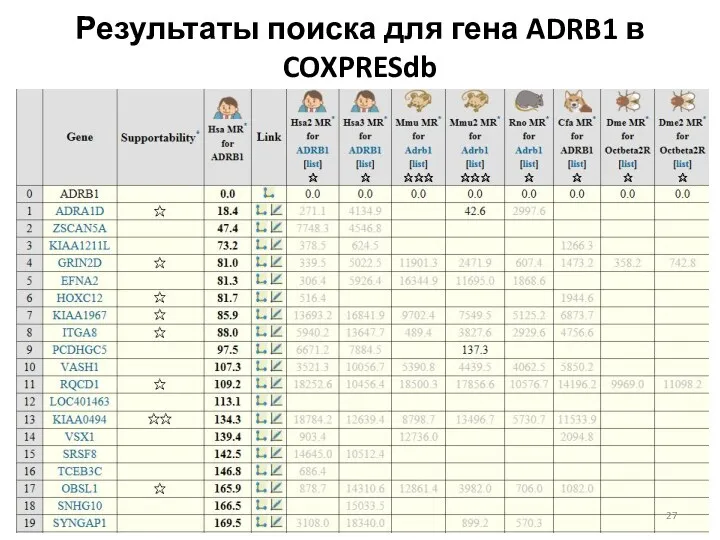
Результаты поиска для гена ADRB1 в COXPRESdb
Результаты поиска для гена ADRB1 в COXPRESdb
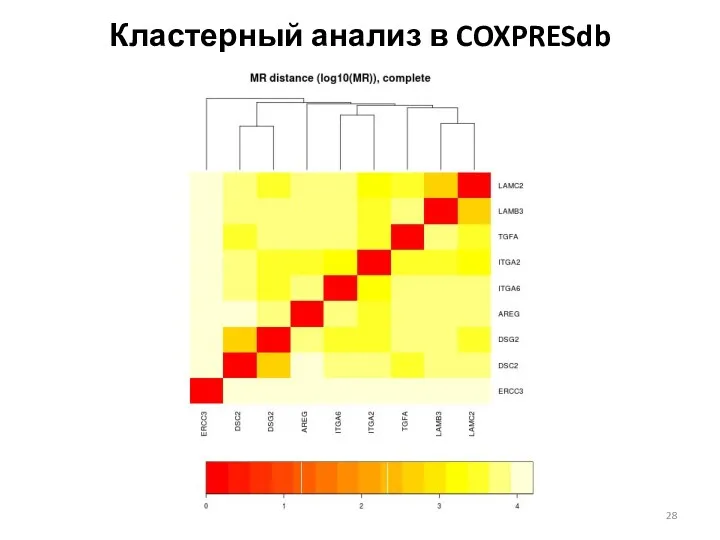
Кластерный анализ в COXPRESdb
Кластерный анализ в COXPRESdb

Сети на основе интеграции информации
Сигнальные сети построены на основе интеграции данных
Сети на основе интеграции информации
Сигнальные сети построены на основе интеграции данных

Примеры средств для визуализации и анализа сетей
Примеры средств для визуализации и анализа сетей
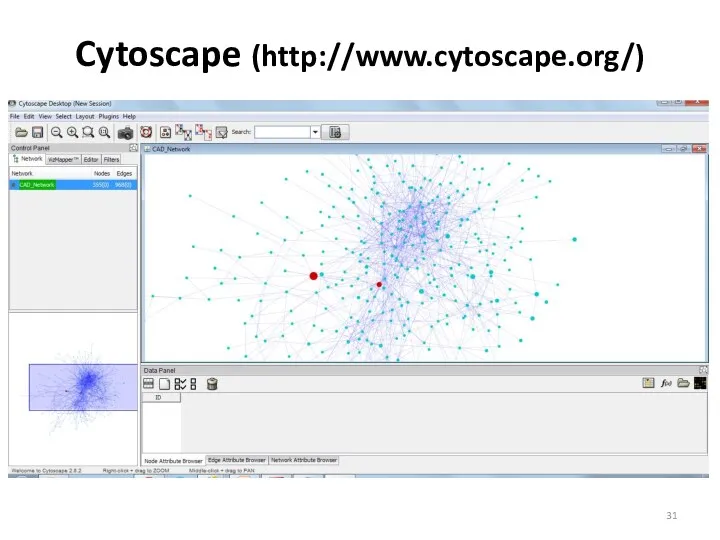
Cytoscape (http://www.cytoscape.org/)
Cytoscape (http://www.cytoscape.org/)

Cytoscape
Платформа с открытым исходным кодом для визуализации и анализа биологических сетей.
Возможность
Cytoscape
Платформа с открытым исходным кодом для визуализации и анализа биологических сетей.
Возможность

Анализ топологии сетей
Анализ топологии сетей
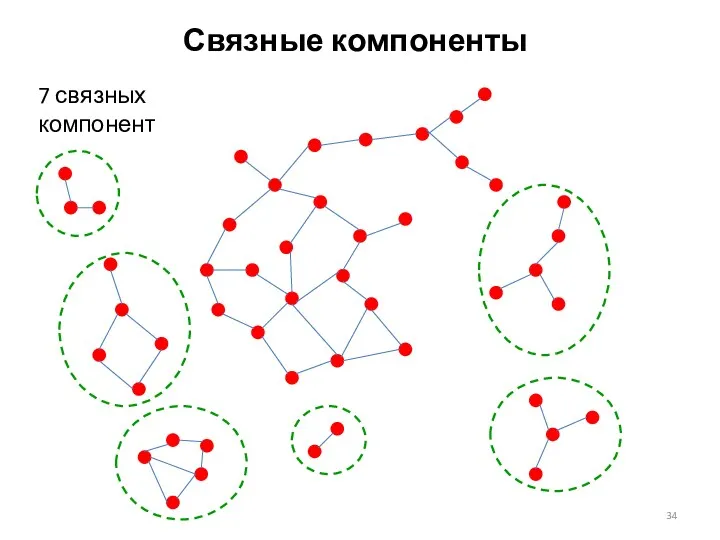
Связные компоненты
7 связных компонент
Связные компоненты
7 связных компонент

Путь в сети (графе)
Путь – это последовательность вершин {x1, x2, …,
Путь в сети (графе)
Путь – это последовательность вершин {x1, x2, …,
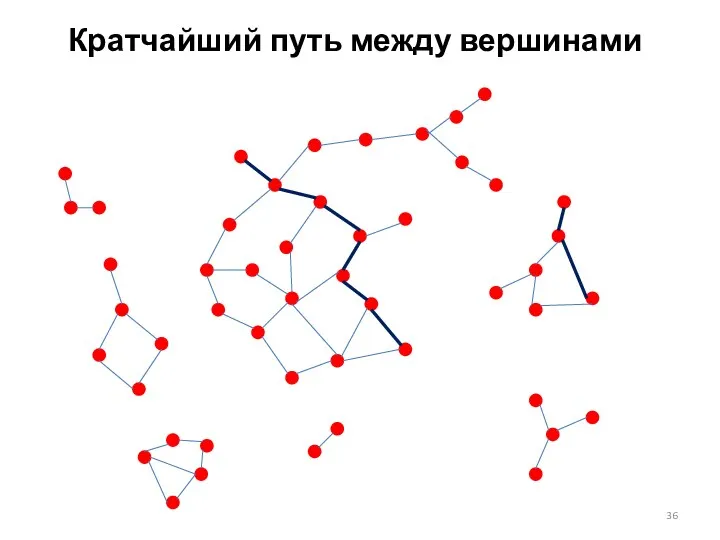
Кратчайший путь между вершинами
Кратчайший путь между вершинами
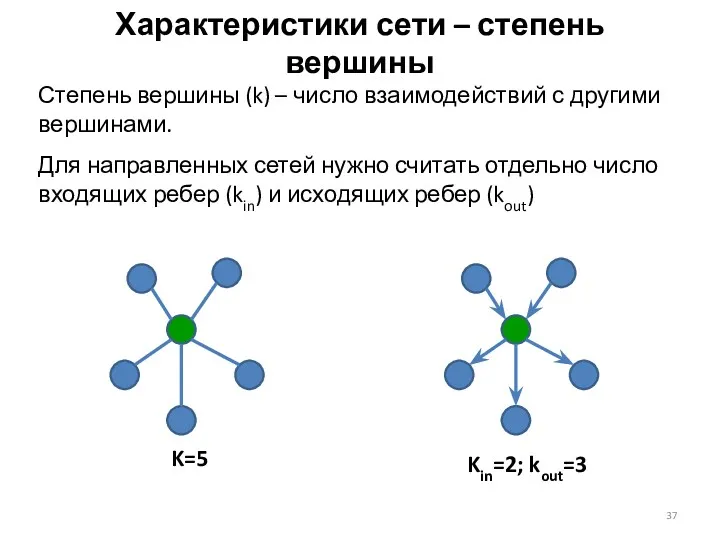
Характеристики сети – степень вершины
Степень вершины (k) – число взаимодействий с
Характеристики сети – степень вершины
Степень вершины (k) – число взаимодействий с
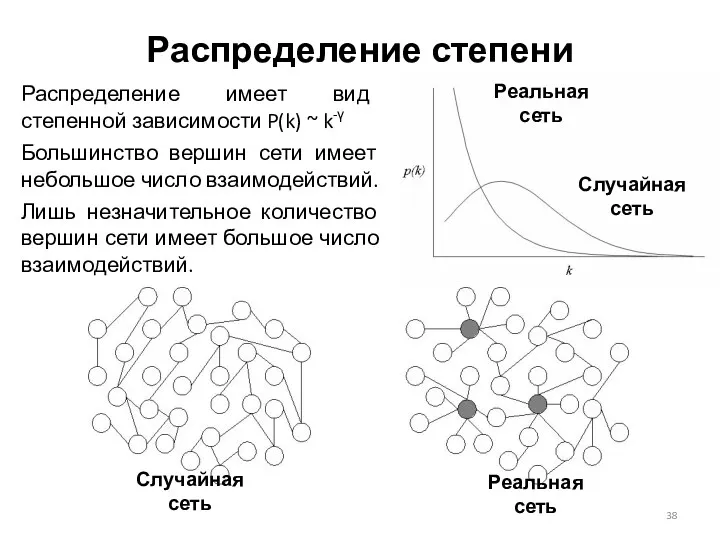
Распределение степени
Распределение имеет вид степенной зависимости P(k) ~ k-γ
Большинство вершин сети
Распределение степени
Распределение имеет вид степенной зависимости P(k) ~ k-γ
Большинство вершин сети

Свойства белков-«хабов»
Белки с большим числом взаимодействий называются белками- «хабами».
Многие «хабы» являются
Свойства белков-«хабов»
Белки с большим числом взаимодействий называются белками- «хабами».
Многие «хабы» являются

Свойства белков-«хабов»
Белки-«хабы» чаще всего не взаимодействуют непосредственно друг с другом
Белки-«хабы» более
Свойства белков-«хабов»
Белки-«хабы» чаще всего не взаимодействуют непосредственно друг с другом
Белки-«хабы» более
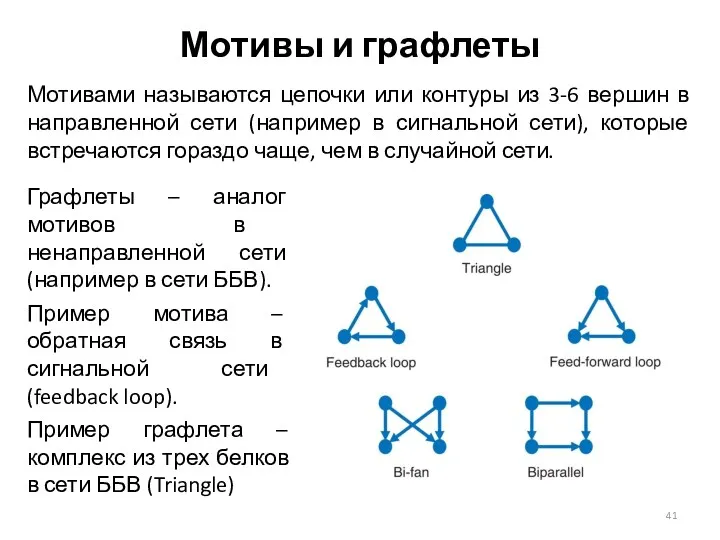
Мотивы и графлеты
Мотивами называются цепочки или контуры из 3-6 вершин в
Мотивы и графлеты
Мотивами называются цепочки или контуры из 3-6 вершин в
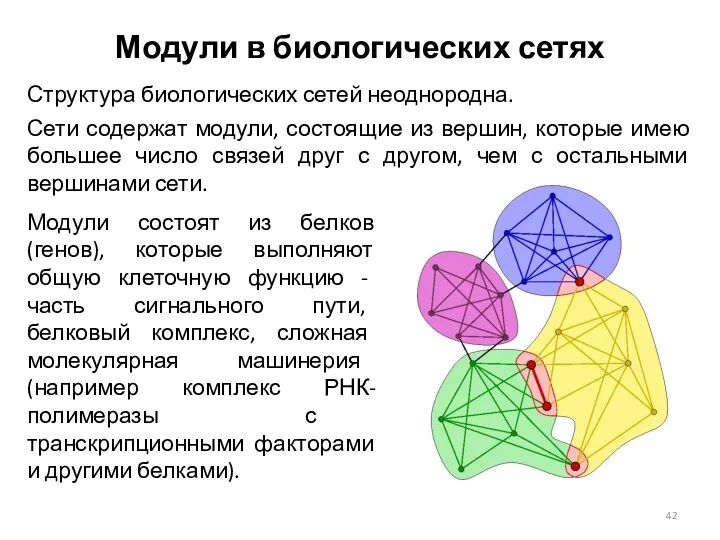
Модули в биологических сетях
Структура биологических сетей неоднородна.
Сети содержат модули, состоящие из
Модули в биологических сетях
Структура биологических сетей неоднородна.
Сети содержат модули, состоящие из
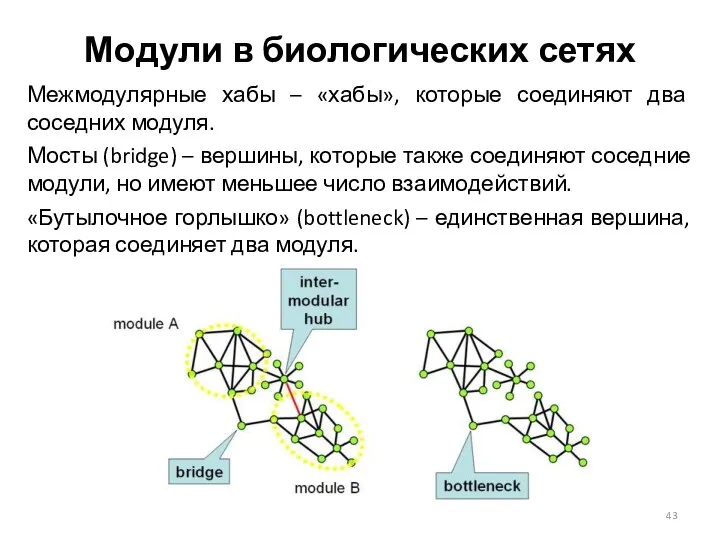
Модули в биологических сетях
Межмодулярные хабы – «хабы», которые соединяют два соседних
Модули в биологических сетях
Межмодулярные хабы – «хабы», которые соединяют два соседних

Модули в биологических сетях
Межмодулярные «хабы» представляют собой временные «хабы», для которых
Модули в биологических сетях
Межмодулярные «хабы» представляют собой временные «хабы», для которых

Предсказание маркеров заболеваний
Белки (гены), связанные с определенным заболеванием, обладают тенденцией формировать
Предсказание маркеров заболеваний
Белки (гены), связанные с определенным заболеванием, обладают тенденцией формировать
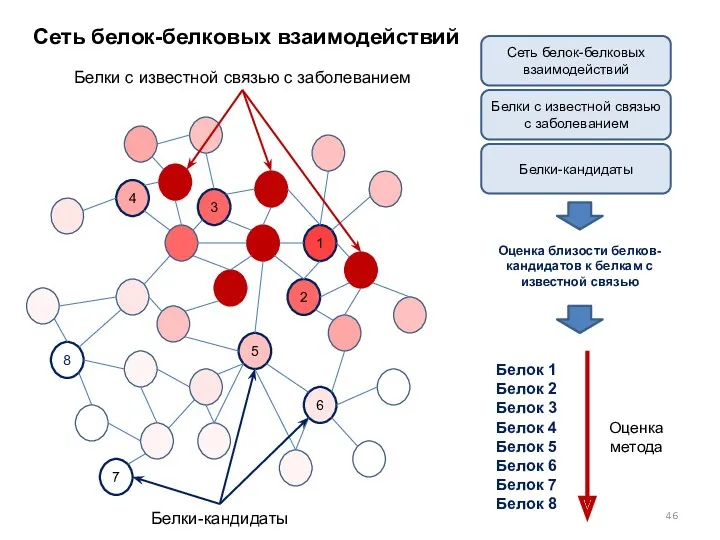
Сеть белок-белковых взаимодействий
Белки с известной связью с заболеванием
Белки-кандидаты
Сеть белок-белковых взаимодействий
Белки с известной связью с заболеванием
Белки-кандидаты
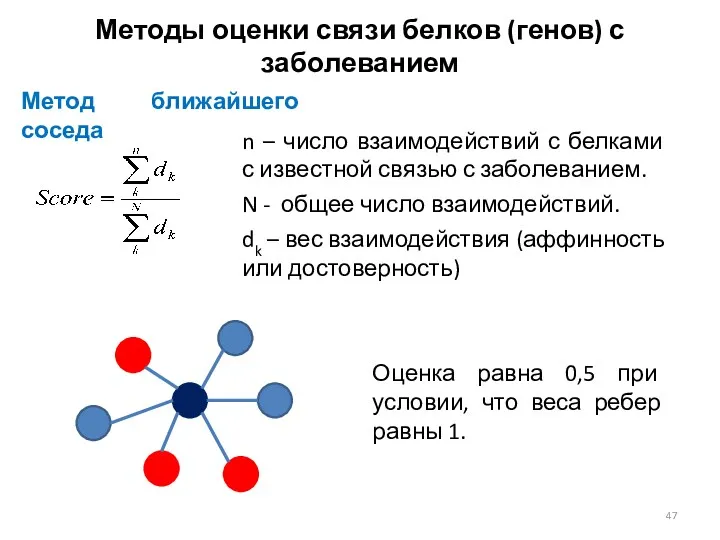
Методы оценки связи белков (генов) с заболеванием
Метод ближайшего соседа
n –
Методы оценки связи белков (генов) с заболеванием
Метод ближайшего соседа
n –

Случайное блуждание с возвратом
Методы оценки связи белков (генов) с заболеванием
Вычисляется
Случайное блуждание с возвратом
Методы оценки связи белков (генов) с заболеванием
Вычисляется

Оценка связи белка (гена) с заболеванием
Широкомасштабные генетические исследования. Оценка корреляции полиморфизма
Оценка связи белка (гена) с заболеванием
Широкомасштабные генетические исследования. Оценка корреляции полиморфизма

Базы данных по связи белков/генов с заболеваниями
Базы данных по связи белков/генов с заболеваниями
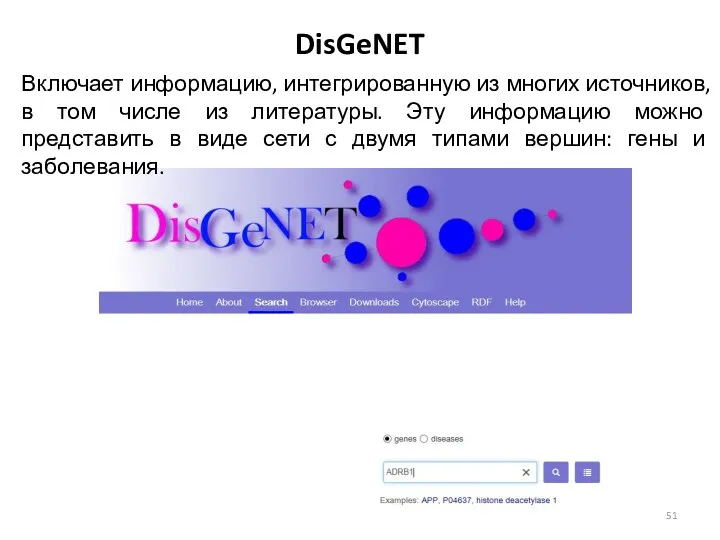
DisGeNET
Включает информацию, интегрированную из многих источников, в том числе из литературы.
DisGeNET
Включает информацию, интегрированную из многих источников, в том числе из литературы.
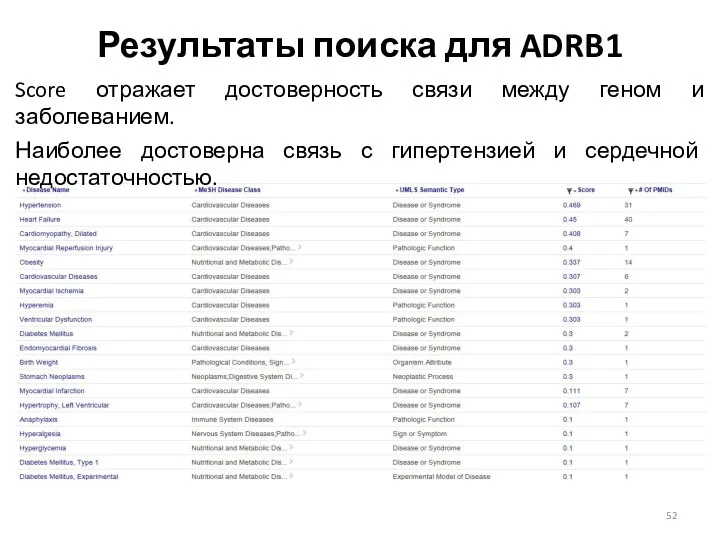
Результаты поиска для ADRB1
Score отражает достоверность связи между геном и заболеванием.
Наиболее
Результаты поиска для ADRB1
Score отражает достоверность связи между геном и заболеванием.
Наиболее

Связь ADRB1 с гипертензией
Указан источник данных. Показаны предложения из Abstarcts PubMed,
Связь ADRB1 с гипертензией
Указан источник данных. Показаны предложения из Abstarcts PubMed,
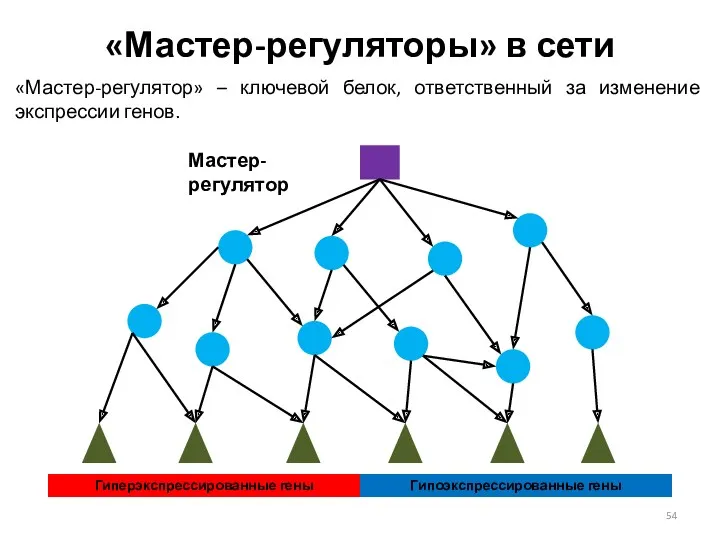
«Мастер-регуляторы» в сети
«Мастер-регулятор» – ключевой белок, ответственный за изменение экспрессии генов.
Гиперэкспрессированные
«Мастер-регуляторы» в сети
«Мастер-регулятор» – ключевой белок, ответственный за изменение экспрессии генов.
Гиперэкспрессированные

Этапы поиска «мастер-регуляторов»
Идентификация дифференциально экспрессирующихся генов (норма-патология, лекарство-растворитель и др.);
Поиск экспериментальной
Этапы поиска «мастер-регуляторов»
Идентификация дифференциально экспрессирующихся генов (норма-патология, лекарство-растворитель и др.);
Поиск экспериментальной

Моделирование динамики сетей
Моделирование динамики сетей

Моделирование динамики сетей
Моделирование изменения концентраций субстратов/продуктов реакций во времени.
В сигнальной сети
Моделирование динамики сетей
Моделирование изменения концентраций субстратов/продуктов реакций во времени.
В сигнальной сети

Моделирование динамики сетей
Дифференциальные уравнения
Логические сети
Сети Петри
Клеточный автомат
Гибридные методы
Моделирование динамики сетей
Дифференциальные уравнения
Логические сети
Сети Петри
Клеточный автомат
Гибридные методы

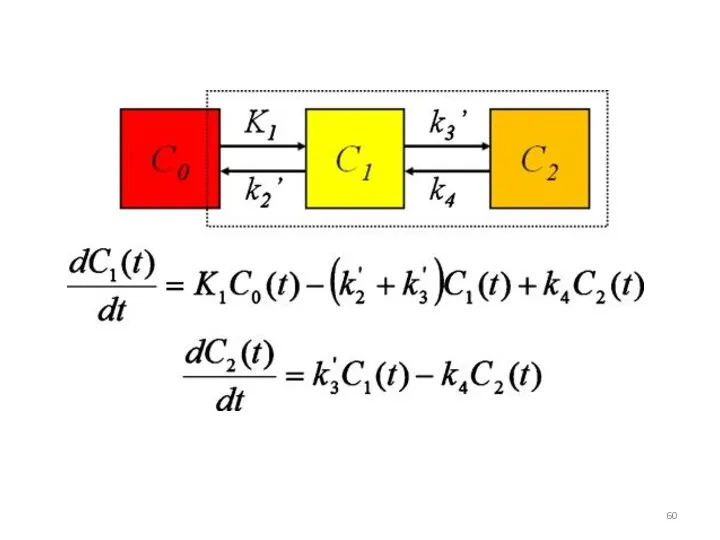
Дифференциальные уравнения
Система дифференциальных уравнений, описывающих изменение концентраций РНК и белков во
Дифференциальные уравнения
Система дифференциальных уравнений, описывающих изменение концентраций РНК и белков во
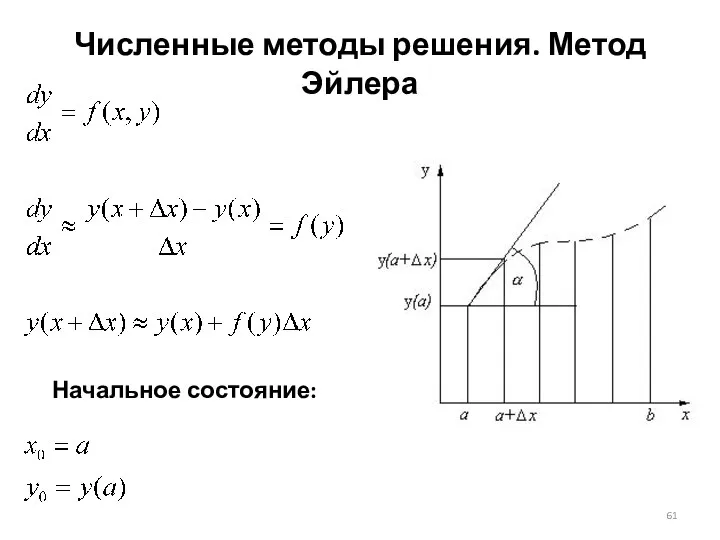
Численные методы решения. Метод Эйлера
Начальное состояние:
Численные методы решения. Метод Эйлера
Начальное состояние:
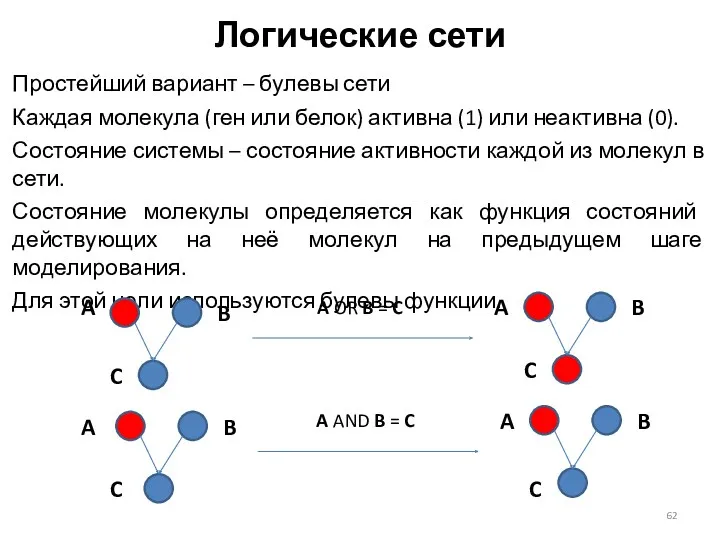
Логические сети
Простейший вариант – булевы сети
Каждая молекула (ген или белок) активна
Логические сети
Простейший вариант – булевы сети
Каждая молекула (ген или белок) активна
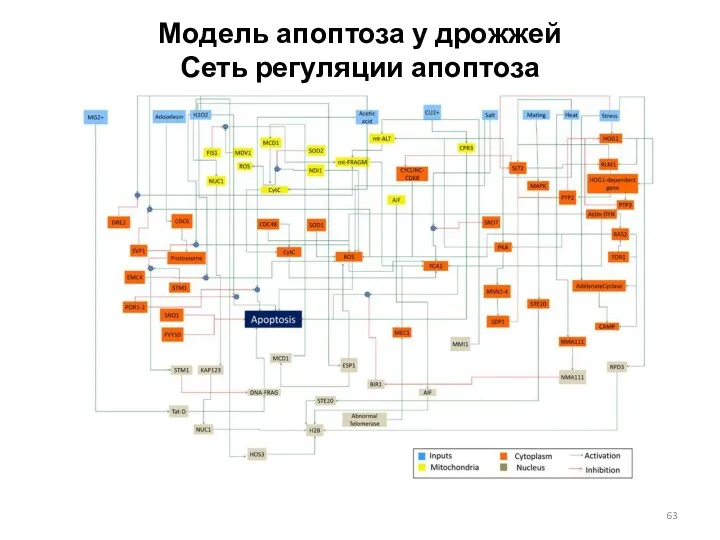
Модель апоптоза у дрожжей
Сеть регуляции апоптоза
Модель апоптоза у дрожжей
Сеть регуляции апоптоза

Для каждого белка записываются булевы функции
Плюсом обозначено логическое «И».
Для каждого белка записываются булевы функции
Плюсом обозначено логическое «И».
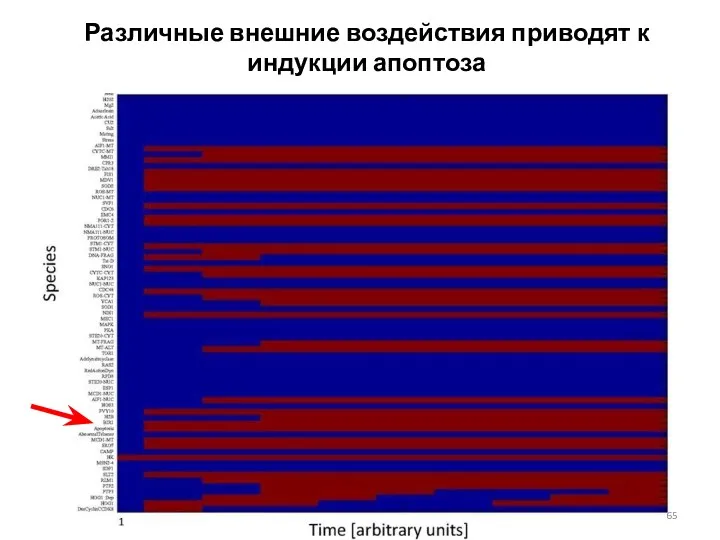
Различные внешние воздействия приводят к индукции апоптоза
Различные внешние воздействия приводят к индукции апоптоза
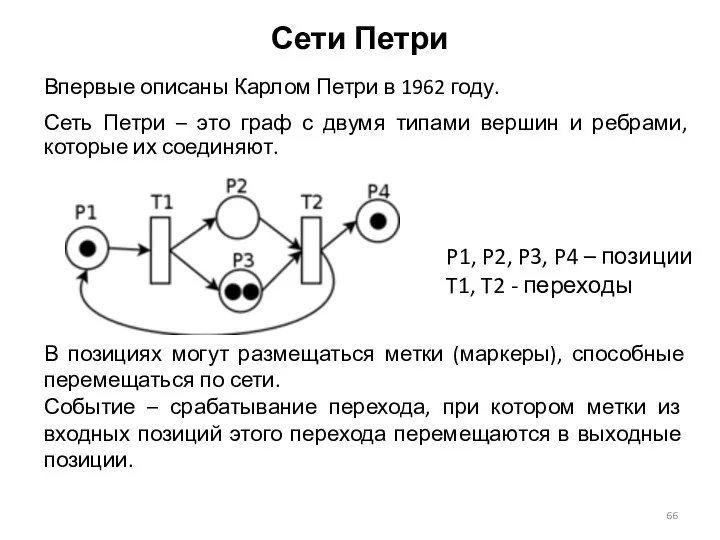
Сети Петри
Впервые описаны Карлом Петри в 1962 году.
Сеть Петри – это
Сети Петри
Впервые описаны Карлом Петри в 1962 году.
Сеть Петри – это
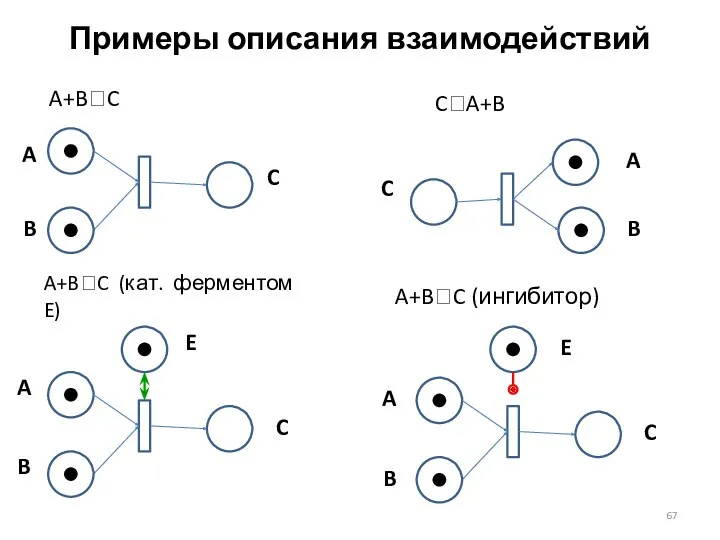
Примеры описания взаимодействий
A+B?C
C?A+B
A+B?C (кат. ферментом E)
A+B?C (ингибитор)
A
B
C
A
B
C
A
B
C
E
A
B
C
E
Примеры описания взаимодействий
A+B?C
C?A+B
A+B?C (кат. ферментом E)
A+B?C (ингибитор)
A
B
C
A
B
C
A
B
C
E
A
B
C
E

Расширение сетей Петри
Временная сеть Петри - такая сеть, где переходы обладают
Расширение сетей Петри
Временная сеть Петри - такая сеть, где переходы обладают

Клеточный автомат
Состоит из регулярной решетки ячеек, каждая из которых может находится
Клеточный автомат
Состоит из регулярной решетки ячеек, каждая из которых может находится

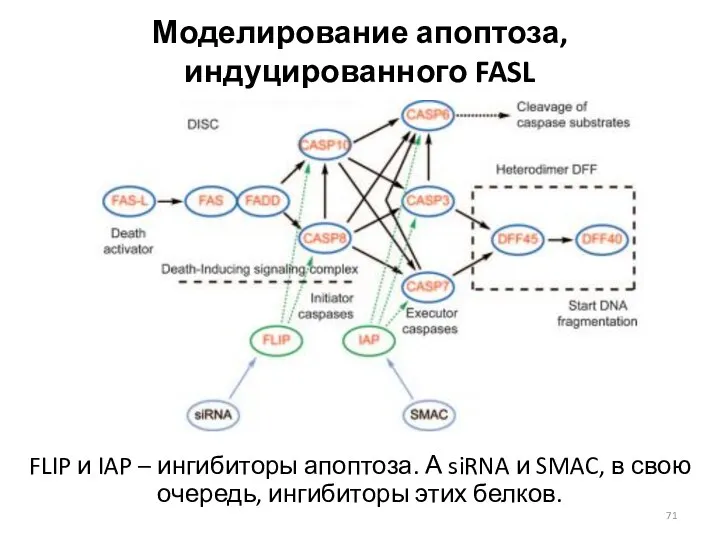
Моделирование апоптоза, индуцированного FASL
FLIP и IAP – ингибиторы апоптоза. А siRNA
Моделирование апоптоза, индуцированного FASL
FLIP и IAP – ингибиторы апоптоза. А siRNA

Описание взаимодействий
Описание взаимодействий
 Иван Петрович Павлов (1849-1936)
Иван Петрович Павлов (1849-1936) Мәмлерің түрлері
Мәмлерің түрлері разработка урока для 7 класса на тему: Тип плоские черви. Белая планария
разработка урока для 7 класса на тему: Тип плоские черви. Белая планария Сердечно-сосудистая система
Сердечно-сосудистая система Саморазвитие экосистемы
Саморазвитие экосистемы Раціональне харчування
Раціональне харчування Вегетативное размножение растений
Вегетативное размножение растений Классификацая органического мира
Классификацая органического мира Презентация Грибы
Презентация Грибы Альтернативные способы обеззараживания семенного материала
Альтернативные способы обеззараживания семенного материала энергетический обмен
энергетический обмен Ямчатость стебля
Ямчатость стебля Цитология. Клетка как биологическая система
Цитология. Клетка как биологическая система Влияние условий хранения хлеба на образование плесени на нём
Влияние условий хранения хлеба на образование плесени на нём Микробиология мяса
Микробиология мяса Деление клетки
Деление клетки Рослини – символи України
Рослини – символи України Тип Членистоногие (Arthropoda)
Тип Членистоногие (Arthropoda) Вирусы. Живые ли вирусы или нет?
Вирусы. Живые ли вирусы или нет? Химиотерапевтические препараты. Антибиотики. Бактериофаги
Химиотерапевтические препараты. Антибиотики. Бактериофаги Строение и функции клетки
Строение и функции клетки Тотығу стрестің өсімдіктерге әсері
Тотығу стрестің өсімдіктерге әсері Экологическая ниша
Экологическая ниша Желудочная секреция
Желудочная секреция Формы размножения организмов
Формы размножения организмов Рекомбинантные антитела для диагностики и терапии
Рекомбинантные антитела для диагностики и терапии Урок на тему: ВОДА – ИСТОЧНИК ЖИЗНИ
Урок на тему: ВОДА – ИСТОЧНИК ЖИЗНИ Порядок Capparales - каперсоцветные
Порядок Capparales - каперсоцветные