- Расширенный материал по лекциям углеводы биоэнергетика

Содержание
- 2. Углеводы – многоатомные альдегидоспирты или кетоспирты Термин введён в 1876 г Шмидтом. Другой термин – глициды.
- 3. Биологическая роль углеводов Энергетическая (распад→СО2 Н2О АТФ) Пластическая (хондроитинсульфат) Резервная (гликоген) Защитная (мембраны, смазка суставов) Регуляторная
- 4. Классификация углеводов В зависимости от сложности строения делят на 3 класса: моносахариды олигосахариды полисахариды
- 6. Моносахариды МОНОСАХАРИД (МОНОЗА) – минимальная структурная единица углеводов, при дроблении которой исчезают свойства сахаров В зависимости
- 7. По строению функциональной группы делят на альдозы и кетозы Самый простой сахар - глицериновый альдегид
- 8. Моносахариды – стереоизомеры. D- и L-форма определяется ориентацией Н- и ОН-групп при атоме С, соседнем с
- 9. Большинство моносахаридов у млекопитающих имеет D-конфигурацию. Именно к ней специфичны фермен-ты, ответственные за метаболизм моноз, поэтому
- 10. Молекулярный состав глюкозы и фруктозы одинаков (С6Н12О6), но структура функцио-нальных групп разная (альдоза и кетоза)
- 11. Основная форма существования углеводов в растворах – циклическая (результат внутримолекулярной реакции) "мутаротация" – взаимопревращение форм глюкозы
- 12. Циклические формы моносахаридов приобретают активные ОН-группы при С1 или С2 или С4 атомах. Гликозидные гидроксилы участвуют
- 13. Образуются при замещении ОН-групп у С2 на NH2-группу. Наиболее важны глюкозамин и галактозамин, их аминогруппа часто
- 14. Сахарные кислоты (глюкаровые, глюконовые, глюкуроновые), аскорбиновая кислота (не у человека!) получаются из глюкозы при воздействии окислителей.
- 15. САХАРНЫЕ СПИРТЫ При восстановлении моносахаров образуются многоатомные спирты: из глюкозы сорбит, из маннозы маннит, из фруктозы
- 16. ОЛИГОСАХАРИДЫ включают от 2 до 10 остатков моносахаридов, соединены 1,4- или 1,2-гликозидными связями, образуются между двумя
- 17. Сахароза (тростниковый или свекловичный сахар) – Это глюкоза и фруктоза, соединённые α-1,2-гликозидной связью. Расщепляет сахарозу фермент
- 18. Мальтоза (фруктовый сахар) 2 молекулы глюкозы, соединены α-1,4-гликозидной связью. Образуется в ЖКТ при гидролизе пищевого крахмала.
- 19. Лактоза (молочный сахар) молекулы глюкозы и галактозы, соединены β-1,4-гликозидной связью. Синтезируется в период лактации. Поступление лактозы
- 20. ПОЛИСАХАРИДЫ Большинство природных углеводов – полимеры, число моносахаридных остатков от 10 до десятков тысяч По функциональным
- 21. По составу: гомо-, гетерополисахариды Гомополисахариды (гомогликаны) состоят из моносахаридных звеньев одного типа. Основные представители – крахмал,
- 22. Амилоза обладает линейной структурой, все глюкозные остатки соединены α(1-4)- гликозидной связью. В молекуле амилозы ≈ 100-1000
- 23. Амилопектин разветвлен, имеет через каждые 24-30 остатков глюкозы, соединённых альфа(1-4)-связями, небольшое число альфа(1-6)-связей. В составе амилопектина
- 24. основная составная часть клеточной стенки растений. Состоит из ≈ 2000-11000 остатков глюкозы, в отличие от крахмала
- 25. Гликоген – животный крахмал Содержит 6.000 – 300.000 остатков глюкозы. Более разветвлен, чем амилопектин: α(1-6)-связи в
- 26. Другие резервные полисахариды Бактерии ротовой полости способны расщеплять сахарозу: сахароза →глюкоза + фруктоза 1) Декстран –
- 27. Вариант СТРОЕНИЯ ДЕКСТРАНА В локализации α(1-6)- и α(1-4)- гликозидных связей в линейной части и местах ветвления
- 28. 2) Леваны – резервные гомополисахариды из остатков D-фруктозы, т.е. являются фруктанами Бактерии ротовой полости, расщепляя сахарозу,
- 29. Нумерация С-атомов фруктозы в составе сахарозы Синтез левана микрофлорой ПР Первая молекула фруктозы присоединяется к молекуле
- 30. Гетерополисахариды (гетерогликаны) сложные углеводы, состоят из 2-х и более типов моносахаридов (аминосахара, уроновые кислоты), чаще всего
- 31. Гиалуроновая кислота – линейный полимер из глюкуроновой кислоты и ацетилглюкозамина. Входит в состав клеточных стенок, синовиальной
- 32. Хондроитинсульфаты – разветвленные сульфатированные полимеры из глюкуроновой кислоты и N-ацетилглюкозамина. Основные структурные компоненты хрящей, сухожилий, роговицы
- 33. Гликопротеины и гликолипиды 1) Комплексы белка с углеводами, существует 2 вида: гликопротеины и протеогликаны. Отличаются по
- 34. Норма углеводов в питании Запас углеводов в организме не превышает 2-3% от массы тела. За счет
- 35. ВНЕШНИЙ ОБМЕН УГЛЕВОДОВ (переваривание) Переваривание – совокупность процессов ферментативного гидролиза поступающих с пищей макромолекул полисахаридов, белков,
- 36. Усвоение олиго- и полисахаридов идёт при их гидролитическом расщеплении до моносахаров. Гликозидазы атакуют 1-4 и 1-6
- 37. ПОЛОСТНОЕ ПИЩЕВАРЕНИЕ Переваривание полисахаридов начинается в ротовой полости, где крахмалы подвергаются хаотичному действию α-амилазы слюны по
- 38. Далее пища идёт в 12-ПК и кишечник (значения рН нейтральные) и вновь подвергается действию 1) амилаз
- 39. ПРИСТЕНОЧНОЕ ПИЩЕВАРЕНИЕ Гидролиз дисахаридов происходит не в просвете кишечника, а на поверхности клеток слизистой оболочки под
- 40. Строение пристеночных ферментов пример: сахаразо-изомальтазный комплекс. 1- сахараза, 2 - изомальтаза, 3 - связывающий домен, 4
- 41. НЕПЕРЕНОСИМОСТЬ УГЛЕВОДОВ Лактоза поступает в организм с молоком. У ребёнка при недостаточной активности «лактазы»: лактоза, которой
- 42. Лечение непереносимости углеводов. Диета с исключением из рациона: в первом случае – молока, во втором случае
- 43. Целлюлоза в организме человека У человека нет ферментов для расщепления β(1-4)-гликозидной связи целлюлозы. Однако микрофлора толстого
- 44. Всасываются в кишечнике только моносахара (глюкоза,галактоза,фруктоза) Их перенос в энтероциты происходит: 1) путём пассивной диффузии по
- 46. Транспорт моносахаров в кишечнике человека
- 47. ДАЛЬНЕЙШИЕ ПРЕВРАЩЕНИЯ САХАРОВ В ОРГАНИЗМЕ В результате переваривания углево-дов и частичной инверсии сахаров в энтероцитах кишечника
- 48. Взаимопревращения сахаров в эпителии кишечника и гепатоцитах Цель создать 1 субстрат (α-D-глюкозу) для экономии ресурсов при
- 49. После печени в общий кровоток поступает только После печени в общий кровоток поступает только
- 50. Роль печени в обмене глюкозы - Регуляция уровня глюкозы в крови - Депонирование глюкозы (гликоген) -
- 51. Если поступление глюкозы в гепатоциты выше их возможностей полимеризовать глюкозу в гликоген, то в крови будет
- 52. Транспорт глюкозы из крови в клетки 1) Пассивная диффузия, т.е. прохождение глюкозы через клеточную мембрану по
- 53. Облегчённая диффузия (ГлюТ) Этот вид транспорта особо важен для эритроцитов и нервных клеток. В клетки мозга
- 54. Влияние инсулина на транспортёры глюкозы (белки-переносчики "ГлюТ") Различают 12 типов транспортеров глюкозы на мембранах различных клеток.
- 55. Облегчённая диффузия имеет принципиальное значение для мышц и жировой ткани, которые имеют белки-переносчики типа ГлюТ-4 для
- 56. Дальнейшая судьба глюкозы в организме В клетке глюкоза прежде всего фосфорили-руется, в результате чего приобретает заряд
- 57. В клетках печени есть два изофермента, которые могут фосфорилировать глюкозу: глюкокиназа и гексокиназа. В других органах
- 58. Пути использования глюкозы в клетке
- 59. Если в клетке образовался избыток Глюкозо-6-Ф, она начинает запасать глюкозу в виде гликогена. Сначала фосфоглюкомутаза (класс
- 61. Гликоген образуется почти во всех клетках орга-низма, но max его концентрация в печени и мышцах. Запасы
- 62. ГЛИКОГЕНОЛИЗ Распад гликогена до молочной кислоты может идти при недостатке кислорода. Гликоген присутствует в клетках в
- 63. Схема гормональной регуляции работы гликоген- синтазы и гликоген- фосфорилазы
- 65. Усиление сигнала адреналина в клетку через инозитолфосфатную систему
- 66. Распад гликогена регулируют гормоны – адреналин, глюкагон, глюкокортикоиды. Инсулин оказывает противоположный эффект – активирует синтез гликогена.
- 67. цАМФ в клетке существует недолго. Фосфоди-эстераза быстро превращает его в АМФ : цАМФ + Н2О АМФ
- 68. В тот момент, когда "включается" фосфорилаза, одновременно происходит "выключение" гликоген-синтазы. Это имеет очень глубокий биологический смысл:
- 69. Обратное превращение фосфорилазы а в неактивную b-форму происходит при действии фосфатазы фосфорилазы, которую стимулирует инсулин. Одновременно
- 70. Отщепившаяся от гликогена молекула глюкозо-1-Ф изомеризуется с образованием глюкозо-6-Ф глюкозо-1-Ф глюкозо-6-Ф - Когда самой клетке нужна
- 71. Гликогенозы – болезни гликогена При наследственной недостаточности или отсутствии ферментов, необходимых для обмена гликогена, в тканях
- 72. АНАЭРОБНЫЕ ПРЕВРАЩЕНИЯ УГЛЕВОДОВ 1) гликогеногенез 2) гликогенолиз 3) гликолиз 4) глюконеогенез 5) спиртовое (и др.) брожение
- 73. ГЛИКОЛИЗ - главный анаэробный путь утилизации глюкозы 1) Протекает во всех клетках 2) Для эритроцитов –
- 75. лимитирующая реакция
- 76. фосфофруктокиназа Весь ход гликолиза лимитируется уровнем активности этого фермента Это аллостерический фермент Его ингибируют: АТФ, лимонная
- 77. Расщепение 6С на 3С+3С Второй этап гликолиза – освобождение энергии, заключённой в ФГА, и запасание её
- 78. фосфорилирование АДФ с образованием АТФ идёт за счет переноса ~Р энергии вместе с фосфатом из состава
- 79. Если окисление глюкозы идёт по анаэробному пути, то акцептором ẽ (и Н+) становится МОЛОЧНАЯ КИСЛОТА Если
- 80. Энергетический баланс гликолиза С6Н12О6 (глюкоза) → 2 С3Н6О3 (лактат) 1) расход по 1 АТФ в ходе
- 81. ЭНЕРГЕТИЧЕСКИЙ БАЛАНС гликогенолиза 4АТФ – 1АТФ = 3АТФ На 1 молекулу АТФ больше, чем в гликолизе
- 82. Спиртовое БРОЖЕНИЕ Микроорганизмы, в первую очередь дрожжи, осуществляют не молочно-кислое, а спиртовое брожение глюкозы в анаэробных
- 83. 2 СН3СОСООН ПИРУВАТ 2 СН3СОН ацетальдегид Энергетический баланс спиртового брожения как в гликолизе: 2 АТФ 2
- 84. Глюконеогенез Глюконеогенез – синтез глюкозы de novo из неуглеводных компонентов. Протекает в печени и ≈10% в
- 85. Места поступления субстратов (предшественников) для глюконеогенеза
- 86. Лактат преобразуется обратно в пируват Жиры (глицерол) включаются на уровне ДОАФ
- 87. ЦИКЛ КОРИ Метаболический груз работающей мышцы (лактат) передается в печень, где соотношение НАДН/НАД+ ниже, чем в
- 88. Источники глюконеогенеза - только гликогенные аминокислоты В пируват – ала, сер, цис, гли Большинство аминокислот преобразуется
- 89. 30% аминокислот, поступающих в печень – аланин Биологический смысл глюкозо-аланинового цикла – транспорт аминного азота из
- 90. Стадии превращения пирувата в фосфоенолпируват
- 91. оксалоацетат В виде малата транспортируется через мембрану митохондрии, а затем снова преобразуется в оксалоацетат
- 92. процесс ГНГ обратный гликолизу, но в местах 3-х необратимых реакций используются 4 обходных фермента: Ферменты обходных
- 94. Синтез глюкозы в результате глюконеогенеза энергозависим При синтезе 1 глюкозы из 2 лактатов лактат + лактат
- 95. АЭРОБНЫЕ ПРОЦЕССЫ углеводного обмена Прямой – пентозофосфатный путь, Непрямой – переключение гликолиза на уровне пирувата на
- 96. При распаде гликогена освобождающийся глюкозо-1-фосфат (Г-1-Ф), фосфоглюкомутаза превращает в Г-6-Ф Свободная глюкоза крови, попадая в клетку,
- 97. Пути использования глюкозо-6-Ф 1) Гл-6-Ф может распадаться в процессе гликолиза и служить источником энергии для клеток
- 98. Пентозофосфатный путь Другие названия: пентозный шунт (или цикл) гексозомонофосфатный путь фосфоглюконатный окислительный путь апотомический («усекновение главы»)
- 99. Роль ПФП: синтез пентоз и НАДФН в процессе окисления глюкозы. Возникает качественно новый тип метаболической энергии
- 101. ПФП состоит из 2 стадий(ветвей) I-я стадия окислительная, её роль: 1) генерация НАДФН + Н+ –
- 102. 6-фосфоглюконолактон 6-фосфоглюконат рибулозо-5-Ф рибозо-5-Ф
- 103. 2-я стадия НЕОКИСЛИТЕЛЬНАЯ : 1) идёт перенос (ферменты - трансферазы) 2-углеродного компонента -СО-СН2ОН транскетолазой 3-углеродного компонента
- 104. ПЕНТОЗОФОСФАТНЫЙ ПУТЬ Конечный продукт перестроек неокислительной ветви – глюкозо-6-Ф
- 105. Для синтезов клеткам нужно много НАДФН, а рибозы для включения в нуклеотиды и нуклеиновые кислоты требуется
- 106. Суммарные реакции ПФП I-я стадия (с образованием рибозы): Глюкозо-6-Ф + 2НАДФ ↓ рибозо-5-Ф + СО2 +
- 107. ГЛЮКУРОНОВЫЙ ПУТЬ «Путь уроновых кислот» идёт в печени и клетках соединительной ткани. Первый этап (до синтеза
- 108. Генетическая недостаточность глюкозо-6Ф-дегидрогеназы - причина лекарственных гемолитических анемий У этих людей в 10 раз меньше активность
- 109. Синдром Вернике-Корсакова У таких людей генетически снижено сродство транскетолазы к витамину В1 в 10 раз, им
- 110. В анаэробных условиях 3-ФГА может превратиться в пируват и далее в лактат (гликолитическая оксиредукция НАД+/НАДН) В
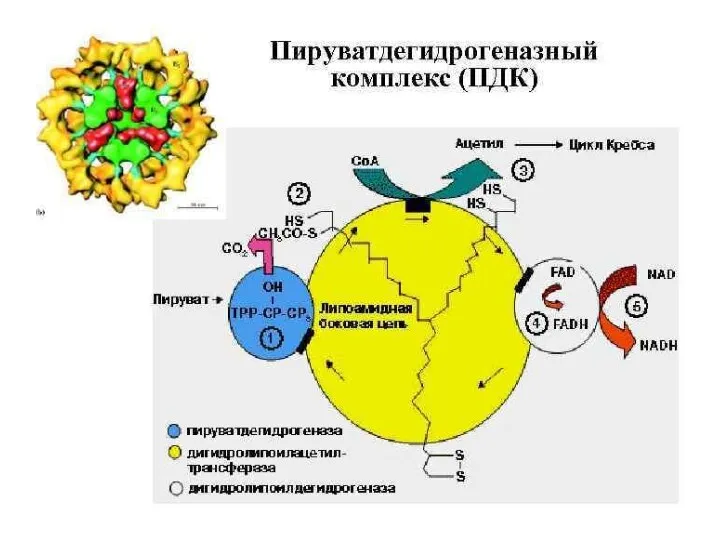
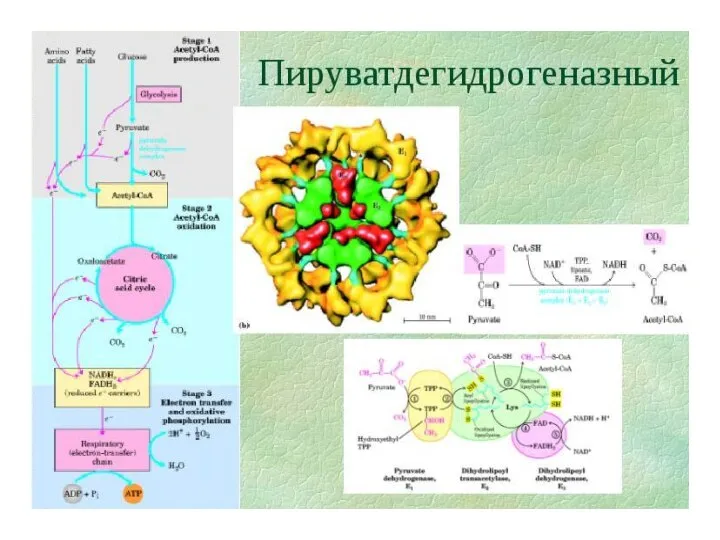
- 111. Пируватдегидрогеназный комплекс 3 фермента: пируватдегидрогеназа, дегидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа 5 коферментов: В1 - тиаминпирофосфат(ТПФ,кокарбоксилаза) N - липоевая кислота(ЛК
- 113. Коферменты ПДГ-комплекса ТПФ ТПФ HS-КоА
- 116. Образование ацетил~КоА из ПВК – ключевой необратимый этап метаболизма Животные не могут превращать ацетил~КоА обратно в
- 117. регуляция активности ПДГ-комплекса 1. Ингибирование продуктами реакции. Ацетил~КоА и НАДН (продукты окисления ПВК) инигибируют ферментный комплекс.
- 118. Биологическая роль окислительного декарбоксилирования ПВК определяется тем, какой будет дальней-шая судьба продуктов этого процесса: НАДН может
- 119. Для запуска ЦТК необходим «катализатор» оксалоацетат Синтез оксалоацетата 1) карбоксилирование ПВК (расход АТФ): 2) окислительное дезаминирование
- 120. Цикл трикарбоновых кислот (=цикл лимонной кислоты, =цикл Кребса) Ганс Кребс – лауреат Нобелевской премии 1953 г
- 121. ЭТАПЫ ЦТК ЭТАПЫ ЦТК
- 123. 1 этап ЦТК: синтез цитрата (этап необратим) 1) реакция конденсации АЦЕТИЛ~SКоА и ОКСАЛОАЦЕТАТА в ЦИТРАТ цитратсинтазой
- 124. 2 этап ЦТК: изомеризация цитрата (2,3 реакции) 2) цис-аконитаза удаляет Н2О с образованием промежу-точной цис-аконитовой формы
- 125. 3 этап ЦТК: окислительное декарбоксилиро- вание изоцитрата (4,5 реакции) фермент изоцитратдегидрогеназа 4) получение НАДН+Н+ Это первая
- 127. 4 этап ЦТК: окислительное декарбоксилиро- вание α-кетоглутарата (6 реакция) 6) процесс окислительного декарбоксилирования α-кетоглутарата катализирует мультиферментный
- 128. 4 этап ЦТК 6 реакция α-кетоглутаратдегидрогеназная (кратко комплекс называют α-кетоглутаратдегидрогеназа и КоЕ не указывают)
- 129. 5 этап ЦТК: получение сукцината - (7 реакция), сукцинаттиокиназная реакция легко обратима. 7) Сукцинил~КоА в сукцинат
- 131. Образование высокоэнергетической фос-фатной связи ГТФ из сукцинил~КоА – есть пример субстратного фосфорилирования. Это единственная реакция ЦТК,
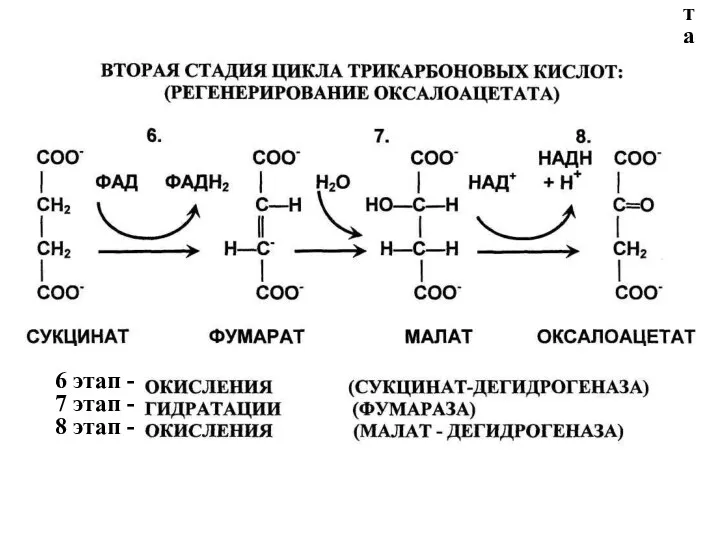
- 132. 6 этап - 7 этап - 8 этап - 6 этап - 7 этап - 8
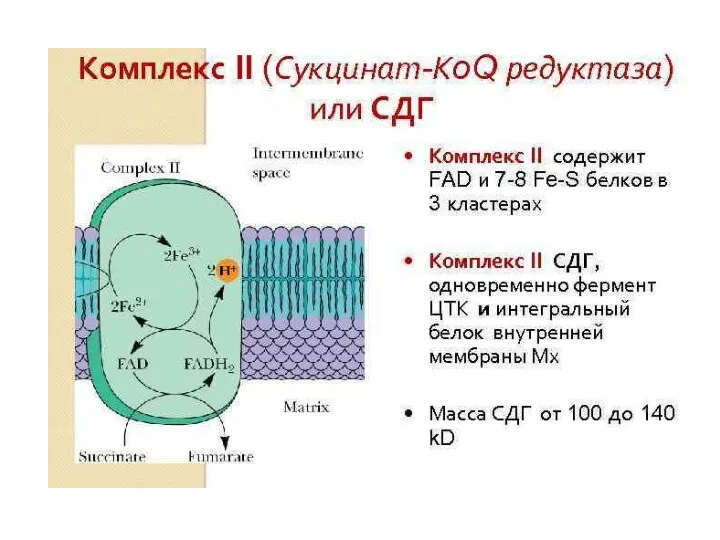
- 133. 6 этап ЦТК: получение сукцината (8 реакция), 3-я редокс-реакция, сукцинат:ФAД-оксидоредуктаза
- 134. 7 этап ЦТК: получение малата (яблочной кислоты) (9 реакция), фумараза гидратирует С=С связь
- 135. 8 этап ЦТК: окисление малата до ОА(10 реакция), 4-я редокс-реакция, малат:НAД-оксидоредуктаза
- 136. Суммарное уравнение ЦТК: CH3CO~SKoA + ГДФ + Н3РО4 + 3НАД+ + ФАД + 2Н2О НSKoA +
- 137. Подсчёт энергетики ЦТК В ЦТК за 1 цикл сгорает 1 ацетил~КоА • при этом образуются 3
- 138. Роль дыхательной цепи МХ: синтез АТФ за счёт окисления Н до воды АТОМ ВОДОРОДА Н =
- 141. НАD+-зависимые дегидрогеназы (в форме НАDН+Н+) передают энергию в виде Н и ȇ на ФМН, забирая из
- 142. От КоQН2 ȇ по цепи цитохромов (в1/в2с1)→с→(аа3), постепенно расходуя энергию, поступают на конечный акцептор ½О2. Энергия
- 143. Энергия, высвобождаемая из ē, идёт на создание градиента Н+ путём их перекачки из матрикса в межмембранное
- 144. АТФ-синтаза расходует энергию создаваемого градиентом Н+ мито- хондриального потенциала (ΔμΗ = ΔΨ + ΔрН) на синтез
- 145. Разнообразие энергетических субстратов и ē-транспортная цепь МХ
- 146. Полное аэробное окисление глюкозы: «-» расход АТФ 2 АТФ на активацию глюкозы «+» синтез АТФ 4
- 147. Роль ЦТК 1) энергетическая ценность общий путь полного окисления топливных молекул – жирных кислот, углево-дов, аминокислот.
- 148. Регуляция ЦТК Скорость производства и утилизации энергии непостоянна в течение жизни клетки. Производ-ство энергии изменяется всегда,
- 149. При активной работе клетки в ней высокий уровень НАД+ и низкий НАДН, поскольку чем быстрее образу-ется
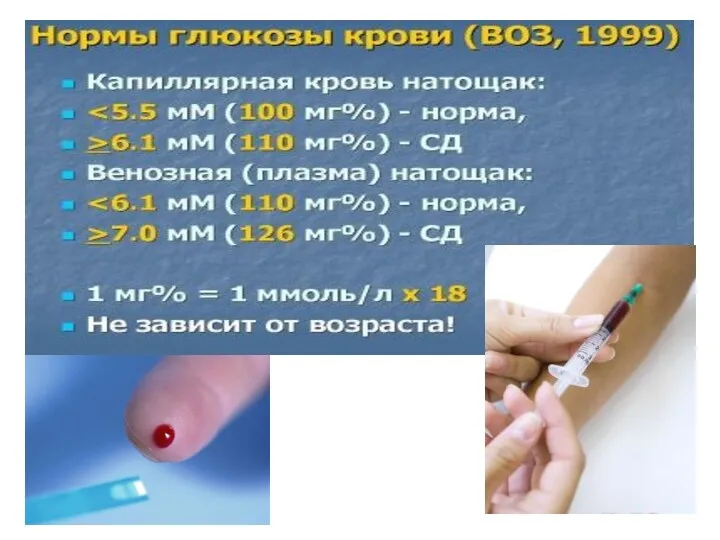
- 151. Регуляция уровня глюкозы крови
- 152. Нарушения углеводного обмена: голодание гипогликемия, глюкагон/адреналин мобилизуют ТАГ и ↑глюконеогенез из глицерола, СЖК идут на образование
- 153. Гипергликемия. После преодоления почечного порога присоединяется глюкозурия Снижен транспорт глюкозы в клетку (в том числе за
- 154. Если человек тебе сделал ЗЛО — ты дай ему конфетку, он тебе ЗЛО — ты ему
- 155. инсулинонезависимый сахарный диабет При длительном переедании и избыточном потреблении лёгких углеводов с возрастом постепенно ↓ способность
- 157. Сахарная кривая при нарушениях углеводного обмена ↓ Экспресс-анализ крови «на сахар» →
- 158. Экспресс-анализ мочи при сахарном диабете
- 160. Скачать презентацию

Углеводы – многоатомные альдегидоспирты или кетоспирты
Термин введён в 1876 г Шмидтом.
Углеводы – многоатомные альдегидоспирты или кетоспирты
Термин введён в 1876 г Шмидтом.

Биологическая роль углеводов
Энергетическая (распад→СО2 Н2О АТФ)
Пластическая (хондроитинсульфат)
Резервная (гликоген)
Биологическая роль углеводов
Энергетическая (распад→СО2 Н2О АТФ)
Пластическая (хондроитинсульфат)
Резервная (гликоген)

Классификация углеводов
В зависимости от сложности строения делят на 3 класса:
Классификация углеводов
В зависимости от сложности строения делят на 3 класса:


Моносахариды
МОНОСАХАРИД (МОНОЗА) – минимальная структурная единица углеводов, при дроблении которой
Моносахариды
МОНОСАХАРИД (МОНОЗА) – минимальная структурная единица углеводов, при дроблении которой
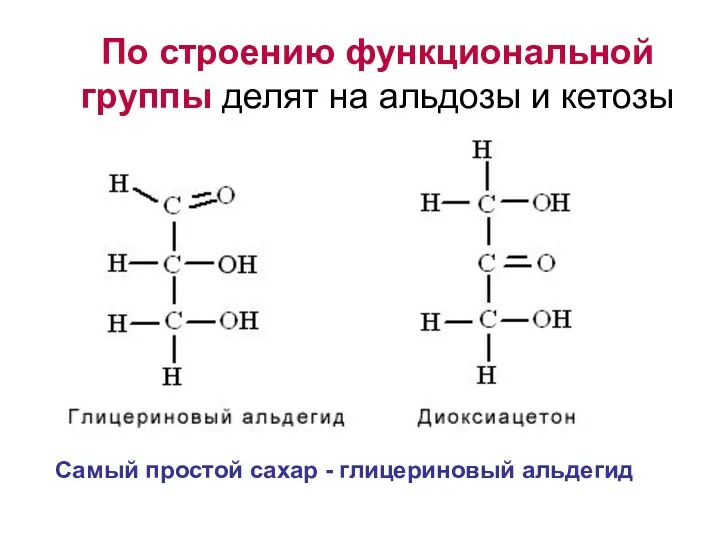
По строению функциональной группы делят на альдозы и кетозы
Самый простой сахар
По строению функциональной группы делят на альдозы и кетозы
Самый простой сахар
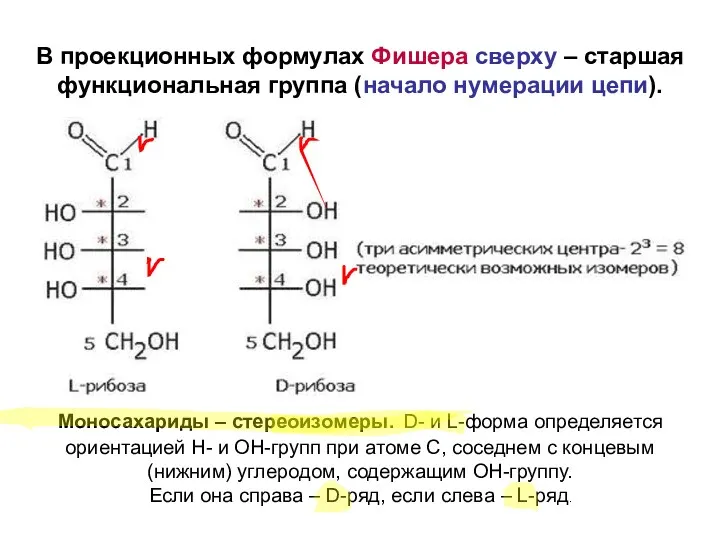
Моносахариды – стереоизомеры. D- и L-форма определяется ориентацией Н- и ОН-групп
Моносахариды – стереоизомеры. D- и L-форма определяется ориентацией Н- и ОН-групп

Большинство моносахаридов у млекопитающих имеет D-конфигурацию. Именно к ней специфичны фермен-ты,
Большинство моносахаридов у млекопитающих имеет D-конфигурацию. Именно к ней специфичны фермен-ты,
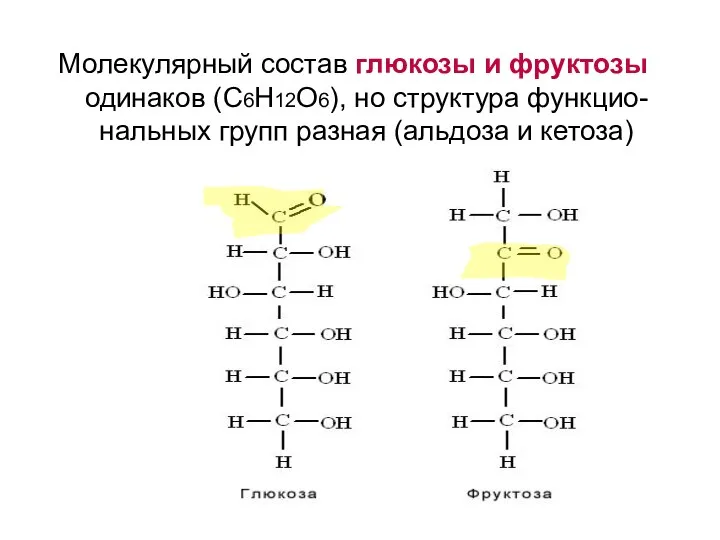
Молекулярный состав глюкозы и фруктозы одинаков (С6Н12О6), но структура функцио-нальных групп
Молекулярный состав глюкозы и фруктозы одинаков (С6Н12О6), но структура функцио-нальных групп
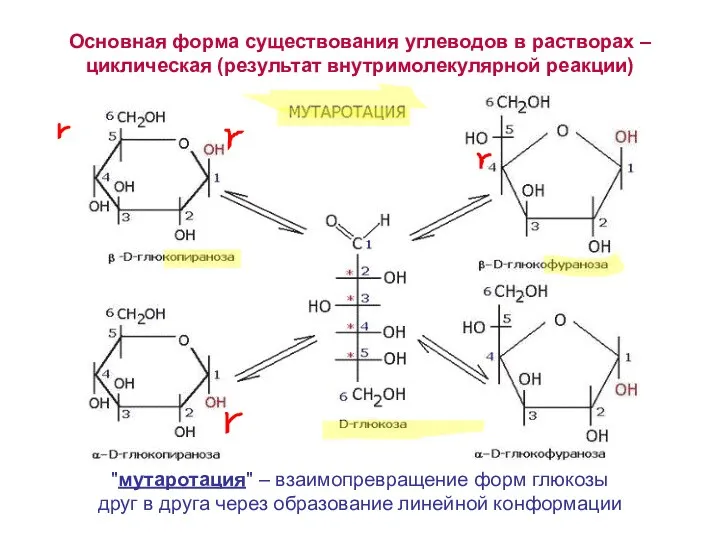
Основная форма существования углеводов в растворах – циклическая (результат внутримолекулярной реакции)
"мутаротация"
Основная форма существования углеводов в растворах – циклическая (результат внутримолекулярной реакции)
"мутаротация"

Циклические формы моносахаридов приобретают активные ОН-группы при С1 или С2 или
Циклические формы моносахаридов приобретают активные ОН-группы при С1 или С2 или
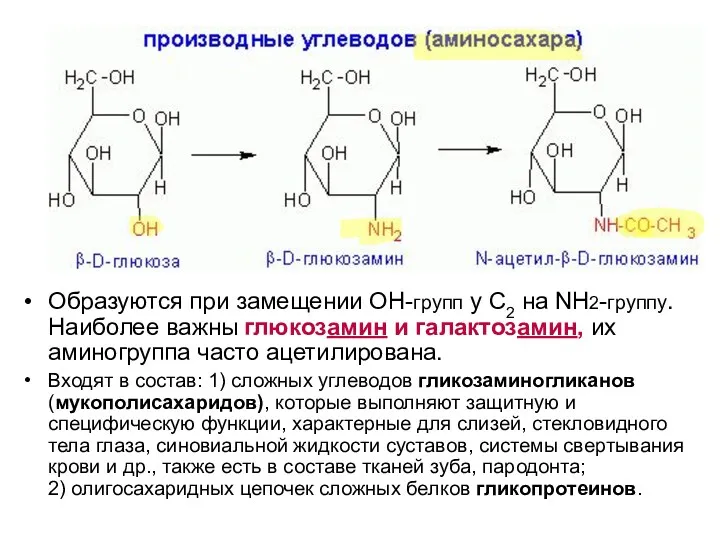
Образуются при замещении ОН-групп у С2 на NH2-группу. Наиболее важны глюкозамин
Образуются при замещении ОН-групп у С2 на NH2-группу. Наиболее важны глюкозамин
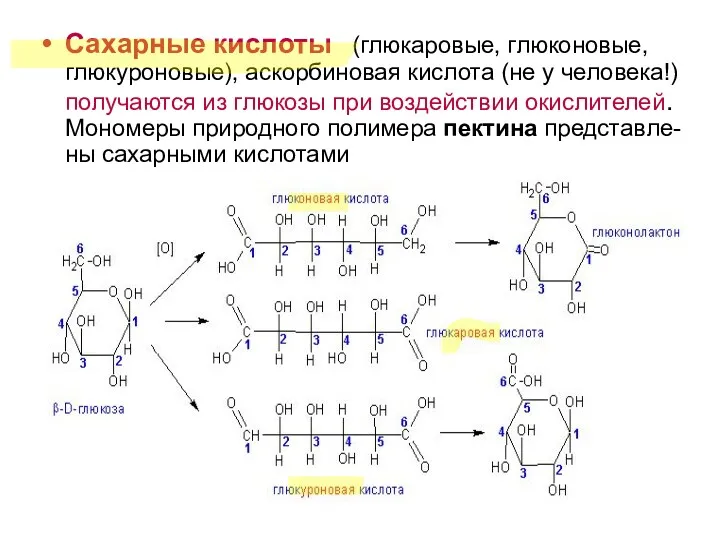
Сахарные кислоты (глюкаровые, глюконовые, глюкуроновые), аскорбиновая кислота (не у человека!) получаются
Сахарные кислоты (глюкаровые, глюконовые, глюкуроновые), аскорбиновая кислота (не у человека!) получаются
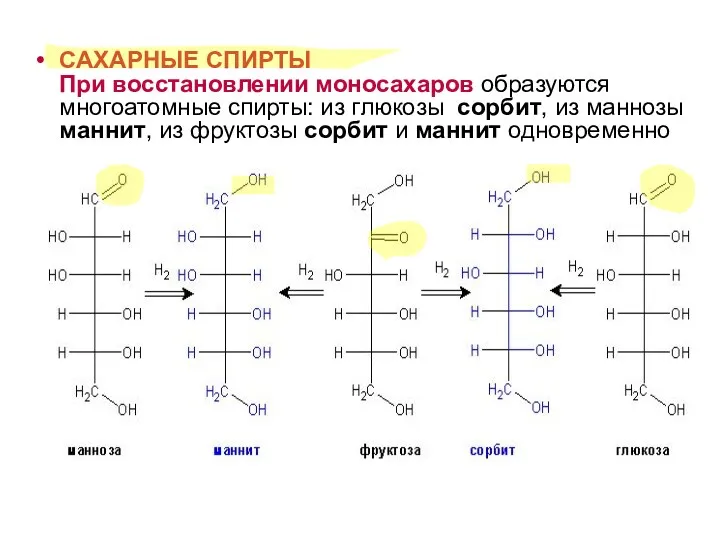
САХАРНЫЕ СПИРТЫ
При восстановлении моносахаров образуются многоатомные спирты: из глюкозы сорбит,
САХАРНЫЕ СПИРТЫ При восстановлении моносахаров образуются многоатомные спирты: из глюкозы сорбит,

ОЛИГОСАХАРИДЫ
включают от 2 до 10 остатков моносахаридов, соединены 1,4- или
ОЛИГОСАХАРИДЫ
включают от 2 до 10 остатков моносахаридов, соединены 1,4- или
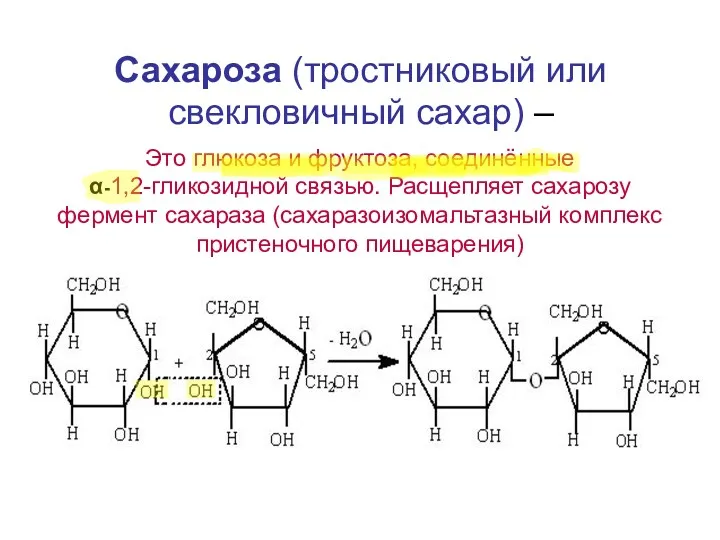
Сахароза (тростниковый или свекловичный сахар) –
Это глюкоза и фруктоза, соединённые
Сахароза (тростниковый или свекловичный сахар) –
Это глюкоза и фруктоза, соединённые
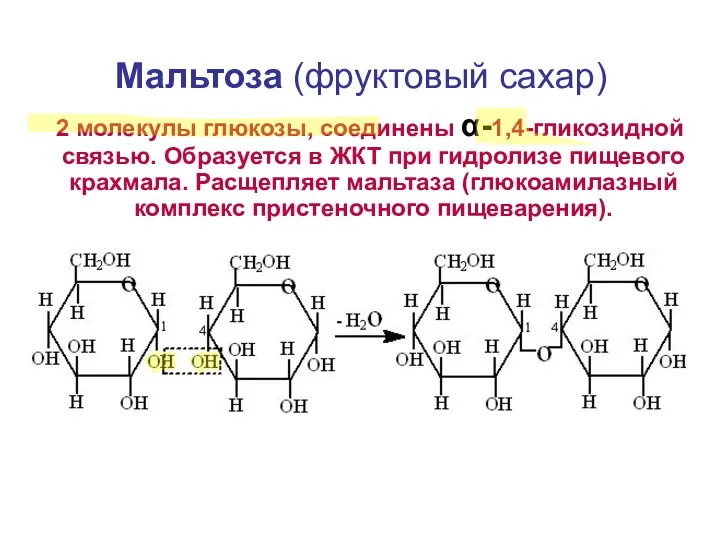
Мальтоза (фруктовый сахар)
2 молекулы глюкозы, соединены α-1,4-гликозидной связью. Образуется в
Мальтоза (фруктовый сахар)
2 молекулы глюкозы, соединены α-1,4-гликозидной связью. Образуется в
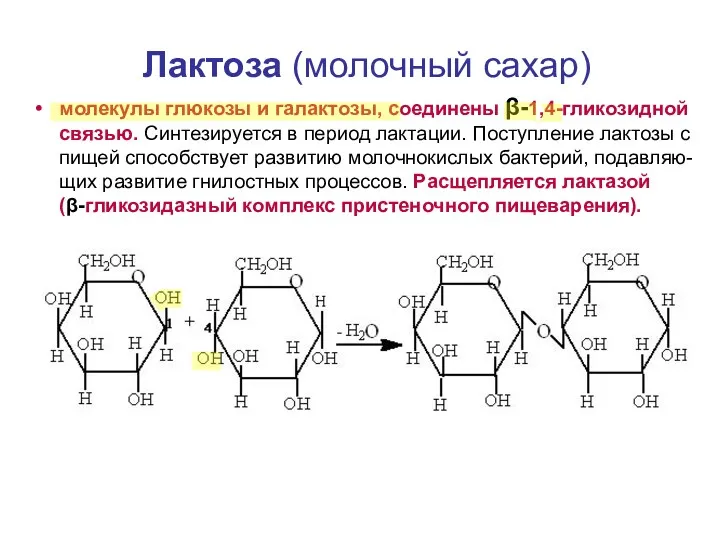
Лактоза (молочный сахар)
молекулы глюкозы и галактозы, соединены β-1,4-гликозидной связью. Синтезируется в
Лактоза (молочный сахар)
молекулы глюкозы и галактозы, соединены β-1,4-гликозидной связью. Синтезируется в

ПОЛИСАХАРИДЫ
Большинство природных углеводов – полимеры, число моносахаридных остатков от 10 до
ПОЛИСАХАРИДЫ
Большинство природных углеводов – полимеры, число моносахаридных остатков от 10 до

По составу: гомо-, гетерополисахариды
Гомополисахариды (гомогликаны) состоят из моносахаридных
По составу: гомо-, гетерополисахариды
Гомополисахариды (гомогликаны) состоят из моносахаридных
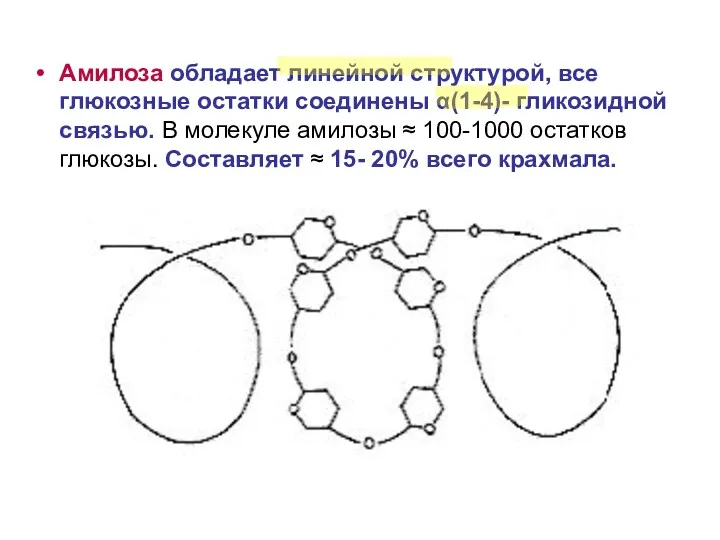
Амилоза обладает линейной структурой, все глюкозные остатки соединены α(1-4)- гликозидной связью.
Амилоза обладает линейной структурой, все глюкозные остатки соединены α(1-4)- гликозидной связью.
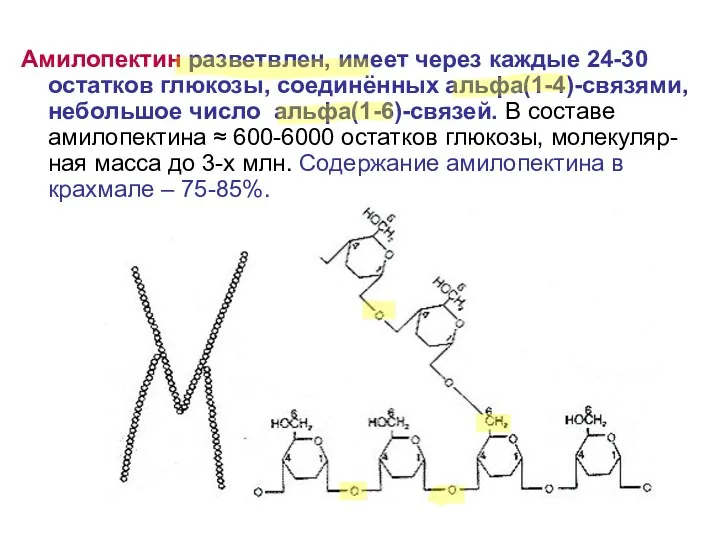
Амилопектин разветвлен, имеет через каждые 24-30 остатков глюкозы, соединённых альфа(1-4)-связями, небольшое
Амилопектин разветвлен, имеет через каждые 24-30 остатков глюкозы, соединённых альфа(1-4)-связями, небольшое
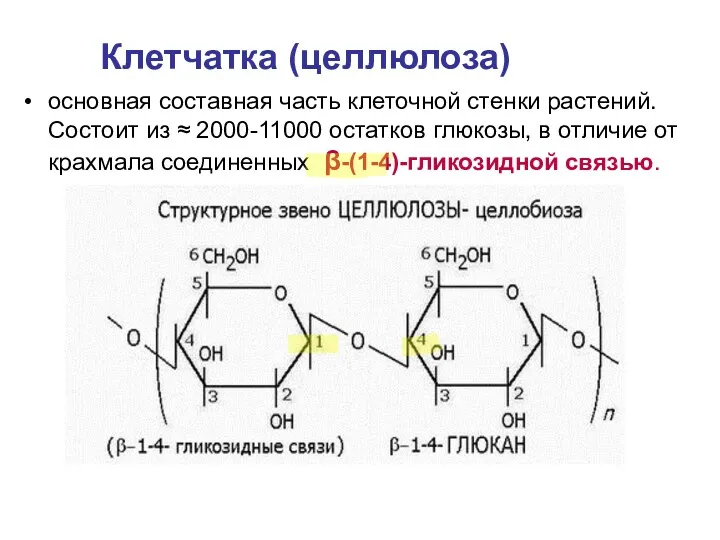
основная составная часть клеточной стенки растений. Состоит из ≈ 2000-11000 остатков
основная составная часть клеточной стенки растений. Состоит из ≈ 2000-11000 остатков
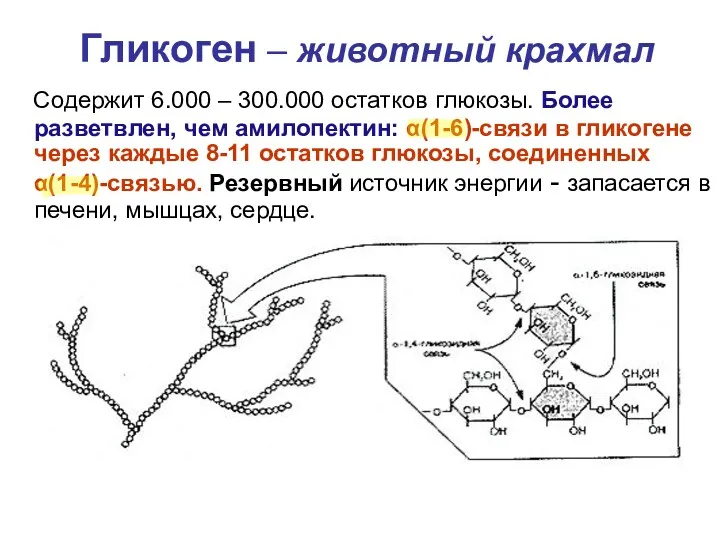
Гликоген – животный крахмал
Содержит 6.000 – 300.000 остатков глюкозы. Более
Гликоген – животный крахмал
Содержит 6.000 – 300.000 остатков глюкозы. Более

Другие резервные полисахариды
Бактерии ротовой полости способны расщеплять сахарозу: сахароза →глюкоза
Другие резервные полисахариды
Бактерии ротовой полости способны расщеплять сахарозу: сахароза →глюкоза
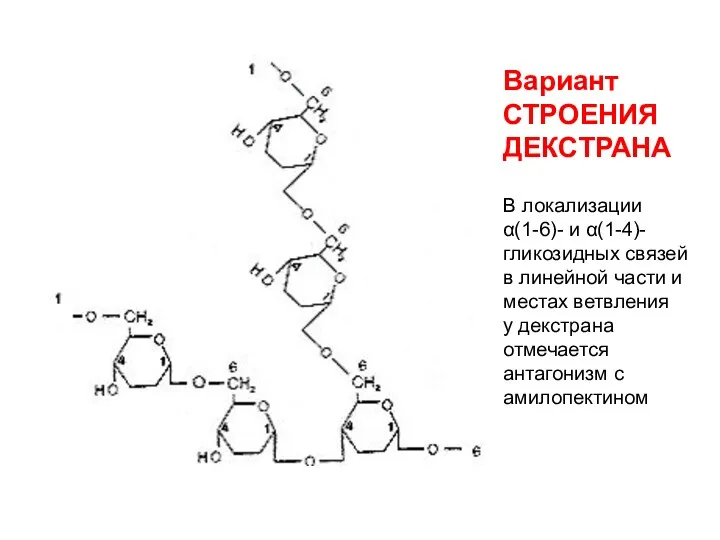
Вариант
СТРОЕНИЯ ДЕКСТРАНА
В локализации
α(1-6)- и α(1-4)-
гликозидных связей
в линейной части и
Вариант СТРОЕНИЯ ДЕКСТРАНА В локализации α(1-6)- и α(1-4)- гликозидных связей в линейной части и

2) Леваны – резервные гомополисахариды из остатков D-фруктозы, т.е. являются фруктанами
2) Леваны – резервные гомополисахариды из остатков D-фруктозы, т.е. являются фруктанами
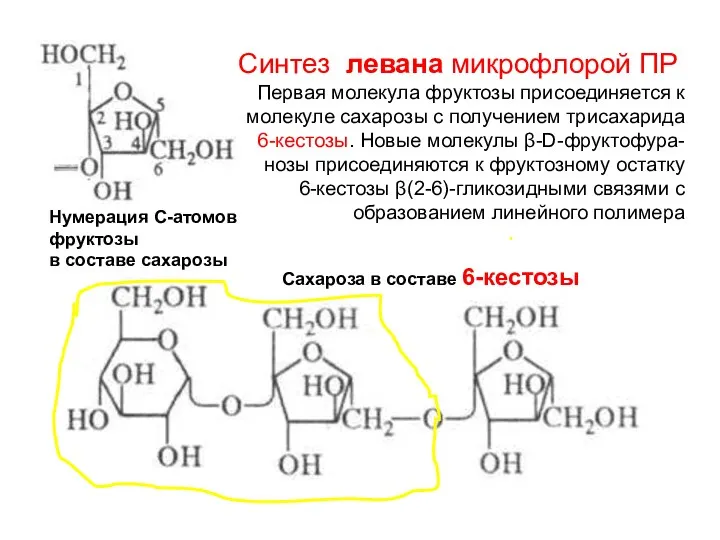
Нумерация С-атомов
фруктозы
в составе сахарозы
Синтез левана микрофлорой ПР
Первая молекула
Нумерация С-атомов
фруктозы
в составе сахарозы
Синтез левана микрофлорой ПР
Первая молекула

Гетерополисахариды (гетерогликаны)
сложные углеводы, состоят из 2-х и более типов моносахаридов
Гетерополисахариды (гетерогликаны)
сложные углеводы, состоят из 2-х и более типов моносахаридов
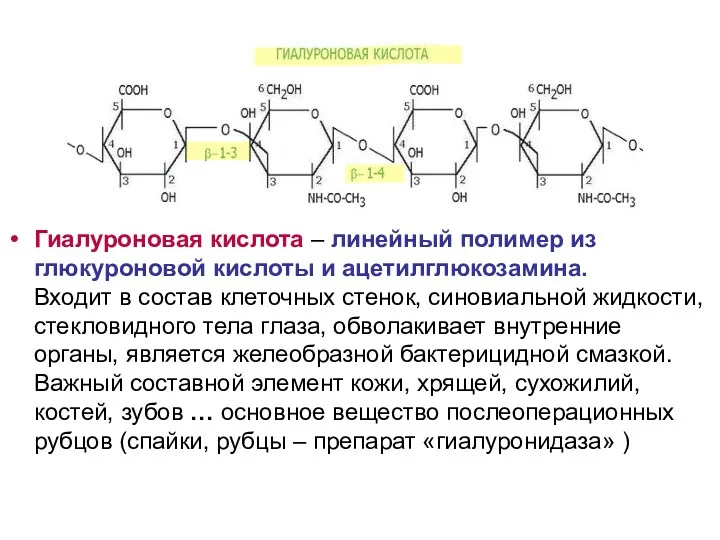
Гиалуроновая кислота – линейный полимер из глюкуроновой кислоты и ацетилглюкозамина.
Входит
Гиалуроновая кислота – линейный полимер из глюкуроновой кислоты и ацетилглюкозамина. Входит
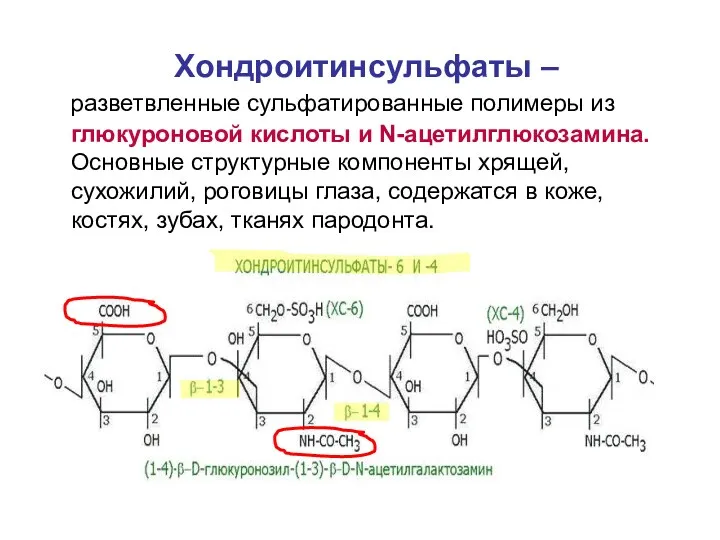
Хондроитинсульфаты –
разветвленные сульфатированные полимеры из глюкуроновой кислоты и N-ацетилглюкозамина. Основные
Хондроитинсульфаты –
разветвленные сульфатированные полимеры из глюкуроновой кислоты и N-ацетилглюкозамина. Основные

Гликопротеины и гликолипиды
1) Комплексы белка с углеводами, существует 2 вида: гликопротеины
Гликопротеины и гликолипиды
1) Комплексы белка с углеводами, существует 2 вида: гликопротеины

Норма углеводов в питании
Запас углеводов в организме не превышает 2-3%
Норма углеводов в питании
Запас углеводов в организме не превышает 2-3%

ВНЕШНИЙ ОБМЕН УГЛЕВОДОВ (переваривание)
Переваривание – совокупность процессов ферментативного гидролиза поступающих
ВНЕШНИЙ ОБМЕН УГЛЕВОДОВ (переваривание)
Переваривание – совокупность процессов ферментативного гидролиза поступающих
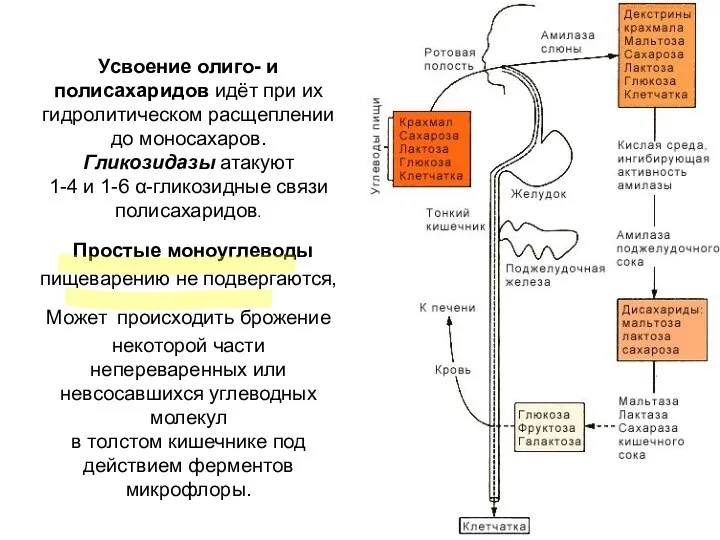
Усвоение олиго- и полисахаридов идёт при их гидролитическом расщеплении до моносахаров.
Усвоение олиго- и полисахаридов идёт при их гидролитическом расщеплении до моносахаров.

ПОЛОСТНОЕ ПИЩЕВАРЕНИЕ
Переваривание полисахаридов начинается
в ротовой полости, где крахмалы подвергаются хаотичному
ПОЛОСТНОЕ ПИЩЕВАРЕНИЕ
Переваривание полисахаридов начинается
в ротовой полости, где крахмалы подвергаются хаотичному

Далее пища идёт в 12-ПК и кишечник (значения рН нейтральные)
Далее пища идёт в 12-ПК и кишечник (значения рН нейтральные)

ПРИСТЕНОЧНОЕ ПИЩЕВАРЕНИЕ
Гидролиз дисахаридов происходит не в просвете кишечника, а на поверхности
ПРИСТЕНОЧНОЕ ПИЩЕВАРЕНИЕ
Гидролиз дисахаридов происходит не в просвете кишечника, а на поверхности
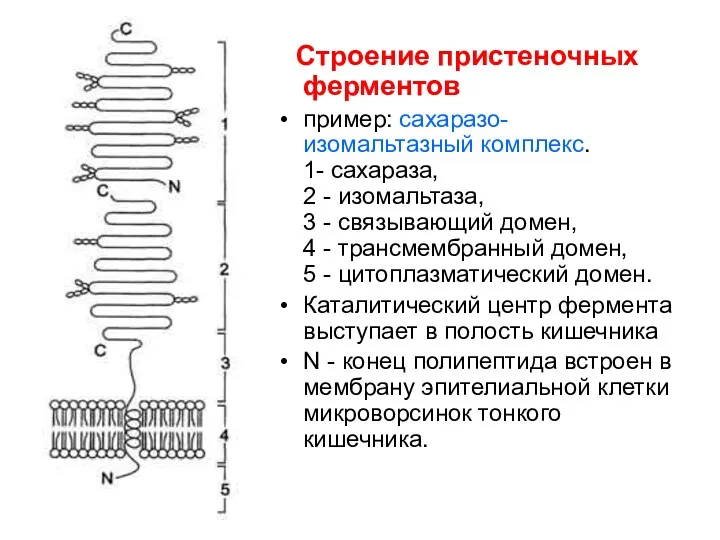
Строение пристеночных ферментов
пример: сахаразо-изомальтазный комплекс.
1- сахараза,
2 - изомальтаза,
Строение пристеночных ферментов
пример: сахаразо-изомальтазный комплекс.
1- сахараза,
2 - изомальтаза,

НЕПЕРЕНОСИМОСТЬ УГЛЕВОДОВ
Лактоза поступает в организм с молоком.
У ребёнка при недостаточной активности
НЕПЕРЕНОСИМОСТЬ УГЛЕВОДОВ
Лактоза поступает в организм с молоком.
У ребёнка при недостаточной активности

Лечение непереносимости углеводов.
Диета с исключением из рациона:
в первом случае –
Лечение непереносимости углеводов. Диета с исключением из рациона: в первом случае –

Целлюлоза в организме человека
У человека нет ферментов для расщепления
β(1-4)-гликозидной
Целлюлоза в организме человека
У человека нет ферментов для расщепления
β(1-4)-гликозидной

Всасываются в кишечнике только моносахара (глюкоза,галактоза,фруктоза)
Их перенос в энтероциты происходит:
1)
Всасываются в кишечнике только моносахара (глюкоза,галактоза,фруктоза)
Их перенос в энтероциты происходит: 1)
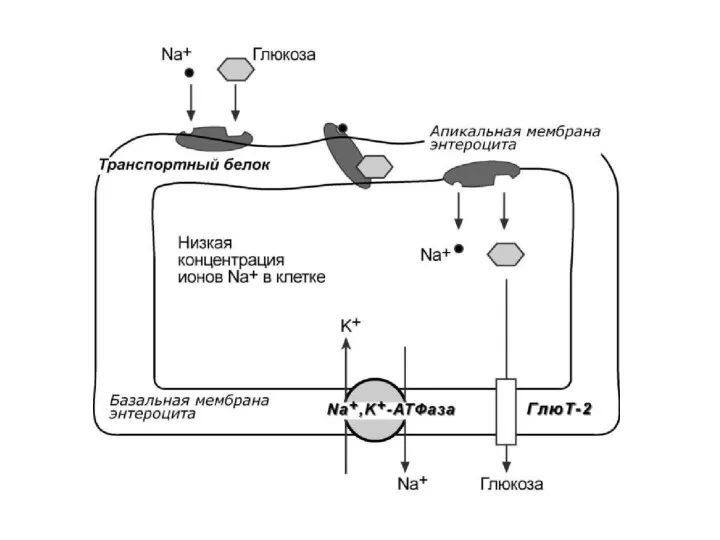
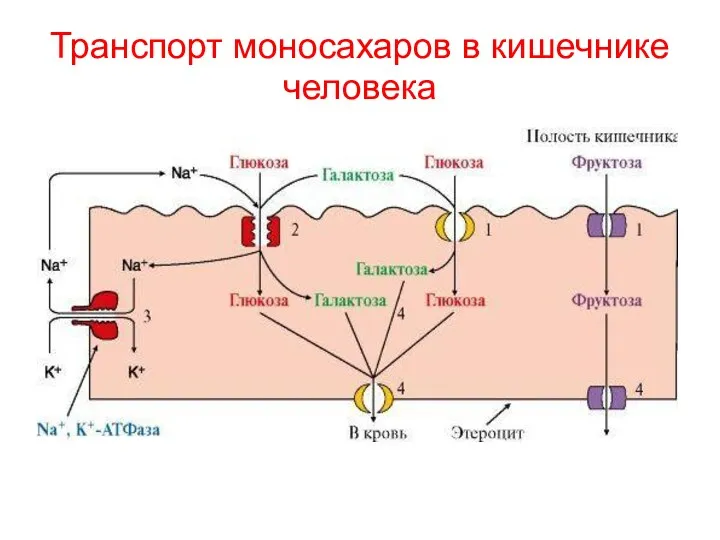
Транспорт моносахаров в кишечнике человека
Транспорт моносахаров в кишечнике человека

ДАЛЬНЕЙШИЕ ПРЕВРАЩЕНИЯ САХАРОВ В ОРГАНИЗМЕ
В результате переваривания углево-дов и частичной инверсии
ДАЛЬНЕЙШИЕ ПРЕВРАЩЕНИЯ САХАРОВ В ОРГАНИЗМЕ
В результате переваривания углево-дов и частичной инверсии
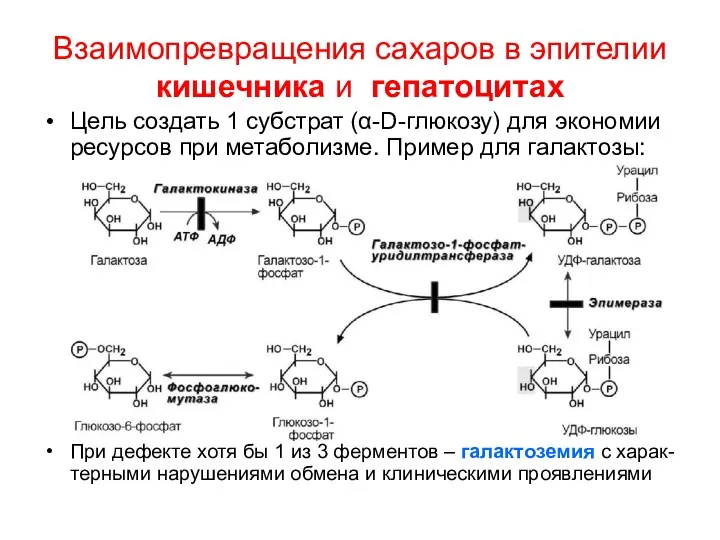
Взаимопревращения сахаров в эпителии кишечника и гепатоцитах
Цель создать 1 субстрат (α-D-глюкозу)
Взаимопревращения сахаров в эпителии кишечника и гепатоцитах
Цель создать 1 субстрат (α-D-глюкозу)
После печени в общий кровоток поступает только
После печени в общий кровоток
После печени в общий кровоток поступает только
После печени в общий кровоток

Роль печени в обмене глюкозы
- Регуляция уровня глюкозы в крови
Роль печени в обмене глюкозы
- Регуляция уровня глюкозы в крови

Если поступление глюкозы в гепатоциты выше их возможностей полимеризовать глюкозу
Если поступление глюкозы в гепатоциты выше их возможностей полимеризовать глюкозу

Транспорт глюкозы из крови в клетки
1) Пассивная диффузия, т.е.
Транспорт глюкозы из крови в клетки
1) Пассивная диффузия, т.е.

Облегчённая диффузия (ГлюТ)
Этот вид транспорта особо важен
для
Облегчённая диффузия (ГлюТ)
Этот вид транспорта особо важен
для

Влияние инсулина на транспортёры глюкозы (белки-переносчики "ГлюТ")
Различают 12 типов транспортеров
Влияние инсулина на транспортёры глюкозы (белки-переносчики "ГлюТ")
Различают 12 типов транспортеров

Облегчённая диффузия имеет принципиальное значение для мышц и жировой ткани,
Облегчённая диффузия имеет принципиальное значение для мышц и жировой ткани,

Дальнейшая судьба глюкозы в организме
В клетке глюкоза прежде всего фосфорили-руется, в
Дальнейшая судьба глюкозы в организме
В клетке глюкоза прежде всего фосфорили-руется, в
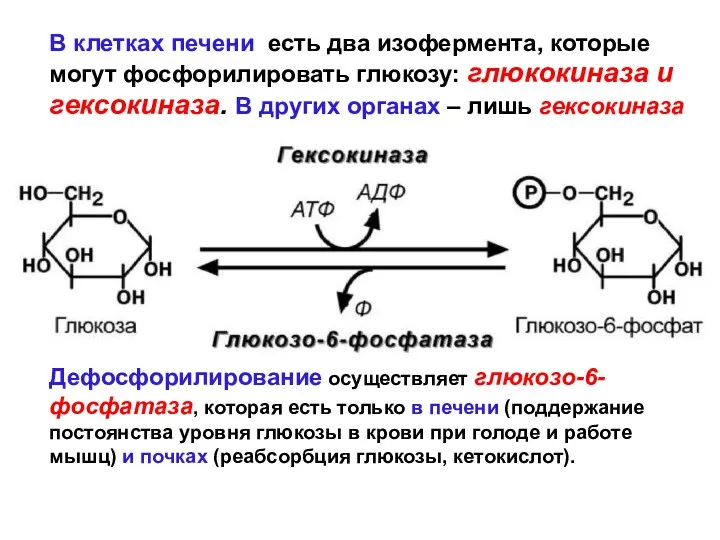
В клетках печени есть два изофермента, которые могут фосфорилировать глюкозу: глюкокиназа
В клетках печени есть два изофермента, которые могут фосфорилировать глюкозу: глюкокиназа
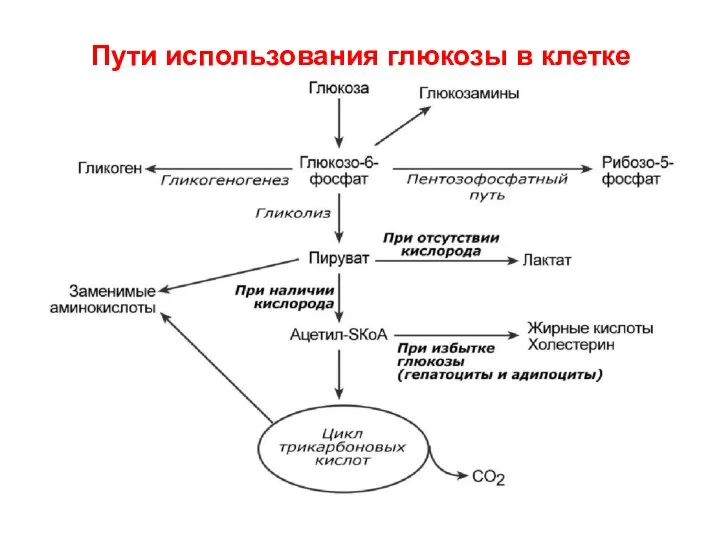
Пути использования глюкозы в клетке
Пути использования глюкозы в клетке

Если в клетке образовался избыток Глюкозо-6-Ф, она начинает запасать глюкозу
Если в клетке образовался избыток Глюкозо-6-Ф, она начинает запасать глюкозу
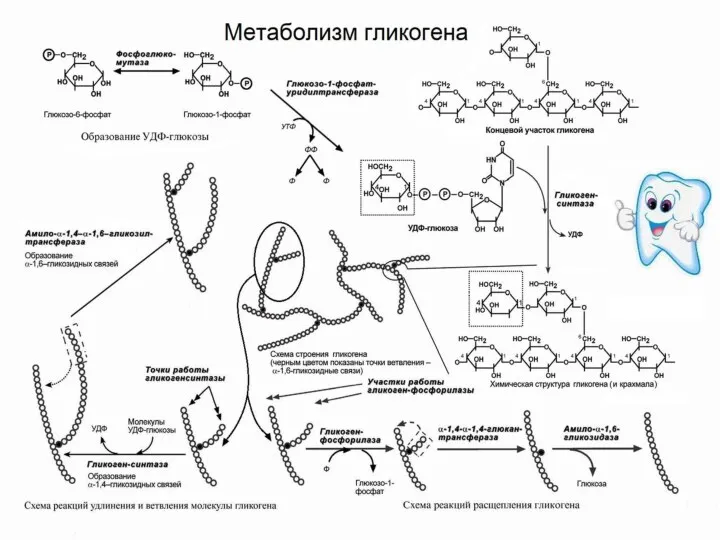

Гликоген образуется почти во всех клетках орга-низма, но max его
Гликоген образуется почти во всех клетках орга-низма, но max его

ГЛИКОГЕНОЛИЗ
Распад гликогена до молочной кислоты
может идти при недостатке кислорода.
Гликоген присутствует
ГЛИКОГЕНОЛИЗ
Распад гликогена до молочной кислоты
может идти при недостатке кислорода.
Гликоген присутствует
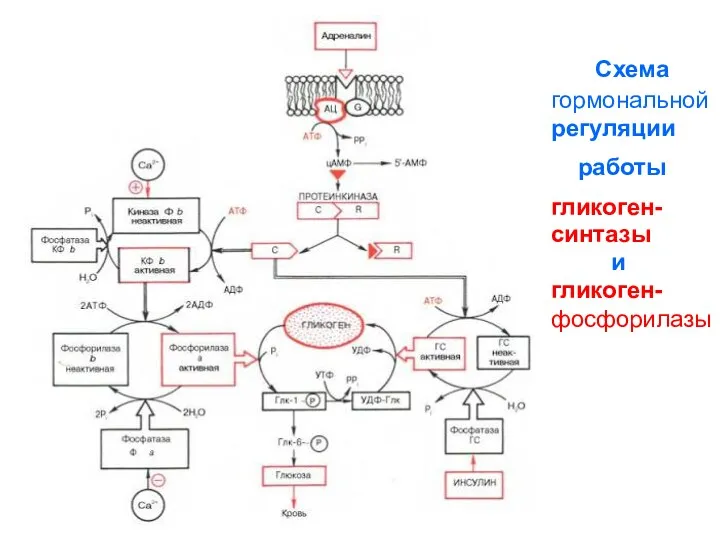
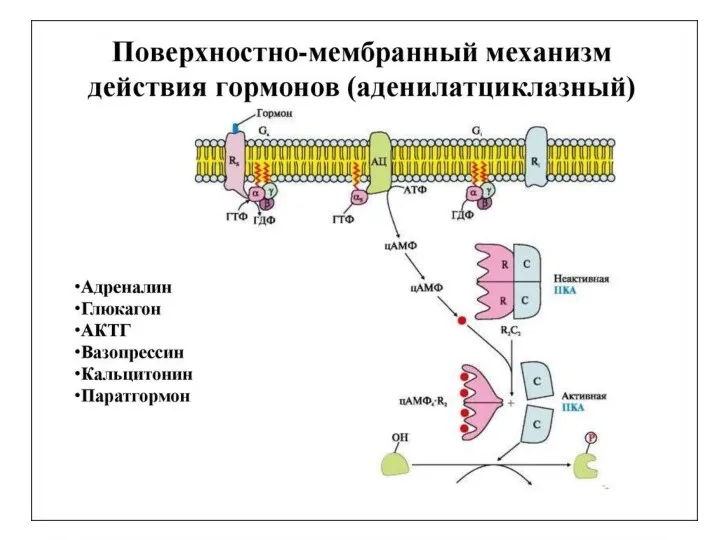
Схема
гормональной
регуляции
работы
гликоген-
синтазы
и
гликоген-
фосфорилазы
Схема
гормональной
регуляции
работы
гликоген-
синтазы
и
гликоген-
фосфорилазы
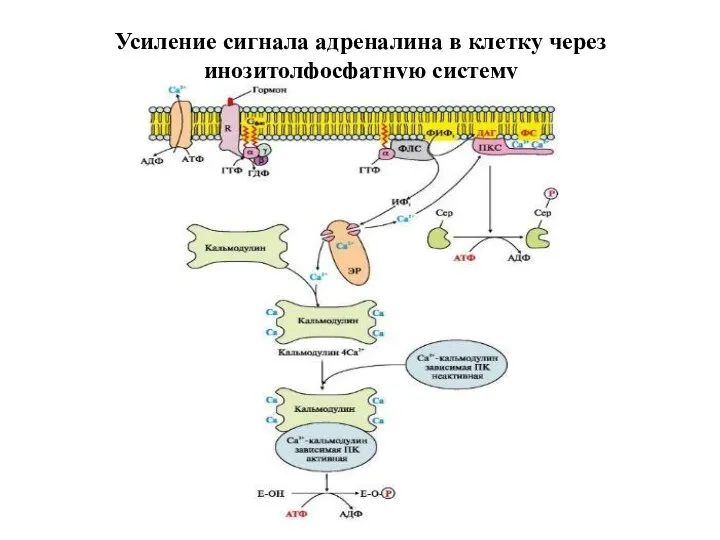
Усиление сигнала адреналина в клетку через инозитолфосфатную систему
Усиление сигнала адреналина в клетку через инозитолфосфатную систему

Распад гликогена регулируют гормоны – адреналин, глюкагон, глюкокортикоиды.
Инсулин оказывает
Распад гликогена регулируют гормоны – адреналин, глюкагон, глюкокортикоиды. Инсулин оказывает

цАМФ в клетке существует недолго. Фосфоди-эстераза быстро превращает его в
цАМФ в клетке существует недолго. Фосфоди-эстераза быстро превращает его в

В тот момент, когда "включается" фосфорилаза, одновременно происходит "выключение" гликоген-синтазы.
Это имеет
Это имеет

Обратное превращение фосфорилазы а в неактивную b-форму происходит при
Обратное превращение фосфорилазы а в неактивную b-форму происходит при

Отщепившаяся от гликогена молекула глюкозо-1-Ф изомеризуется
с образованием глюкозо-6-Ф
глюкозо-1-Ф
глюкозо-6-Ф
-
Отщепившаяся от гликогена молекула глюкозо-1-Ф изомеризуется
с образованием глюкозо-6-Ф
глюкозо-1-Ф
глюкозо-6-Ф
-

Гликогенозы – болезни гликогена
При наследственной недостаточности или
Гликогенозы – болезни гликогена
При наследственной недостаточности или

АНАЭРОБНЫЕ
ПРЕВРАЩЕНИЯ УГЛЕВОДОВ
1) гликогеногенез
2) гликогенолиз
3) гликолиз
4) глюконеогенез
5)
АНАЭРОБНЫЕ ПРЕВРАЩЕНИЯ УГЛЕВОДОВ 1) гликогеногенез 2) гликогенолиз 3) гликолиз 4) глюконеогенез 5)

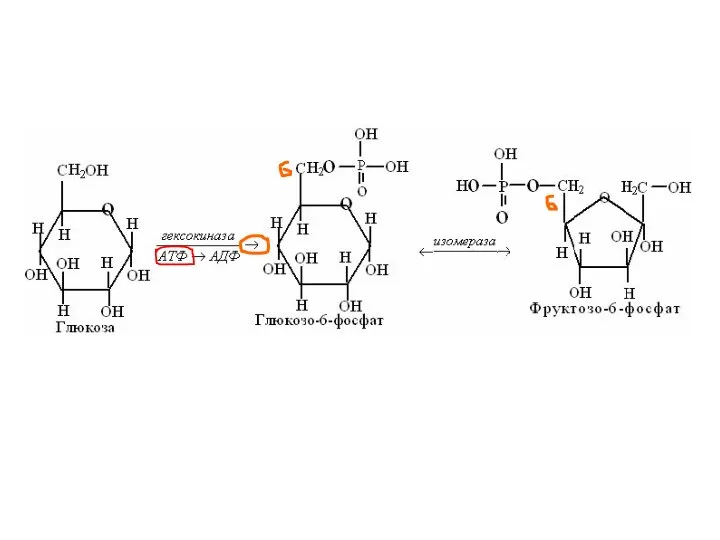
ГЛИКОЛИЗ
- главный анаэробный путь утилизации глюкозы
1) Протекает во
ГЛИКОЛИЗ
- главный анаэробный путь утилизации глюкозы
1) Протекает во
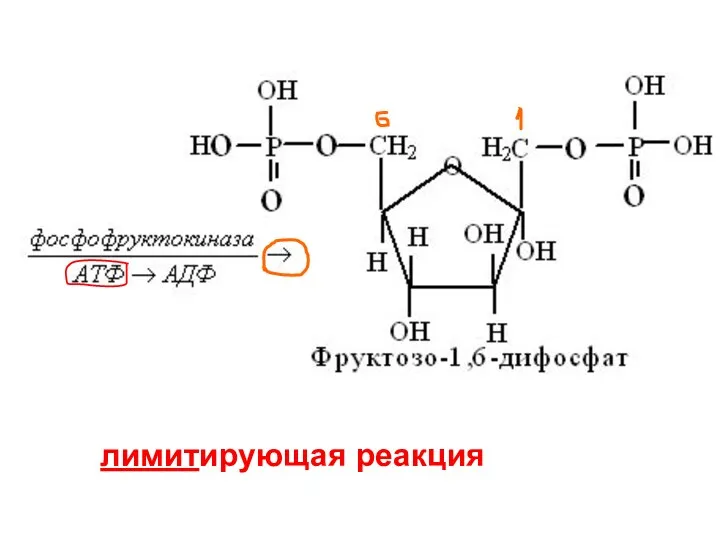
лимитирующая реакция
лимитирующая реакция

фосфофруктокиназа
Весь ход гликолиза лимитируется уровнем активности этого фермента
Это
фосфофруктокиназа
Весь ход гликолиза лимитируется уровнем активности этого фермента
Это
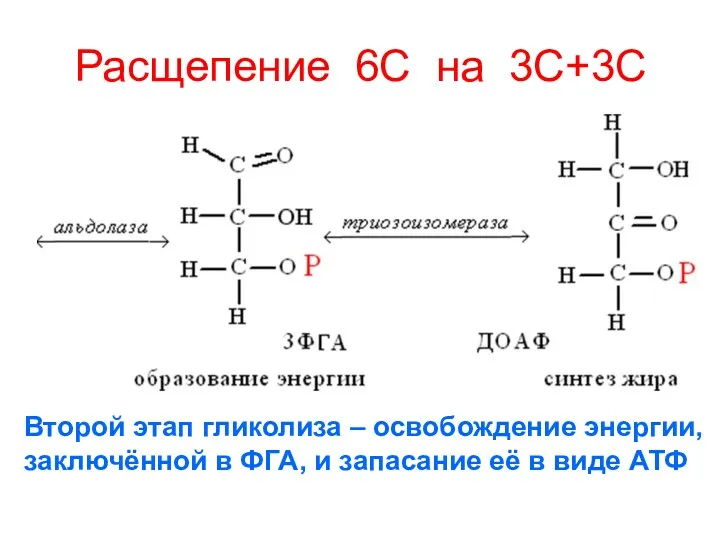
Расщепение 6С на 3С+3С
Второй этап гликолиза – освобождение энергии,
заключённой в
Расщепение 6С на 3С+3С
Второй этап гликолиза – освобождение энергии, заключённой в
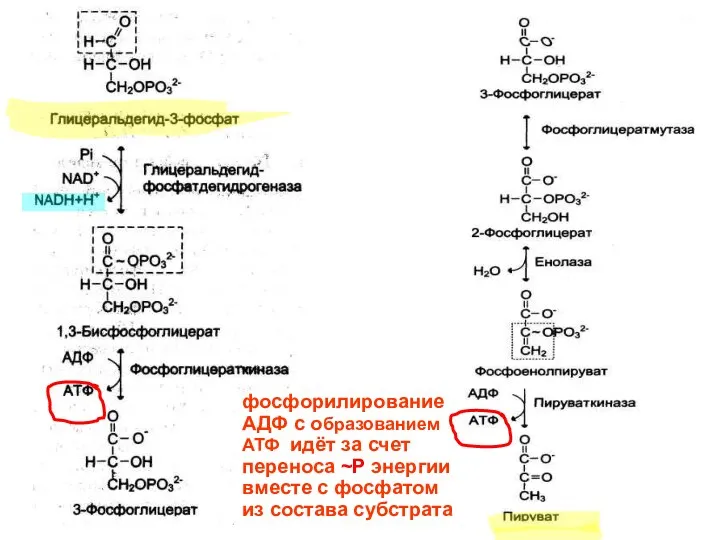
фосфорилирование АДФ с образованием АТФ идёт за счет переноса ~Р энергии
фосфорилирование АДФ с образованием АТФ идёт за счет переноса ~Р энергии
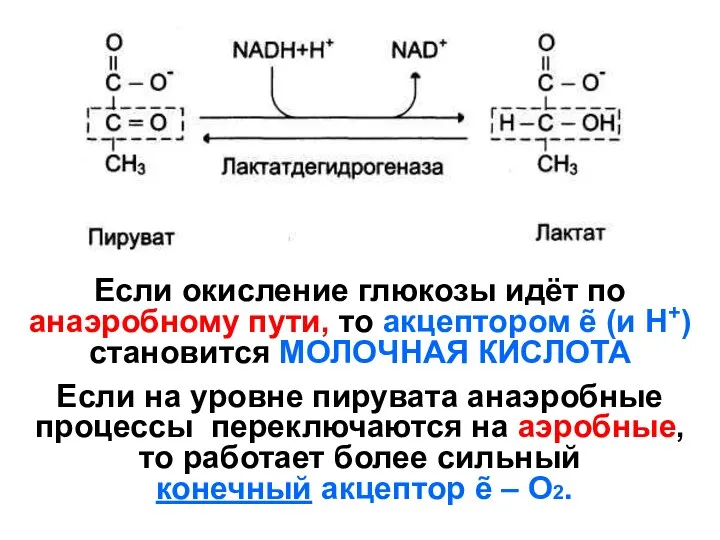
Если окисление глюкозы идёт по анаэробному пути, то акцептором ẽ (и
Если окисление глюкозы идёт по анаэробному пути, то акцептором ẽ (и

Энергетический баланс гликолиза
С6Н12О6 (глюкоза) → 2 С3Н6О3 (лактат)
1) расход по 1
Энергетический баланс гликолиза
С6Н12О6 (глюкоза) → 2 С3Н6О3 (лактат)
1) расход по 1

ЭНЕРГЕТИЧЕСКИЙ БАЛАНС гликогенолиза
4АТФ – 1АТФ = 3АТФ
На 1 молекулу
ЭНЕРГЕТИЧЕСКИЙ БАЛАНС гликогенолиза
4АТФ – 1АТФ = 3АТФ
На 1 молекулу

Спиртовое БРОЖЕНИЕ
Микроорганизмы, в первую очередь дрожжи, осуществляют не молочно-кислое, а спиртовое
Спиртовое БРОЖЕНИЕ
Микроорганизмы, в первую очередь дрожжи, осуществляют не молочно-кислое, а спиртовое

2 СН3СОСООН
ПИРУВАТ
2 СН3СОН
ацетальдегид
Энергетический баланс спиртового
брожения как
2 СН3СОСООН
ПИРУВАТ
2 СН3СОН
ацетальдегид
Энергетический баланс спиртового брожения как

Глюконеогенез
Глюконеогенез – синтез глюкозы
de novo из неуглеводных компонентов.
Протекает
Глюконеогенез
Глюконеогенез – синтез глюкозы de novo из неуглеводных компонентов. Протекает
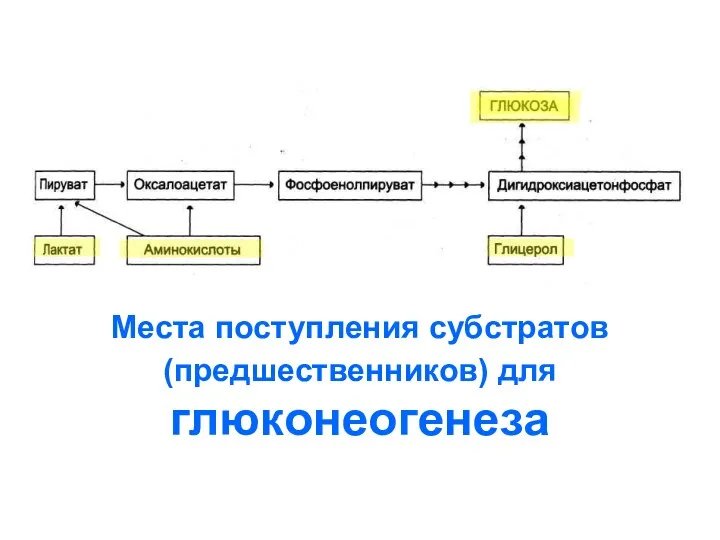
Места поступления субстратов (предшественников) для
глюконеогенеза
Места поступления субстратов (предшественников) для
глюконеогенеза
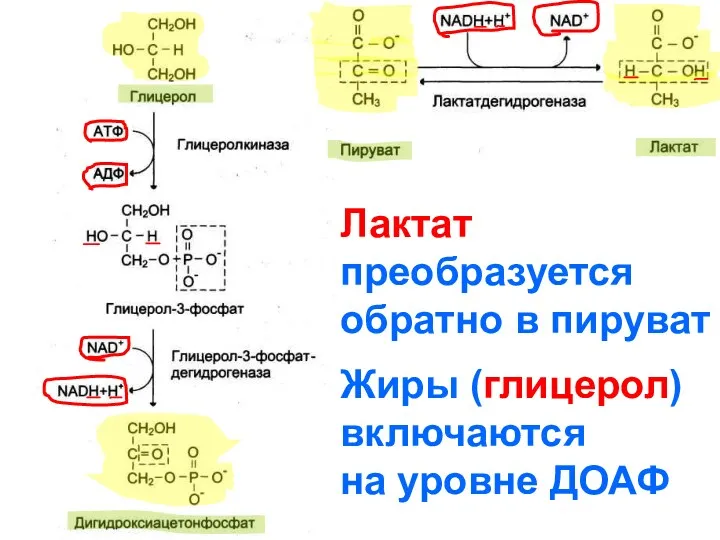
Лактат
преобразуется
обратно в пируват
Жиры (глицерол)
включаются
на уровне ДОАФ
Лактат
преобразуется
обратно в пируват
Жиры (глицерол)
включаются
на уровне ДОАФ
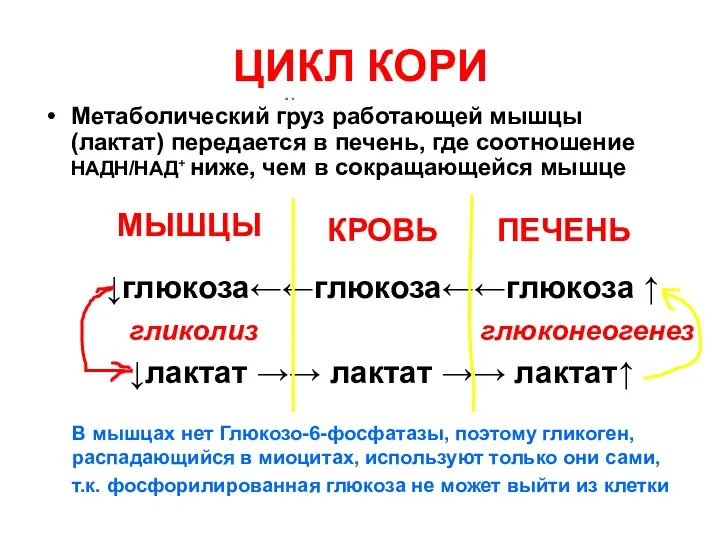
ЦИКЛ КОРИ
Метаболический груз работающей мышцы (лактат) передается в печень, где соотношение
ЦИКЛ КОРИ
Метаболический груз работающей мышцы (лактат) передается в печень, где соотношение

Источники глюконеогенеза - только
гликогенные аминокислоты
В пируват – ала, сер,
Источники глюконеогенеза - только
гликогенные аминокислоты
В пируват – ала, сер,
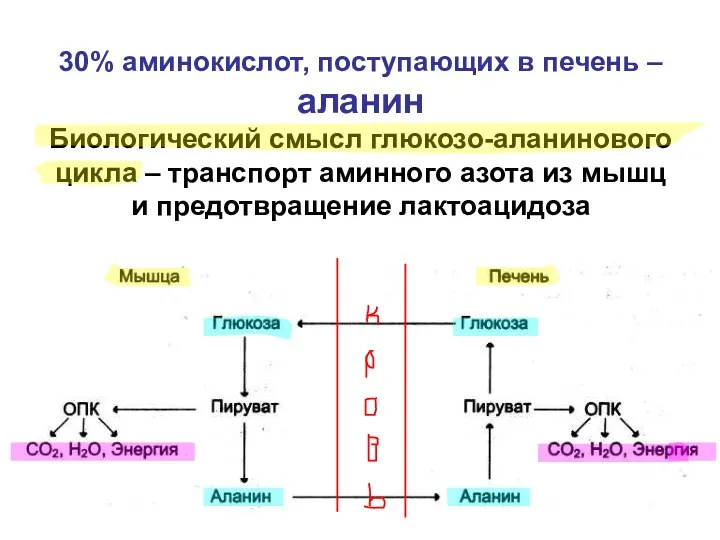
30% аминокислот, поступающих в печень – аланин
Биологический смысл глюкозо-аланинового цикла –
30% аминокислот, поступающих в печень – аланин Биологический смысл глюкозо-аланинового цикла –
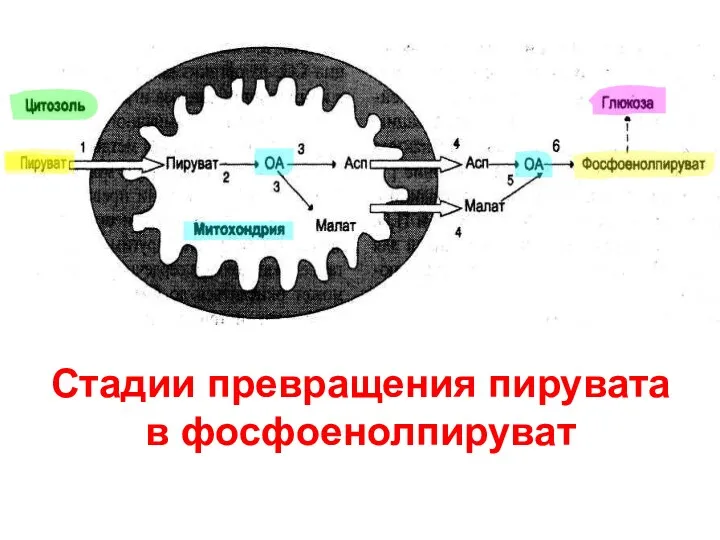
Стадии превращения пирувата
в фосфоенолпируват
Стадии превращения пирувата
в фосфоенолпируват
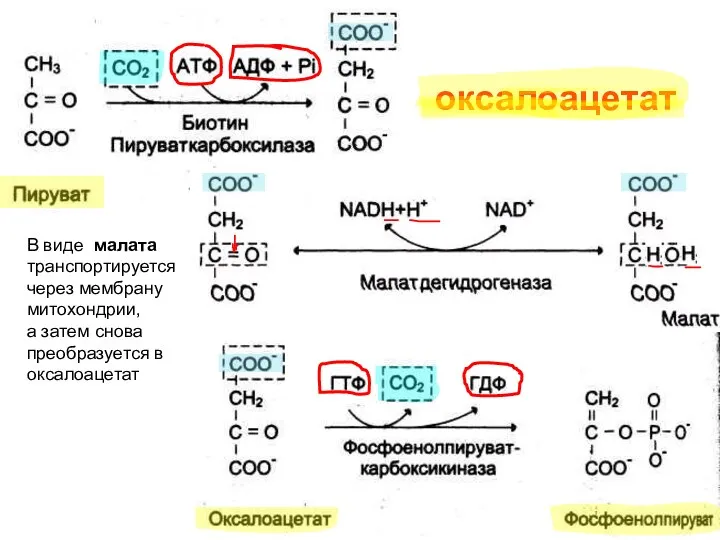
оксалоацетат
В виде малата
транспортируется
через мембрану
митохондрии,
а затем снова преобразуется в оксалоацетат
оксалоацетат
В виде малата
транспортируется
через мембрану
митохондрии,
а затем снова преобразуется в оксалоацетат
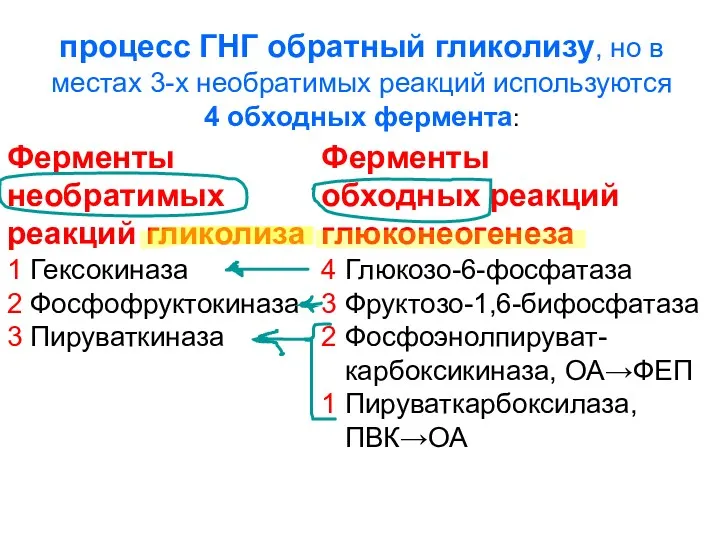
процесс ГНГ обратный гликолизу, но в местах 3-х необратимых реакций используются
процесс ГНГ обратный гликолизу, но в местах 3-х необратимых реакций используются
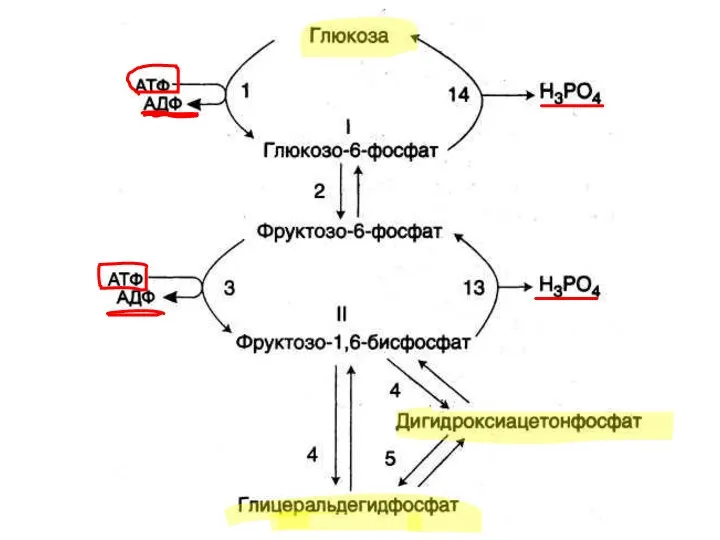

Синтез глюкозы
в результате глюконеогенеза энергозависим
При синтезе 1 глюкозы
Синтез глюкозы
в результате глюконеогенеза энергозависим
При синтезе 1 глюкозы

АЭРОБНЫЕ ПРОЦЕССЫ
углеводного обмена
Прямой – пентозофосфатный путь,
Непрямой – переключение гликолиза
на
АЭРОБНЫЕ ПРОЦЕССЫ
углеводного обмена
Прямой – пентозофосфатный путь,
Непрямой – переключение гликолиза
на
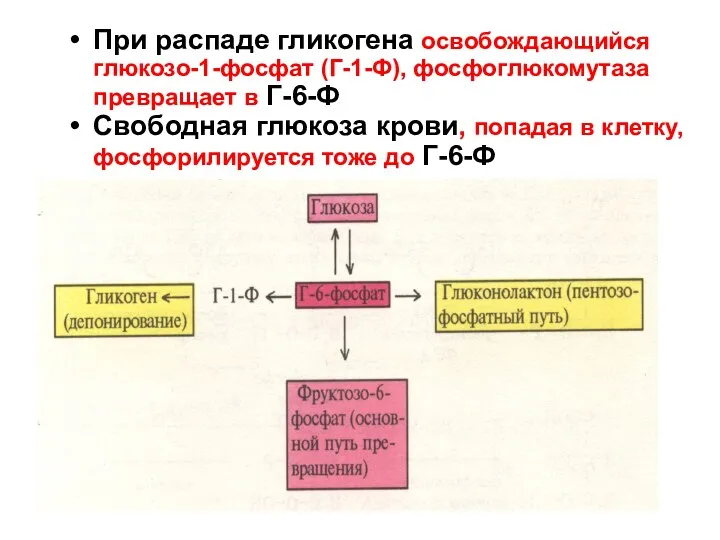
При распаде гликогена освобождающийся
глюкозо-1-фосфат (Г-1-Ф), фосфоглюкомутаза
превращает в Г-6-Ф
Свободная
Свободная

Пути использования глюкозо-6-Ф
1) Гл-6-Ф может распадаться в процессе гликолиза и служить
Пути использования глюкозо-6-Ф
1) Гл-6-Ф может распадаться в процессе гликолиза и служить

Пентозофосфатный путь
Другие названия:
пентозный шунт (или цикл)
гексозомонофосфатный путь
Пентозофосфатный путь
Другие названия:
пентозный шунт (или цикл)
гексозомонофосфатный путь

Роль ПФП: синтез пентоз и НАДФН
в процессе окисления глюкозы.
Возникает качественно
Роль ПФП: синтез пентоз и НАДФН
в процессе окисления глюкозы.
Возникает качественно
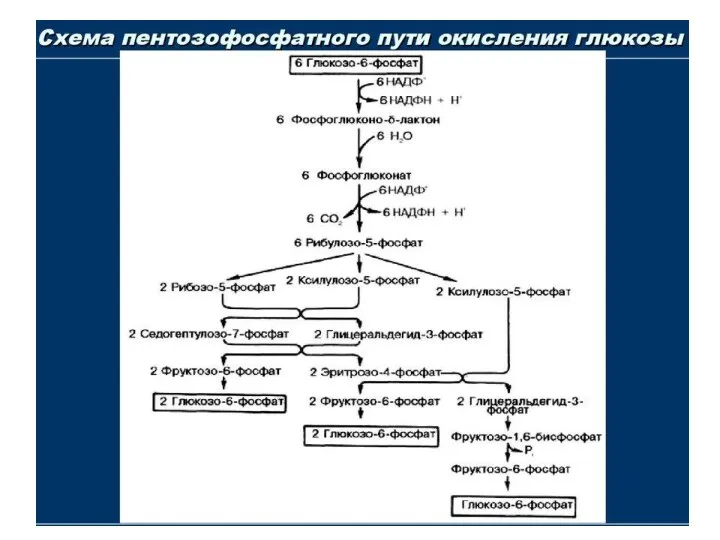

ПФП состоит из 2 стадий(ветвей)
I-я стадия окислительная, её роль:
1)
ПФП состоит из 2 стадий(ветвей)
I-я стадия окислительная, её роль:
1)
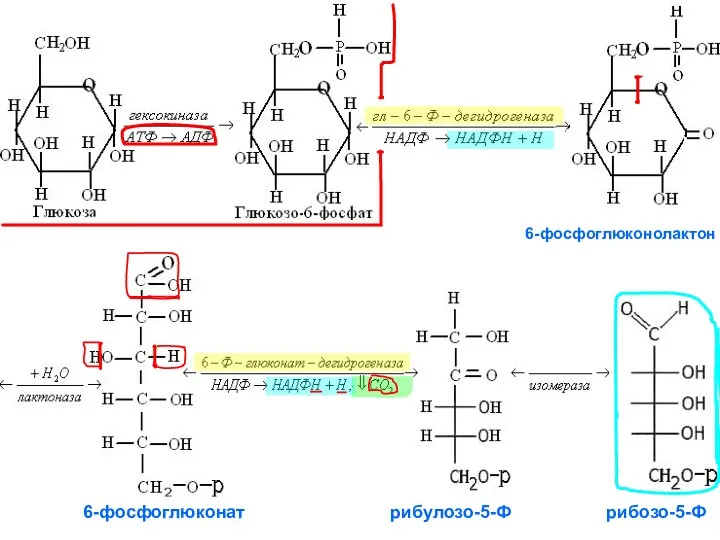
6-фосфоглюконолактон
6-фосфоглюконат рибулозо-5-Ф рибозо-5-Ф
6-фосфоглюконолактон
6-фосфоглюконат рибулозо-5-Ф рибозо-5-Ф

2-я стадия НЕОКИСЛИТЕЛЬНАЯ :
1) идёт перенос (ферменты - трансферазы)
2-углеродного компонента
2-я стадия НЕОКИСЛИТЕЛЬНАЯ :
1) идёт перенос (ферменты - трансферазы)
2-углеродного компонента

ПЕНТОЗОФОСФАТНЫЙ ПУТЬ
Конечный продукт перестроек неокислительной ветви – глюкозо-6-Ф
ПЕНТОЗОФОСФАТНЫЙ ПУТЬ
Конечный продукт перестроек неокислительной ветви – глюкозо-6-Ф

Для синтезов клеткам нужно много НАДФН,
а рибозы для включения
Для синтезов клеткам нужно много НАДФН, а рибозы для включения

Суммарные реакции ПФП
I-я стадия (с образованием рибозы):
Глюкозо-6-Ф + 2НАДФ ↓
Суммарные реакции ПФП
I-я стадия (с образованием рибозы): Глюкозо-6-Ф + 2НАДФ ↓
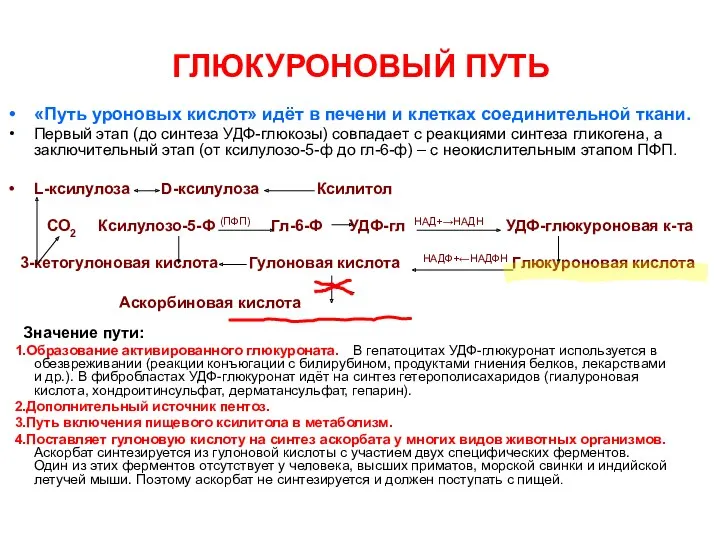
ГЛЮКУРОНОВЫЙ ПУТЬ
«Путь уроновых кислот» идёт в печени и клетках соединительной ткани.
ГЛЮКУРОНОВЫЙ ПУТЬ
«Путь уроновых кислот» идёт в печени и клетках соединительной ткани.

Генетическая недостаточность
глюкозо-6Ф-дегидрогеназы - причина
лекарственных гемолитических анемий
У этих людей
Генетическая недостаточность
глюкозо-6Ф-дегидрогеназы - причина
лекарственных гемолитических анемий
У этих людей

Синдром Вернике-Корсакова
У таких людей генетически снижено сродство транскетолазы к витамину В1
Синдром Вернике-Корсакова
У таких людей генетически снижено сродство транскетолазы к витамину В1
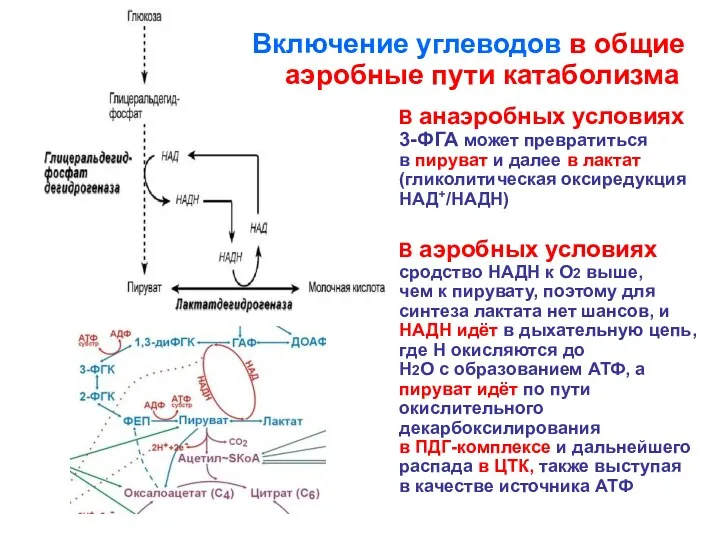
В анаэробных условиях
3-ФГА может превратиться
в пируват и далее в
В анаэробных условиях 3-ФГА может превратиться в пируват и далее в

Пируватдегидрогеназный комплекс
3 фермента:
пируватдегидрогеназа, дегидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа
5 коферментов:
В1 - тиаминпирофосфат(ТПФ,кокарбоксилаза)
Пируватдегидрогеназный комплекс
3 фермента:
пируватдегидрогеназа, дегидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа
5 коферментов:
В1 - тиаминпирофосфат(ТПФ,кокарбоксилаза)
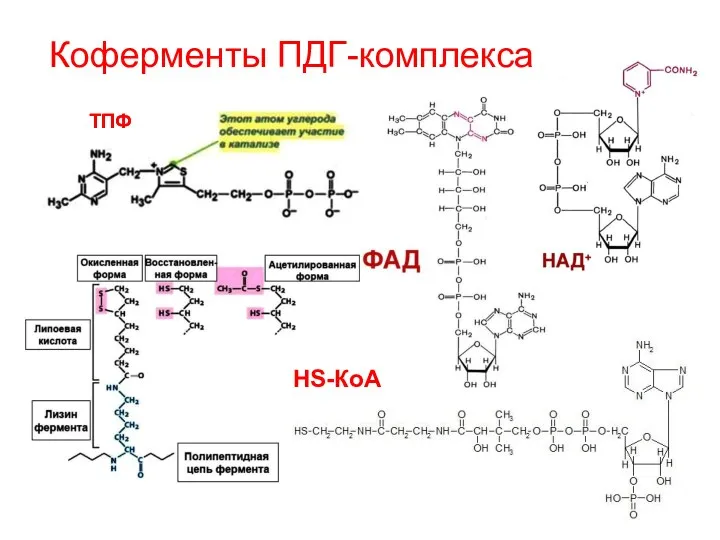
Коферменты ПДГ-комплекса
ТПФ
ТПФ
HS-КоА
Коферменты ПДГ-комплекса
ТПФ
ТПФ
HS-КоА
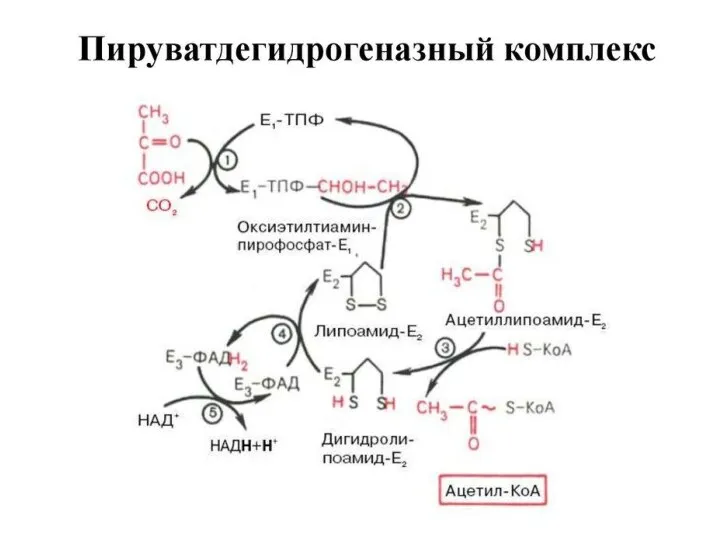
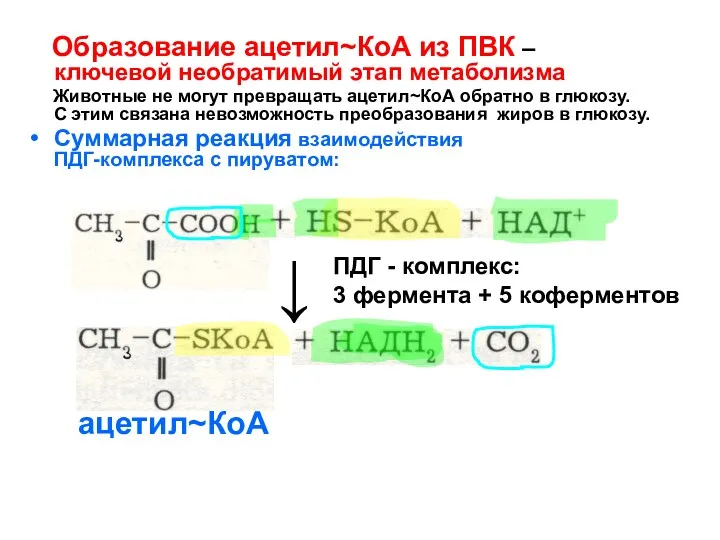
Образование ацетил~КоА из ПВК –
ключевой необратимый этап метаболизма
Образование ацетил~КоА из ПВК –
ключевой необратимый этап метаболизма

регуляция активности ПДГ-комплекса
1. Ингибирование продуктами реакции.
Ацетил~КоА и НАДН (продукты
регуляция активности ПДГ-комплекса
1. Ингибирование продуктами реакции. Ацетил~КоА и НАДН (продукты

Биологическая роль окислительного декарбоксилирования ПВК
определяется тем, какой будет дальней-шая
Биологическая роль окислительного декарбоксилирования ПВК определяется тем, какой будет дальней-шая
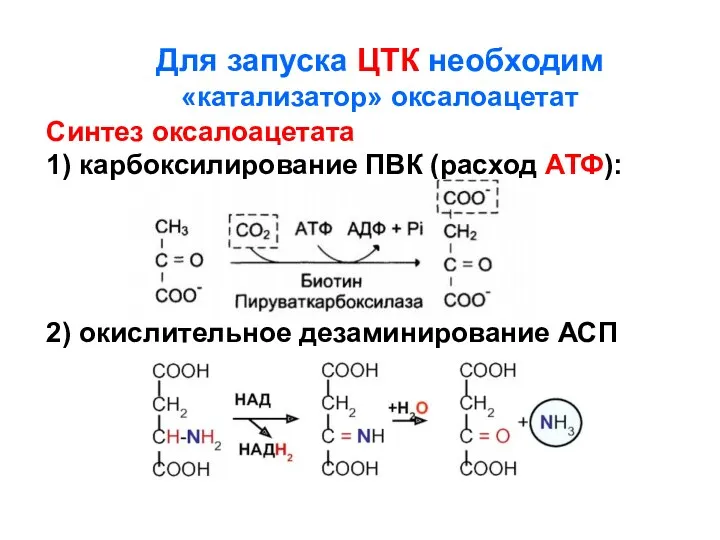
Для запуска ЦТК необходим
«катализатор» оксалоацетат
Синтез оксалоацетата
1) карбоксилирование ПВК (расход АТФ):
2)
Для запуска ЦТК необходим
«катализатор» оксалоацетат
Синтез оксалоацетата
1) карбоксилирование ПВК (расход АТФ):
2)

Цикл трикарбоновых кислот
(=цикл лимонной кислоты,
=цикл Кребса)
Ганс Кребс – лауреат Нобелевской
Цикл трикарбоновых кислот
(=цикл лимонной кислоты,
=цикл Кребса)
Ганс Кребс – лауреат Нобелевской
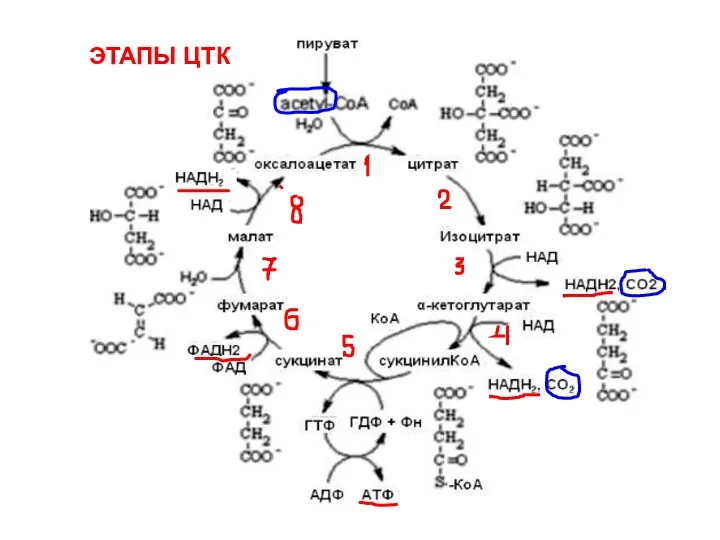
ЭТАПЫ ЦТК
ЭТАПЫ ЦТК
ЭТАПЫ ЦТК
ЭТАПЫ ЦТК
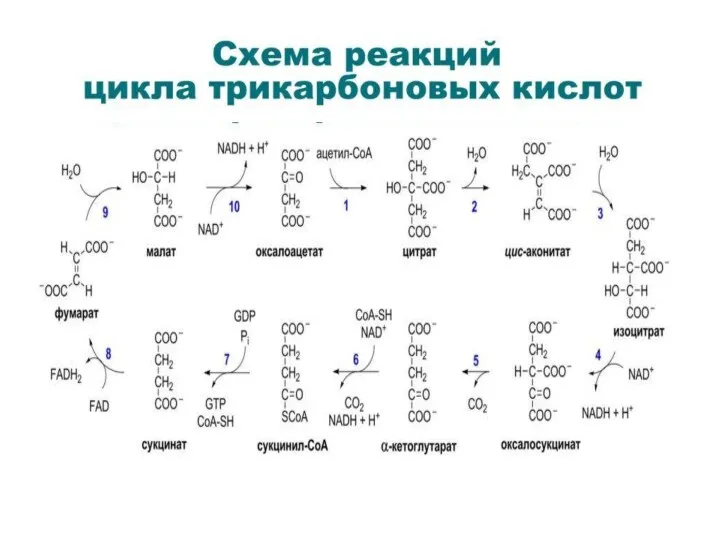
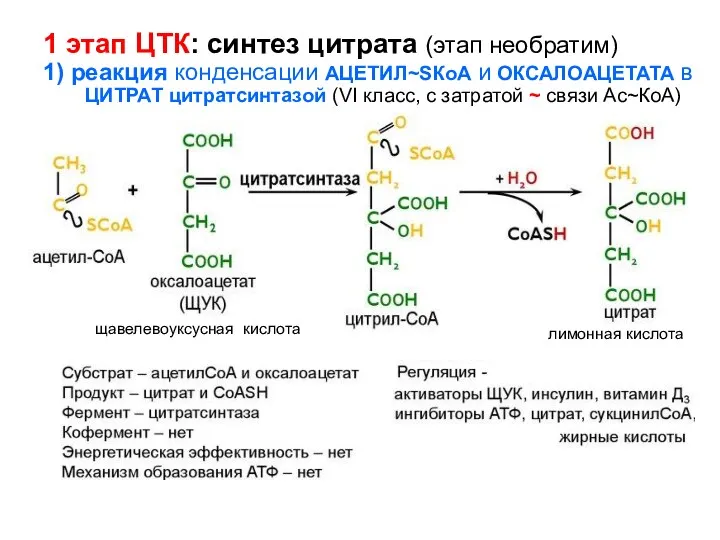
1 этап ЦТК: синтез цитрата (этап необратим)
1) реакция конденсации АЦЕТИЛ~SКоА
1 этап ЦТК: синтез цитрата (этап необратим)
1) реакция конденсации АЦЕТИЛ~SКоА
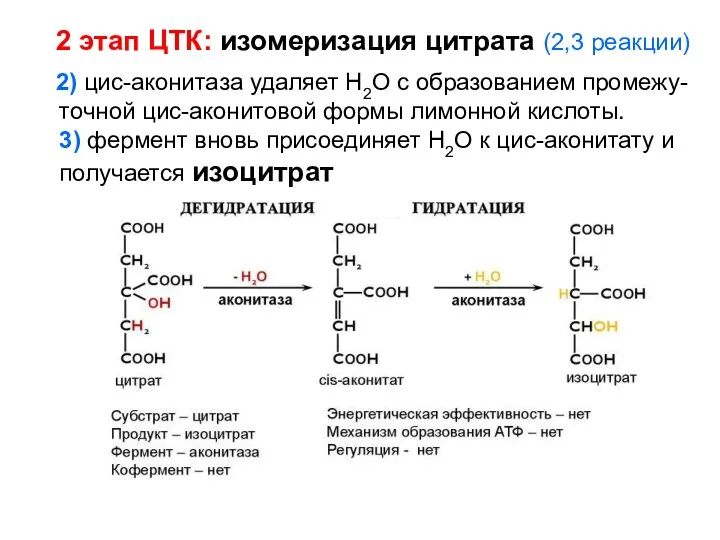
2 этап ЦТК: изомеризация цитрата (2,3 реакции)
2) цис-аконитаза удаляет
2 этап ЦТК: изомеризация цитрата (2,3 реакции)
2) цис-аконитаза удаляет

3 этап ЦТК: окислительное декарбоксилиро- вание изоцитрата (4,5 реакции)
3 этап ЦТК: окислительное декарбоксилиро- вание изоцитрата (4,5 реакции)
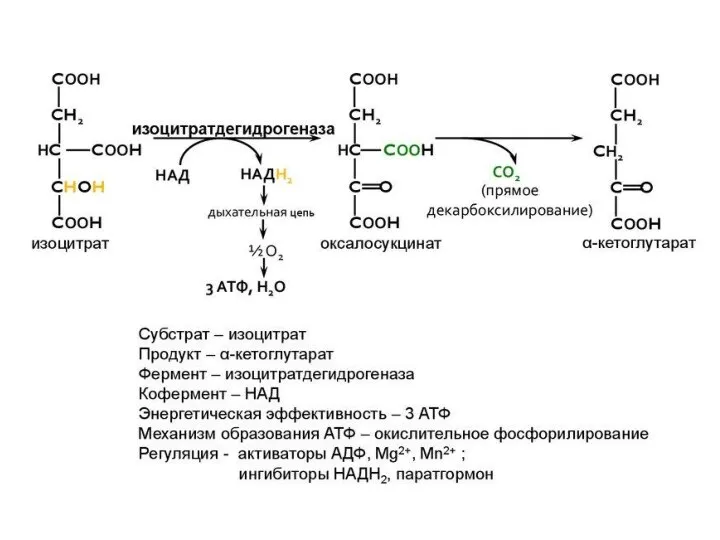

4 этап ЦТК: окислительное декарбоксилиро-
вание α-кетоглутарата (6 реакция) 6)
4 этап ЦТК: окислительное декарбоксилиро- вание α-кетоглутарата (6 реакция) 6)
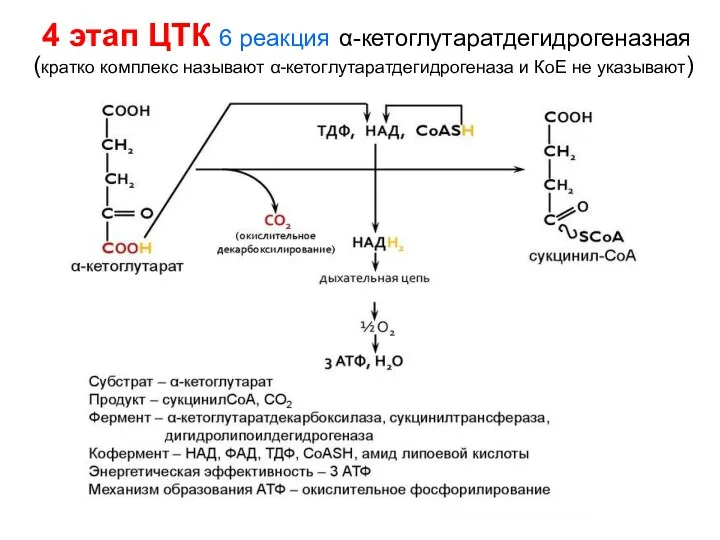
4 этап ЦТК 6 реакция α-кетоглутаратдегидрогеназная
(кратко комплекс называют α-кетоглутаратдегидрогеназа
4 этап ЦТК 6 реакция α-кетоглутаратдегидрогеназная
(кратко комплекс называют α-кетоглутаратдегидрогеназа

5 этап ЦТК: получение сукцината - (7 реакция), сукцинаттиокиназная реакция
5 этап ЦТК: получение сукцината - (7 реакция), сукцинаттиокиназная реакция
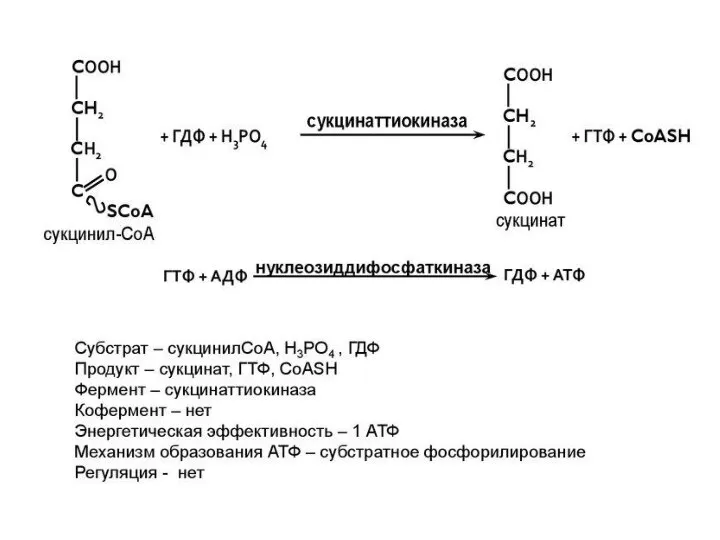

Образование высокоэнергетической фос-фатной связи ГТФ из сукцинил~КоА – есть пример
Образование высокоэнергетической фос-фатной связи ГТФ из сукцинил~КоА – есть пример
6 этап -
7 этап -
8 этап -
6 этап -
7 этап -
8
6 этап -
7 этап -
8 этап -
6 этап - 7 этап - 8
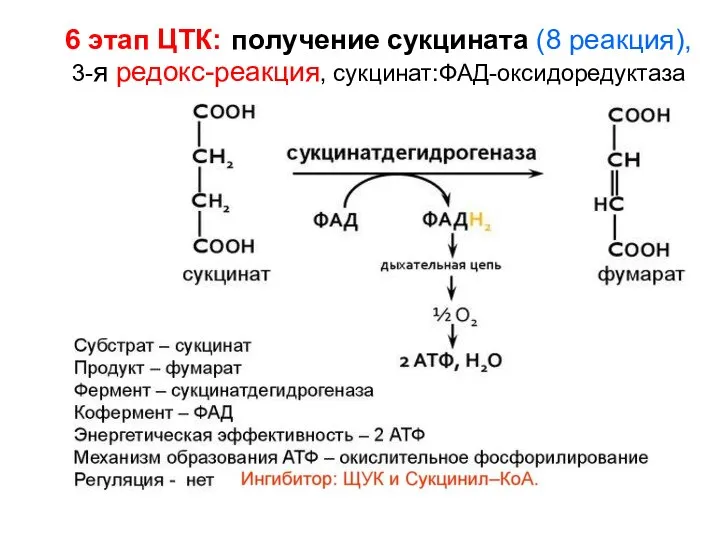
6 этап ЦТК: получение сукцината (8 реакция),
3-я редокс-реакция, сукцинат:ФAД-оксидоредуктаза
6 этап ЦТК: получение сукцината (8 реакция),
3-я редокс-реакция, сукцинат:ФAД-оксидоредуктаза
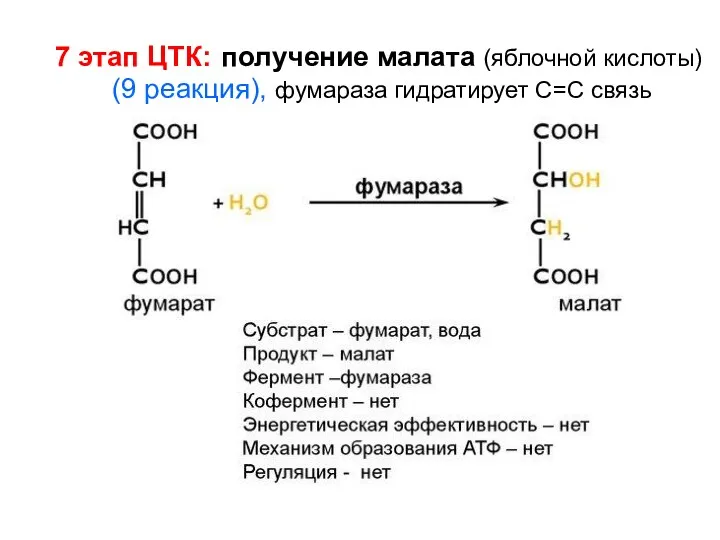
7 этап ЦТК: получение малата (яблочной кислоты) (9 реакция), фумараза гидратирует
7 этап ЦТК: получение малата (яблочной кислоты) (9 реакция), фумараза гидратирует
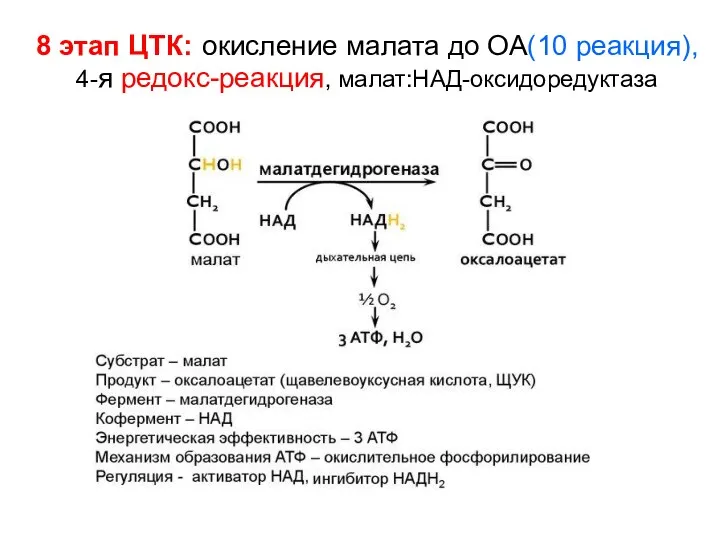
8 этап ЦТК: окисление малата до ОА(10 реакция),
4-я редокс-реакция, малат:НAД-оксидоредуктаза
8 этап ЦТК: окисление малата до ОА(10 реакция),
4-я редокс-реакция, малат:НAД-оксидоредуктаза

Суммарное уравнение ЦТК:
CH3CO~SKoA + ГДФ + Н3РО4 +
3НАД+
Суммарное уравнение ЦТК:
CH3CO~SKoA + ГДФ + Н3РО4 +
3НАД+

Подсчёт энергетики ЦТК
В ЦТК за 1 цикл сгорает 1 ацетил~КоА
Подсчёт энергетики ЦТК
В ЦТК за 1 цикл сгорает 1 ацетил~КоА
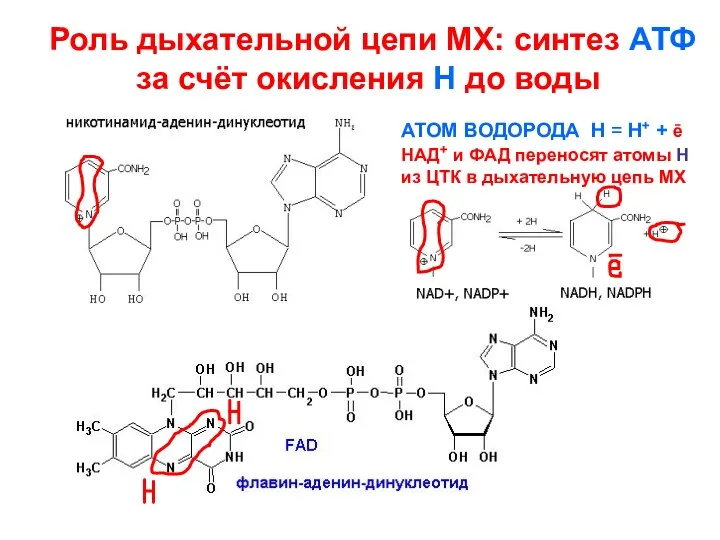
Роль дыхательной цепи МХ: синтез АТФ за счёт окисления Н до
Роль дыхательной цепи МХ: синтез АТФ за счёт окисления Н до
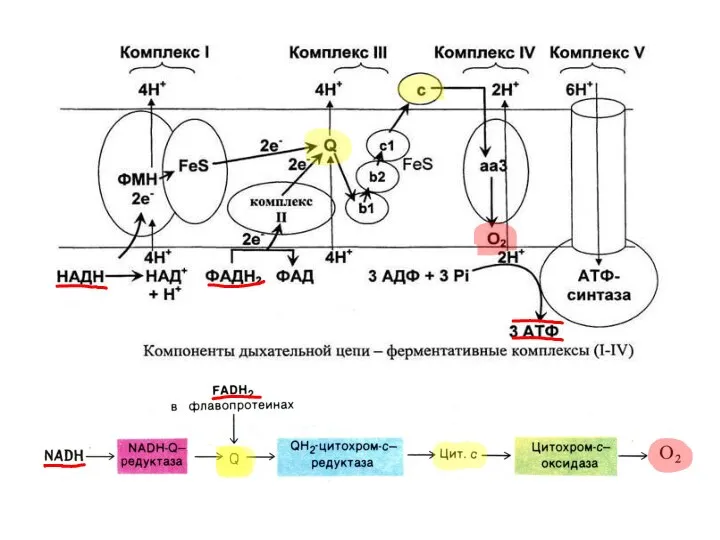
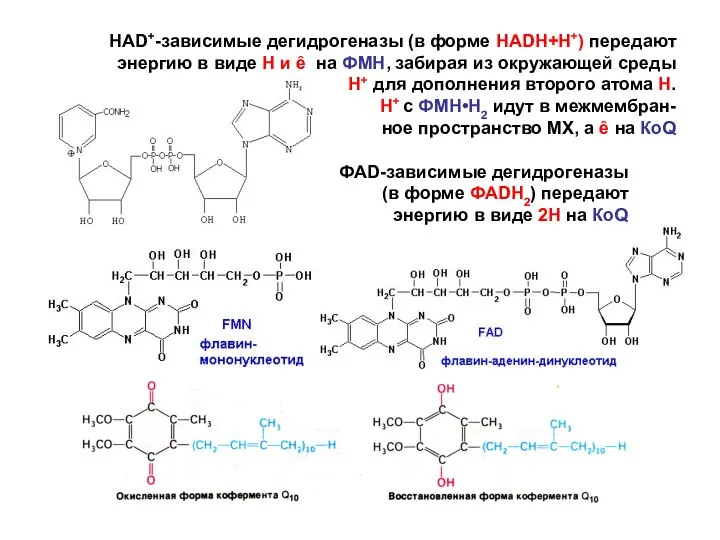
НАD+-зависимые дегидрогеназы (в форме НАDН+Н+) передают энергию в виде Н и
НАD+-зависимые дегидрогеназы (в форме НАDН+Н+) передают энергию в виде Н и
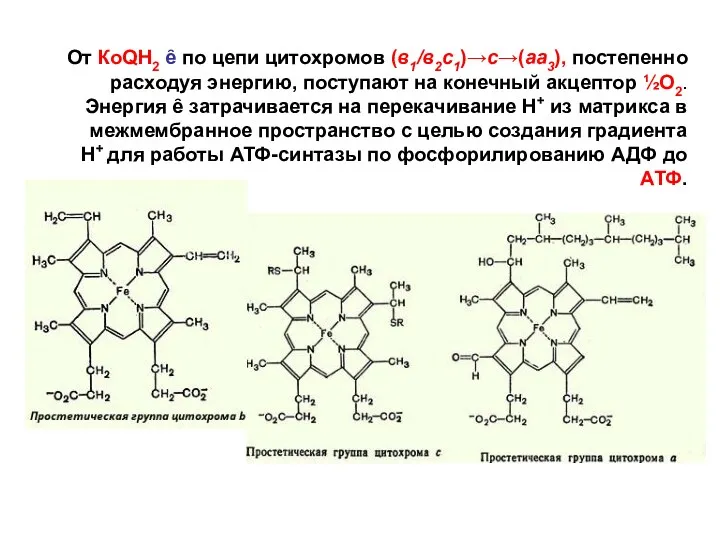
От КоQН2 ȇ по цепи цитохромов (в1/в2с1)→с→(аа3), постепенно расходуя энергию,
От КоQН2 ȇ по цепи цитохромов (в1/в2с1)→с→(аа3), постепенно расходуя энергию,
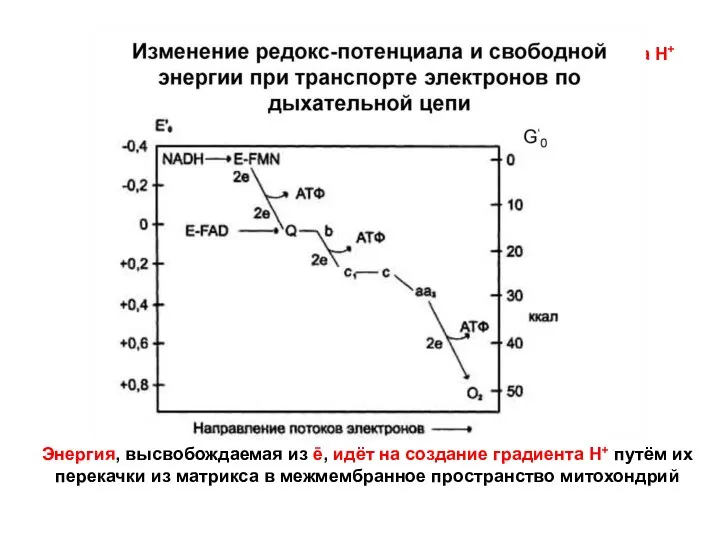
Энергия, высвобождаемая из ē, идёт на создание градиента Н+ путём их
Энергия, высвобождаемая из ē, идёт на создание градиента Н+ путём их
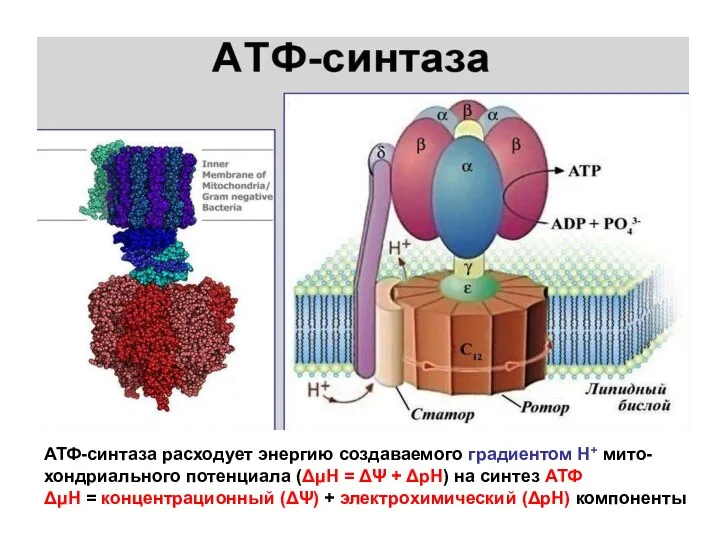
АТФ-синтаза расходует энергию создаваемого градиентом Н+ мито-
хондриального потенциала (ΔμΗ = ΔΨ
АТФ-синтаза расходует энергию создаваемого градиентом Н+ мито- хондриального потенциала (ΔμΗ = ΔΨ
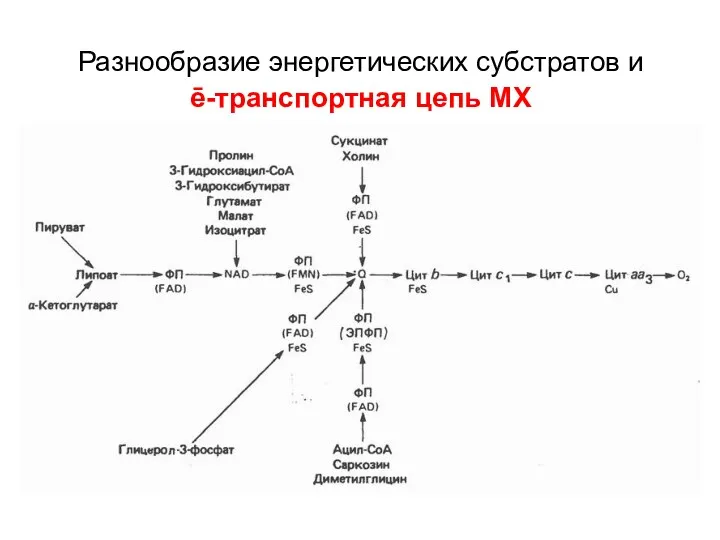
Разнообразие энергетических субстратов и
ē-транспортная цепь МХ
Разнообразие энергетических субстратов и
ē-транспортная цепь МХ
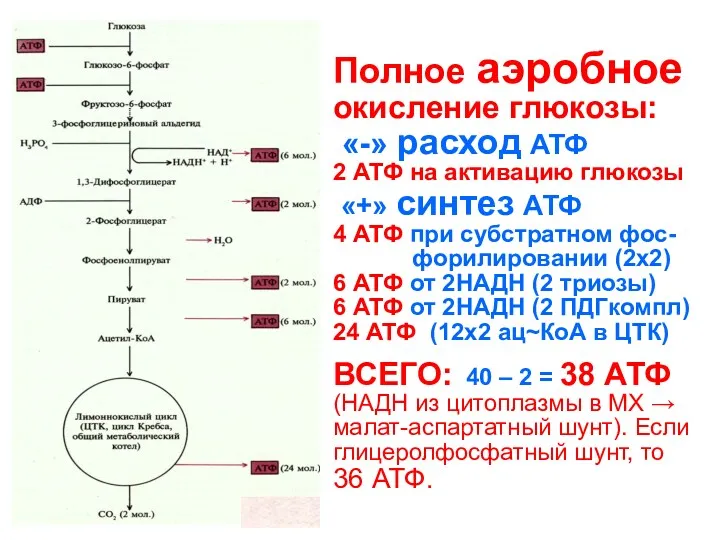
Полное аэробное окисление глюкозы:
«-» расход АТФ
2 АТФ на активацию глюкозы
Полное аэробное окисление глюкозы: «-» расход АТФ 2 АТФ на активацию глюкозы

Роль ЦТК
1) энергетическая ценность общий путь полного окисления топливных молекул
Роль ЦТК
1) энергетическая ценность общий путь полного окисления топливных молекул

Регуляция ЦТК
Скорость производства и утилизации энергии непостоянна в течение жизни клетки.
Регуляция ЦТК
Скорость производства и утилизации энергии непостоянна в течение жизни клетки.

При активной работе клетки в ней высокий уровень НАД+
При активной работе клетки в ней высокий уровень НАД+
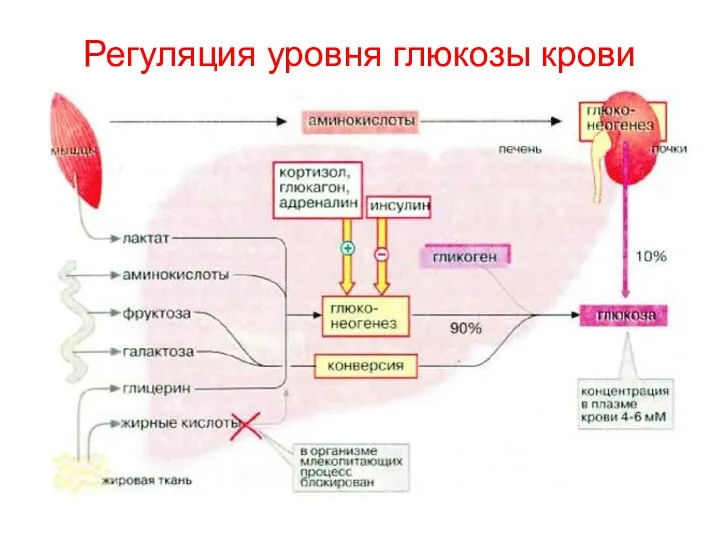
Регуляция уровня глюкозы крови
Регуляция уровня глюкозы крови

Нарушения углеводного обмена:
голодание
гипогликемия, глюкагон/адреналин мобилизуют ТАГ и ↑глюконеогенез из глицерола,
Нарушения углеводного обмена:
голодание гипогликемия, глюкагон/адреналин мобилизуют ТАГ и ↑глюконеогенез из глицерола,

Гипергликемия. После преодоления почечного порога присоединяется глюкозурия
Снижен транспорт глюкозы в клетку
Гипергликемия. После преодоления почечного порога присоединяется глюкозурия
Снижен транспорт глюкозы в клетку

Если человек тебе сделал ЗЛО — ты дай ему конфетку, он
Если человек тебе сделал ЗЛО — ты дай ему конфетку, он

инсулинонезависимый сахарный диабет
При длительном переедании и избыточном потреблении лёгких углеводов с
инсулинонезависимый сахарный диабет
При длительном переедании и избыточном потреблении лёгких углеводов с
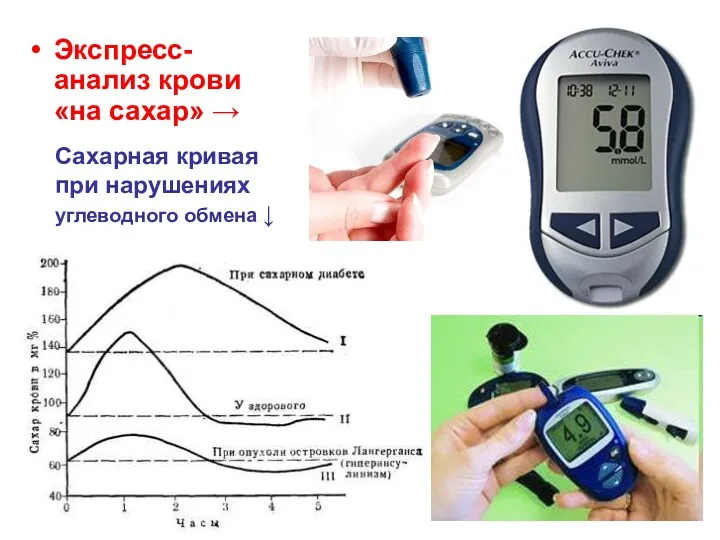
Сахарная кривая
при нарушениях
углеводного обмена ↓
Экспресс-анализ крови «на сахар» →
Сахарная кривая
при нарушениях
углеводного обмена ↓
Экспресс-анализ крови «на сахар» →
Экспресс-анализ мочи при сахарном диабете
Экспресс-анализ мочи при сахарном диабете
 Взаимодействие света с веществом. (Лекция 12)
Взаимодействие света с веществом. (Лекция 12) Амурский тигр
Амурский тигр презентация к уроку в 9 классе ФОТОСИНТЕЗ
презентация к уроку в 9 классе ФОТОСИНТЕЗ Cell Membrane Organization Detail
Cell Membrane Organization Detail Про растения из Красной книги Красноярского края
Про растения из Красной книги Красноярского края Метаболизм белков и аминокислот. Образование и обезвреживание аммиака. Тема 13.а
Метаболизм белков и аминокислот. Образование и обезвреживание аммиака. Тема 13.а Окружающая среда
Окружающая среда Анатомия внутреннего уха
Анатомия внутреннего уха Возрастные особенности зрительной сенсорной системы. Особенности рефлекторных реакций в разные возрастные периоды
Возрастные особенности зрительной сенсорной системы. Особенности рефлекторных реакций в разные возрастные периоды Сердечно-сосудистая система
Сердечно-сосудистая система Водоросли
Водоросли Мутационная и комбинативная изменчивость
Мутационная и комбинативная изменчивость Мутагенные факторы
Мутагенные факторы Обмен углеводов в организме человека
Обмен углеводов в организме человека Учение о популяции. Общая экология
Учение о популяции. Общая экология Обмен веществ и энергии в клетке
Обмен веществ и энергии в клетке Предмет ботаники. Растительная клетка. Протопласт и его производные
Предмет ботаники. Растительная клетка. Протопласт и его производные Витамины и их роль в жизни людей
Витамины и их роль в жизни людей Продукты, полезные для глаз
Продукты, полезные для глаз Введение в генетику микроорганизмов
Введение в генетику микроорганизмов Осьминоги. Головоногие моллюски
Осьминоги. Головоногие моллюски Комнатные растения и уход за ними
Комнатные растения и уход за ними Soluble mediators of Immunity
Soluble mediators of Immunity Ткани растений и животных
Ткани растений и животных Проведение опыта по определению pH талой, водопроводной и дистиллированной воды с помощью лакмусовой бумаги
Проведение опыта по определению pH талой, водопроводной и дистиллированной воды с помощью лакмусовой бумаги Физиологические механизмы слуховой и вестибулярной сенсорных систем
Физиологические механизмы слуховой и вестибулярной сенсорных систем Урок по теме Грибы в биосфере и жизни человека
Урок по теме Грибы в биосфере и жизни человека Основные особенности и методы изучения наследственности человека
Основные особенности и методы изучения наследственности человека